





-
九龙山榧Torreya jiulongshanensis为红豆杉科Taxaceae榧属Torreya高大乔木,雌雄异株,仅分布于浙江中部和西南部,模式产地为遂昌九龙山,被列为浙江省极小种群保护植物,最近一次被评估为极危(CR)植物[1]。2021年颁布的《国家重点保护野生植物名录》中,其被列为国家Ⅱ级保护植物(
http://www.gov.cn/zhengce/zhengceku/2021-09/09/content_5636409.htm )。目前,九龙山榧的种群数量极少,仅17株,其中模式产地仅1雌2雄共3株,每年结籽量很少,无幼树和幼苗,更新困难,而且生境破坏严重,人为干扰程度大[2-4]。九龙山榧作为榧属的古老孑遗植物,对研究古植物学和古地理学具有重要意义,其对于改良香榧T. grandis ‘Merrillii’品质也可能具有很大的潜在价值[5]。植物的有性生殖发育异常是濒危的主要原因之一,珍稀濒危植物或多或少存在生殖障碍[6-7]。研究发现:云南红豆杉Taxus yunnanensis不仅生殖周期长,且在1个雌配子体上有多个颈卵器,但最终只有1个或者少数几个颈卵器能得到精子,传粉效率低及雌雄性生殖系统发育不同步是造成其濒危的主要原因[8];香果树Emmenopterys henryi在胚发育过程中存在明显胚后熟现象,致使种子萌发率低,更新困难[9];羊角槭Acer yangjuechi在雌配子体发育过程中存在严重的生殖障碍,出现胚珠败育、胚囊退化及珠心组织细胞死亡等现象,是其濒危的重要原因[10];崖柏Thuja sutchuenensis从大、小孢子叶球形成至种子成熟的整个发育过程中均存在败育,而胚珠败育及雌配子体游离核时期至幼胚发育期间的败育是其生殖障碍的主要原因[11]。
自20世纪初以来,国内外学者先后对榧属几种植物生殖生物学开展了研究[12-14]。然而,关于九龙山榧生殖生物学的研究至今未见报道。本研究采用石蜡切片法,对九龙山榧大、小孢子的发生和雌、雄配子体的发育过程进行研究,并与红豆杉科其他植物加以比较,旨在从生殖生物学角度探讨九龙山榧的种子发育及结籽率低下是否与其大、小孢子的发生和雌、雄配子的发育异常有关,以期为九龙山榧保育措施的制定提供生殖生物学依据。
-
九龙山榧取自模式产地:丽水市遂昌县王村口镇西坑下村(28°20′01″N,118°55′22″E,海拔501 m)。以林缘仅有的1雌株和与雌株相近的1雄株为样株。
-
2018年6月底至2019年4月初,在小孢子叶球(雄球花)的芽分化到散粉期间,对其外部形态发育过程进行观测、拍照,并记录发育的各个时间段和重要时间节点。2018年6月28日至2019年2月13日,隔3 d采1次;2019年2月中旬至2019年4月初散粉,每天采样。每次取样5个。
2018年11月12日至2019年11月30日,在大孢子叶球(雌球花)的芽分化到种子发育期间,对其外部形态发育过程进行观测、拍照,记录各个发育时间段和重要时间节点。2018年11月12日至2019年4月4日,隔6 d采1次;2019年4月5—30日,每天取样。每次取样5个。
2019年4月5—6日的散粉期间,用毛笔对吐露传粉滴的胚珠进行人工授粉,授粉2 h后进行观察,对仍有传粉滴吐露的胚珠进行补粉,直到该大孢子叶球不再吐露传粉滴。2019年5月1日至2019年11月30日,隔6 d对大孢子叶球进行采样。每次采集3~5个。
采集的大、小孢子叶球浸泡固定于FAA溶液(体积分数为38%甲醛溶液∶冰醋酸∶体积分数为70%乙醇溶液=1∶1∶18,体积比)中,置于4 ℃冰箱中冷藏保存。采用常规石蜡切片法进行制片,切片厚度为5~7 μm[15],用改良爱氏苏木精染色[16],中性树胶封片,在Motic BA410E显微镜下观察并拍照。
-
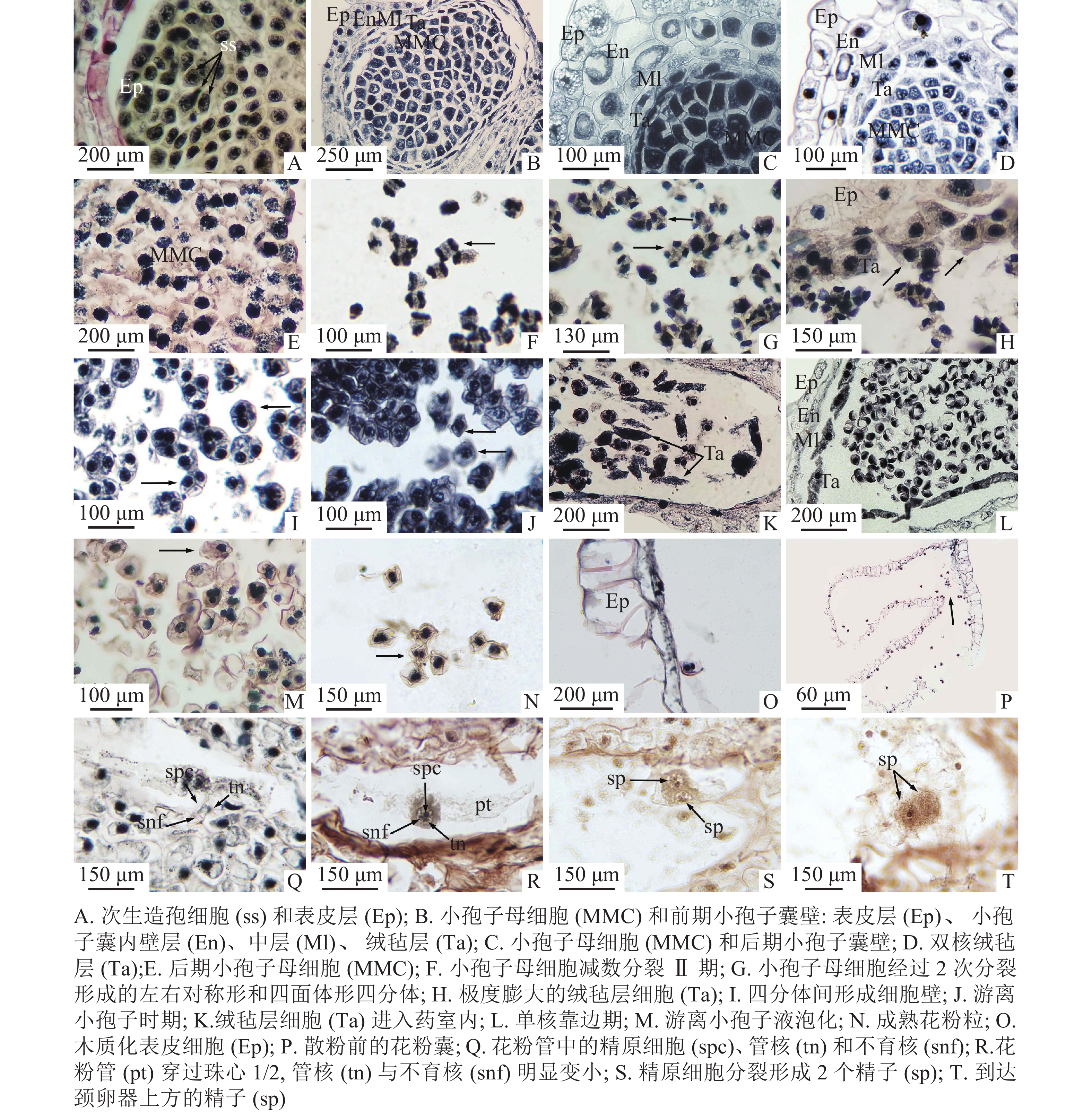
2018年6月29日,九龙山榧雄株的多数枝条叶腋处已经可见新生的混合芽,长为2.78~3.96 mm[(2.40±0.11) mm],宽为1.18~1.58 mm[(1.38±0.14) mm],混合芽着生于当年生枝条上,偶见于2年生枝条(图1A)。2018年9月22日,幼嫩的小孢子叶球体积明显变大,长为2.24~2.55 mm[(2.40±0.11) mm],宽为1.18~1.58 mm[(1.36±0.16) mm],外裹鳞片叶,深绿色,顶端扁而宽;营养芽的体积变化不大,顶端与茎端相似,保持较尖的圆锥形(图1B)。此时,从外观上很容易将小孢子叶球与营养芽区分出来。2018年10月12日,小孢子叶球呈圆锥形(图1C),外裹绿色鳞片叶,基部着生4枚苞片,2轮鳞片。小孢子叶球单生,长为2.78~3.96 mm[(3.37±0.47) mm],宽为1.01~1.91 mm[(1.62±0.38) mm],有一短轴,轴上螺旋状紧密排列着30~40枚小孢子叶,小孢子叶背面常着生4个(稀为3或5个)小孢子囊。2019年2月26日,小孢子叶球中下部变圆,呈浅绿色,芽鳞逐渐张开,露出小孢子囊(图1D)。2019年3月25日,小孢子叶球叶轴伸长,小孢子囊突破苞片、鳞片,逐渐伸到芽鳞外(图1E),紧密的小孢子叶变得松散,小孢子囊完全暴露在空气中,开裂后花粉散出,此时小孢子叶球成熟(图1F)。成熟的小孢子叶球呈长圆柱形,饱满,小孢子叶逐渐变成黄绿色。

图 1 大、小孢子叶球(雌、雄球花)的发育
Figure 1. Development of microstrobilus and macrostrobilus (male and female strobili)
2019年4月4日,小孢子囊开始散粉,散粉时间持续4 d,时间较快。散粉前小孢子囊开裂,花粉散出,颜色呈黄色。同一小孢子叶球中,叶轴基部的小孢子囊散粉通常比叶轴上部的早开裂(图1G)。散粉后,小孢子叶球迅速干缩(图1H)。
-
2018年11月12日,1年生枝条顶端存在普通芽和生殖芽,外观难以区分(图1I)。此时,生殖芽内部的苞叶叶腋出现珠被原基的隆起(图2A)。此后珠被不断生长,2019年3月25日,珠被生长至珠心上方,包被珠心(图2B)。此时,大孢子叶球2个,成对生于叶腋,外观呈圆锥形,长为0.38~0.49 mm[(0.47±0.12) mm],宽为0.26~0.29 mm[(0.28±0.09) mm],两侧微微隆起,外裹绿色鳞片状叶,着生于1年生雌株枝条顶端。每一大孢子叶球的短轴上紧密排列着4枚两两交互对生的苞片和最基部1枚侧生的苞片,具1个直立胚珠。2019年3月28日,在小孢子叶球即将散粉前,大孢子叶球逐渐从外包苞片和鳞片中突破,伸到芽鳞外,胚珠开始暴露在空气中,为接收花粉做准备(图1J)。2019年4月6日,假种皮开始露出(图2C)。此时,大孢子叶球呈圆球形,长为1.48~2.12 mm[(1.86±0.24) mm],宽为1.07~1.47 mm[(1.37±0.28) mm]。2019年4月4—7日为散粉期,胚珠的珠孔端吐露出传粉滴,授粉后传粉滴消失(图1K)。2019年4月23日,假种皮的长度逐渐生长至珠被1/2处,可见明显的珠孔和珠心上部的溶解腔(图2D)。2019年4月30日,胚珠个体逐渐变大,外层的珠被和假种皮逐渐伸长,但还未完全包裹住胚珠的珠孔(图1L)。2019年6月8—29日,成对的大孢子叶球仅1个发育,发育的假种皮长度逐渐超过珠被,突破鳞片,包裹住胚珠(图1M和图1N),先端较尖。2019年8月29日,胚珠长度增大,深绿色,胚珠着生的枝条顶端出现新生的小孢子叶球(图1O)。2019年9月22日,大孢子叶球呈卵球形,长为6.18~7.28 mm[(6.85±0.42) mm],宽为3.15~3.50 mm[(3.32±0.11) mm],胚珠底部变黄,假种皮先端变红(图1P)。
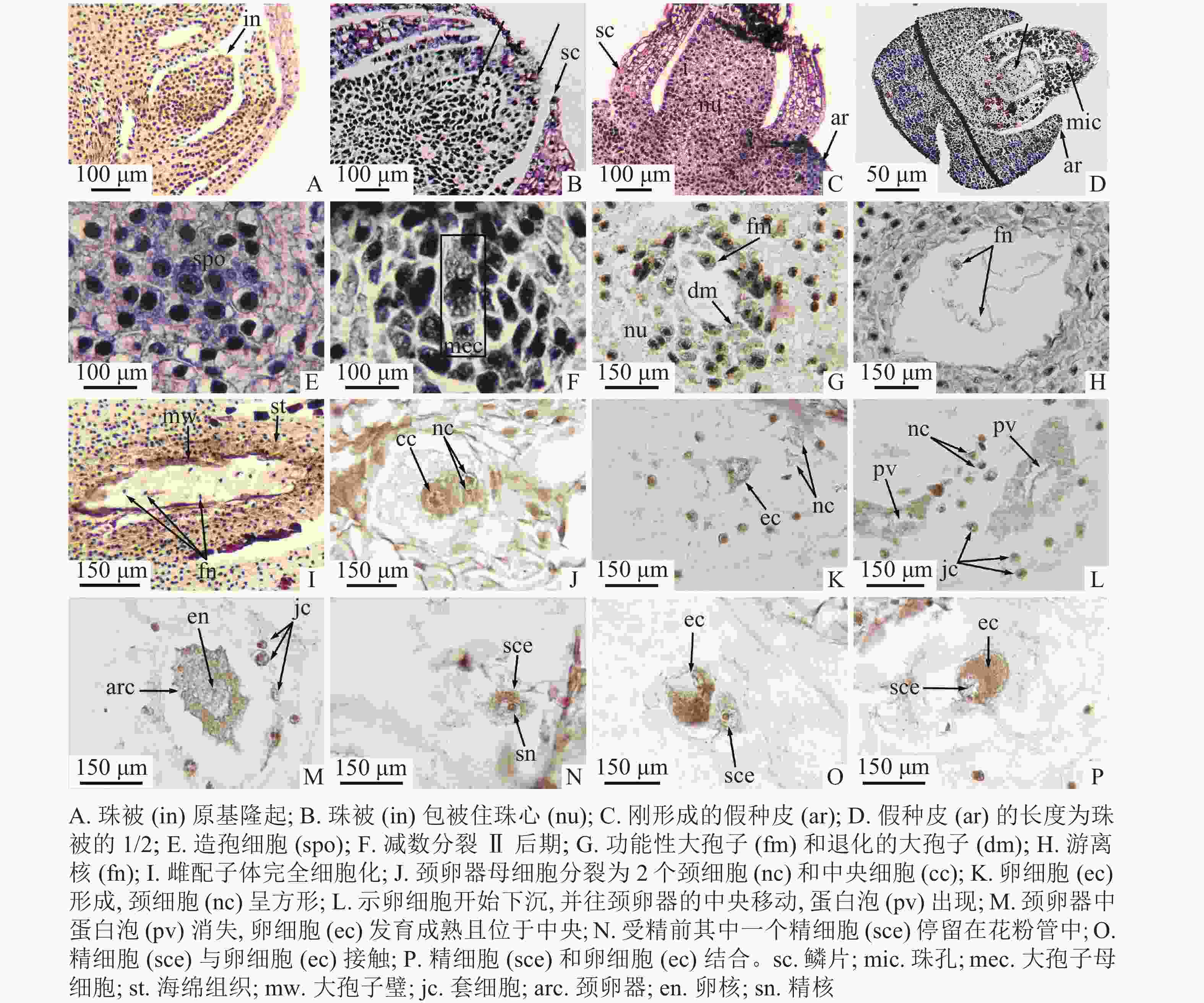
图 2 大孢子发生和雌配子体发育(含受精作用)
Figure 2. Megasporogenesis and female gametophyte development (including fertilization)
-
2018年8月20日,小孢子叶原基表皮下方的孢原细胞已经分化形成次生造孢细胞。次生造孢细胞紧密相连,呈多边形(图3A)。2018年11月12日,外层的次生壁细胞开始分裂、分化,小孢子囊由外而内最终形成矩形的表皮层,呈椭圆形的小孢子囊内壁,呈不规则散状排列的2层中层细胞,以及最内呈长条形的绒毡层(图3B)。小孢子囊发育类型为基本型。2018年11月15日,造孢细胞分化形成小孢子母细胞,最初的小孢子母细胞由于体积较大、排列紧密,呈多边形,胞质浓厚,细胞核大(图3C)。2019年1月3日,在小孢子母细胞不断形成时期,最外层的表皮细胞经垂周分裂后,垂周壁加厚,径向壁延长,细胞内液泡化,细胞核和核仁逐渐消失,木质化加强,以适应内部小孢子母细胞数目的增加;内壁细胞径向延长,并纤维化加厚;中层细胞被挤压,切向壁延长呈扁平状紧贴内壁;绒毡层细胞在小孢子母细胞时期最初呈长条形单核延长,发育后期细胞个体逐渐变大,细胞质变浓,并以单核或双核形式存在(图3D)。2019年2月2日,排列紧密的小孢子母细胞逐渐从胼胝质中解离,变成游离的小孢子母细胞,形状从多边形变为圆形(图3E)。

图 3 小孢子发生和雄配子体发育(含精子形成)
Figure 3. Microsporogenesis and male gametophyte development (including spermatogenesis)
2019年2月26日至3月6日为小孢子母细胞减数分裂期。小孢子母细胞经过第1次减数分裂,形成2个子核(图3F),2个子核之间不形成细胞壁直接进入减数分裂Ⅱ期,再次分裂之后形成四面体形、左右对称形2种类型的四分体(图3G)。小孢子囊壁和中层细胞开始解体,绒毡层细胞多为双核,由绒毡层出现的位置判断绒毡层细胞为周原质团细胞,绒毡层类型为变形绒毡层。小孢子囊内约7%绒毡层出现异常膨大,堆叠在一起(图3H)。2019年3月12日,四分体中的4个小孢子之间开始形成各自的细胞壁(图3I)。2019年3月16日,四分体开始解体,胼胝质壁消失,小孢子的细胞壁逐渐加厚,形成游离小孢子细胞(图3J),绒毡层部分细胞进入药室内部(图3K)。2019年3月19日为小孢子细胞单核靠边期,表皮细胞木质化加强,内壁继续解体,中层细胞仅留下残迹,绒毡层仍以双核或单核形式存在(图3L)。一些小孢子囊内游离小孢子内液泡化导致细胞变形,约占11%(图3M)。
2019年3月31日,单核靠边的小孢子细胞壁开始逐渐增厚,细胞核经过有丝分裂形成2个核,细胞质也在2个核之间形成细胞板将2个核隔开,其中大的核为管核,小的核为生殖核。此时,成熟花粉粒形成(图3N)。成熟花粉粒的直径为30 μm,双核,表面褶皱成不规则状,无气囊。花粉成熟期时,小孢子囊内壁与中层细胞仅剩残迹,绒毡层消失,表皮细胞完全木质化并且带状加厚(图3O)。2019年4月4—7日,开始散粉时,小孢子囊囊基部囊壁没有加厚的开裂口破裂,从开裂口散出(图3P)。2019年5月30日,在胚珠上方花粉粒萌发成花粉管,伸入珠心组织1/3处,花粉管中可清晰看到管细胞、生殖细胞和不育细胞,生殖细胞比不育细胞稍大(图3Q)。2019年6月24日,花粉管入侵至珠心1/2处,生殖核明显增大,管核与不育核明显变小,且即将消失(图3R)。2019年7月27日,花粉管抵达雌配子体壁,此时管核和不育核已消失,位于花粉管先端的精原细胞已分裂成2个形状相似、大小相同的精细胞。精细胞核大、细胞质浓,形状为椭圆形或圆形(图3S)。2019年11月29日,花粉管伸长至颈卵器上方,精细胞明显增大,细胞质浓厚(图3T)。九龙山榧小孢子及雄配子体发育进程见表1。
表 1 九龙山榧的有性生殖过程
Table 1. Process of sexual reproduction in T. jiulongshanensis
发育时期(年-月-日) 小孢子和雄配子体发育 大孢子和雌配子体发育 2018-08-20 次生造孢细胞 2018-11-15 小孢子母细胞形成 2019-02-26—2019-03-06 小孢子母细胞减数分裂期 2019-03-12—2019-03-16 四分体时期 2019-03-16 小孢子从四分体中相互分离 2019-03-19 单核靠边期 2019-04-06 散粉与传粉 造孢组织 2019-04-23 减数分裂Ⅱ后期 2019-05-18 功能大孢子 2019-05-30 管细胞、生殖细胞和不育细胞 2019-06-06 游离核时期 2019-06-24 管核与不育核即将消失 2019-07-27 精原细胞分裂形成两个大小相同的精细胞 2019-08-11 细胞化阶段 2019-09-22 颈卵器母细胞 2019-09-22—2019-11-29 颈卵器阶段 2019-11-29 受精 -
2019年4月6日,在散粉期,珠心下方与珠被齐平的水平线上出现一团核大、质浓、呈多边形的造孢细胞(图2E)。①大孢子发生时期:2019年4月23日,造孢细胞分化,在近中央位置形成大孢子母细胞,并进行减数分裂Ⅰ,形成大孢子二分体,二分体再进行减数分裂Ⅱ(图2F),最终形成纵向直列的4个大孢子。2019年5月18日,合点端的大孢子不断发育,最终形成功能性大孢子,近珠孔的3个大孢子逐渐退化(图2G)。②游离核时期:2019年6月6日,合点端的功能大孢子继续进行多次有丝分裂,形成多核的雌配子体,但不形成细胞壁,细胞核呈游离状态,分布在雌配子体的细胞质中(图2H)。游离核之间由染色质丝相连。③细胞化时期:2019年8月11日,当游离核分裂8次,形成256个游离核后,在连接游离核间原生质丝的基础上,开始向心式形成细胞壁,最后整个雌配子体完全细胞化(图2I)。④颈卵器时期:当雌配子体不断发育到一定程度,近珠孔端的一些细胞开始逐渐膨大,形成颈卵器母细胞。2019年9月22日,颈卵器母细胞进行平周和垂周分裂,最终形成4个较小的颈细胞和1个较大的中央细胞(图2J)。刚形成的颈细胞呈圆形,细胞核明显,个体较周围细胞小,中央细胞呈圆形,细胞核较大,周围细胞质浓。中央细胞进行1次不均等分裂,形成卵核和腹沟核,腹沟核很快消失。由于此过程发生较快,因此未捕捉到正在退化的腹沟核。2019年9月30日,初形成的卵核位于颈卵器的近珠孔端,周围有少量细胞质包围,卵核下方有1个大液泡(图2K)。2019年10月13日,卵核开始下沉,往颈卵器的中央开始移动,同时蛋白泡出现(图2L)。2019年11月12日,受精前,成熟的颈卵器中蛋白泡消失,细胞质变浓,卵细胞发育成熟且位于中央,等待受精(图2M)。九龙山榧的颈卵器为椭圆形,多数位于雌配子体的近珠孔端,同一胚珠中有2个颈卵器。颈卵器的周围通常紧密排列着1层套细胞,套细胞的细胞核大、体积较小(图2L~M)。本研究观察到九龙山榧每个胚珠中只产生1个雌配子体,含2个单生型颈卵器,与香榧相同。九龙山榧的大孢子及雌配子体发育进程见表1。
-
九龙山榧的受精作用发生于2019年11月29日,从传粉到受精约7个月,在花粉管中产生2个大小相似的精细胞(图3T),受精前其中一个精细胞进入颈卵器中与卵细胞结合,另一个则停留在花粉管中(图2N)。在精细胞接触卵细胞前,精细胞边缘整齐,细胞核明显,两者接触时,精细胞边缘开始变得模糊,细胞质因变得蓬松而染色较浅(图2O)。在精细胞和卵细胞逐渐融合的过程中,精细胞核区逐渐消失,细胞质相融(图2P)。
-
裸子植物小孢子母细胞发育节律主要分为4种类型[17-20]:①小孢子母细胞的减数分裂过程起始于初冬,进入休眠期停止减数分裂,在翌年春季解除休眠后完成后续发育,如侧柏Platycladus orientalis。②翌年春天形成造孢细胞,再分化成小孢子母细胞,小孢子母细胞不经过休眠直接开始进行减数分裂,如穗花杉Amentotaxus argotaenia。③小孢子母细胞于当年年底前已经分化形成,经过翌年春季才开始后续的减数分裂过程,如欧洲赤松Pinus sylvestris。④小孢子母细胞进行减数分裂并形成游离小孢子再越冬,翌年春季继续发育为成熟花粉,红豆杉属Taxus加拿大红豆杉T. canadenesis、短叶红豆杉T. brevifolia、南方红豆杉T. wallichiana var. mairei和云南红豆杉T. yunnanensis等属此类型。九龙山榧与香榧小孢子母细胞的发育方式相同[21],这种发育方式避免了其减数分裂过程受到寒冷冬季低温影响,降低了减数分裂发生异常的风险。九龙山榧在减数分裂时期部分小孢子囊的绒毡层发生异常增生和膨大现象。陈祖铿等[22]对穗花杉研究发现:绒毡层细胞发生异常膨大在裸子植物发育中为异常现象,其结果会导致小孢子母细胞受到挤压,在减数分裂过程中发生异常,最终引起花粉败育。这也曾在太白红杉Larix chinensis [23]的小孢子发育过程中有过报道。九龙山榧花粉囊中的绒毡层从小孢子母细胞时期一直持续存在到单核靠边期,直到形成成熟花粉粒才完全降解。绒毡层的延迟降解可能会争夺游离小孢子细胞发育所需的营养物质和空间。单核靠边期时,少部分小孢子囊中出现小孢子细胞内液泡化现象,导致一些细胞形状变化、破裂。但由于花粉粒能在雌花胚珠的珠心上方萌发出花粉管,且雄配子体的发育基本正常,说明花粉粒具备正常的生理活性和后期生殖功能,这与本研究对其花粉活力测定的结果一致。由此可见,小孢子的发生和雄配子体的发育均正常。
裸子植物的花粉萌发普遍迟缓且花粉管的生长非常缓慢[24]。穗花杉于5月底开始散粉,7月中下旬形成精子,历时约2个月[25];短叶红豆杉于4月底萌发花粉管,6月初形成精子,历时1个多月[26];南方红豆杉2月中下旬散粉,4月中旬体细胞迅速分裂形成2个精子,历时约2个月[20];香榧于4月下旬散粉,7月中下旬花粉管中体细胞核分裂出2个大小相等的精核,过程近3个月[27]。九龙山榧4月初散粉,4月底花粉才开始萌发出花粉管,7月底雌配子体上方产生2个精原细胞。因此,九龙山榧从传粉到形成精子,整个过程历时近4个月,发育周期均较红豆杉科其他种更长。这可能是雄配子体在花粉管中发育缓慢以等待颈卵器中的卵细胞发育成熟以完成受精[20]。受精延迟现象在裸子植物中普遍存在。白豆杉Pseudotaxus chienii于4月17日传粉,5月下旬陆续发生受精作用,两者相隔1个多月[28];穗花杉于5月25日至6月15日传粉,7月20—29日受精,两者相隔约2个月[22];云南红豆杉的受精作用发生于3月底至4月初,传粉与受精相隔约4个月[8];香榧于4月下旬授粉,9月上旬发生受精作用,从传粉到受精间隔4~5个月[27]。在本研究中,九龙山榧于4月初开始传粉,11月下旬发生受精作用,从传粉到受精需要约7个月,晚于红豆杉科已经报道的大多数植物。
植物有性生殖过程的任何一个环节出现障碍,都会造成生殖失败,种子减少,更新困难从而致濒[29]。九龙山榧小孢子叶球8月中旬开始发生,至2019年7月底才形成精子,历时11个月余;大孢子叶球11月中上旬开始发生,至2019年11月底才进行受精作用,过程历时12个月余,雌雄生殖系统发育的周期均较红豆杉科以往报道的其他种更长。生殖过程历时久、环节多,增加花粉败育、胚珠死亡的概率[8],且从传粉到受精时间跨度大,加之个体数量极少,仅1雌株2雄株,胚珠发育严重滞后于小孢子叶球的散粉期,可能导致受精率降低甚至受精作用受阻。此外,陈佳妮等[30]发现:香榧结籽高于榧树,在于前者成熟叶片的氮含量和光合能力显著高于后者。九龙山榧为喜光的阳生树种[31],其结籽率低的原因可能还与雌株树体老化、光合生理特性、外界营养不足等有关。
可见,九龙山榧冗长的生殖周期、复杂的生殖过程和雌性生殖系统发育明显滞后于雄性生殖系统,加之人为干扰强、树体老化、营养不足、个体数量少等均可能造成其结籽率底、自然更新困难,进而致濒。从实际出发,对九龙山榧的保育可以借鉴香榧的培育技术:①由于九龙山榧雌雄异株,小种群中的花粉密度很难达到较高的水平,胚珠传粉滴的吐露容易受不良天气的影响,因此在散粉期收集足够的花粉,将花粉合理优化保存,分期在晴天进行人工授粉以提高授粉率[32];②加强雌株的科学管理,平衡施肥,追施磷、钾复合肥[33],浅根多次施肥,繁殖期叶面施淡肥[34],从而促进胚的发育和提高坐果率。
-
材料采集过程中得到浙江遂昌王村口镇西坑下村严东根先生的无私帮助。在此深表谢意!
Microsporogenesis, megasporogensis and development of male and female gametophytes of Torreya jiulongshanensis, a critically endangered plant
-
摘要:
目的 九龙山榧Torreya jiulongshanensis为浙江特有的古老孑遗植物,数量极少,为国家Ⅱ级重点保护野生植物,也被列为浙江省极小种群保护植物。系统观测九龙山榧大、小孢子的发生和雌、雄配子体的发育过程,有助于发现其配子是否发育异常及导致结籽率低下的原因。 方法 以九龙山榧模式产地的雌、雄植株为研究对象,通过定期观测和取样,采用石蜡切片法对其大、小孢子的发生,雌、雄配子体的发育过程进行切片、染色和观察。 结果 ①小孢子叶球于8月中旬开始分化,8月下旬分化形成次生造孢细胞,11月中旬形成小孢子母细胞,翌年2月下旬进入减数分裂期,3月中旬形成游离小孢子细胞,于3月末逐渐成熟;成熟花粉粒为双核,无气囊。小孢子囊的发育类型为基本型,孢质分裂方式为同时型,四分体有四面体形和左右对称形。②雄配子体从3月中下旬开始发生,到7月底形成精子,整个过程历时约4个月。③翌年3月下旬,大孢子叶球每个苞叶的叶腋基部直立着生2枚胚珠;大孢子母细胞于4月下旬经过减数分裂Ⅰ和减数分裂Ⅱ形成4个直列大孢子,5月中旬珠孔端的3个大孢子退化,合点端的大孢子最终发育成为雌配子体。④九龙山榧有2个单生型椭圆形的颈卵器,从传粉到受精历时约7个月。 结论 九龙山榧雌雄生殖系统发育的周期均较红豆杉科Taxaceae其他种更长,其冗长的生殖周期、复杂的生殖过程和雌性生殖系统发育明显滞后于雄性生殖系统,加之种群个体数量极度稀少(1雌株、2雄株)是造成结籽率低下、自然更新困难的主要原因。图3表1参34 Abstract:Objective Torreya jiulongshanensis, an ancient relict plant endemic to Zhejiang Province, is a national Class Ⅱ key protected wild plant and also listed as a protected plant of extremely small population in Zhejiang Province. The purpose of this study is to systematically observe the occurrence of megaspores and microspores of T. jiulongshanensis, as well as the development of male and female gametophytes , so as to find out whether the gametes are abnormal and the reasons for low seed-setting rate. Method The male and female individuals of T. jiulongshanensis from the type locality were collected as the research objects. Through regular observation and sampling, the occurrence of megaspores and microspores and the development of female and male gametophytes were sectioned, stained and observed by paraffin sectioning method. Result (1) The microsporophyll differentiated in mid-August, differentiated into secondary sporogenesis cells in late August and formed microspore mother cells in mid-November. In late February of the next year, it entered the meiosis stage, formed free microspore cells in mid-March, and gradually matured at the end of March. The mature pollen grains were binucleate without air sacs. The development type of microsporangium was basic type, and the spore division model was simultaneous type. The tetrad had tetrahedral and zygomorphic forms. (2) Male gametophytes began to develop in mid-to-late March, and formed sperm at the end of July. The whole process lasted about 4 months. (3) In late March of the next year, 2 erect ovules were born at the base of leaf axils of each bract in megasporophyll. In late April, the megaspore mother cell underwent meiosis Ⅰ and Ⅱ, and formed 4 linear megaspores. In mid-May, 3 megaspores at the micropylar end degenerated and the megaspores at the chalazal end finally developed into female gametophytes. (4) T. jiulongshanensis had 2 solitary oval archegonia, which lasted about 7 months from pollination to fertilization. Conclusion The development cycle of male and female reproductive system of T. jiulongshanensis is longer than that of other species in the family Taxaceae. The main reasons for low seed-setting rate and difficult natural regeneration of T. jiulongshanensis include long reproductive cycle, complicated reproductive process, the development of female reproductive system which lags behind that of the male reproductive system, as well as the extremely limited population (1 female plant and 2 male ones). [Ch, 3 fig. 1 tab. 34 ref.] -
Key words:
- Torreya jiulongshanensis /
- microsporogenesis /
- male gametophyte /
- megasporogenesis /
- female gametophyte
-
图 1 大、小孢子叶球(雌、雄球花)的发育
Figure 1 Development of microstrobilus and macrostrobilus (male and female strobili)
图 2 大孢子发生和雌配子体发育(含受精作用)
Figure 2 Megasporogenesis and female gametophyte development (including fertilization)
图 3 小孢子发生和雄配子体发育(含精子形成)
Figure 3 Microsporogenesis and male gametophyte development (including spermatogenesis)
表 1 九龙山榧的有性生殖过程
Table 1. Process of sexual reproduction in T. jiulongshanensis
发育时期(年-月-日) 小孢子和雄配子体发育 大孢子和雌配子体发育 2018-08-20 次生造孢细胞 2018-11-15 小孢子母细胞形成 2019-02-26—2019-03-06 小孢子母细胞减数分裂期 2019-03-12—2019-03-16 四分体时期 2019-03-16 小孢子从四分体中相互分离 2019-03-19 单核靠边期 2019-04-06 散粉与传粉 造孢组织 2019-04-23 减数分裂Ⅱ后期 2019-05-18 功能大孢子 2019-05-30 管细胞、生殖细胞和不育细胞 2019-06-06 游离核时期 2019-06-24 管核与不育核即将消失 2019-07-27 精原细胞分裂形成两个大小相同的精细胞 2019-08-11 细胞化阶段 2019-09-22 颈卵器母细胞 2019-09-22—2019-11-29 颈卵器阶段 2019-11-29 受精  下载: 导出CSV
下载: 导出CSV
-
[1] 覃海宁, 赵莉娜, 于胜祥, 等. 中国被子植物濒危等级的评估[J]. 生物多样性, 2017, 25(7): 745 − 757. QIN Haining, ZHAO Lina, YU Shengxiang, et al. Evaluating the endangerment status of China’s angiosperms through the red list assessment [J]. Biodiversity Sci, 2017, 25(7): 745 − 757. [2] 孙孟军, 邱瑶德. 浙江林业自然资源: 野生植物卷[M]. 北京: 中国农业科学技术出版社, 2002: 75 − 176. SUN Mengjun, QIU Yaode. Zhejiang Forestry Natural Resources: Wild Plants[M]. Beijing: China Agricultural Science and Technology Press, 2002: 75 − 176. [3] 滕童莹, 潘成椿, 刘菊莲, 等. 浙江榧属植物叶片变异及其分类学意义[J]. 杭州师范大学学报(自然科学版), 2017, 16(1): 9 − 12. TENG Tongyin, PAN Chengchun, LIU Julian, et al. Leaf variation of Torreya plants in Zhejiang and its taxonomic implications [J]. J Hangzhou Norm Univ Nat Sci, 2017, 16(1): 9 − 12. [4] JIANG Ming, WANG Junfeng, ZHANG Huijuan. Characterization of the complete plastome sequence of Torreya grandis var. jiulongshanensis (Taxaceae), a rare and endangered plant species endemic to Zhejiang Province, China [J]. Mitochondrial DNA Part B, 2020, 5(1): 834 − 836. [5] 康宁, 汤仲埙. 榧属分类学研究[J]. 植物研究, 1995, 15(3): 349 − 362. KANG Ning, TANG Zhongxun. Studies on the taxonomy of the genus Torreya [J]. Bull Bot Res, 1995, 15(3): 349 − 362. [6] 张文辉, 祖元刚, 刘国彬. 10种濒危植物的种群生态学特征及致危因素分析[J]. 生态学报, 1512, 22(9): 1512 − 1520. ZHANG Wenhui, ZU Yuangang, LIU Guobin. Population ecological characteristics and analysis on endangered cause of ten endangered plant species [J]. Acta Ecol Sin, 1512, 22(9): 1512 − 1520. [7] 徐刚标, 刘雄盛, 梁文斌. 极度濒危植物水松大孢子发生、雌配子体发育及胚形成[J]. 林业科学, 2015, 51(6): 50 − 62. XU Gangbiao, LIU Xiongsheng, LIANG Wenbin. Megasporogenesis, female gametophyte development and embryogenesis in critically endangered Glyptostrobus pensilis [J]. Sci Silv Sin, 2015, 51(6): 50 − 62. [8] 王兵益. 云南红豆杉生殖生态学研究[D]. 北京: 中国林业科学研究院, 2008. WANG Bingyi. Reproductive Ecology of Taxus yunnanensis[D]. Beijing: Chinese Academy of Forestry, 2008. [9] 李利平, 李争艳, 王玉兵, 等. 香果树花及胚胎发育的细胞学研究[J]. 植物研究, 2012, 32(6): 646 − 650. LI Liping, LI Zhengyan, WANG Yubing, et al. Cytology research on the flower and embryo development of Emmenopterys henryi Oliv. [J]. Bull Bot Res, 2012, 32(6): 646 − 650. [10] 刘杨. 珍稀濒危植物羊角槭的繁殖生物学研究[D]. 南昌: 江西农业大学, 2014. LIU Yang. Study on Reproductive Biology of the Rare Plants Acer yanjuechi[D]. Nanchang: Jiangxi Agricultural University, 2014. [11] 金江群, 任风鸣, 夏鹰, 等. 极小种群野生植物崖柏的生殖物候、传粉及胚胎发育研究[J]. 植物科学学报, 2020, 38(5): 696 − 706. JIN Jiangqun, REN Fengmin, XIA Ying, et al. Research on reproductive phenology, pollination, and embryonic development of Thuja sutchuenensis Franch., a plant species with extremely small populations [J]. Plant Sci J, 2020, 38(5): 696 − 706. [12] COULTER J, LAND W. Gametophytes and embryo of Torreya taxifolia [J]. Bot Gaz, 1905, 39(3): 161 − 178. [13] 刘志敏, 赵宏波, 黄有军, 等. 香榧雌配子体发育和原胚形成的组织学观察[J]. 果树学报, 2017, 34(2): 231 − 237. LIU Zhimin, ZHAO Hongbo, HUANG Youjun, et al. Histological studies on megagametophyte and embryogeny development in Torreya grandis ‘Merrillii’ [J]. J Fruit Sci, 2017, 34(2): 231 − 237. [14] 高岚, 吕治平, 乐佳兴, 等. 中国特有裸子植物巴山榧树的保护生物学研究进展[J]. 重庆师范大学学报(自然科学版), 2019, 36(2): 127 − 132. GAO Lan, LÜ Zhiping, YUE Jiaxing, et al. Research progress of conservation biology on Torreya fargesii, a rare and eddangered plant endemic to China [J]. J Chongqing Norm Univ Nat Sci, 2019, 36(2): 127 − 132. [15] 李正理. 植物制片技术[M]. 北京: 科学出版社, 1987: 138 − 148. LI Zhengli. Plant Slicing Technology[M]. Beijing: Science Press, 1987: 138 − 148. [16] 徐明堂, 何春年, 张秀智, 等. 苏木精染液的配制及染色方法的改进[J]. 临床与实验病理学杂志, 2008, 24(3): 371 − 372. XU Mingtang, HE Chunnian, ZHANG Xiuzhi, et al. Improvement of preparation and dyeing method of Sumu fine [J]. Chin J Clin Exp Pathol, 2008, 24(3): 371 − 372. [17] LUOMAJOKI A. Temperature and dates of male meiosis in trees [J]. Hereditas, 2008, 97(2): 167 − 178. [18] 曹玉芳, 许方, 姚敦义. 松杉类植物雌雄配子体的发育与胚胎发生[J]. 莱阳农学院学报, 1995, 12(3): 206 − 212. CAO Yufang, XU Fang, YAO Dunyi. Development and embryogenesis of male and female gametophytes in Coniferae [J]. J Laiyang Agric Coll, 1995, 12(3): 206 − 212. [19] 曹志勇, 邵芬娟, 刘亚娟, 等. 杜松小孢子的发生和雄配子体的发育[J]. 林业科学, 2009, 45(3): 74 − 78. CAO Zhiyong, SHAO Fenjuan, LIU Yajuan, et al. Microsporogenesis and male gametophyte development of Juniperus rigida [J]. Sci Silv Sin, 2009, 45(3): 74 − 78. [20] 肖玉菲, 徐刚标, 梁文斌, 等. 南方红豆杉小孢子发生与雄配子体发育[J]. 广西植物, 2014, 34(6): 754 − 762. XIAO Yufei, XU Gangbiao, LIANG Wenbin, et al. Microsporogenesis and development of male gametophyte in Taxus chinensis var. mairei [J]. Guihaia, 2014, 34(6): 754 − 762. [21] 余象煜, 李平, 林亚康. 香榧小孢子叶球的发育及花粉超微结构研究[J]. 杭州大学学报(自然科学版), 1987, 14(3): 343 − 347. YU Xiangyu, LI Ping, LIN Yakang. Studies on the development of male cone and the ultrastructure of pollen in Torreya grandis [J]. J Hangzhou Univ Nat Sci, 1987, 14(3): 343 − 347. [22] 陈祖铿, 王伏雄. 穗花杉传粉和受精作用的研究[J]. 植物学报, 1985, 27(3): 239 − 245. CHEN Zukeng, WANG Fuxiong. A contribution to pollination and fertilization of Amentotaxus argotaenia [J]. Acta Bot Sin, 1985, 27(3): 239 − 245. [23] 俞晓敏, 赵桂仿. 太白红杉小孢子的发生和雄配子体的发育[J]. 植物学通报, 2003, 20(5): 576 − 584. YU Xiaomin, ZHAO Guifang. Microsporogenesis and male gametogenesis of Larix chinensis Beissn [J]. Chin Bull Bot, 2003, 20(5): 576 − 584. [24] FERNANDO D, LAZZARO M, OWENS J. Growth and development of conifer pollen tubes [J]. Sex Plant Rep, 2005, 18(2): 149 − 162. [25] 陈祖铿, 王伏雄. 穗花杉的胚珠结构与雌、雄配子体的发育[J]. 植物学报, 1985, 27(1): 19 − 25. CHEN Zukeng, WANG Fuxiong. The ovule structure and development of male and female gametophytes in Amentotaxus [J]. Acta Bot Sin, 1985, 27(1): 19 − 25. [26] ADERSON E D, OWENS J N. Megagametophyte development, fertilization, and cytoplasmic inheritance in Taxus brevifolia [J]. Int J Plant Sci, 1999, 160(3): 459 − 469. [27] 刘志敏. 香榧雌配子体的发育过程和败育研究[D]. 杭州: 浙江农林大学, 2015. LIU Zhimin. Megagametophyte Development and Abortion in Torreya grandis[D]. Hangzhou: Zhejiang A&F University, 2015. [28] 陈祖铿, 王伏雄. 白豆杉的配子体发育和受精作用[J]. 植物学报, 1979, 21(1): 9 − 29. CHEN Zukeng, WANG Fuxiong. The gametophytes and fertilization in Pseudotaxus [J]. Acta Bot Sin, 1979, 21(1): 9 − 29. [29] 徐刚标, 肖玉菲, 刘雄盛, 等. 濒危植物南方红豆杉大孢子发生和雌配子体发育[J]. 植物科学学报, 2015, 33(3): 271 − 280. XU Gangbiao, XIAO Yufei, LIU Xiongsheng, et al. Studies on megasporogenesis and development of megagametophytes in Taxus chinensis var. mairei [J]. Plant Sci J, 2015, 33(3): 271 − 280. [30] 陈佳妮, 廖亮, 黄增冠, 等. 香榧与榧树叶片光合特性及其光保护机制的比较[J]. 林业科学, 2015, 51(10): 134 − 141. CHEN Jiani, LIAO Liang, HUANG Zengguan, et al. A comparative study on photosynthetic characteristics and photoprotective mechanisms between Torreya grandis ‘Merrilli’ and Torreya grandis [J]. Sci Silv Sin, 2015, 51(10): 134 − 141. [31] 郑英茂, 周钰鸿, 潘成椿, 等. 濒危植物九龙山榧与其同属广布种榧树叶绿素荧光特性的比较[J]. 林业资源管理, 2021(1): 86 − 93. ZHENG Yingmao, ZHOU Yuhong, PAN Chengchun, et al. Comparison of chlorophyll fluorescence characteristics between Torreya jiulongshanensis leaves and its widespread congener T. grandis [J]. For Resour Manage, 2021(1): 86 − 93. [32] 梅方旭, 杨勤松, 张军. 香榧低产林改造技术要点[J]. 南方农业, 2021, 15(2): 141 − 142. MEI Fangxu, YANG Qinsong, ZHANG Jun. Key technical points of low-yield forest reconstruction of Torreya grandis ‘Merrillii’ [J]. South China Agric, 2021, 15(2): 141 − 142. [33] 刘萌萌, 曾燕如, 江建斌, 等. 香榧生长期叶片和种子中矿质元素动态变化研究[J]. 浙江农林大学学报, 2014, 31(5): 724 − 729. LIU Mengmeng, ZENG Yanru, JIANG Jianbin, et al. Mineral elements in leaves and seeds of Torreya grandis ‘Merrillii’ during seed development [J]. J Zhejiang A&F Univ, 2014, 31(5): 724 − 729. [34] FAGERIA N K, FILHO M P B, MOREIRA A, et al. Foliar fertilization of crop plants [J]. J Plant Nutr, 2009, 32(6): 1044 − 1064. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220181
 点击查看大图
点击查看大图

计量
- 文章访问数: 1865
- HTML全文浏览量: 609
- PDF下载量: 87
- 被引次数: 0
