




-
同一块土地连续种植同一作物或近缘作物,即使在正常的栽培管理状况下,也会出现植株长势变弱、病虫害严重、作物产量降低、品质下降等的现象为连作障碍[1],很多作物都存在这一现象[2−5]。目前,中国连作危害程度高的地块面积超过10%,当季作物损失占20%~80%,严重的甚至几乎绝产。连作障碍每年造成数百亿元的经济损失,同时还降低了农作物的安全性[6]。作物轮作、间作、套作等增加生物多样性的方法是解决连作障碍的有效措施,但都仅是短期内的改善,且操作上存在诸多不便,推广应用不理想。中国土地利用指数高、经营强度大,同一地区集中连作1~2种植物是当下农业生产的主要模式[7]。因此,目前提出的解决连作障碍的技术与方法,难以从本质上解决连作障碍问题[8]。以“作物连作障碍”为关键词,统计1989—2018年的结果表明:突现度(某变量出现频次在较短时间内突然增加)排名前二位的分别是黄豆Glycine max (35.3094)、马铃薯Solanum tuberosum(28.9866)[9],说明这2种植物连作障碍问题严重。而禾本科植物连作障碍的研究报道极少,说明实际生产中禾本科Poaceae植物很少出现连作障碍。禾本科作物还常常作为连作障碍首选的间作或轮作对象,如对枸杞Lycium chinense与禾草间作研究发现:土壤理化性质、土壤酶活性、枸杞生长速率和产量以及禾草生物量等指标均表现为间作高于枸杞单独连作,间作对枸杞分枝的促进作用尤为显著[10]。胡麻Linum usitatissimum与小麦Triticum aestivum轮作可减弱土壤水提液对胡麻种子萌发以及生长的自毒作用,利于胡麻的生长[11]。可见,不同植物耐受连作障碍的能力不同。
土壤酶是大部分物质转化过程的执行者[12],土壤酶活性是衡量土壤肥力的重要指标之一[13]。在植物生长过程中,根系不断分泌代谢产物到根际土壤,对土壤微生物活动产生显著影响,从而影响土壤酶活性,由此改变了根际环境土壤的理化性质,特别是各种养分的生物有效性以及养分的转化速率[14]。随着连作时间的延长,植株细胞膜的通透性增加,养分物质运输功能下降,引发自毒作用,作物的正常生长也受到影响[15],虽然对连作障碍这一农业生产老大难问题已经开展了大量研究[9],但同时比较多科植物连作过程中生物化学变化的研究未见报道。为此,本研究通过比较多科植物连作过程中土壤酶活性的差异变化,假设植物遗传特性是耐连作障碍的重要因子,为耐连作障碍的发生机制研究提供证据。
-
以浙江农林大学实习基地的森林土壤和育苗基质1∶1的体积比混合均匀后为盆栽本底土,该土结构疏松、质地优良,具有良好的保水保肥效果。森林土壤基本理化性质:pH 5.21,速效钾为193.33 mg·kg−1,碱解氮为51.17 mg·kg−1,有效磷为6.00 mg·kg−1,全氮为1.33 g·kg−1,全碳为28.50 g·kg−1。育苗基质基本理化性质:有机质质量分数大于45%,pH 5.50~7.00,含少量的氮磷钾物质。混合后的土pH 5.42。植物包括:豆科Fabaceae花生Arachis hypogaea、黄豆Glycine max,葫芦科Cucurbitaceae西瓜Citrullus lanatus、南瓜Cucurbita moschata,茄科Solanaceae番茄Solanum lycopersicum、马铃薯Solanum tuberosum,禾本科Poaceae早熟禾亚科Pooideae二穗短柄草Brachypodium distachyon (Bd-21,模式植物)、黑麦草Lolium perenne、小麦Triticum aestivum,黍亚科Pamicinae玉米Zea mays、高粱Sorghum bicolor。选择饱满,大小均匀的种子,所有种子在播种前均经过氧化氢灭菌消毒和催芽。
-
进行单一作物长期连作的盆栽试验,每种植物至少6盆(即重复),连续种植3季。考虑不同植物的生物量尽量一致的原则,花生、黄豆、西瓜、南瓜、番茄、马铃薯、二穗短柄草、黑麦草、小麦、玉米、高梁分别栽培5、5、5、5、7、2、10、40、10、5、5 株·盆−1,所选植物生长均匀,长势一致。
-
于浙江农林大学人工气候箱进行试验,气候箱栽培条件为25 ℃、光照16 h,18 ℃、黑暗 8 h的昼夜循环,生长旺季时温度调至30 ℃。土壤水分基本保持在75%的田间持水量,其他栽培管理措施按一般要求进行,各盆保持一致。注意抗旱、防病虫,保证全苗及植株正常生长发育。
种植时间和采样时间如表1所示,每茬植物的收获时间处于该植物营养生长时期,根据植物生长的情况可适当调整采样时间,栽培时长为38~55 d,共采集3茬,鉴于本底土壤速效钾质量分数高、而氮磷质量分数低的实际情况,在第1季时选择磷酸二铵[(NH4)2HPO4]作为追肥[氮磷钾质量比(N∶P2O5∶K2O)为18∶46∶0]。肥料按照肥质量比1∶50进行配置,每盆每次浇50 mL,均以每季植物能正常生长为依据适当补施肥料,每季浇施3次。选择生长一致的4个重复,分别采集第1季和第3季的土壤样品,分析土壤酶以及土壤性质。收获时植物根系基本充满整个盆,密布于土壤之间,植物根系对土壤的影响涉及大部分土壤,可以认为盆中土壤均为根区土壤。将植物取出后的盆中土壤充分混匀后得到混合样品,风干待测。
表 1 植物栽培和收获时间表
Table 1. Schedule for transplanting and harvesting
栽植季度 栽培日期
(年-月-日)采样日期
(年-月-日)采收植物 第1季 2020-12-02 2021-02-02
2021-02-07
2021-02-20
2021-03-01
2021-03-08黄豆、花生
高粱、玉米
马铃薯、小麦
二穗短柄草、黑麦草
番茄、西瓜、南瓜第2季 2021-03-18 2021-04-29
2021-05-02
2021-05-07
2021-05-14
2021-05-17黄豆、玉米
花生、高粱
西瓜、南瓜、番茄
马铃薯、二穗短柄草
黑麦草、小麦第3季 2021-05-31 2021-07-14
2021-07-20
2021-07-21
2021-07-23玉米、番茄
黄豆、花生、马铃薯
西瓜、南瓜、高粱
黑麦、小麦、二穗短柄草 -
测定土壤碳、氮、磷循环的3类土壤酶:①碳循环土壤酶,包括α-葡萄糖苷酶(AG)、β-葡萄糖苷酶(BG)、纤维二糖水解酶(CB)、β-木糖苷酶(XYL);②氮循环土壤酶,包括亮氨酸氨基肽酶(LAP)、N-乙酰-β-氨基葡萄糖苷酶(NAG);③ 磷循环土壤酶,为酸性磷酸酶(PHOS)。酶活性测定采用SAIYA-CORK等[16]的荧光微孔板检测技术,具体测定方法:称取2 g土壤,加入醋酸缓冲液进行浸提,取200 μL浸提液与96孔板后立即加入反应底物,25 ℃ 培养箱中黑暗培养3 h,使用多功能酶标仪(SynergyTM H1,Biotek)在365和450 nm波长下测定吸光度并计算土壤酶活性。
-
重点分析与土壤碳、氮、磷循环相关酶活关联最大的pH、碱解氮和有效磷等3个指标(表2)。与本底土壤pH 5.42相比,除玉米土壤外,其他植物种植1季后,土壤pH普遍下降,其中豆科植物下降幅度最小,而西瓜最大;第3季又比第1季下降,所有土壤pH均在4.5以下,土壤呈强酸性。连作后土壤碱解氮及有效磷明显增加,第1季土壤碱解氮质量分数西瓜最低、玉米最高,有效磷质量分数则是南瓜最高、小麦最低。第3季时土壤有效磷质量分数比第1季大幅增加,而碱解氮则有增有减。
表 2 第1季和第3季植物收获后不同植物根区土壤化学性质比较
Table 2. Comparison of soil chemical properties in root zone of different plants after harvest in the first and third seasons
植物种类 pH 碱解氮/(mg·kg−1) 有效磷/(mg·kg−1) 第1季 第3季 第1季 第3季 第1季 第3季 花生 5.03±0.05 bc 4.20±0.08 abc 561.65±60.70 cde 705.83±58.59 a 149.89±17.64 e 651.52±21.10 a 黄豆 5.23±0.17 ab 4.10±0.08 abc 567.42±34.93 bcd 577.84±30.07 ab 161.44±30.06 de 355.58±63.73 ef 西瓜 4.23±0.25 e 3.97±0.05 bc 492.05±46.00 e 672.01±96.79 ab 161.36±30.12 de 393.66±34.33 def 南瓜 4.90±0.22 cd 4.13±0.21 abc 664.13±22.70 ab 553.72±108.19 ab 323.48±17.76 a 577.35±93.74 ab 番茄 4.93±0.09 bc 4.13±0.12 abc 647.74±56.66 abc 520.56±131.32 abc 292.47±18.44 ab 478.03±0.02 bcde 马铃薯 4.77±0.05 cd 3.97±0.05 bc 530.86±10.41 de 360.96±12.77 c 267.34±49.73 abc 304.40±51.93 f 二穗短柄草 4.57±0.17 d 3.93±0.12 c 540.13±39.37 de 356.81±20.47 c 245.17±16.65 bc 425.49±40.00 cdef 黑麦 4.73±0.12 cd 4.23±0.12 ab 593.97±29.02 bcd 667.47±82.33 ab 218.97±46.98 cd 546.70±60.91 abc 小麦 4.83±0.05 cd 4.23±0.12 ab 567.93±26.81 bcd 687.14±71.14 ab 114.48±11.85 e 524.25±41.53 abc 玉米 5.43±0.17 a 4.27±0.21 a 699.27±75.11 a 546.80±114.25 ab 288.63±45.34 ab 599.87±42.06 ab 高粱 4.77±0.09 cd 4.13±0.05 abc 397.89±24.82 f 509.08±23.38 bc 124.26±3.98 e 485.14±24.71 bcd 说明:不同小写字母表示不同植物间差异显著(P<0.05)。 -
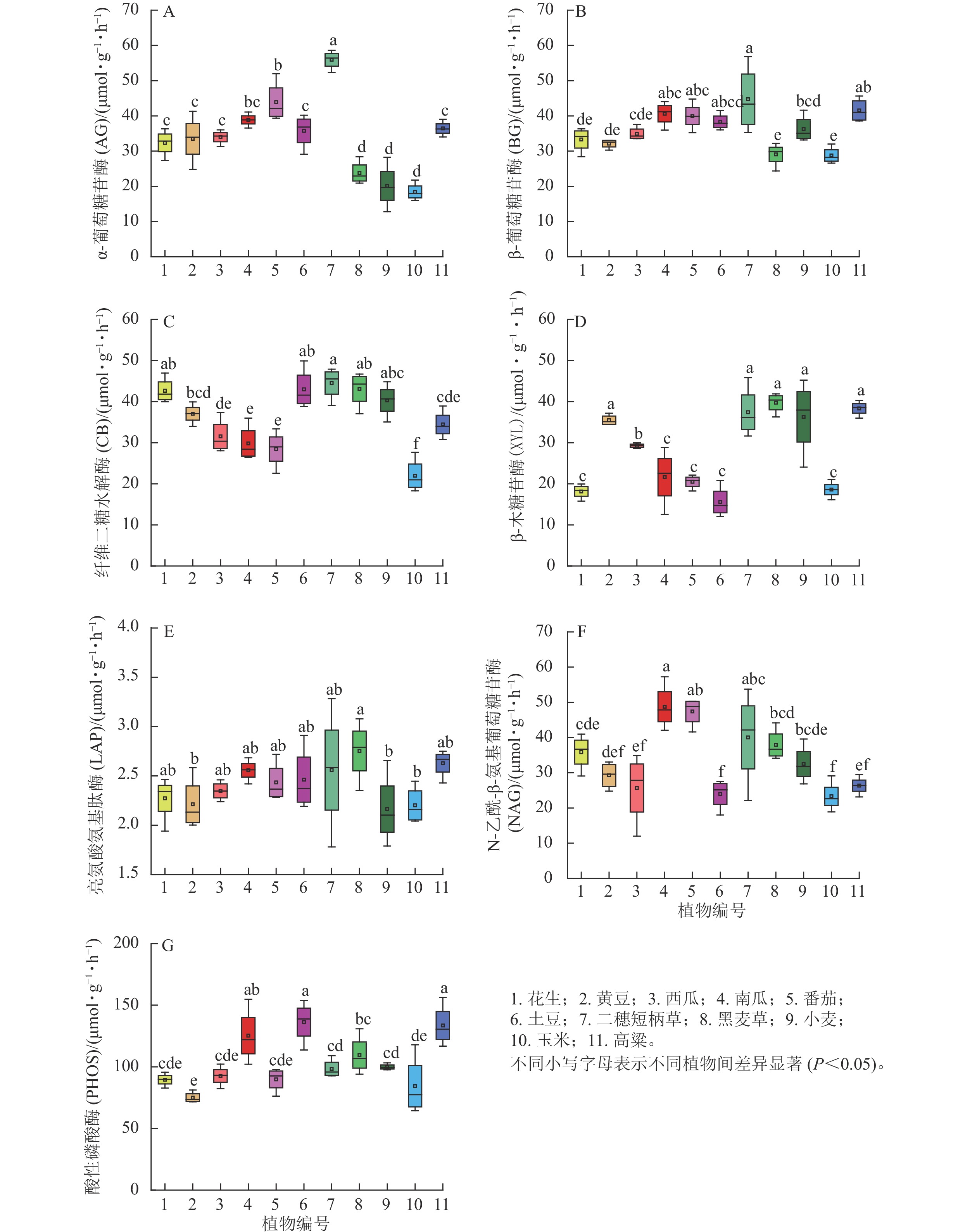
第1季土壤碳循环相关的AG和BG活性水平相近(图1 A和B), AG活性较高的有南瓜、番茄、黑麦草和玉米,而小麦和高粱最低;BG活性较低为黄豆、马铃薯和玉米,较高组为花生、南瓜、黑麦草和高粱,同科植物均存在较大差异,甚至达显著水平(P<0.05)。CB活性总体低于AG和BG,不同植物之间的高低水平与AG基本一致; XYL活性最高的是马铃薯,最低是黑麦草(图1C和D)。综上所知,种植1季后,不同植物之间4种土壤碳循环酶活性高低顺序没有显著分异,相对而言,几种禾本植物的波动较大,特别是玉米在所有植物中的排序处于最高(AG和CB)或最低(BG)的位置。与土壤氮循环相关的LAP和NAG活性其重复之间的变异比碳循环酶活性小,但南瓜土壤LAP活性异常高,其他植物之间没有显著差异(图1E);NAG活性最高的是高粱,最低是马铃薯和二穗短柄草(图1F)。PHOS活性在不同植物之间的差异是所有酶中最小的,总体而言,禾本科植物高于非禾本科植物,其中最高的还是高粱(图1G)。
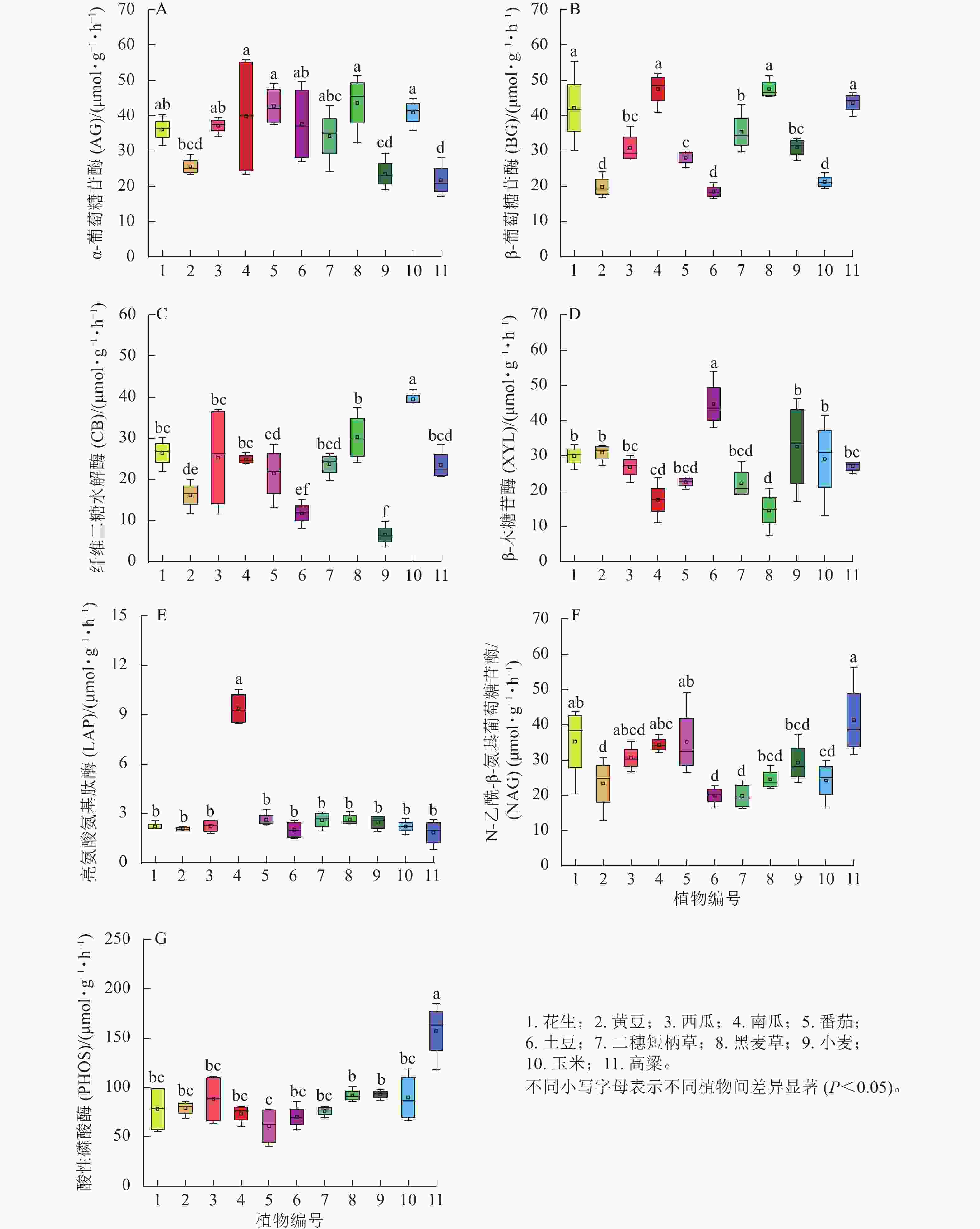
图 1 第1季不同植物土壤根区酶活性
Figure 1. Soil rhizosphere enzyme activities of different plants after the first season
分析第1季所有植物土壤酶活性之间的相关性结果发现:显著相关性只存在于AG与PHOS(R=−0.647)、BG与XYL (R=−0.605)及LAP (R=0.653)、CB与XYL (R=−0.704),说明不同植物对土壤酶活性影响强度、甚至方向不相同,显著负相关(3组)多于显著正相关(1组)。
-
第3季植物土壤酶活性,除XYL外,土壤酶活性的重复之间变异也显著缩小(图2),其主要规律为非禾本科植物AG高于禾本科植物、而CB则相反;不同植物之间土壤BG活性差异不显著;禾本科植物XYL活性总体高于非禾本科植物。综上所知,种植3季植物后,二穗短柄草土壤几种碳循环相关酶活性总体较高,而玉米则总体较低。不同植物土壤2种氮循环酶活性没有一致的差异规律。土壤磷循环相关的PHOS活性则是同科的不同种出现一高一低相反的结果,二穗短柄草、黑麦草和小麦之间差异较小。
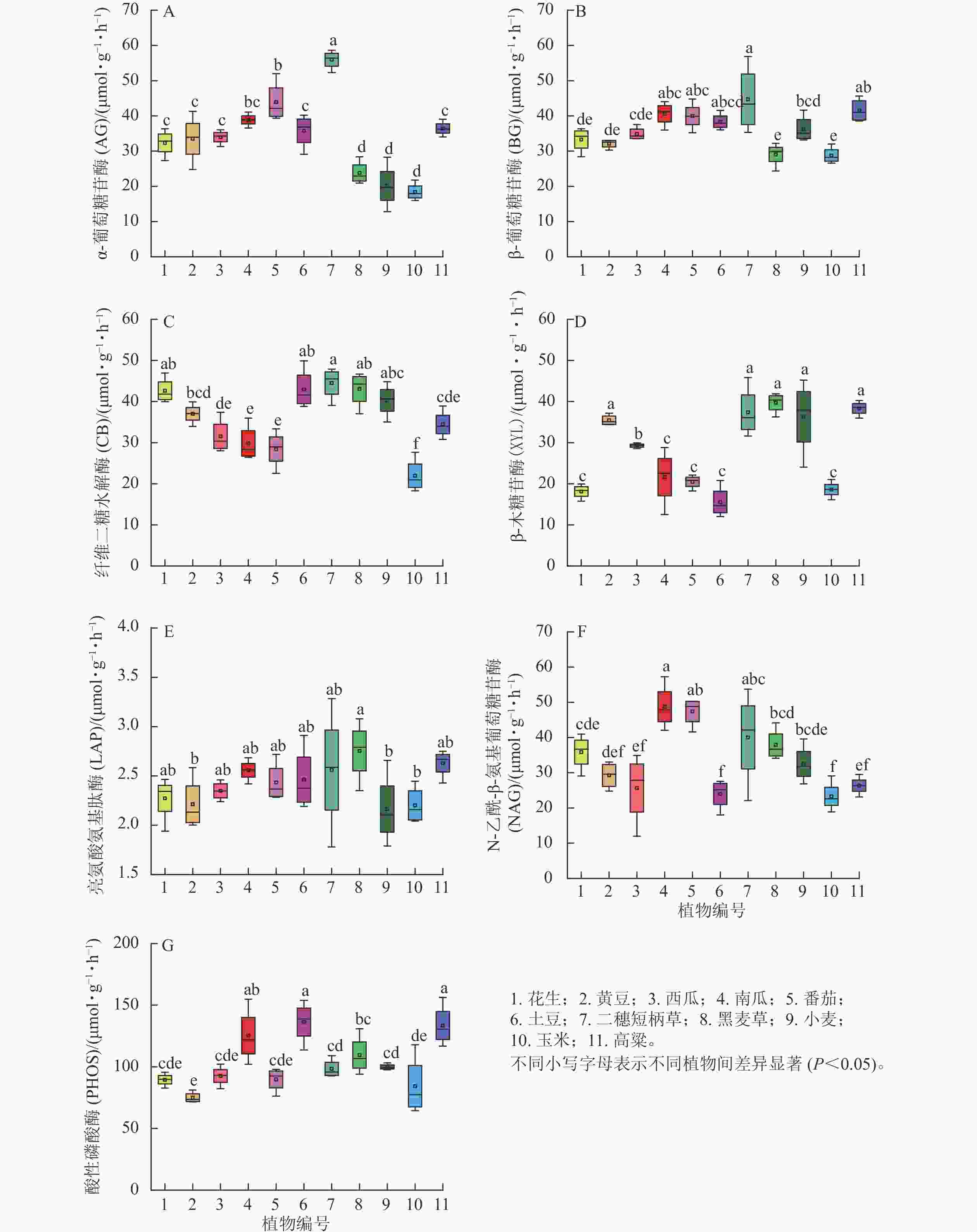
图 2 第3季不同植物土壤根区酶活性
Figure 2. Soil rhizosphere enzyme activities of different plants after the third season
分析第3季所有植物7种土壤酶活性之间的相关性结果发现:只有AG与BG (R=0.763)、LAP与PHOS (R=0.642)之间显著相关,说明植物对不同土壤酶活性影响分异性增加。
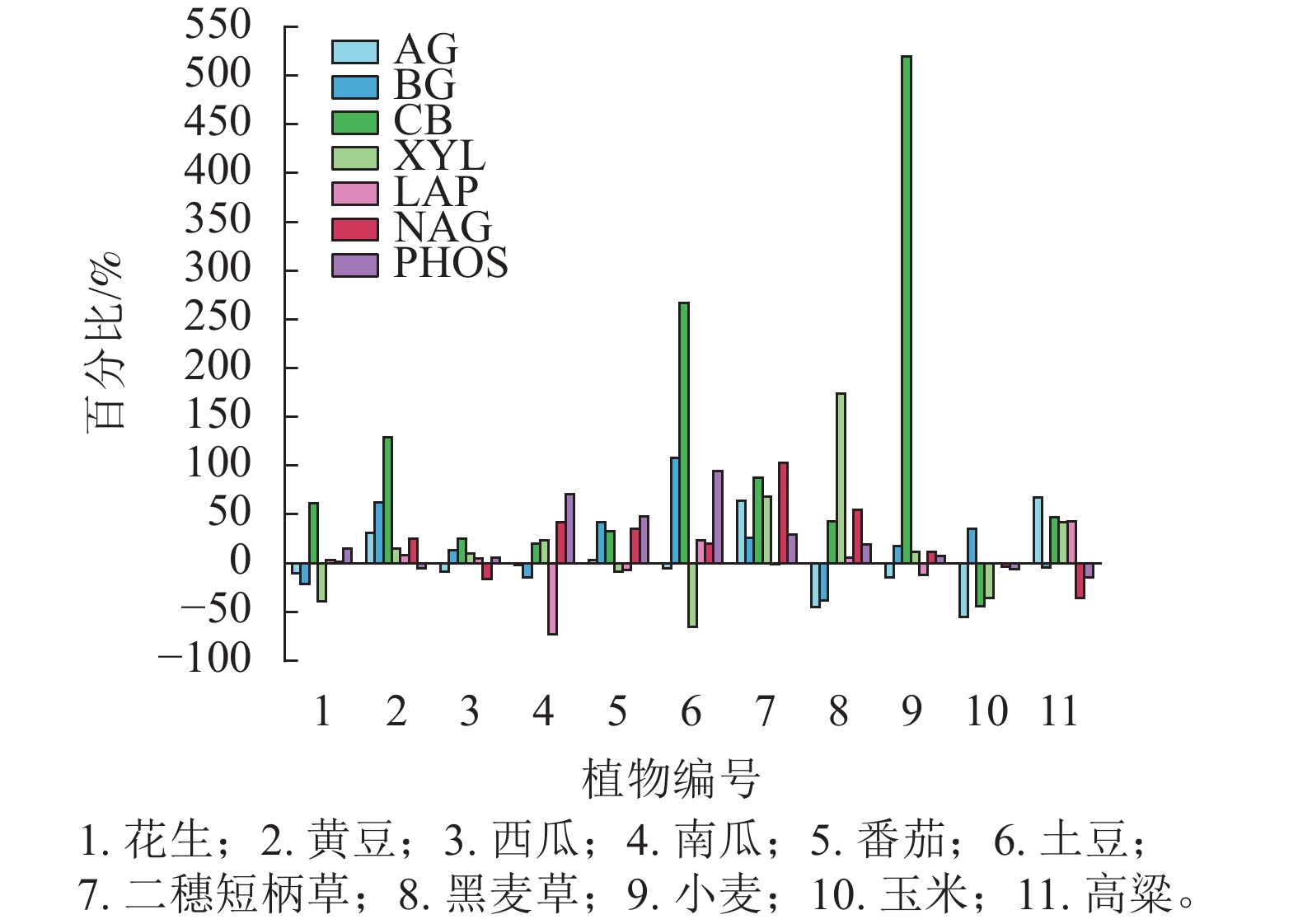
分析土壤酶的动态变化(图3)发现:与第1季相比,第3季植物收获后土壤酶活性多数呈增加趋势,其中CB活性的增幅最明显,而玉米则除BG外其余土壤酶活性均出现降幅。
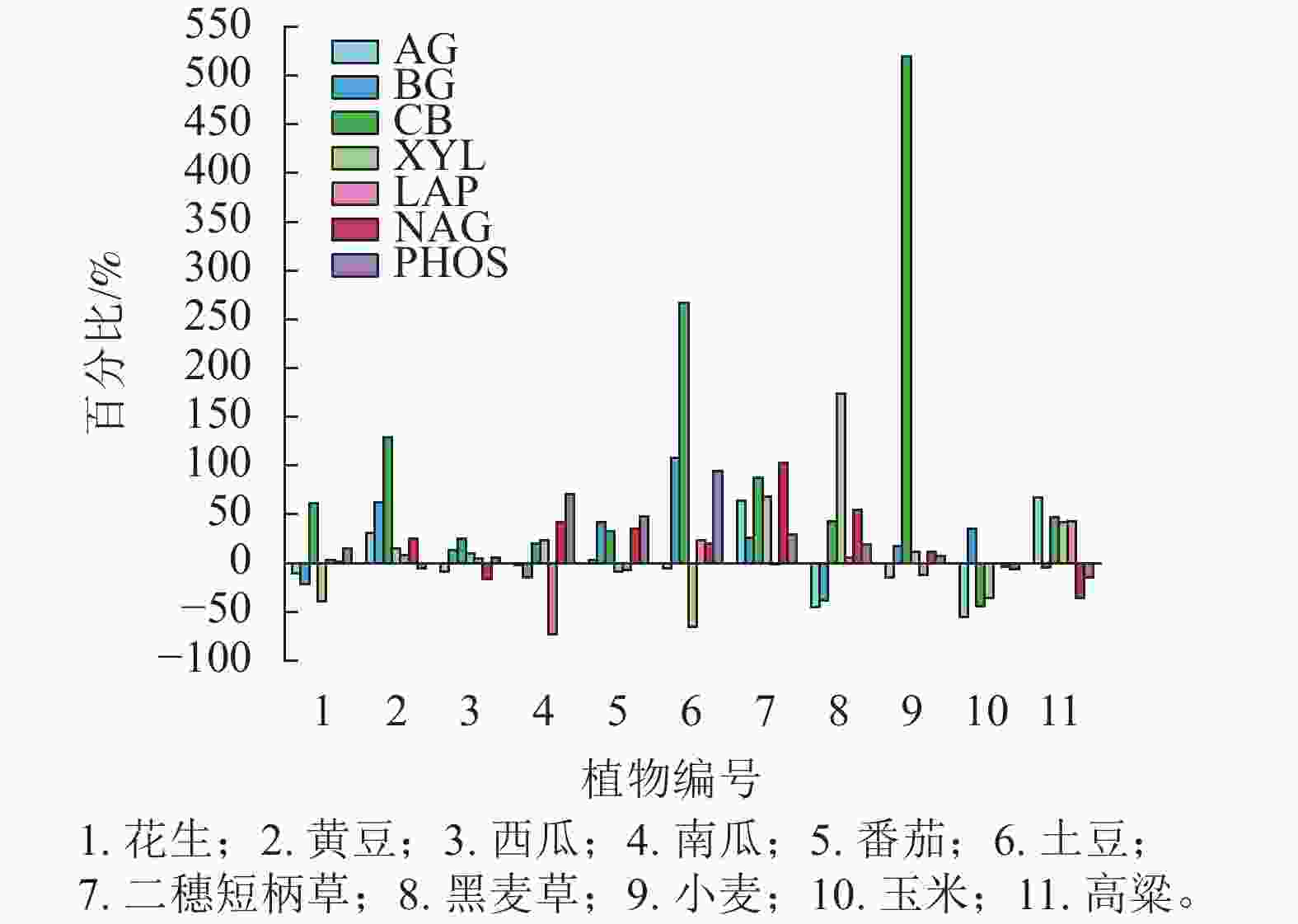
图 3 不同植物第1季和第3季土壤酶活性平均增幅或减幅
Figure 3. Average increase or decrease percentage of soil enzyme activities in the first and third plant seasons
-
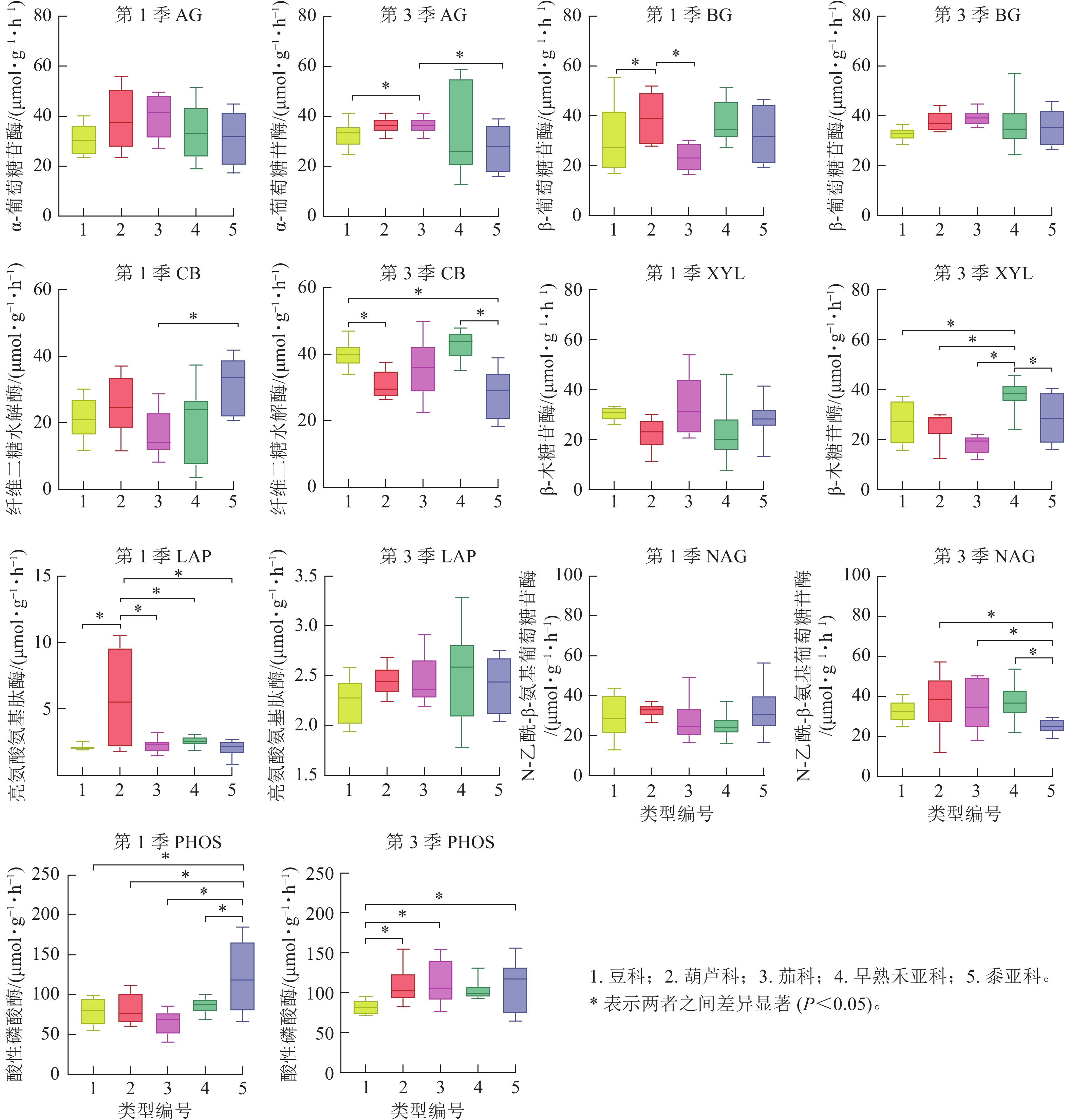
将非禾本科植物以科为单位、禾本科以亚科为单位进行比较。禾本科包括二穗短柄草、黑麦和小麦等组成的早熟禾亚科(C3植物)以及由玉米和高粱组成的黍亚科(C4植物)。由图4发现:第1季不同科或亚科之间AG活性无显著差异,而第3季的茄科显著高于豆科和黍亚科(P<0.05)。BG活性第3季各科、亚科之间无显著差异,第1季的葫芦科显著高于豆科和茄科(P<0.05),茄科的平均值最低。 第1季土壤的CB活性表现为黍亚科显著高于茄科(P<0.05),而第3季则是黍亚科显著低于早熟禾亚科及豆科(P<0.05),豆科显著高于葫芦科(P<0.05)。XYL活性第1季无差异,而第3季早熟禾亚科显著高于其他科(P<0.05)。土壤氮循环酶LAP活性第1季的葫芦科显著高于其他科(P<0.05),而第3季却没有显著差异;NAG活性第1季无差异,第3季黍亚科显著低于豆科、葫芦科和早熟禾亚科(P<0.05)。PHOS活性第1季时黍亚科显著高于其他科(P<0.05),第3季则是豆科显著低于其他科(P<0.05)。
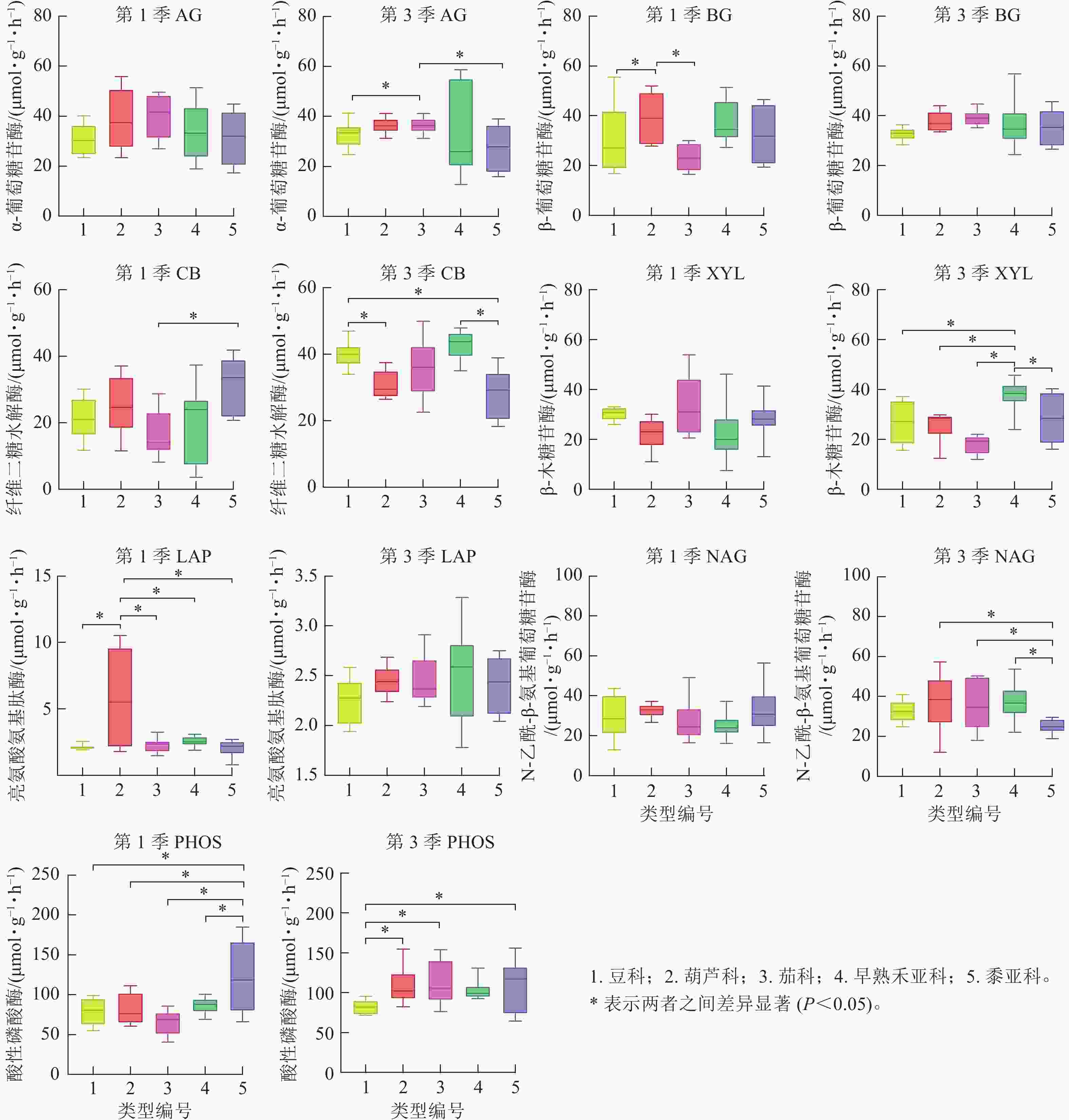
图 4 不同科及禾本科亚科第1季和第3季植物根区土壤碳循环酶活性的差异
Figure 4. Difference of carbon cycling enzyme activity between family or subfamily of rhizosphere soil in the first and third seasons
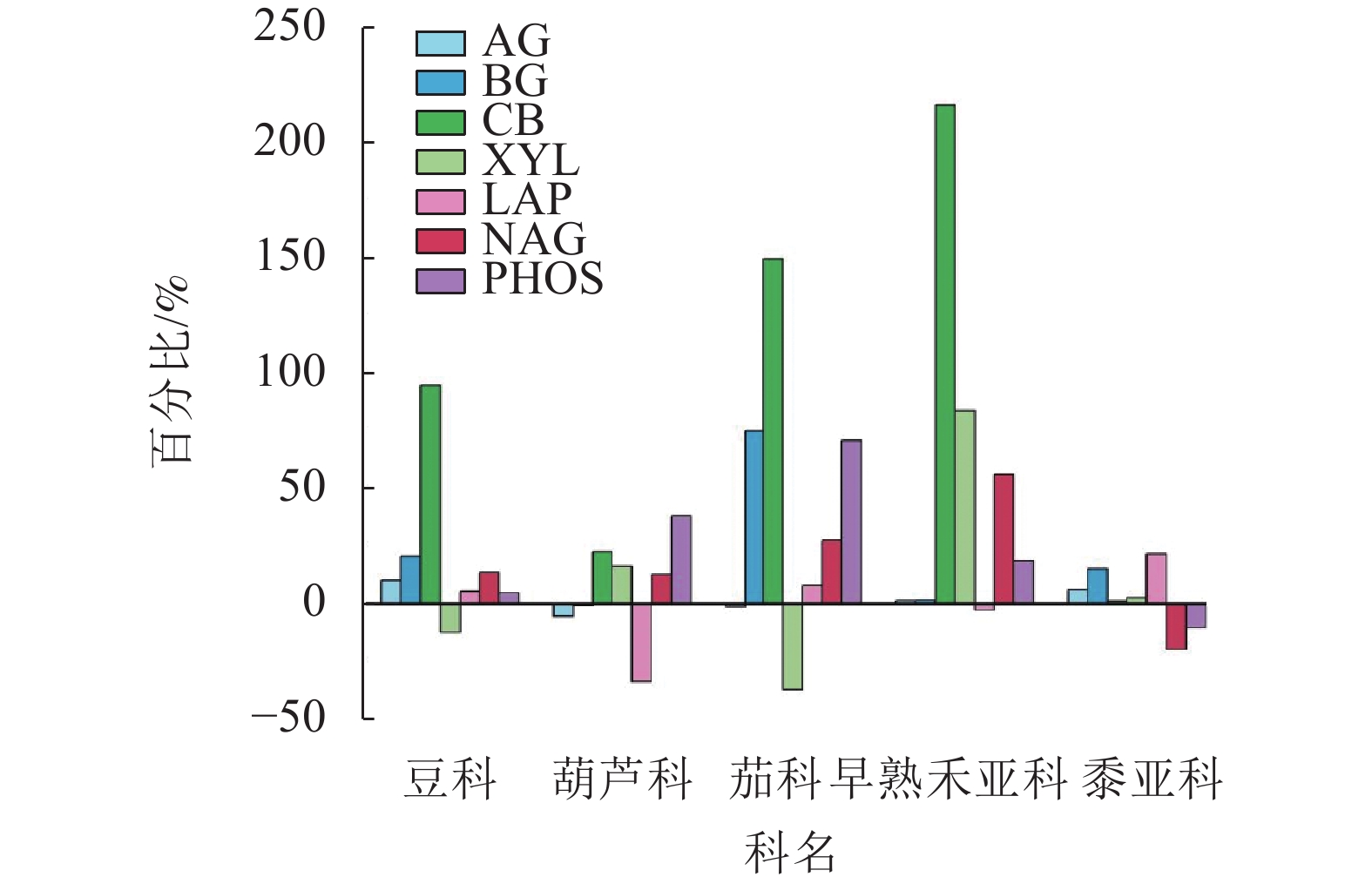
以科或亚科为单位统计分析的动态变化结果(图5)表明:多科植物不同酶活性大多增加;豆科、茄科和早熟禾亚科增幅最大的是CB,葫芦科增幅最大的是PHOS,黍亚科增幅最大的是LAP。
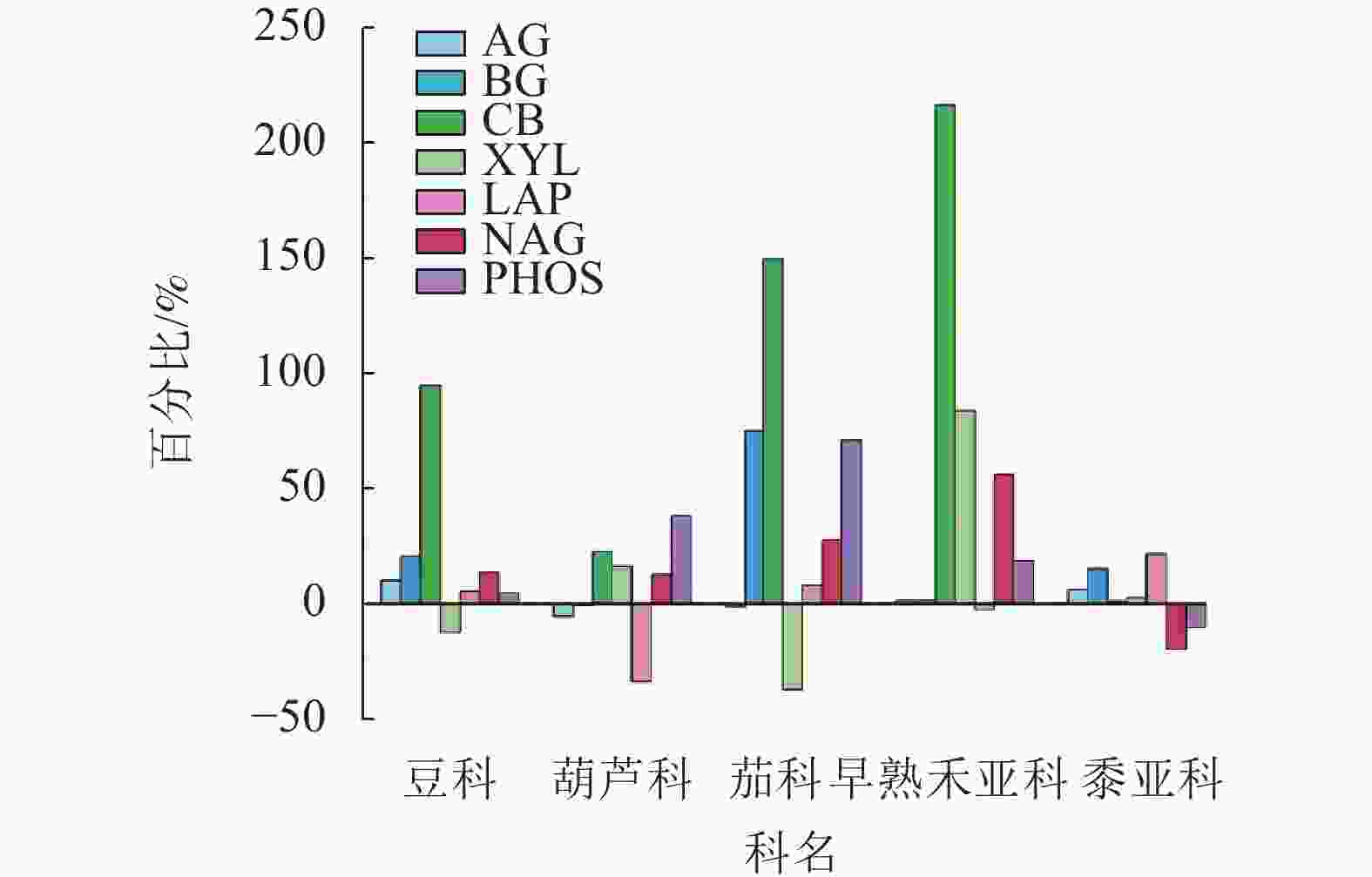
图 5 不同科植物第1季和第3季土壤酶活性平均增幅或减幅
Figure 5. Average increase or decrease percentage of soil enzyme activities in the first and third plant seasons based on plant family
-
种植1季植物后不同植物土壤的多项性质产生差异变化,pH 5.00以上的花生、黄豆和玉米,其中玉米pH最高,而西瓜和二穗短柄草pH较低,分别为4.23和4.57。土壤碱解氮的差异规律与pH基本一致,说明植物对氮利用越多,则土壤pH下降越大(西瓜和高粱),这一结果符合氮肥引起土壤酸化的结论[17]。第1季后玉米土壤各项养分指标及pH均处于较高水平,可能是其吸收利用养分相对较少的结果,与高粱、小麦土壤差异明显。另外,除BG外,玉米土壤的其他3种碳循环酶活性均较高,而小麦则较低。玉米土壤酶活性与土壤养分呈正相关,与已有研究结果[18]一致。玉米和小麦土壤AG活性与BG活性相反,说明高水平土壤养分不利于BG活性发挥。BG水解结合于末端非还原性的β-D-葡萄糖键,与纤维素降解有关,由于纤维素的贫营养特征,因此,分解纤维素的微生物能够长期适应低养分水平土壤环境[19]。番茄和马铃薯土壤也是AG活性高,BG活性低,土壤氮磷水平总体与玉米相近。除了土壤养分外,植物也能通过根系分泌物影响土壤微生物,从而间接影响土壤酶活性。土壤LAP活性除南瓜异常高外,其他植物之间差异不大。南瓜土壤LAP活性异常高的原因可能与其根系分泌物存在诱导LAP活性提高的成分。南瓜被广泛用作砧木以缓解西瓜、黄瓜、甜瓜、冬瓜和苦瓜等多种植物连作障碍[20−24],这与南瓜土壤较高的LAP活性有关。高粱土壤的NAG活性最高,BG活性也最高,NAG与几丁质降解有关,几丁质是真菌细胞壁的主要成分。高粱土壤可能存在较丰富的纤维素[25],促进降解纤维素的真菌大量繁殖,从而积累含几丁质的大量真菌生物残体,诱导NAG活性增加。高粱土壤PHOS活性显著高于其他植物,已有研究表明:土壤有效磷较低可诱导PHOS活性[26−28],由此推测,高粱土壤PHOS活性显著高是因为其土壤有效磷低于其他植物。本研究土壤PHOS活性总体均低于其他研究报道[29−30]。
种植3季作物后,土壤pH较第1季下降0.50~1.16,玉米仍然是所有植物中最高(4.27),而西瓜、马铃薯和二穗短柄草则低于4.0;土壤碱解氮、有效磷显著增加,特别是有效磷增加了2~3倍,原因是本底土壤有效磷很低(6.00 mg·kg−1),氮素水平也偏低(碱解氮51.17 mg·kg−1),而速效钾丰富(193.33 mg·kg−1),因此种植过程中追施高磷的磷酸二铵肥料以保证植物养分需求。第3季时土壤养分水平过高对土壤酶活性产生一定的负面影响,而二穗短柄草虽然其土壤pH仅3.93,但土壤氮磷水平较低,反而其各种土壤酶活性均较高;马铃薯土壤pH、养分水平与二穗短柄草相近,除XYL和NAG酶外,其他5种酶活性也较高。相反的情况发生在玉米,土壤养分水平较高但土壤各种酶活性相对较低,说明高水平养分对玉米土壤酶活性的负面影响比其他植物强。从第3季土壤酶的增减幅度结果证明:玉米土壤仅BG活性有小幅增加,其他6种酶均下降;而CB活性总体增幅最大,其中小麦和马铃薯分别增加519%和267%。CB水解纤维素释放纤维二糖,其增幅最大的原因可能:一是连作后真菌数量增加[31−32],真菌是分解纤维素的最主要类群; 二是随着种植次数增加,土壤中纤维素不断积累,诱导微生物分泌CB。这与王雨晴等[33]的研究一致,即CB活性与纤维素含量及真菌基因丰度呈极显著正相关。
-
禾本科植物存在着广泛的基因序列和功能的保守性[34],其他同科植物也有共性的遗传基因。一般而言,遗传相近的植物具有相似的根系分泌物、对养分的需求也较相似,由此推测,同一科植物对土壤微生物以及土壤酶活性可能有共性影响。第1季植物收获后,只有BG (葫芦科显著高于豆科和茄科)和CB (黍亚科显著高于茄科) 2种酶存在科之间的显著差异,而第3季收获后仅BG和LAP不存在显著差异,说明随着种植季数的增加,同科植物对土壤的共性影响逐渐显现。BG和CB都是降解纤维素的酶,最初的本底土壤微生物较少,而种植1季后土壤微生物大量繁殖,由于不同植物根系分泌物以及根系残体化学组成的差异,特别是根系残体中纤维素较多,由此引起不同科植物纤维素降解相关酶的差异。第3季时不同科植物土壤酶活性之间的差异加大,虽同为早熟禾亚科植物,但二穗短柄草在遗传上与小麦及黑麦草距离相对较远,因此第3季时其土壤AG和BG活性明显高于小麦及黑麦草。 第3季时非禾本科植物土壤AG活性明显高于禾本科植物,最高是茄科,而XYL则正好相反,其中最高的是早熟禾亚科。AG和XYL都是大分子有机物水解的末端酶,非禾本科植物土壤AG活性高,说明土壤中其上游大分子有机物中α-葡萄糖苷链接的多糖(如淀粉)较多,而禾本科植物土壤XYL活性高,说明土壤中上游大分子有机物中β-木糖苷酶链接的半纤维素多糖较多。也可能是禾本科植物的根系分泌物与非禾本科植物存在差异[35]。禾本科植物中早熟禾亚科土壤CB活性较高,但黍亚科则明显低于非禾本科植物,可能C3和C4植物的碳代谢物存在差异,需要后继分析根系分泌物。不同科植物第3季土壤CB和XYL活性的高低与第1季正好相反,豆科和茄科土壤CB活性明显升高,而黍亚科下降,茄科土壤XYL活性明显下降,而早熟禾亚科升高,说明CB和XYL可能存在互补关系。另外,禾本科植物土壤的XYL活性显著高于非禾本科植物,可能是禾本科植物土壤中半纤维素含量较丰富。XYL水解半纤维素产生种类丰富的五碳糖和六碳糖,是否有利于土壤微生物活动值得深入研究。黍亚科土壤PHOS活性在第1季和第3季土壤中均为最高,而其土壤NAG活性在第3季为最低。分科统计则土壤酶活性总体以上升为主,其中CB活性增幅最大,而豆科和茄科的XYL活性、葫芦科的LAP活性出现下降,黍亚科植物土壤的NAG活性和PHOS活性都出现下降,说明黍亚科植物土壤的氮、磷循环能力下降,可能是氮、磷养分增加显著,加上黍亚科植物对氮、磷的需求不高,因此,相关循环酶活性下降。
-
通过不同科11种植物第1季和第3季土壤碳、氮、磷循环相关酶活性分析发现:同科的2种植物不同土壤酶、同种土壤酶不同科植物的动态变化规律并不一致,第1季和第3季黍亚科植物土壤PHOS活性均为最高,其中,高粱又显著高于比玉米;第3季时禾本科植物土壤的XYL活性显著高于非禾本科植物,而3种早熟禾亚科植物土壤又显著高于黍亚科植物。禾本科植物土壤PHOS和XYL活性较高是否与其耐连作有关,需要深入研究连作次数和土壤微生物指标等。
Differential response of soil enzyme activity to continuous cropping of different plants
-
摘要:
目的 连作障碍给农业生产造成巨大的经济损失,而不同植物耐受连作障碍能力存在差异,特别是禾本科 Poaceae植物很少有连作障碍的研究报道。为此,从植物角度出发,分析不同植物(科)连作3季后7种土壤酶活性的变化规律,旨在探究不同植物耐连作障碍的机制。 方法 分别选择豆科Fabaceae、葫芦科Cucurbitaceae、茄科Solanaceae每科各2种以及5种禾本科植物模拟连作的盆栽试验,分别采集第1季和第3季植物收获后的根区土壤,分析土壤pH,有效氮、磷,碳、氮、磷循环相关酶活性。 结果 不同植物3季连作后土壤pH呈下降趋势,第1季和第3季玉米Zea mays pH始终最高,而西瓜Citrullus lanatus最低。土壤有效氮、磷质量分数明显增加,大部分植物土壤第3季时有效氮、磷质量分数均在500 mg·kg−1以上。第1季植物土壤酶活性没有呈现差异规律,第3季非禾本科植物土壤α-葡萄糖苷酶(AG)高于禾本植物,而纤维二糖水解酶(CB)正好相反。不同科植物之间、甚至同科的2种植物之间土壤酶变化规律并不一致。比较明确的规律是:第1季和第3季,黍亚科Panicinae植物(玉米和高粱Sorghum bicolor)土壤酸性磷酸酶(PHOS)活性均最高,第3季禾本科植物土壤β-木糖苷酶(XYL)活性显著高于(P<0.05)非禾本科植物,而早熟禾亚科Pooideae植物(二穗短柄草Brachypodium distachyon、小麦Triticum aestivum、黑麦草Lolium perenne)土壤又普遍高于黍亚科植物。土壤酶总体呈动态增加趋势,其中CB、PHOS和亮氨酸氨基肽酶(LAP)活性分别为早熟禾亚科、葫芦科和黍亚科增幅最大;同属禾本科的黍亚科和早熟禾亚科植物土壤PHOS活性分别呈下降和上升的相反趋势。 结论 禾本科植物土壤PHOS和XYL活性较高,其他酶活性并没有表现出明显的同科变化规律。图5表2参35 Abstract:Objective Continuous cropping obstacles have caused huge economic losses to agricultural production. There are differences in the ability of plants to tolerate continuous cropping obstacles. Few studies have been reported on continuous cropping obstacles of Poaceae plants. This study tries to explore the mechanisms of different plants’ resistance to continuous cropping by investigating 7 soil enzymes in different plants (families) after 3 seasons of continuous cropping. Method Two species of plants from each family of Fabaceae, Cucurbaceae, Solanaceae and 5 species of Poaceae family were selected for simulated continuous pot experiment. Soil in the root zone of the plants in the 1st and 3rd seasons was collected after harvest to analyze the soil pH, available nitrogen (AN) and phosphorus (AP) and the activities of enzymes related to carbon, nitrogen and AP cycling. Result The soil pH showed a decreasing trend after continuous cropping of different plants for 3 seasons, among which the soil pH of Zea mays in the 1st and 3rd seasons was always the highest, while that of Citrullus lanatus was the lowest. The contents of soil AN and AP increased significantly, and AN and AP contents in the soil of most plants in the 3rd season were above 500 mg kg−1. There was no difference in soil enzyme activities among the treatments in the 1st season, while in the 3rd season, soil α-glucosidase (AG) of plants of other families was higher than that of Poaceae plants, and cellobiohydrolase (CB) was the opposite. The changes of soil enzymes in different families and even in the 2 plants of the same family were not consistent. A clear rule was that in 1st and 3rd seasons the acid phosphatas (PHOS) activity in the soil of Panicinae subfamily plants was the highest, and the activity of β-1,4-xylosidase (XYL) in Poaceae family was significantly higher (P<0.05) than other families in the third season. The soil of Pooideae was generally higher than that of Panicinae. Soil enzyme showed a dynamic increasing trend in general. The activities of CB, PHOS and leucine aminopeptidase (LAP) enzymes increased most in Pooideae, Cucurbaceae and Panicinae, respectively. Soil PHOS enzyme activities of Panicinae and Pooideae, which belonged to the same family of Poaceae, exhibited the opposite trend of decreasing and increasing. Conclusion The soil PHOS and XYL activities of Poaceae plants are higher, and other enzyme activities do not show obvious changes in the same family. [Ch, 5 fig. 2 tab. 35 ref.] -
Key words:
- continuous cropping obstacle /
- Poaceae /
- soil enzyme activity
-
图 1 第1季不同植物土壤根区酶活性
Figure 1 Soil rhizosphere enzyme activities of different plants after the first season
图 2 第3季不同植物土壤根区酶活性
Figure 2 Soil rhizosphere enzyme activities of different plants after the third season
图 3 不同植物第1季和第3季土壤酶活性平均增幅或减幅
Figure 3 Average increase or decrease percentage of soil enzyme activities in the first and third plant seasons
图 4 不同科及禾本科亚科第1季和第3季植物根区土壤碳循环酶活性的差异
Figure 4 Difference of carbon cycling enzyme activity between family or subfamily of rhizosphere soil in the first and third seasons
图 5 不同科植物第1季和第3季土壤酶活性平均增幅或减幅
Figure 5 Average increase or decrease percentage of soil enzyme activities in the first and third plant seasons based on plant family
表 1 植物栽培和收获时间表
Table 1. Schedule for transplanting and harvesting
栽植季度 栽培日期
(年-月-日)采样日期
(年-月-日)采收植物 第1季 2020-12-02 2021-02-02
2021-02-07
2021-02-20
2021-03-01
2021-03-08黄豆、花生
高粱、玉米
马铃薯、小麦
二穗短柄草、黑麦草
番茄、西瓜、南瓜第2季 2021-03-18 2021-04-29
2021-05-02
2021-05-07
2021-05-14
2021-05-17黄豆、玉米
花生、高粱
西瓜、南瓜、番茄
马铃薯、二穗短柄草
黑麦草、小麦第3季 2021-05-31 2021-07-14
2021-07-20
2021-07-21
2021-07-23玉米、番茄
黄豆、花生、马铃薯
西瓜、南瓜、高粱
黑麦、小麦、二穗短柄草 下载: 导出CSV
下载: 导出CSV
表 2 第1季和第3季植物收获后不同植物根区土壤化学性质比较
Table 2. Comparison of soil chemical properties in root zone of different plants after harvest in the first and third seasons
植物种类 pH 碱解氮/(mg·kg−1) 有效磷/(mg·kg−1) 第1季 第3季 第1季 第3季 第1季 第3季 花生 5.03±0.05 bc 4.20±0.08 abc 561.65±60.70 cde 705.83±58.59 a 149.89±17.64 e 651.52±21.10 a 黄豆 5.23±0.17 ab 4.10±0.08 abc 567.42±34.93 bcd 577.84±30.07 ab 161.44±30.06 de 355.58±63.73 ef 西瓜 4.23±0.25 e 3.97±0.05 bc 492.05±46.00 e 672.01±96.79 ab 161.36±30.12 de 393.66±34.33 def 南瓜 4.90±0.22 cd 4.13±0.21 abc 664.13±22.70 ab 553.72±108.19 ab 323.48±17.76 a 577.35±93.74 ab 番茄 4.93±0.09 bc 4.13±0.12 abc 647.74±56.66 abc 520.56±131.32 abc 292.47±18.44 ab 478.03±0.02 bcde 马铃薯 4.77±0.05 cd 3.97±0.05 bc 530.86±10.41 de 360.96±12.77 c 267.34±49.73 abc 304.40±51.93 f 二穗短柄草 4.57±0.17 d 3.93±0.12 c 540.13±39.37 de 356.81±20.47 c 245.17±16.65 bc 425.49±40.00 cdef 黑麦 4.73±0.12 cd 4.23±0.12 ab 593.97±29.02 bcd 667.47±82.33 ab 218.97±46.98 cd 546.70±60.91 abc 小麦 4.83±0.05 cd 4.23±0.12 ab 567.93±26.81 bcd 687.14±71.14 ab 114.48±11.85 e 524.25±41.53 abc 玉米 5.43±0.17 a 4.27±0.21 a 699.27±75.11 a 546.80±114.25 ab 288.63±45.34 ab 599.87±42.06 ab 高粱 4.77±0.09 cd 4.13±0.05 abc 397.89±24.82 f 509.08±23.38 bc 124.26±3.98 e 485.14±24.71 bcd 说明:不同小写字母表示不同植物间差异显著(P<0.05)。
下载: 导出CSV
-
[1] FAN Yanli, LIU Junjie, LIU Zhuxiu, et al. Chitin amendments eliminate the negative impacts of continuous cropping obstacles on soil properties and microbial assemblage [J/OL]. Frontiers in Plant Science, 2022, 13: 1067618[2022-06-10]. doi: 10.3389/fpls.2022.1067618. [2] WANG Feiyang, ZHANG Xuemin, WEI Meitang, et al. Appropriate crop rotation alleviates continuous cropping barriers by changing rhizosphere microorganisms in Panax notoginseng [J/OL]. Rhizosphere, 2022, 23: 100568[2022-06-10]. doi: 10.1016/j.rhisph.2022.100568. [3] QIN Shuhao, STEPHEN Yeboah, LI Cao, et al. Breaking continuous potato cropping with legumes improves soil microbial communities, enzyme activities and tuber yield [J/OL]. PLoS One, 2017, 12(5): e175934[2022-06-10]. doi: 10.1371/journal.pone.0175934. [4] 张冬明, 郑道君, 曾建华, 等. 西瓜连作对土壤主要微生物数量、酶活性及果实品质的影响[J]. 北方园艺, 2017(4): 155 − 158. ZHANG Dongming, ZHENG Daojun, ZENG Jianhua, et al. Effects of continuous cropping on microorganism and enzymes in the soil and the quality of watermelon fruit [J]. Northern Horticulture, 2017(4): 155 − 158. [5] TAN Ge, LIU Yongjun, PENG Shuguang, et al. Soil potentials to resist continuous cropping obstacle: three field cases [J/OL]. Environmental Research, 2021, 200: 111319[2022-06-10]. doi: 10.1016/j.envres.2021.111319. [6] 李天来, 杨丽娟. 作物连作障碍的克服——难解的问题[J]. 中国农业科学, 2016, 49(5): 916 − 918. LI Tianlai, YANG Lijuan. Overcoming continuous cropping obstacles: the difficult problem [J]. Scientia Agricultura Sinica, 2016, 49(5): 916 − 918. [7] 赵利坤, 张英. 作物连作障碍的影响因素及防治对策[J]. 黑龙江农业科学, 2013(12): 18 − 20. ZHAO Likun, ZHANG Ying. Influencing factors and countermeasures of continuous cropping obstacles [J]. Heilongjiang Agricultural Sciences, 2013(12): 18 − 20. [8] 杜家方. 地黄连作障碍发生的生物机制及其消减措施研究[D]. 武汉: 华中农业大学, 2020. DU Jiafang. Study on the Biological Mechanism of Continuous Cropping Obstacles in Rehmannia glutinosa and Its Reduction Measures [D]. Wuhan: Huazhong Agricultural University, 2020. [9] 夏梅梅, 钟宛凌, 欧阳里山, 等. 1989—2018年国内作物连作障碍研究现状——基于CNKI的文献计量学分析和科学知识图谱研究[J]. 农学学报, 2021, 11(3): 46 − 54. XIA Meimei, ZHONG Wanling, OUYANG Lishan, et al. Research status of continuous cropping obstacles in China from 1989 to 2018: based on bibliometric analysis and knowledge mapping of CNKI [J]. Journal of Agriculture, 2021, 11(3): 46 − 54. [10] ZHU Lizhen, HE Jun, TIAN Ying, et al. Intercropping wolfberry with Gramineae plants improves productivity and soil quality [J/OL]. Science Direct, 2022, 292(7): 110632[2022-06-10]. doi: 10.1016/J.SCIENTA.2021.110632. [11] 王静, 王磊, 李永青, 等. 3种桉树叶提取物除草活性研究[J]. 中国农学通报, 2013, 29(6): 191 − 194. WANG Jing, WANG Lei, LI Yongqing, et al. Study on herbicidal activity of three eucalyptus leaf extracts [J]. Chinese Agricultural Science Bulletin, 2013, 29(6): 191 − 194. [12] 赵军, 耿增超, 尚杰, 等. 生物炭及炭基硝酸铵对土壤微生物量碳、氮及酶活性的影响[J]. 生态学报, 2016, 36(8): 2355 − 2362. ZHAO Jun, GENG Zengchao, SHANG Jie, et al. Effects of biochar and biochar-based ammonium nitrate fertilizers on soil microbial biomass carbon and nitrogen and enzyme activities [J]. Acta Ecologica Sinica, 2016, 36(8): 2355 − 2362. [13] 丁维婷, 武雪萍, 张继宗, 等. 长期有机无机配施对暗棕壤土壤酶活性及春麦产量品质的影响[J]. 中国土壤与肥料, 2020(6): 1 − 8. DING Weiting, WU Xueping, ZHANG Jizong, et al. Effects of long-term organic-inorganic combined application on enzyme activity of dark brown soil and yield, quality of spring wheat [J]. Soil and Fertilizer Sciences in China, 2020(6): 1 − 8. [14] 鲁莽. 植物根系及其分泌物对微生物生长及活性的影响[J]. 化学与生物工程, 2012, 29(3): 18 − 21. LU Mang. Effects of plant roots and root exudates on growth and activity of microorganism [J]. Chemistry &Bioengineering, 2012, 29(3): 18 − 21. [15] 陈玲, 董坤, 杨智仙, 等. 连作障碍中化感自毒效应及间作缓解机理[J]. 中国农学通报, 2017, 33(8): 91 − 98. CHEN Ling, DONG Kun, YANG Zhixian, et al. Allelopathy autotoxcity effect of successive cropping obstacle its alleviate mechanism by intercropping [J]. Chinese Agricultural Science Bulletin, 2017, 33(8): 91 − 98. [16] SAIYA-CORK K R, SINSABAUGH R I, ZAK D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil [J]. Soil Biology &Biochemistry, 2002, 34(9): 1309 − 1315. [17] 徐仁扣, 李九玉, 周世伟, 等. 我国农田土壤酸化调控的科学问题与技术措施[J]. 中国科学院院刊, 2018, 33(2): 160 − 167. XU Renkou, LI Jiuyu, ZHOU Shiwei, et al. Scientific issues and controlling strategies of soil acidification of croplands in China [J]. Bulletin of Chinese Academy of Sciences, 2018, 33(2): 160 − 167. [18] 李忠, 江立庚, 唐荣华, 等. 连作对花生土壤酶活性、养分含量和植株产量的影响[J]. 土壤, 2018, 50(3): 491 − 497. LI Zhong, JIANG Ligeng, TANG Ronghua, et al. Effects of long-term continuous peanut cropping on dry matter weight of different peanut varieties, soil nutrient contents and enzyme activities [J]. Soils, 2018, 50(3): 491 − 497. [19] KEVIN Z, MGANG A, BAHAR S, et al. Microbial and enzymes response to nutrient additions in soils of Mt. Kilimanjaro region depending on land use [J]. European Journal of Soil Biology, 2015, 69: 33 − 40. [20] QIN Yaoguo, YANG Cuiqin, XIA Jialong, et al. Effects of dual/threefold rootstock grafting on the plant growth, yield and quality of watermelon [J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2014, 42(2): 495 − 500. [21] MIAO Li, QIN Xing, GAO Lihong, et al. Selection of reference genes for quantitative real-time PCR analysis in cucumber (Cucumis sativus L.), pumpkin ( Cucurbita moschata Duch.) and cucumber-pumpkin grafted plants [J/OL]. PeerJ, 2019, 7: e6536[2022-06-10]. doi: 10.7717/peerj.6536. [22] 张芊, 乜兰春, 赵文圣, 等. 甜瓜耐冷砧木筛选及嫁接苗耐冷性评价[J]. 中国蔬菜, 2022(7): 85 − 91. ZHANG Qian, NIE Lanchun, ZHAO Wensheng, et al. Screening of melon cold resistant rootstocks and evaluation on cold tolerance of grafted seedlings [J]. China Vegetables, 2022(7): 85 − 91. [23] 符厚隆, 廖道龙, 刘子凡, 等. 南瓜作砧木嫁接冬瓜根系分泌物对土壤微生态的影响[J]. 中国瓜菜, 2022, 35(6): 56 − 61. FU Houlong, LIAO Daolong, LIU Zifan, et al. Root exudates affect soil microecology of grafted wax gourd with pumpkin as rootstock [J]. China Cucurbits and Vegetables, 2022, 35(6): 56 − 61. [24] TAMILSELVI N A, PUGALENDHI L. Role of cucurbitaceous rootstocks on vegetative growth, fruit yield and quality of bitter gourd (Momordica charantia L. ) scions through grafting [J]. Journal of Animal and Plant Sciences, 2018, 28(3): 811 − 818. [25] 李勋, 崔宁洁. 大渡河干旱河谷地区2个饲用甜高粱品种农艺性状的研究[J]. 农业与技术, 2022, 42(13): 18 − 21. LI Xun, CUI Ningjie. Agronomic characters of two forage sweet sorghum varieties in arid valley area of Dadu River [J]. Agriculture and Technology, 2022, 42(13): 18 − 21. [26] 王伟伟, 齐冰洁, 赵攀衡, 等. 低磷胁迫下不同磷效率燕麦品种酸性磷酸酶活性差异[J]. 北方农业学报, 2017, 45(5): 55 − 58. WANG Weiwei, QI Bingjie, ZHAO Panheng, et al. The acid phosphatase activities of different phosphorus efficiency of oat varieties under low phosphorus stress [J]. Journal of Northern Agriculture, 2017, 45(5): 55 − 58. [27] 张丽梅, 郭再华, 张琳, 等. 缺磷对不同耐低磷玉米基因型酸性磷酸酶活性的影响[J]. 植物营养与肥料学报, 2015, 21(4): 898 − 910. ZHANG Limei, GUO Zaihua, ZHANG Lin, et al. Effects of phosphate deficiency on acid phosphatase activities of different maize genotypes tolerant to low-P stress [J]. Journal of Plant Nutrition and Fertilizers, 2015, 21(4): 898 − 910. [28] WU Aijiao, FANG Yan, LIU Shuo, et al. Root morphology and rhizosheath acid phosphatase activity in legume and graminoid species respond differently to low phosphorus supply [J/OL]. Rhizosphere, 2021, 19: 100391[2022-06-10]. doi: 10.1016/j.rhisph.2021.100391. [29] 洪彪, 李明, 黄结雯, 等. 连作对广藿香扦插苗土壤微生物及酶活性的影响[J]. 江西农业大学学报, 2020, 42(3): 438 − 447. HONG Biao, LI Ming, HUANG Wenjie, et al. Effects of continuous cropping on microorganism and enzyme activities in the restubble soil of patchouli [J]. Acta Agriculturae Universitatis Jiangxiensis, 2020, 42(3): 438 − 447. [30] 张丽琼, 郝明德, 臧逸飞, 等. 苜蓿和小麦长期连作对土壤酶活性及养分的影响[J]. 应用生态学报, 2014, 25(11): 3191 − 3196. ZHANG Liqiong, HAO Mingde, ZANG Yifei, et al. Effects of continuous cropping of wheat and alfalfa on soil enzyme activities and nutrients [J]. Chinese Journal of Applied Ecology, 2014, 25(11): 3191 − 3196. [31] WANG Yan, ZHANG Yu, LI Zhenzhou, et al. Effect of continuous cropping on the rhizosphere soil and growth of common buckwheat [J]. Plant Production Science, 2020, 23(1): 81 − 90. [32] KANG Yalong, JING Feng, SUN Wenqing, et al. Soil microbial communities changed with a continuously monocropped processing tomato system [J]. Acta Agriculturae Scandinavica,Section B:Soil &Plant Science, 2017, 68(2): 149 − 160. [33] 王雨晴, 陈香碧, 董明哲, 等. 红壤丘陵区旱地和水旱轮作地土壤中纤维素降解功能微生物群落特征[J]. 农业环境科学学报, 2017, 36(10): 2071 − 2079. WANG Yuqing, CHEN Xiangbi, DONG Mingzhe, et al. Characteristics of cellulose-degrading microbial communities in upland and paddy-upland rotation land soils in red soil hilly region [J]. Journal of Agro-Environment Science, 2017, 36(10): 2071 − 2079. [34] 赖先军. 玉米、高粱和谷子基因表达和调控元件以及人工选择的比较研究[D]. 成都: 四川农业大学, 2018. LAI Xianjun. Comparative Study of Gene Expression, Gene Regulatory Elements, and Artificial Selection in Poaceae Crops: Maize, Sorghum, and Setaria [D]. Chengdu: Sichuan Agricultural University, 2018. [35] 尹晓童, 杨浩, 于瑞鹏, 等. 根系分泌物在作物多样性体系中对种间地下部互作的介导作用[J]. 中国生态农业学报, 2022, 30(8): 1215 − 1227. YIN Xiaotong, YANG Hao, YU Ruipeng, et al. Interspecific below-ground interactions driven by root exudates in agroecosystems with diverse crops [J]. Chinese Journal of Eco-Agriculture, 2022, 30(8): 1215 − 1227. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220494
 点击查看大图
点击查看大图

计量
- 文章访问数: 3240
- HTML全文浏览量: 680
- PDF下载量: 212
- 被引次数: 0
