





-
近年来,由于全球气温升高,浙江省极端高温干旱天气频发,特别是当高温和少雨相伴发生时,高温胁迫发生的频率和范围便会增大[1]。光合作用作为植物对外界环境因子最为敏感的生理过程[2]也相应受到不同程度的影响。植物生长季的平均气温升高1 ℃,作物产量可减少高达17%[3]。当植物遭受中度高温胁迫(30~42 ℃)时,光合作用受抑是可逆的,只有当植物遭受严重高温胁迫(Ta>45 ℃)后,光合机构会受到永久性伤害,使光合作用的抑制不可逆转[4]。植物光合作用本身是一个复杂的生理过程,光合能力的强弱在一定程度上取决于物种的遗传特性[5]。对不同树种光合作用生理生态因子已有较多研究[6-13],对不同树种逆境中表现出的光合特性的研究有助于我们比较不同树种应对极端气候的差异,对亚热带高温区域绿化树种选育起到参考作用。
HTML
-
本研究试验区选在浙江省临安市,位于浙江省西北部,东临杭州市余杭区。地理坐标为29°56′~30°23′N,118°51′~119°52′E,属中亚热带季风气候区,温暖湿润光照充足,雨水充沛。本试验区全年降水量为1 628.6 mm,全年平均气温16.4 ℃,全年日照时数1 847.3 h左右。选取浙江省5个常见绿化造林树种(表 1),实验区设置在浙江农林大学校园内,试验测定树种的环境、立地和管理条件一致,所测树木均为移栽实生苗,属同期栽植。树龄均为10~15 a,无明显病虫害影响,生长状况良好。
序号 科 属 树种 1 银杏科Ginkgoaceae 银杏属 银杏Ginkgo biloba 2 棒科Lauraceae 樟属Cinnamomum 樟树Cinnamomum camphora 3 木兰科Magnoliaceae 含笑属4icheha 乐昌含笑Michelia chapensis 4 木兰科Magnoliaceae 木兰属Magnolia 广玉兰Magnolia grandiflora 5 木兰科Magnoliaceae 木兰属Magnolia 玉兰Magnolia denudata Table 1. Five common tree species in northern Zhejiang
-
利用英国ADC Bioscientific公司生产的LCPRO+便携式光合作用仪在2013年极端高温8月和2014年正常气温8月连续观测环境因子和5树种光合作用生理参数,光合特征参数为银杏、樟树、乐昌含笑、广玉兰、玉兰等的净光合速率(Pn),蒸腾速率(Tr),气孔导度(Gs),胞间二氧化碳摩尔分数(Ci)等,环境参数为光合有效辐射(RPAR),叶片温度(T1),空气湿度(HRH),大气二氧化碳摩尔分数(Ca),大气温度(Ta)等。选择3株·树种-1长势相近的样木,各取枝条3段·样木-1分别测定不同枝条上的3片不同叶片,测定叶片取朝向一致,无病虫害,从植株顶部展开叶往下数第5~7叶近叶尖位置,每片叶子在Pn变化幅度小于0.5时连续记录5个值。试验选择无风晴朗天气进行,从6:00-18:00隔2 h测定1次。试验于2014年8月利用相同测量方法,对同株试验样木进行重复测量。
利用LCPRO+便携式光合作用仪测定5树种的光响应曲线。测量设定温度30.0 ℃,二氧化碳摩尔分数为500.00 μmol·mol-1,空气相对湿度为50%~70%,叶室光照强度设置梯度为1 600.00,1 400.00,1 200.00,1 000.00,800.00,600.00,400.00,300.00,250.00,200.00,150.00,100.00,50.00和20.00 μmol·m-2·s-1,测量叶面积6 cm2,测量结果取3株样木的平均值,利用LCPRO+便携式光合作用测定仪测定。净光合速率温度响应曲线测定设定光强600.00 μmol·m-2·s-1,二氧化碳摩尔分数为500.00 μmol·mol-1,空气相对湿度为50%~70%,测量叶面积6 cm2,测量结果取3株样木的平均值,利用LCPRO+便携式光合作用测定仪测定。对低光量子通量密度(DPFD)(0.00~200.00 μmol·m-2·s-1)下Pn进行线性回归分析,求得光补偿点(PLCP)。
-
所测得的光响应数据运用Excel 2013和SPSS 21等软件进行处理与分析。
1.1. 研究区概况和试验材料
1.2. 试验方法
1.3. 数据分析
-
植物的生理指标如光合速率、蒸腾速率等的变化均是对外界环境因子变化而产生的自适应性调整,在极端气候条件下,如大气温度、湿度、二氧化碳摩尔分数及光合有效辐射等环境因子发生较大改变时植物会产生抗逆性反应,从而导致其光合和蒸腾作用表现出更为复杂的变化规律。
分别记录2014年8月和2013年8月环境因子日变化数据(图 1),RPAR和Ta均呈单峰曲线。2014年和2013年,RPAR均在12:00有最高值(1 728.16,1 812.46 μmol·m-2·s-1);Ta的变化主要受到RPAR影响(R2014=0.77,R2013=0.63),最高值分别出现在14:00和10:00,极端高温天气大气温度较一般夏季提前4 h达到当日最高值,日平均温度比一般夏季高7.9%。HRH在8月6:00第1次测量之后呈下降趋势,大气Ca的变化与RPAR变化呈负相关(R2014=-0.74,R2013=-0.71),在12:00-14:00达到当日最小值。

Figure 1. Diurnal courses of environmental factors in August 2014 and August 2013
-
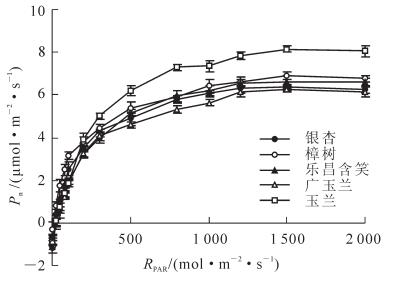
当气温、大气二氧化碳摩尔分数、大气相对湿度等环境因子相同,5树种叶片净光合速率对光合有效辐射的响应如图 2所示。随着RPAR的增加Pn在弱光强(RPAR<200.00 μmol·m-2·s-1范围内,光合速率随着光强的上升迅速上升。各树种光补偿点(PLCP)集中于9.63~18.95 μmol·m-2·s-1之间,樟树(9.63)耐阴性比其他树种高,木兰科植物耐阴性较为接近;乐昌含笑和银杏光饱和点(PLSP)集中于1 200.00 μmol·m-2·s-1左右,樟树、广玉兰、玉兰光饱和点集中于1 500.00 μmol·m-2·s-1左右,达到最适光强后玉兰Pn(8.14 μmol·m-2·s-1)高于其他树种。5树种叶片净光合速率对温度的响应呈指数变化如图 3所示。叶片Pn在25~35 ℃变化范围内随温度的增加而增加,当温度达到最适温度35 ℃时Pn有最大值,此时樟树(9.09 μmol·m-2·s-1)光合速率高于其他树种。

Figure 2. Response of photosynthetic rate to light intensity of five species

Figure 3. Response of photosynthetic rate to temperatures of five species
-
日Pn能够反映植物光合能力的大小[14]。根据图 4:树种Pn均于8:00出现第1个峰值,但2014年5个树种在12:00(樟树在10:00)出现明显光合午休现象(图 4A1),之后Pn开始回升,在14:00-16:00达到第2个峰值,不同树种Pn日变化趋势较相似,日均值差异不明显,Pn日均值樟树(5.02 μmol·m-2·s-1)>银杏(4.96 μmol·m-2·s-1)>玉兰(4.06 μmol·m-2·s-1)>乐昌含笑(3.76 μmol·m-2·s-1)>广玉兰(3.56 μmol·m-2·s-1)。2013年,在14:00气温下降之后Pn无明显回升(图 4A2),试验树种未出现明显光合午休现象。Pn日均值表现为广玉兰(1.08 μmol·m-2·s-1)>银杏(0.78 μmol·m-2·s-1)>樟树(0.55 μmol·m-2·s-1)>玉兰(0.63 μmol·m-2·s-1)>乐昌含笑(0.31 μmol·m-2·s-1)。持续的高温条件造成试验树种光合速率日变化平均值减少了70%~92%,高温灾害性天气已经对植物正常生理活动造成了显著影响(P<0.05)。

Figure 4. Diumal changes in net photosynthesis (Pn), transpirational rate (Tr), intercellular CO2 concentration (Ci), stomatal conductance (Gs), and water use efficiency (EWUE) in leaves of five species under high temperature conditions
同一树种Tr变化与Gs变化较接近,在不同年份夏季试验树种Tr和Gs多呈双峰变化(除2014年广玉兰和玉兰)。2014年,不同树种的Tr和Gs变化存在较大差异(图 4B1,4C1),第1个峰值在8:00-10:00,第2个峰值在14:00-16:00,在12:00左右出现当日最低值。随着植物光合午休现象结束Pn上升,Tr和Gs在14:00左右出现第2个峰值。2013年,不同树种的Tr和Gs变化呈相似性(图 4B2,4C2),在8:00出现第1个峰值(除广玉兰Tr),在14:00-16:00左右出现第2个峰值,10:00-14:00之间Tr和Gs均呈较低水平。在极端高温天气试验树种Tr和Gs均呈较低水平,持续高温影响下Tr和Gs的树种间差异被削弱。
正常气温下植物叶片Ci变化与Pn变化相反(图 4D1),当光合作用升高时叶肉细胞固定的二氧化碳增多Ci下降。2014年,Ci第1个波谷出现在8:00-10:00(图 4D2),第2个波谷出现在16:00,随着光合午休现象的出现,Ci在12:00-14:00出现波峰。2013年,Ci变化呈不规则波动趋势(乐昌含笑数据采集期可能仪器和测量叶室温度过高导致测量误差),且Ci水平明显高于正常气温,这与Pn值在高温条件下平均值低于正常年份有关。
水分利用率(EWUE)为光合作用与蒸腾作用的比值,一般情况下作为评价植物适应干旱胁迫能力的一个重要生理指标[15]。2014年,试验树种EWUE在8:00和12:00左右出现2个峰值(图 4E1),EWUE日平均值为樟树(2.57 mmol·mol-1)>银杏(2.56 mmol·mol-1)>广玉兰(2.41 mmol·mol-1)>玉兰(2.17 mmol·mol-1)>乐昌含笑(2.15 mmol·mol-1)。2013年,试验树种在8:00和16:00出现2个峰值(图 4E2),EWUE日平均值为银杏(1.17 mmol·mol-1)>玉兰(1.16 mmol·mol-1)>广玉兰(1.30 mmol·mol-1)>樟树(1.06 mmol·mol-1)>乐昌含笑(0.49 mmol·mol-1)。正常年份和极端高温下试验树种表现出不同的耐旱性。
-
植物的光合作用是一个复杂的生理过程,叶片光合速率与光照强度、气温、二氧化碳摩尔分数、水分等因子有关[16]。植物在高温条件下的光合速率明显不同于其他生长季,随着光照强度的增高大气温度和湿度产生明显变化,植物自身的生理因子也会产生相应的变化。根据表 2:在极端高温天气时RPAR,Ta,T1与Pn呈负相关。Pn与Ci呈负相关关系,樟树、玉兰的Pn与Ci呈极显著相关关系(R1=-0.91**,R2=-0.90**);玉兰的Pn与Tr呈极显著相关关系(R=1.00**);Pn与Gs呈正相关关系,广玉兰Pn与Gs呈极显著相关关系(R=0.90**),银杏、樟树、玉兰Pn与Gs呈显著相关关系(R1=0.76*,R2=0.82*,R3=0.81*)从相关性系数的大小及出现的概率来区分,对Pn影响最大的是Gs,其次是Ci,其他因素影响并不明显。根据表 3:正常年份银杏、樟树Pn与Tr呈显著相关关系(R1=0.83*,R2=0.86*),玉兰Pn与Tr呈极显著相关关系(R=0.88**);乐昌含笑、广玉兰Pn与Gs呈显著相关关系(R1=0.76*,R2=0.87*),玉兰Pn与Gs呈极显著相关关系(R=0.97**);从相关性系数的大小及出现的概率来区分,对Pn影响最大的是Gs,其次是Tr,其他因素影响并不明显。
因子 树种 RPAR Ta T1 Ci Tr Gs 银杏 -0.118 -0.384 -0.442 -0.484 0.512 0.755* 樟树 0.140 -0.578 -0.527 -0.910** -0.162 0.817* Pn 乐昌含笑 -0.180 -0.571 -0.544 -0.666 -0.731 0.071 广玉兰 -0.137 -0.372 -0.360 -0.020 0.569 0.901** 玉兰 -0.221 -0.139 -0.162 -0.904** 0.995** 0.812* 银杏 0.203 -0.390 -0.269 -0.362 1 0.933** 樟树 -0.507 0.343 0.245 0.210 1 0.012 Tr 乐昌含笑 -0.284 0.053 -0.005 0.267 1 0.323 广玉兰 0.477 0.274 0.299 -0.145 1 0.691 玉兰 -0.189 -0.084 -0.105 -0.881** 1 0.804* 说明:*表示相关性达显著水平(P<0.05);**表示相关性达极显著水平(P<0.01)。 Table 2. Correlative coefficient between photosynthetic rate in leaves of 5 species phy-ecological factors and transpiration rate in 2013
因子 树种 RPAR Ta T1 Ci Tr Gs 银杏 0.290 0.414 0.431 -0.120 0.827* 0.318 樟树 0.103 -0.279 -0.225 0.516 0.855* 0.544 Pn 乐昌含笑 -0.180 -0.571 -0.544 -0.666 -0.731 0.071 广玉兰 0.223 -0.463 -0.431 0.098 0.456 0.870* 玉兰 0.575 0.549 0.558 0.423 0.883** 0.973** 银杏 0.711 0.323 0.447 0.117 1 0.427 樟树 0.317 0.084 0.009 0.408 1 0.613 Tr 乐昌含笑 0.884** 0.476 0.599 0.174 1 0.904** 广玉兰 0.625 0.221 0.266 -0.528 1 0.642 玉兰 0.758* 0.817* 0.826* 0.288 1 0.864* 说明:*表示相关性达显著水平(P<0.05);**表示相关性达极显著水平(P<0.01)。 Table 3. Correlative coefficient between photosynthetic rate in leaves of 5 species phy-ecological factors and transpiration rate in 2014
-
通过相关性分析发现:2013年2014年,环境因子与树种光合生理参数相关性较高,同一树种各生理参数相关性明显。对树种间各光合生理参数进行单因素方差分析发现(表 4),不同年份夏季不同树种间Pn,Tr,Ci差异不显著,高温条件下不同树种Ci差异显著(P<0.05),结合其他学者的研究这部分差异被认为主要来自于不同树种叶片结构特征差异[17]。2013年2014年,环境因子对树种的光合生理参数单因素方差分析如表 5。环境因子对不同树种光合作用参数Pn,Tr,Gs造成的影响显著(P<0.05),对银杏和广玉兰Ci变化造成的影响不明显(P>0.05)。
树种 自由度 Pn Tr Gs Ci n-1 f p f p f p f p 2014年5个树种 4 1.020 0 0.413 0 0.526 0 0.717 0 0.367 0 0.830 0 1.490 0 0.230 0 2013年5个树种 4 1.645 0 0.189 0 1.388 0 0.262 0 2.248 0 0.087 0 7.958 0 < 0.000 1 Table 4. Different species of photosynthetic physiological parameters ANOVA
树种 Pn Tr Gs Ci f p f p f p f p 银杏 53.137 0 < 0.000 1 7.768 0 0.016 0 6.684 0 0.024 0 2.465 0 0.142 0 樟树 23.452 0 < 0.000 1 28.501 0 < 0.000 1 19.016 0 0.001 0 15.321 0 0.002 0 乐昌含笑 33.750 0 < 0.000 1 20.537 0 0.001 0 16.552 0 0.002 0 15.702 0 0.002 0 广玉兰 11.101 0 0.006 0 4.848 0 0.048 0 8.824 0 0.0120 2.865 0 0.116 0 玉兰 24.742 0 < 0.000 1 18.180 0 0.001 0 17.338 0 0.001 0 8.764 0 0.012 0 Table 5. 2013-2014 environmental factors on ANOVA analysis of species photosynthetic parameters
2.1. 环境因子日变化规律
2.2. 不同树种光响应曲线和温度响应曲线

2.3. 极端高温与正常年份树种光合作用生理参数比较

2.4. 极端高温与正常年份叶片生理因子差异分析
2.5. 2013年2014年环境因子与树种生理参数相关性分析
-
光合作用是植物对高温最敏感的部分之一,5个树种叶片在极端高温下净光合速率变化与正常年份有相似性,均在8:00达到第1个峰值,但极端高温下植物叶片净光合速率强度明显低于正常年份,未出现明显光合“午休”现象,在14:00之后未有明显回升。对于光合“午休”现象产生的原因,有学者[18-19]认为:影响因素主要有生理和生化因素2个方面。生理因素主要是由于环境因素变化,如高温、强光、二氧化碳浓度降低所导致的叶片气孔关闭,导致光合速率下降;生化因素主要是由于叶肉细胞自身羧化酶活性降低所导致的光合速率降低。极端高温下大气温度在8:00升高到42.4 ℃,5个树种光合速率均迅速下降,光合有效辐射和气温的迅速变化导致植物在极端高温条件下蒸腾作用变化幅度高于正常年份,试验期间极端高温下5个树种叶片胞间二氧化碳摩尔分数均高于正常气温,快速的失水造成叶片气孔迅速闭合,气孔快速闭合说明光合作用光抑制作用未发生,但增高的胞间二氧化碳摩尔分数和较低的光合速率说明此时叶片光合作用被抑制,之后表现出进入光合“午休”但无午休恢复现象,一系列的迹象表明高温条件下5树种光合速率降低主要因素可能是叶片光反应酶活性降低所导致的[20]。
有学者对不同土壤水分条件的银杏叶片研究发现,土壤水分越低,Ci变化越大,植物“午休”现象越明显[21-22]。研究发现同种植株在不同生境条件下叶片Pn变化主导因素不同[23],试验测得在高温条件下对Pn影响最大的是Gs,其次是Ci,高温条件使叶肉细胞自身羧化酶活性降低,部分酶促反应速率降低使Pn有明显下降[24]。正常夏季5个树种Pn在12:00左右出现低谷,在14:00-16:00有明显回升,Tr在10:00和14:00有2个峰值;极端高温下5个树种Pn出现波谷的时间提前到10:00,在10:00-18:00处于较低水平,5个树种在高温条件下产生类似周期延长的光合“午休”的抗逆性反应。
Pn与Tr的变化反映了EWUE的变化,EWUE可以反映植物对逆境的适应能力强弱[25]。通过研究发现:极端高温下5个树种EWUE日平均值为银杏>玉兰>广玉兰>樟树>乐昌含笑,正常气温下5个树种EWUE日平均值为樟树>银杏>广玉兰>玉兰>乐昌含笑,EWUE变化幅度最小的是樟树(20.32%),变化幅度最大的是玉兰(50.80%),在对5个树种Pn值温度响应曲线中发现,达到35 ℃最适温度时樟树Pn值明显高于其他树种,结合Pn值光响应曲线樟树相比其他试验树种更能够适应极端高温天气。
光合作用和蒸腾作用均在保卫细胞调节气孔开度这一共同路径下完成,但彼此却是相对独立的过程,当温度达到植物光合作用最适值之后,光合作用随着温度继续升高而下降。极端高温下,RPAR在6:00(596.34 μmol·m-2·s-1)开始上升,5个树种叶片Ci与Pn均有所上升,当RPAR在8:00达到1 008.70 μmol·m-2·s-1之后,Pn开始下降,Ci与Pn变化相反。试验发现当光强大致为1 000.00 μmol·m-2·s-1时,5个树种达到光饱和状态。
通过对试验树种种间、种内,及不同年份光合生理参数分析发现,不同树种光合参数Ci存在显著性差异,环境因子对不同树种光合作用参数Pn(P<0.01),Tr(P<0.05),Gs(P<0.02)造成的影响显著。不同树种间光合生理差异受到树种叶片胞间二氧化碳摩尔分数的影响显著,不同树种光合速率、蒸腾速率、气孔导度受到环境因子的显著影响。
 DownLoad:
DownLoad: