







-
大气氮沉降一定范围内可促进植物生长,过量则削弱植物的抗逆性,抑制植物正常生长,甚至造成生态系统的退化[1]。李德军等[2]发现:100 kg·hm-2·a-1氮强度下木荷Schima superba和黄果厚壳桂Cryptocarya concinna的净光合速率(Pn)和气孔导度(Gs)达到最大,超过该强度则表现为抑制。ZHANG等[3]发现:氮沉降抑制了毛竹Phyllostachys edulis的最大光化学效率(Fv/Fm)和实际量子产量[Y(Ⅱ)],促进了非光化学猝灭系数(qN)。当氮沉降持续增加时,毛竹的叶绿素含量、气孔导度和胞间二氧化碳浓度也受到抑制[4]。有研究发现:600 kg·hm-2的生物质炭可改善植物光合性能[5]。由此推测施加生物质炭能缓解氮沉降对毛竹光合特性的消极影响。生物质炭(biochar)是由有机物原料在彻底或部分缺氧的环境下经过热解炭化生成的一种具有高度芳香化结构的固态物质,理化性质独特,被普遍地用作改良剂来提高作物产量和品质[6];但自身营养价值不高,其增产效应还受到作物类型、土壤种类及施用量的影响[7]。研制生物质炭与肥料复合成的新型生物质炭肥已成为近年来的研究热点,这种新型生物质炭肥具有肥料缓释功效,在调节地温,改善土壤结构,保水保肥上具有显著促进效果[7-8]。当前关于生物质炭肥环境功能与理化特性及其对农作物产量影响等方面的报道较多[9],有关生物质炭在林业经营管理中的应用研究较少[10],生物质炭添加对毛竹光合生理的影响还未见报道。毛竹是亚热带广泛分布的主要竹种,经济价值高,种植面积连年扩大[11]。中国现有毛竹林面积4.43 Mhm2,占全球毛竹林分布面积的84.02%[12-13];毛竹生长快,再生能力强,在固碳和缓解气候变化上潜力巨大[12, 14]。在中国,毛竹林分布区也是氮沉降最严重的地区[11]。金晓春等[15]发现:换叶期间施肥对毛竹光合影响具有显著促进作用;但是在氮沉降条件下,施加生物质炭是否影响毛竹新老叶片的光合及叶绿素荧光特性还未见报道。本研究以2龄(2016年出笋)毛竹为试验对象,采取人工喷氮的方法模拟大气氮沉降,随后施加生物质炭,分析毛竹新老叶的光合和叶绿素荧光参数的变化规律,旨在研究施加生物质炭肥的毛竹对氮沉降的响应,为气候变化背景下提高毛竹林的生产力提供科技支撑。
HTML
-
本试验在浙江省杭州市临安区青山镇毛竹林试验基地(30°14′ N,119°42′ E)进行。该区为亚热带季风气候,四季分明,年平均气温为15.6 ℃,年平均降水量为1 420.0 mm,夏季多高温降雨天气,土壤类型为酸性黄壤。试验区内的毛竹林有约14 a的集约经营历史,为笋材两用林,经营措施主要为去除毛竹林下的杂草和灌木。每年9月施用450 kg·hm-2复合肥(氮67.5 kg·hm-2·a-1,磷11.8 kg·hm-2·a-1,钾74.7 kg·hm-2·a-1)[15]。毛竹林样地初始立竹密度为(3 362±309)株·hm-2,立竹胸径为(10.16±0.13)cm,初始土壤容重为(0.97±0.07)g·cm-3,土壤有机质为(23.73±0.24)mg·g-1,土壤全氮为(1.11±0.04)mg·g-1,土壤全磷为(0.52±0.01)mg·g-1,土壤pH值为4.46±0.01。
-
在研究区内建立12个林分条件和环境状况相似的代表性样方(20 m × 20 m),样方之间设置20 m以上的缓冲间隔以免相互干扰。以当前中国亚热带地区实际大气氮沉降量和未来增长趋势为依据[16],以当地氮沉降率[17]为基础(30.5 kg·hm-2·a-1)设置氮沉降水平,即低氮(30.0 kg·hm-2·a-1,N30),中氮(60.0 kg·hm-2·a-1,N60)和高氮(90.0 kg·hm-2·a-1,N90)。自2013年1月至2017年3月,每月月初配置氮沉降强度所需的硝酸铵(NH4NO3)溶液,对样地进行模拟喷氮;同时对照组(0 kg·hm-2·a-1,N0)喷洒等量自来水[18],各组重复3次。氮沉降21个月(2014年9月)后,在各氮沉降样地内建立3个大小为10 m × 10 m的生物质炭处理次样地,一次性分别施用0(BC0),20(BC20)和40 t·hm-2(BC40)生物质炭[19];均匀撒在地表后,翻耕30 cm与土壤混合均匀。
-
每个样地中心搭建高约10 m的观测架,用于竹叶光合作用的原位观测。2017年3月26-28日9:00-11:00在各样地中选取1株生长良好、大小(胸径10~11 cm,株高约13 m)相近的2龄毛竹,选取树体中部冠层第12~14轮枝上,向南伸展的枝条上部、发育良好的成熟新叶老叶各3片。采用LI-6400便携式光合测定仪测定各株新叶与同株原生老叶的气体交换参数,仪器参数为0.5 L·min-1空气流速,25 ℃环境温度,70%相对湿度,380 μmol·mol-1二氧化碳摩尔分数和1 200 μmol·m-2·s-1光合有效辐射(PAR);测量最大净光合参数(Pmax),气孔导度(Gs),胞间二氧化碳浓度(Ci)和蒸腾速率(Tr)等参数。采用SPAD-502叶绿素含量测定仪测定叶色值(SPAD),重复5次取平均值。用PAM-2500便携式叶绿素荧光分析仪测定叶绿素荧光参数,每次测定前,用仪器配备的叶夹使叶片暗适应20 min,然后照射饱和脉冲光,测量最大荧光(Fm),初始荧光(Fo),光能潜在活性(Fv/Fo),最大光化学效率(Fv/Fm),实际量子产量[Y(Ⅱ)]和非光化学猝灭系数(qN)等参数。
-
利用SPSS 17.0软件进行统计分析;利用单因素方差分析(one-way ANOVA)和最小显著差法(LSD)检验各处理下参数的差异显著性;根据方差齐性检验要求,对氮沉降和生物炭的影响进行可重复双因素方差分析。图均由SigmaPlot 12.5软件制作。
1.1. 试验区概况
1.2. 试验设计
1.3. 数据采集
1.4. 数据处理与分析
-
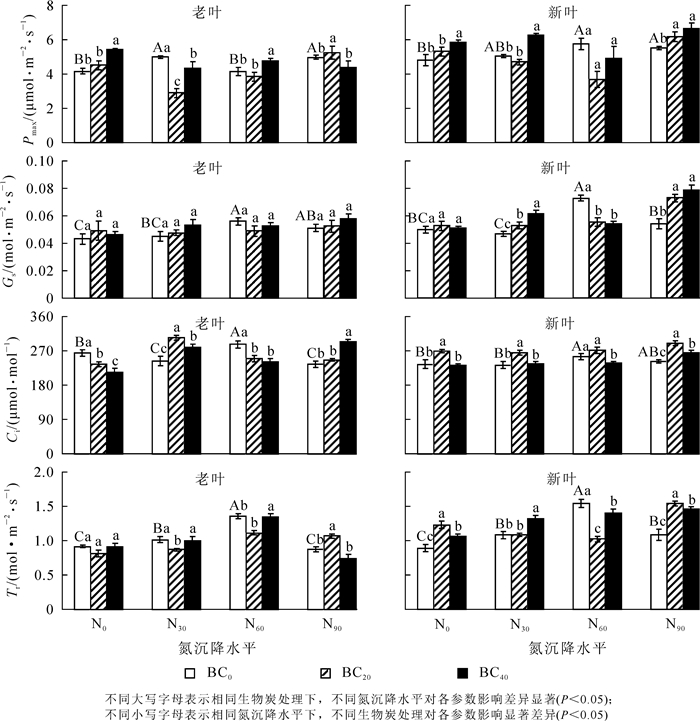
相比对照组,氮沉降处理对毛竹老叶和新叶的Pmax(图 1A,图 1B),Gs(图 1C,图 1D)和Tr(图 1G,图 1H)均具有显著的促进作用(P<0.05)。除N60处理外,氮沉降对老叶Ci(图 1E)均有显著抑制作用(P<0.05)。对照组施用生物质炭后,新叶、老叶的Pmax和新叶的Tr均显著增加;老叶Ci显著降低,新叶Ci呈现先升高后下降趋势(P<0.001)。与不施生物质炭(BC0)相比,N30处理下施加生物质炭,老叶Ci和新叶Pmax,Gs,Ci,Tr均显著增加,但老叶Pmax,Tr显著降低(P<0.05)。N60下施加生物质炭,毛竹新老叶的Ci,Tr和新叶的Gs显著降低,老叶Pmax显著增加(P<0.05),新叶Pmax则无显著差异(P>0.05)。N90下施加生物质炭,毛竹新老叶Pmax,Ci,Tr和新叶的Gs显著增加(P<0.01);且新叶的Pmax的促进效果要高于老叶。

Figure 1. Effects of nitrogen deposition and biochar amendment on the gas exchange parameters of the new and old leaves of bamboo
-
相比对照组,老叶和新叶的SPAD值随氮沉降强度的增加而上升,但无显著差异(图 2A,图 2B)(P>0.05);对照组施用生物质炭,毛竹新叶和老叶SPAD均无显著性差异(P>0.05)。与不施生物质炭(BC0)相比,N30和N60下施加生物质炭的老叶和N60下施加生物质炭的新叶SPAD差异极显著(P<0.001),随生物质炭施加量的增加新老叶SPAD值均呈现出先降低后上升趋势,且对新叶SPAD值的促进效果要高于老叶。

Figure 2. Effects of nitrogen deposition and biochar amendment on the SPAD of the new and old leaves of bamboo
-
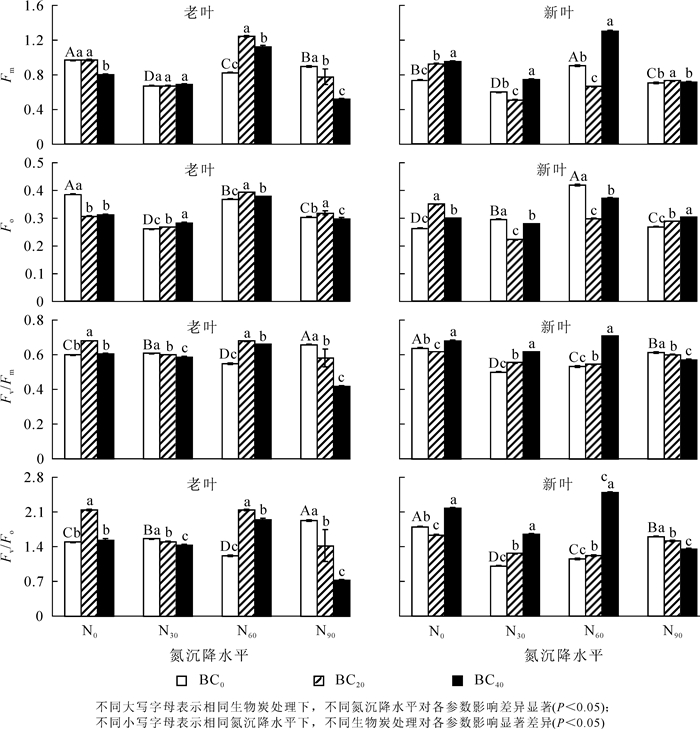
相比对照组,氮沉降处理对毛竹老叶的Fm(图 3A),Fo(图 3C)及新叶的Fv/Fm(图 3F),Fv/Fo(图 3H)具有显著的抑制作用(P<0.05)。除N60处理外,氮沉降处理对老叶的Fv/Fm(图 3E),Fv/Fo(图 3G)和新叶Fo(图 3D)具有显著的促进作用(P<0.05)。对照组施用生物质炭,老叶的Fm和Fo均显著降低,新叶的Fm(图 3B)和Fo显著增加,老叶Fv/Fo和Fv/Fm显著增加(P<0.05)。与不施生物质炭(BC0)相比,N30下施加生物质炭,老叶Fo和新叶Fv/Fm,Fv/Fo显著增加,新叶Fo和老叶Fv/Fm,Fv/Fo显著降低,新叶Fm随着生物质炭施加量的增加呈先下降后上升趋势(P<0.05)。N60下施加生物质炭,老叶Fm,Fo,Fv/Fm,Fv/Fo及新叶Fv/Fm,Fv/Fo显著增加,而新叶Fo显著降低,新叶的Fm随着生物质炭施加量的增加呈先下降后上升趋势(P<0.05)。N90下施加生物质炭,老叶Fm,Fv/Fm,Fv/Fo及新叶Fv/Fm和Fv/Fo显著降低,而新叶Fm和Fo显著增加,同时老叶Fo随着生物质炭施加量的增加呈先上升后下降趋势(P<0.05)。

Figure 3. Effects of Nitrogen deposition and biochar amendment on Fm, Fo, Fv/Fo and Fv/Fm of new and old leaves of bamboo
-
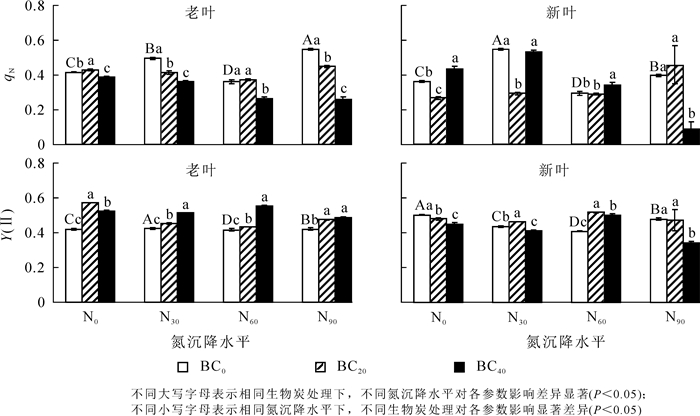
除N60处理外,氮沉降对毛竹新老叶qN(图 4A,图 4B)及老叶的Y(Ⅱ)(图 4C)具有显著促进作用(P<0.05)。相比对照组,氮沉降对新叶Y(Ⅱ)(图 4D)具有显著抑制作用(P<0.05)。对照组施用生物质炭,老叶的Y(Ⅱ)显著增加而新叶的Y(Ⅱ)显著降低,老叶qN随着生物炭施加量的增加呈先上升后下降趋势,而新叶qN呈相反趋势(P<0.05)。与不施生物质炭(BC0)相比,N30下施加生物质炭,新叶和老叶的qN显著降低,老叶的Y(Ⅱ)显著增加,新叶的Y(Ⅱ)随着生物炭施加量的增加呈先上升后下降趋势(P<0.05)。N60下施加生物质炭,老叶的Y(Ⅱ),新叶的Y(Ⅱ)和qN均显著增加,老叶的qN显著降低(P<0.05);N90下施加生物质炭,老叶的Y(Ⅱ)显著增加,老叶qN和新叶的Y(Ⅱ),qN则显著降低(P<0.05)。

Figure 4. Effects of nitrogen deposition and biochar amendment on qN and Y(Ⅱ) of new and old leaves of bamboo
-
双因素方差分析(表 1)表明:除新叶Tr和叶色值外,氮沉降显著影响了毛竹新老叶的光合和叶绿素荧光参数(P<0.001);除老叶Gs和新老叶Tr外,生物质炭显著影响了毛竹新老叶的光合和叶绿素荧光参数(P<0.01);除老叶Gs外,氮沉降和生物质炭的交互作用对毛竹新老叶的光合和叶绿素荧光参数影响显著(P<0.01)。
差异源 叶龄 氮沉降 生物质炭 交互 F P F P F P 最大净光合速率(Pmax) 老叶 6.182 *** 5.901 ** 8.150 *** 新叶 8.787 *** 8.857 *** 4.596 *** 气孔导度(Gs) 老叶 8.872 *** 2.210 0.113 1.856 0.091 新叶 15.569 *** 3.983 ** 10.708 *** 胞间二氧化碳浓度(Ci) 老叶 13.651 *** 4.106 ** 17.510 *** 新叶 8.291 *** 10.376 *** 3.162 ** 蒸腾速率(Tr) 老叶 18.114 *** 2.959 0.055 3.129 ** 新叶 2.601 0.054 1.308 0.273 10.030 *** 叶色值(SPAD) 老叶 7.691 *** 9.055 *** 5.456 *** 新叶 1.985 0.126 7.487 *** 3.292 ** 最大荧光(Fm) 老叶 4 674.660 *** 3 965.420 *** 1 644.273 *** 新叶 378.992 *** 71.460 *** 102.018 *** 初始荧光(Fo) 老叶 436.019 *** 72.843 *** 197.174 *** 新叶 3 601.767 *** 79.188 *** 396.305 *** 最大光化学效率(Fv/Fm) 老叶 2 737.899 *** 4 618.213 *** 1 896.683 *** 新叶 50.106 *** 59.594 *** 75.543 *** PO!潜在活性(Fv/Fo) 老叶 3 001.495 *** 6 337.901 *** 2 436.052 *** 新叶 37.190 *** 52.256 *** 64.359 *** 非光化学猝灭系数(qN) 老叶 255.861 *** 1 009.686 *** 181.946 *** 新叶 38.997 *** 15.537 *** 57.401 *** 实际量子产量[Y(Ⅱ)] 老叶 199.855 *** 1 808.386 *** 311.431 *** 新叶 16.009 *** 29.128 *** 21.589 *** 说明:**表示P<0.01,***表示P<0.001 Table 1. Two-way ANOVA analysis of N deposition and biochar amendment on photosynthesis and chlorophyll fluorescence characteristics of bamboo leaves
2.1. 氮沉降和生物质炭对光合气体交换参数的影响
2.2. 不同处理组毛竹的叶绿素荧光特性差异
2.2.1. 氮沉降和生物质炭对毛竹新老叶叶色值(SPAD)的影响

2.2.2. 氮沉降和生物质炭对毛竹新老叶Fm,Fo,Fv/Fo和Fv/Fm的影响
2.2.3. 氮沉降和生物质炭对毛竹新老叶qN和Y(Ⅱ)的影响
2.3. 氮沉降和生物质炭的交互作用对毛竹光合与叶绿素荧光参数的影响
-
植物有机物质合成、能量储存与转化以光合作用为基础,以Pn对光合生产力的影响最为重要[20-21]。本次研究于春季进行,结果发现:氮沉降处理后2龄毛竹换叶期叶片的Pmax,Gs和Tr均升高,说明氮沉降增强了毛竹叶片的光合作用,原因可能是氮沉降促进了叶片氮含量的增加,导致叶片中的光合色素增加,从而提高了光合能力[12]。与前期在夏季进行的研究[3]结果一致,说明不同季节、不同叶片发育阶段毛竹的气体交换能力对氮沉降的响应无差异。新叶Pmax高于老叶,可能原因是Pmax随着叶片的逐渐衰老而降低[22]。叶片光合作用受气孔和非气孔因素影响,Pn下降,Ci上升,气孔限制值(Ls)减小,由此认为叶肉细胞的光合活性减小是光合速率降低的决定因素[23];N60处理毛竹老叶片Pmax降低,Ci升高,Fv/Fm,Fv/Fo和Y(Ⅱ)也相应降低,很有可能是叶片细胞的光合活性降低造成的。除N60外,氮沉降提高了老叶的Fv/Fm,Fv/Fo和Y(Ⅱ),说明氮沉降提高了毛竹老叶的光强适应性和光能利用率,这与曲道春等[24]发现氮沉降提高樟树Cinnamomum camphora叶片Y(Ⅱ)的结果类似。有研究表明[25]:由于气温较低,快速生长前期(3月底)1龄和2龄毛竹叶片易受到环境胁迫,Fv/Fm均低于0.8;快速生长中期(4月)2,4和6龄毛竹叶片Fv/Fm均低于0.8,且上层叶片(8.5 m)低于中层(7.1 m)和下层(5.6 m)[26]。本研究同样发现叶片Fv/Fm低于0.8,可能是2017年3月研究区持续阴雨低温天气使得叶片受到光胁迫所致。叶绿素含量与SPAD值存在显著正相关[24],本试验发现不同氮沉降处理下2龄竹新老叶SPAD值均有上升,但差异不显著。
-
与不施生物质炭相比,施加生物质炭后毛竹新老叶的Pmax和新叶的Tr均有提高,而Ci都降低,说明生物质炭一定程度上促进了毛竹的光能利用率。有研究发现:适量的生物质炭肥提高了黄连木Pistacia chinensis的净光合速率、气孔导度及蒸腾速率,抑制了胞间二氧化碳浓度[27];但是生物质炭并未显著影响毛竹新老叶叶绿素的合成[7]。本试验也观测到生物质炭提高了毛竹老叶的Fv/Fm,Fv/Fo,Y(Ⅱ)和新叶Fm,推测原因为适量的生物质炭改善了土壤结构和微生物环境[6],增加了毛竹新叶PSⅡ反应中心潜在活性,提高了老叶PSⅡ的电子传递活性和实际光能转化效率。
-
研究发现:不同氮强度下施加生物质炭,毛竹新老叶的Pmax,Ci和Gs均不同程度地增加,老叶Tr总体下降,而新叶总体上升,说明毛竹叶片光合能力被提高,与李艳梅等[28]发现生物质炭与氮肥配施提高了番茄Lycopersicon esculentum净光合速率和蒸腾速率。刘领等[29]发现生物质炭与氮肥配施提高了烤烟Nicotiana tabacum的光能利用率的结果一致。N30处理下施加生物质炭后老叶Pmax,N60处理下施加生物质炭后新叶Pmax呈现出先降低后增加的趋势,且新叶Pmax高于老叶,与相对应处理的新老叶SPAD值变化趋势一致;说明氮沉降和生物质炭的复合作用下,新老叶片内氮、磷、钾等元素含量发生变化,造成SPAD值波动,进而影响毛竹叶片的净光合速率。
N30处理下施加生物质炭提高了毛竹新叶的Fv/Fm,Fv/Fo和老叶Y(Ⅱ),降低了新老叶的qN;N60处理下施加生物质炭提高了毛竹老叶Fv/Fm,Fv/Fo和Y(Ⅱ),毛竹新叶的Fv/Fm和Y(Ⅱ),降低了老叶的qN,原因可能是生物质炭改良了酸性土壤[8],促进了土壤微生物的生长;作为营养元素的缓释载体,生物质炭增加了土壤肥力,促进植物吸收营养元素(氮、磷、钾等),从而增强了毛竹光合荧光特性[6, 27]。说明氮沉降下适量施加生物质炭有利于增加毛竹PSⅡ反应中心潜在活性,提高最大光化学效率和实际光能转化效率,降低毛竹新叶老叶的光能热耗散率,使叶片所捕获的光能充分地用于光合作用[27, 29]。N90强度下施加生物质炭,毛竹老叶的Fv/Fm,Fv/Fo及毛竹新叶的Fv/Fm,Fv/Fo和Y(Ⅱ)均降低,则说明过量的氮沉降抑制了毛竹新老叶的光合性能[4, 25],降低了新老叶PSⅡ反应中心内禀光能转化效率,不利于毛竹叶片有机物的积累。总体来说,氮沉降持续增加的趋势下施加生物质炭有利于提高毛竹新老叶光合荧光特性。
3.1. 氮沉降对毛竹新老叶叶片光合作用的影响
3.2. 生物质炭对毛竹新老叶光合作用的影响
3.3. 氮沉降背景下施加生物质炭对毛竹新老叶光合作用的影响
-
一定程度的氮沉降提高了毛竹的荧光动力学参数,提升了毛竹PSⅡ反应中心开放程度,增强了毛竹最大光化学效率,有利于光合速率的提高,但对叶片叶色值影响不显著。生物质炭促进了毛竹的光合作用,提高了新老叶的荧光动力学参数,但并未显著影响叶绿素的合成;相比老叶,新叶反应更加敏感。不同氮沉降强度下施加生物质炭一定程度上促进了毛竹新老叶片的气体交换参数及荧光参数,氮沉降和生物质炭的交互作用对毛竹新老叶的光合和叶绿素荧光特性具有极显著的影响,表明在氮沉降日益增强背景下毛竹林里施加适量生物质炭有利于毛竹的生长。
 DownLoad:
DownLoad: