







-
合理平衡施肥对于促进苗木生长具有重要意义,能够提高苗木对外界不良环境包括干旱、低温以及病害等的抵抗力[1-4]。植物的稳态矿质营养理论是通过营养液栽培系统试验提出的,使植物生长速率与营养状态保持平衡[5]。TIMMER等[6]在稳态矿质营养理论的应用研究基础上提出指数施肥。指数施肥是通过指数递增的养分添加方式适应植物在各生长阶段相对生长率的施肥方法,能够区分营养不足、营养充足以及奢养消耗,并且能够通过诱导奢养消耗提高苗木的营养储备[7]。指数施肥的应用能够提高苗木质量,且成功应用于多种树种,例如黑云杉Picea mariana[8-9]、杉木Cunninghamia lanceolata[10]、西南桦Betula alnoides[11]、卢茨云杉Picea × lutzii[12]。闽楠Phoebe bournei为亚热带常绿阔叶树种,是国家重点保护的珍贵用材树种。培育优质苗木是闽楠人工林培育的物质基础,苗期营养管理是影响苗木质量的关键技术环节。王东光等[13]研究表明:磷素供应量为30 mg·株−1最有利于闽楠苗木的生长及抗低温胁迫能力的提高。王艺等[14]研究了缓释肥加载对闽楠容器苗生长及养分库构建的影响,结果表明:缓释肥加载有助于提高闽楠苗氮含量,且施肥量为3.0 kg·m−2时实现氮氧分库的最大积累量。但这些研究都局限于采用一种固定的施肥量、施肥时间去研究闽楠的养分需求,忽略了苗木异速生长的特性。不同时期苗木生长速度不同,其对养分的需求量也不相同,按苗木异速生长规律定期供应恰当的施肥量既满足了苗木需求,又能节约使用肥料。指数施肥是苗木稳态营养加载一种较为先进的施肥方式,指在苗木培育过程中,根据苗木生长对养分的需求规律,每次施肥量呈指数增加,提供植物养分的量同步于植物生长量,把肥料尽可能多地固定在苗木体内以形成养分库,保持苗木体内尽可能多的养分浓度,造林后苗木就会利用这一养分库促进根系和顶梢生长。苗期提高苗木氮含量能够促进造林后的早期生长,尤其是营养亏缺以及存在竞争的环境,因此苗期的氮素施用是苗木培育的重点[15-16]。为此,本研究以闽楠1年生闽楠苗为研究材料,采用盆栽试验,设置不同的氮素供应水平开展指数施肥试验,以了解不同指数施氮量对闽楠苗的生长生理以及养分积累的影响,采用分析闽楠苗生物量、氮素质量分数、株含量以及施氮量之间的关系和临界值法确定本试验条件下的最适指数施氮量,以期为生产中按闽楠指数生长需求进行氮素供应,最大限度发挥肥料效应,提高苗木质量提供理论依据。
-
供试闽楠苗为生长正常且均匀一致的1年生实生苗,苗高为(13.8±0.5) cm,地径为(2.09±0.08) mm。闽楠苗栽培基质为灭菌后的腐殖质、泥炭土按1∶1(质量比)的混合基质,基质理化性质见表1。供试肥料为普罗丹高浓度水溶性复合肥,其主要养分质量分数为氮200.000 g·kg−1,五氧化二磷(P2O5)200.000 g·kg−1,氧化钾(K2O)200.000 g·kg−1,螯合铁(Fe)1.000 g·kg−1,螯合锰(Mn)0.500 g·kg−1,螯合锌(Zn)0.500 g·kg−1,螯合铜(Cu)0.500 g·kg−1,硼(B)0.200 g·kg−1,钼(Mo)0.005 g·kg−1,EDTA螯合体10.000 g·kg−1。
腐殖质∶泥炭土 全氮/
(g·kg−1)水解氮/
(mg·kg−1)全磷/
(g·kg−1)速效磷/
(mg·kg−1)全钾/
(g·kg−1)速效钾/
(mg·kg−1)有机质/
(g·kg−1)pH 1∶1 1.0 1.5 62.8 69.4 2.0 5 076.4 336.4 4.2 Table 1. Basic physical and chemical properties of the substance
-
在贵州大学林学院苗圃采用盆栽法进行施肥试验。植苗容器为外口径20 cm,深15 cm的塑料花盆,每盆装入1 kg腐殖质、泥炭(1∶1,质量比)混合基质。为了防止水肥流失,花盆底部套有双层白色塑料袋。植苗时间为3月初,缓苗4周后开始施肥。为了减小边际效应,试验过程中,隔2周移动1次花盆。盆栽过程中浇水量根据基质最大持水量并视天气和苗木生长状况进行适当调整。
采用指数施肥模型来确定施肥量。指数施肥模型为NT=NS(erT−1)。其中:NT为需施养分总量,NS为施肥前苗木体内养分含量,T为施肥总次数,r为相对添加速率,公式的具体意义参照CHEN等[17]和USCOLA等[18]的研究。
试验开始前,随机抽取20株幼苗,确定幼苗初始生物量(1.281 g·株−1)及初始氮(5.165 g·kg−1)、磷(0.810 g·kg−1)、钾(4.327 g·kg−1)质量分数。设置施氮量0、1、2、3、4、5 g·株−1共6个处理,分别用ck、N1、N2、N3、N4、N5表示,每个处理30株苗,苗木移栽后第4周开始施肥,每周1次,共12次。氮素指数施肥量见表2。
处理 施氮量/(g·株−1·周−1) 总量/(g·株−1) 4 5 6 7 8 9 10 11 12 13 14 15周 ck 0 0 0 0 0 0 0 0 0 0 0 0 0 N1 0.003 0.005 0.008 0.012 0.018 0.028 0.042 0.077 0.086 0.149 0.235 0.336 1 N2 0.004 0.006 0.010 0.017 0.027 0.044 0.070 0.113 0.182 0.293 0.472 0.760 2 N3 0.004 0.007 0.019 0.014 0.034 0.056 0.094 0.156 0.260 0.433 0.721 1.201 3 N4 0.005 0.008 0.014 0.023 0.040 0.067 0.115 0.196 0.334 0.570 0.972 1.657 4 N5 0.005 0.008 0.015 0.026 0.044 0.077 0.134 0.233 0.405 0.704 1.223 2.125 5 Table 2. Schedule of fertilizer additions by seedling age for P. bournei
-
试验期间,分别于移苗第6、9、12、15周末尾,测定全株养分含量,试验结束时(即移苗后第15周末尾),测定苗高、地径、叶面积、生物量、叶绿素质量分数、根系活力及叶片养分质量分数。
生物量及全株养分含量测定:每个处理随机取3株苗木,用游标卡尺测量地径,精度0.01 mm;钢卷尺测量苗高,精度0.1 cm;将苗木用自来水洗净晾干后置于烘箱中105 ℃杀青20 min,然后75 ℃下烘48 h至恒量后称干质量,精度0.001 g;然后将每处理苗的地上和地下部分干质量充分混合,研磨后分别采用扩散法、钼锑抗比色法和火焰光度计法测定全株全氮、全磷、全钾质量分数,详见LY/T 1270−1999《森林植物与森林枯枝落叶层全硅、铁、铝、钙、镁、钾、钠、磷、硫、锰、铜、锌的测定》。
叶绿素质量分数及根系活力测定:每个处理随机抽取3株苗木,每株摘取第3、4片初展叶,用去离子水洗净晾干,用体积分数为80%丙酮与95%乙醇1∶1混合浸提叶片,采用紫外-可见分光光度计测定叶绿素质量分数;取每株苗木根部,去离子水洗净,采用改良TCC法测定苗木根系活力[19]。
叶片养分质量分数测定:每个处理随机取3株苗木,每株摘取第3、4片初展叶,用去离子水洗净晾干后于75 ℃下烘48 h至恒量后研磨成粉,测定叶片全氮、全磷、全钾质量分数,测定方法同全株养分测定。
-
数据采用Excel 2007处理,采用SPSS 22.0进行差异显著性分析。
-
施肥促进闽楠苗高、地径、叶面积的生长以及生物量的积累(表3)。与对照相比,5个施肥处理闽楠苗苗高、地径、叶面积、生物量分别增加134.1%~157.7%、62.9%~88.5%、104.3%~366.8%、174.9%~269.9%。不同施肥处理之间,各生长指标随指数施氮量的增加均呈现先增加后降低的趋势,N3处理苗高、地径、叶面积、生物量均最大,且地径、叶面积、生物量与其他施肥处理差异显著(P<0.05)。不同处理闽楠苗根冠比随指数施氮量的增加而减小。
处理 苗高/cm 地径/mm 叶面积/cm2 生物量/g 根冠比 ck 18.20±0.78 c 2.880±0.091 e 140.83±6.70 e 1.923±0.011 f 2.200±0.162 a N1 42.60±0.74 b 4.690±0.075 d 424.87±12.00 bc 5.287±0.006 e 1.150±0.018 b N2 44.60±0.96 b 4.990±0.044 c 472.22±28.50 b 6.355±0.009 d 1.060±0.031 bc N3 46.90±0.64 a 5.430±0.050 a 657.39±38.70 a 7.114±0.010 a 0.940±0.042 c N4 45.10±0.32 ab 5.240±0.066 b 380.88±22.90 c 6.758±0.010 b 0.880±0.043 cd N5 43.70±0.68 b 5.070±0.098 c 287.67±4.40 d 6.699±0.010 c 0.820±0.023 d 说明:数据为平均值±标准差。同列不同字母表示同一指标不同处理间差异显著(P<0.05) Table 3. Growth indices of P. bournei seedlings in different treatments
-
施肥可提高闽楠苗叶绿素a和叶绿素b质量分数,不同指数施肥效果不同(图1A)。叶绿素a、叶绿素b以及总叶绿素质量分数从大到小的顺序为N3、N2、N4、N5、N1,N3处理叶绿素a与叶绿素b质量分数分别为16.532和3.641 mg·g−1,与其他处理差异显著(P<0.05)。最低与最高施氮量处理之间叶绿素b质量分数差异不显著(P>0.05)。施氮量超过3 g·株−1,叶绿素a和叶绿素b质量分数下降明显,低于N1、N2处理。不同指数施氮量处理闽楠苗根系活力从大到小依次为N3、N4、N5、N2、N1(图1B),其中,N3处理闽楠苗根系活力最大,比N4处理高1.33%,两者差异不显著(P>0.05)。N1、N2处理闽楠苗根系活力较低,但两者差异不显著(P>0.05)。

Figure 1. Chlorophyll content and root activityof P. bournei seedlings in different treatments
-
施肥促进闽楠苗生物量及养分积累,不同处理表现出的积累规律不同(表4)。移苗12周之前,各处理闽楠苗生物量随指数施氮量的增加而增加;12周之后,N3处理生物量增加速率高于其他处理,且在15周苗木生物量最大,与N4、N5处理差异不显著。氮质量分数随指数施氮量的增加及施肥次数的增加而增加,N5处理氮质量分数在15周最高,显著高于其他处理(P<0.05)。随指数施氮量及施肥次数的增加,闽楠苗磷质量分数及磷株含量均呈增加趋势,N5处理闽楠苗磷质量分数在整个施肥阶段均高于其他处理。钾素积累在移苗12周前与磷素积累规律一致,移苗12周后N5处理闽楠苗磷株含量增加速率明显降低,15周之后钾株含量低于N3及N4处理,三者差异显著(P<0.05)。
移栽后周数/周 处理 生物量/(g·株−1) 养分质量分数/(mg·g−1) 养分株含量/(mg·株−1) 氮 磷 钾 氮 磷 钾 6 ck 1.43±0.01 d 8.4±0.2 d 0.47±0.07 c 0.64±0.01 f 12.05±0.44 f 0.67±0.10 c 9.15±0.09 f N1 1.59±0.02 c 8.8±0.1 d 0.57±0.06 b 0.79±0.01 e 14.03±0.17 e 0.91±0.09 b 12.54±0.09 e N2 1.64±0.01 b 11.1±0.2 c 0. 62±0.09 b 0.84±0.01 d 17.33±0.30 d 1.01±0.14 b 13.78±0.10 d N3 1.63±0.00 b 11.1±0.2 c 0.64±0.04 b 0.94±0.01 c 17.89±0.28 c 1.04±0.07 b 15.34±0.07 c N4 1.69±0.02 a 11.2±0.4 b 0.65±0.03 b 0.98±0.01 b 20.64±0.07 b 1.10±0.05 b 16.58±0.11 b N5 1.71±0.01 a 11.4±0.3 a 0.74±0.04 a 1.00±0.02 a 24.57±0.10 a 1.26±0.07 a 17.00±0.09 a 9 ck 1.55±0.01 e 9.8±0.1 e 0.57±0.05 e 0.68±0.01 f 14.22±0.14 f 0.88±0.07 e 10.48±0.14 f N1 1.95±0.01 d 13.0±0.3 d 0.64±0.03 d 0.82±0.01 e 25.39±0.12 e 1.25±0.04 d 15.97±0.10 e N2 2.23±0.01 b 13.2±0.1 d 0.72±0.05 c 0.89±0.01 d 29.41±0.16 d 1.60±0.07 c 19.89±0.15 d N3 2.12±0.01 c 13.8±0.1 c 0.76±0.03 c 0.95±0.01 c 29.23±0.10 c 1.61±0.06 c 20.22±0.12 c N4 2.24±0.01 b 14.2±0.1 b 0.82±0.06 b 1.06±0.01 b 31.86±0.11 b 1.84±0.10 b 23.81±0.12 b N5 2.32±0.01 a 14.8±0.1 a 0.88±0.04 a 1.12±0.01 a 34.38±0.10 a 2.04±0.06 a 26.02±0.14 a 12 ck 1.77±0.01 f 10.6±0.1 c 0.66±0.06 e 0.76±0.01 f 1.88±0.01 f 1.17±0.10 f 13.42±0.07 f N1 4.22±0.01 e 15.0±0.8 b 0.74±0.02 d 0.90±0.01 e 61.58±0.32 e 3.12±0.09 e 37.78±0.04 e N2 4.57±0.01 d 15.2±0.1 b 0.82±0.02 c 1.00±0.01 d 69.51±0.32 d 3.75±0.07 d 45.91±0.15 d N3 4.61±0.01 c 15.6±0.1 b 0.86±0.03 c 1.07±0.04 c 71.88±0.29 c 3.96±0.11 c 49.16±0.11 c N4 4.93±0.01 b 16.6±0.2 a 0.92±0.01 b 1.12±0.01 b 81.77±0.11 b 4.53±0.06 b 55.31±0.10 b N5 5.08±0.01 a 17.2±0.3 a 0.98±0.03 a 1.18±0.01 a 87.36±0.07 a 4.98±0.12 a 60.15±0.08 a 15 ck 1.92±0.01 d 11.4±0.2 e 0.72±0.04 f 0.78±0.01 e 21.92±0.32 e 1.38±0.08 f 14.94±0.09 f N1 5.29±0.01 c 15.2±0.2 d 0.78±0.01 e 1.01±0.03 d 80.36±0.14 d 4.12±0.04 e 53.14±0.07 e N2 6.36±0.01 b 15.8±0.4 cd 0.88±0.02 d 1.07±0.03 c 100.41±0.08 c 5.59±0.10 d 67.72±0.04 d N3 7.11±0.01 a 16.4±0.6 bc 0.94±0.01 c 1.10±0.04 c 116.67±0.10 b 6.69±0.07 c 78.08±0.10 b N4 6.96±0.01 a 16.8±0.4 b 1.04±0.02 b 1.15±0.03 b 116.89±0.16 b 7.03±0.10 b 77.63±0.08 c N5 7.10±0.01 a 17.6±0.2 a 1.26±.001 a 1.23±0.02 a 124.94±0.11 a 8.44±0.07 a 82.16±0.09 a 说明:数据为平均值±标准差。同列不同字母表示同一指标不同处理间差异显著(P<0.05) Table 4. Nutrient accumulation of P. bournei seedlings in different treatments
-
由图 2可以看出:氮素供应为0~3 g·株−1时,闽楠苗生物量、氮质量分数、氮株含量随氮素供应量的增加而增加,营养状况表现为养分亏缺。氮素供应增加至3 g·株−1时,闽楠苗生物量最大,为7.114 g·株−1,养分供应能够满足生长需求。氮素供应超过3 g·株−1时,闽楠苗生物量未发生显著变化,但氮株含量仍增加且达差异显著水平(P<0.05),表现为奢养消耗状态。养分供应过剩产生毒害通常表现为生物量及养分质量分数的减少[20]。试验未产生养分毒害现象。

Figure 2. Responses of biomass, N concentration and N content of P. bournei seedlings to increasing N supply
-
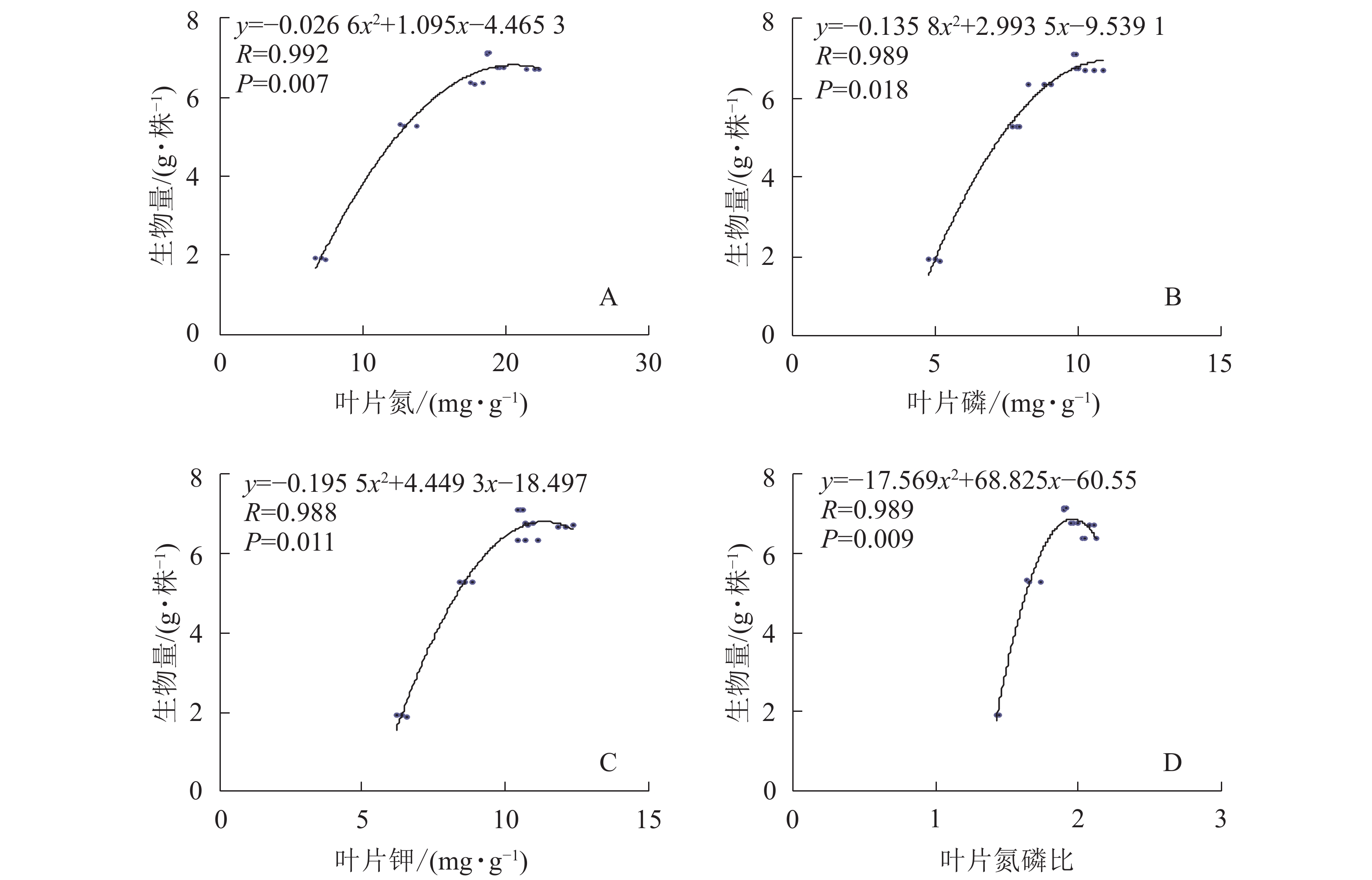
绘制闽楠苗叶片氮、磷、钾质量分数及氮磷质量分数比值与生物量的散点图,添加趋势线可以看出两者呈抛物线关系(图3)。以抛物线方程中最大生物量的90%相对应的2个养分质量分数作为临界值,两者之间的值为最适范围[11],则闽楠苗叶片氮、钾质量分数及氮磷质量分数比值临界值分别为16.88、9.78 mg·g−1和1.86,最适临界范围分别16.88~20.58、9.78~11.38 mg·g−1和1.86~1.96。叶片磷质量分数虽与生物量呈抛物线关系,但是从图3B可以看出:磷质量分数与生物量关系图难以准确估算临界值和最适质量分数范围,不适宜用于确定适宜施氮量。因此,根据叶氮和钾质量分数以及氮磷质量分数比值的最适范围,确定闽楠幼苗的适宜施氮量范围分别是2~4、2~4和3~4 g·株−1,综合三者最适质量分数范围得出,闽楠幼苗的最适氮素量为3~4 g·株−1。

Figure 3. Parabolic relationship of leaf nutrient concentration and biomass of P. bournei seedlings
-
苗木培育中养分供应方式对于苗木形态、生理、养分状况有重要影响,进而也影响培育苗木的质量及出圃后的生长表现。苗木生长不同阶段需肥量不同,供肥太快太多,会造成土壤养分因来不及被植物吸收而流失,相反如果供肥速度太慢太少则不能满足植物的生长需要。通过合理的施肥手段为植物提供充足的养分,创造良好的营养环境,提高植物营养效率,最终提高植物质产量是植物营养学研究的目的之一[21]。指数施肥以适应苗木养分需求规律的方式为苗木生长提供养分,一定程度上有利于苗木生长及质量提高。POKHAREL等[22]的研究中指数施肥较常规施肥方式显著提高苗木生物量及氮素质量分数。本试验以指数施肥方式施用氮素,探讨对闽楠苗生长生理以及养分积累的影响。结果表明:不同指数施氮量对闽楠苗生长的影响表现为随施氮量的增加各生长指标先增加后降低,以3 g·株−1生长表现最好,规律与叶绿素质量分数与根系活力表现一致。植物对氮、磷、钾的吸收会随供氮量的增加而增加[23]。试验中闽楠苗氮、磷、钾质量分数在整个施肥阶段随施氮量的增加而增加,与ZHAO等[24]、刘欢等[25]的研究结果一致。
植物氮素奢养消耗表现为生物量不变,氮素含量增加[26-27]。对西南桦Betula alnoides的指数施肥研究表明:施氮量为100~400 mg·株−1时表现为奢养消耗[17]。本试验奢养消耗表现在施氮量3~5 g·株−1。当施肥量超过奢养消耗所需养分的最大值时,通常会造成养分毒害而抑制植物生长。红橡木Quercus rubra[28]施氮量超过100 mg·株−1,黑云杉Picea mariana[20]施氮量超过64 mg·株−1会造成毒害。本试验中施氮量最高处理未出现生物量下降现象,未造成养分毒害。VILLAR-SALVADOR等[29]对橡树Quercus coccifera和Q. faginea的研究也未观察到养分毒害现象,原因可能是包括闽楠在内的这些植物具有较其他植物更高的养分需求。
林木叶片的养分含量与林木生长状况密切相关,是林木营养诊断的主要指标[30-31]。临界值法是植物养分诊断中分析叶片养分浓度的常用方法,容易受环境条件、取样方法的影响而使得某些元素的临界浓度无法诊断出。本试验中叶片氮钾质量分数比、磷钾质量分数比与生物量不呈抛物线关系,磷质量分数与生物量虽呈抛物线关系,但试验范围内未达到最大值,故不适合计算临界质量分数值以及最适施氮量。尾叶桉Eucalyptus urophylla 的营养诊断研究中由于试验环境影响,钾的临界浓度未诊断出[32]。综合氮、钾临界质量分数以及氮磷质量分数比临界值,闽楠适宜施氮量为3~4 g·株−1。氮素供应与生物量、养分质量分数之间的关系分析表明:此施氮量范围闽楠苗表现为养分的奢养消耗。造林后,苗木的成活和生长由苗木质量和造林地条件共同决定[29]。造林初期,由于苗木根系吸收能力较弱,其所需要的养分主要依赖储存器官养分内转移,因此,处于奢养状态的苗木有较强的抗性和较高成活率[33-34]。闽楠苗生长生理指标在3 g·株−1处理中表现最好,但养分积累表现以4 g·株−1优于3 g·株−1,且部分生长生理指标2个施肥量之间差异不显著,因此,综合闽楠生长生理以及养分积累情况,以指数施氮量4 g·株−1更有利于苗木培育过程中的养分储备以及出圃后的生长。
Growth, physiology and nutrient accumulation of Phoebe bournei seedlings under different amount of exponential nitrogen fertilization
doi: 10.11833/j.issn.2095-0756.20190259
- Received Date: 2019-04-09
- Rev Recd Date: 2019-12-12
- Available Online: 2020-05-21
- Publish Date: 2020-06-01
-
Key words:
- silviculture /
- exponential fertilization /
- Phoebe bournei /
- nitrogen /
- nutrient accumulation
Abstract:
| Citation: | WANG Xiao, WANG Yinglin, WEI Xiaoli, et al. Growth, physiology and nutrient accumulation of Phoebe bournei seedlings under different amount of exponential nitrogen fertilization[J]. Journal of Zhejiang A&F University, 2020, 37(3): 514-521. DOI: 10.11833/j.issn.2095-0756.20190259

|

 DownLoad:
DownLoad: