







-
马尾松Pinus massoniana曾经是南方集体林区的主要造林树种,在荒山绿化、生态服务和发展经济中起过不可替代的重要作用。但长期大面积纯林营造,林地出现地力衰退、生产力降低、生态服务功能下降等不良现象,特别是马尾松林经营长期遭受松材线虫Bursaphelenchus xylophlius病的困扰。据国家林业和草原局统计,近30 a来,全国因松材线虫病损失的松树累计达数十亿株,造成的直接经济损失和生态服务价值损失高达上千亿元,松材线虫病已成为全球森林生态系统中最具危险性、毁灭性的病害之一[1]。对于松材线虫病发生的疫区疫点,相关办法都要求“加强疫情科学除治,全面实施以清理病死、枯死、濒死松树为核心措施”。各地据此采取了皆伐、择伐等干扰方式清除疫木,以防止松材线虫的进一步扩散。大多相关研究集中在松材线虫病入侵后皆伐形成的除治迹地上马尾松林群落演替、林分结构、植物多样性和种群生态位的评价上[1-4]。但采伐疫木对受害马尾松林综合林分结构长期影响的相关研究较少。林分结构不但包括树种、胸径、树高、年龄、林木个体大小差异等非空间结构,也包括林木分布格局、单木竞争指数等空间结构,是林分未来生长变化的决定性因素之一[5-6]。在生产实践中,采伐疫木后的马尾松群落结构将会发生哪些变化,有必要对受害马尾松林的健康经营开展研究。本研究在浙江省杭州市临安区选择松材线虫病入侵过的马尾松人工纯林,以未受松材线虫病危害的林分为对照,探讨疫木卫生伐后的天然更新对马尾松人工纯林林分结构的影响,比较2种类型的林分在树种组成、胸径结构、树高结构、林木个体大小差异、聚集指数和竞争指数等方面的差异,以期为松材线虫病入侵后的马尾松林分的经营管理提供参考。
HTML
-
研究样地位于浙江省杭州市临安区(29°56′~30°23′N,118°51′~119°52′E)。该地属中亚热带季风气候,温暖湿润,四季分明,具有春多雨,夏湿热,秋气爽,冬干燥的气候特征,全年平均气温16.4 ℃,年均降水量1 628.6 mm,无霜期年平均为237 d[7]。该地先锋群落为马尾松林,地带性植被为亚热带常绿阔叶林,优势种主要有青冈Cyclobalanopsis glauca、苦槠Castanopsis sclerophylla等。研究区气候条件适宜马尾松的生长,但长期遭受松材线虫病危害。
-
在杭州市临安区河桥镇选择1994年同一批造林的马尾松纯林作为研究对象。21世纪初该马尾松人工纯林有部分遭受松材线虫病,为防止病害蔓延,2005年开始每年进行卫生伐,其他马尾松林木均予以保留,保护天然更新的非松科Pinaceae植株,持续7 a,共采伐约1 500株·hm−2。疫木伐除后,任其自然恢复更新。2017年8月,按照立地环境基本因子一致的原则,从中分别选择未入侵过的小块马尾松林(对照)和卫生伐过的马尾松林作为调查对象,2种类型的林分分别设置了5块30 m×30 m的标准样地。样地土壤为黄壤,土层厚度为0.8 m,坡向为西北坡,坡度35°~40°,立地条件差,pH 6.2。样地边界和林木坐标用全站仪测定,对样地进行了每木检尺,起测胸径为5 cm,调查因子包括树种、胸径、坐标、树高等。样地基本情况见表1。
处理 密度/(株·hm−2) 胸径/cm 树高/m 郁闭度 马尾松 阔叶树 马尾松 阔叶树 马尾松 阔叶树 对照 1 800 13.0±2.4 12.5±1.7 0.9 卫生伐 300 1 000 17.6±2.8 7.1±1.8 12.5±2.2 5.6±1.1 0.6 Table 1. Characteristics of P. massoniana stands after sanitation thinning diseased trees
-
物种丰富度是指样地内出现的物种数目。树种重要值是研究某个种在群落中的地位和作用的综合数量指标,物种丰富度和树种重要值反映了植物多样性。重要值计算采用的公式为:重要值
$= $ (相对多度+相对频度+相对显著度)/3。其中:相对多度$= $ (某个种的多度/所有物种的多度之和)×100%;相对频度$= $ (某个种的频度/所有物种的频度之和)×100%;相对显著度$= $ (某个种的胸高断面积之和/所有物种的胸高断面积之和)×100%。 -
林木个体大小差异在植物种群中普遍存在,相邻林木个体的大小会影响目标个体的胸径、高度的生长,在群落发展和维持群落物种多样性方面也具有深远意义,反映植物个体大小差异的指数采用库兹涅茨系数(KC)、基尼系数(GC)和洛伦茨不对称系数(LAC)[8]。
用二元材积公式[9]计算马尾松和阔叶树的单株林木材积,计算公式为[4]:
式(1)~(2)中:V为单株材积(m3);D为胸径(cm);H为树高(m)。
库兹涅茨系数(KC)是指单株材积为林分前20%的林木个体的材积之和在林分蓄积量中所占的比例,这个指数的数值越大,表示林木分化越严重,个体差异越大。基尼系数( GC)可用于研究林分内各林木之间差异性的大小[4, 8],计算公式为:
式(3)中:n为林分内林木的株数;xi为林木个体按材积大小升序排列时对应的第i株林木的材积;u为林分平均材积。洛伦茨不对称系数(LAC)用于反映林木个体大小等级对不一致性程度的贡献,当LAC>1时,表示不一致性的产生主要在少数较大的林木个体中;当LAC<1时,表示不一致性的产生主要在大多数较小的林木个体中[4, 8]。将林分n株林木按单株材积从小到大排序(x1,x2,…,xn),LAC的计算公式为:
式(4)~(5)中:m为恰好小于林木平均材积μ的林木序号;Lm为前m株林木个体材积之和;Ln为n株林木个体材积之和;θ为参数;Xm为排序第m株林木的材积;Xm+1为排序第m+1株林木的材积。
-
聚集指数是由CLARK等[12]提出的,用于检验种群空间分布格局的常用指数,是最早采用的与距离有关的空间格局指数[13]。采用最近邻单株距离的平均值与随机分布下的期望平均距离之比来表示:
式(6)中:R为聚集指数;ri为第i株树木到其最近邻木的距离;N为样地内林木株数;S为样地面积。若R>1,林木呈均匀分布;若R<1,则林木趋于聚集分布;若R
$= $ 1,林木则表现为随机分布。 -
竞争指数采用HEGYI[14]的计算公式:
式(7)中:Ci为对象木i的竞争指数;Lij为对象木i与竞争木j之间的距离;di为对象木i的胸径;dj为竞争木j的胸径;n为对象木i的竞争木株数,本研究采用4邻近树法,n
$= $ 4。 -
采用Excel 2003进行数据处理与图表制作,SPSS 20.0进行单因素方差分析(one-way ANOVA),LSD方法进行显著性检验(α
$= $ 0.05)。利用ArcGIS和ForStat软件对2种林分进行空间分析。数据为平均值±标准差。
2.1. 样地调查
2.2. 非空间结构指数及计算方法
2.2.1. 植物多样性指数




2.2.2. 林木个体大小差异指数
2.3. 空间结构指数及计算方法
2.3.1. 聚集指数

2.3.2. 竞争指数

2.4. 数据处理

-
由表2可知:马尾松人工纯林的重要值仍然为100.00%。在受害马尾松纯林疫木卫生伐后的林分中,马尾松的重要值下降至38.41%,仍然为群落的第1优势种;出现了大量天然更新的阔叶树种,短柄枹Quercus glandulifera的重要值为35.59%,是群落的第2优势种;檵木Loropetalum chinense的重要值为10.91%,是群落下层林分中主要的伴生树种;白栎Quercus fabri的重要值为10.54%,是继短柄枹之后的又一主要乔木树种;另外还有少量的无患子Sapindus mukorossi、枫香Liquidambar formosana、薄叶山矾Symplocos anomala和合欢Albizia julibrissin等乔木树种分布。可见,松材线虫入侵后的马尾松人工纯林在疫木卫生伐后经过长期恢复已演替为马尾松落叶阔叶混交林。而且该混交林分没有发现马尾松的小树和幼树,预示着未来该林分的阔叶树重要值将超过马尾松,形成阔叶树马尾松混交林。
处理 树种 重要值/% 对照 马尾松 100.00 卫生伐 马尾松 38.41 短柄枹 35.59 檵木 10.91 白栎 10.54 无患子 2.24 枫香 1.25 薄叶山矾 0.55 合欢 0.52 Table 2. Tree species composition of P. massoniana stands after sanitation thinning infected trees
-
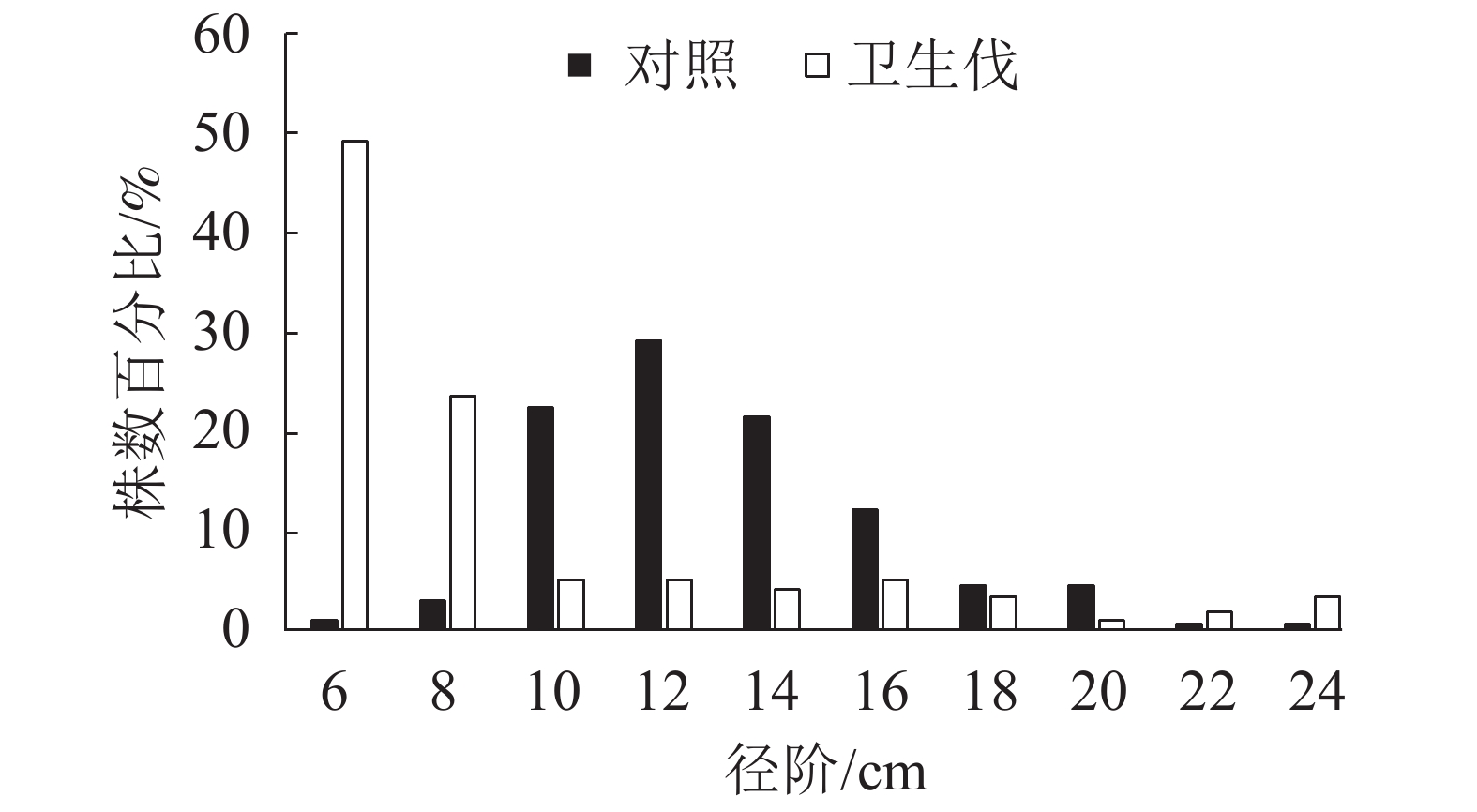
由图1可知:马尾松人工纯林的林分平均胸径为13.0 cm,其中,10、12和14 cm径阶的林木株数最多,分别为22.8%、29.6%和21.8%,呈单峰状左偏正态分布,小径阶和大径阶林木各占一小部分。疫木卫生伐后形成的马尾松阔叶混交林林分平均胸径为9.1 cm,林木大多数分布在6和8 cm径阶,株数分别占49.2%和23.4%,而且随着胸径的增大呈下降趋势,呈现反J型曲线,较大径阶22和24 cm的株树比例比马尾松人工纯林高。未遭受松材线虫入侵的马尾松林依然保持单层林;疫木卫生伐后经过长期恢复由单层纯林演变为由马尾松占据的主林层与阔叶树组成的次林层的复层混交林分(表1)。
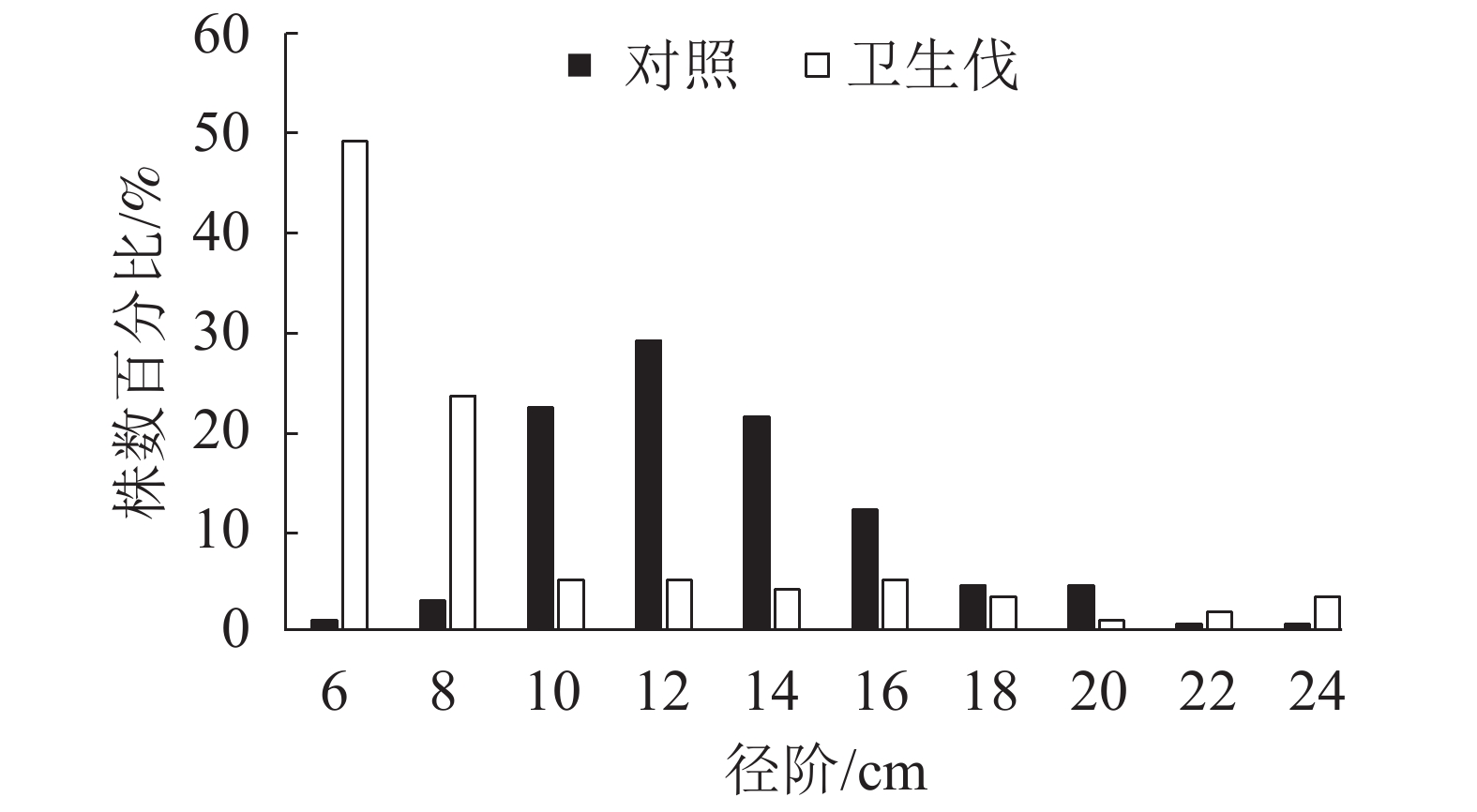
Figure 1. DBH structure of P. massoniana stands after sanitation thinning infected trees
-
由表3可知:马尾松人工纯林疫木卫生伐后,库兹涅茨系数(KC)增加,说明较大个体林木和较小个体林木分化更严重;基尼系数(GC)增加,说明林木个体大小不一致性增大;洛伦茨不对称系数(LAC)由略小于1增加为略大于1,说明林木个体差异的产生主要在于大多数较小的林木个体变为少数较大的林木个体。
处理 KC GC LAC 对照 0.35±0.08 a 0.26±0.05 a 0.97±0.15 a 卫生伐 0.74±0.13 b 0.66±0.09 b 1.04±0.23 a 说明:同列不同字母表示同一指标不同处理间差异显著 (P<0.05) Table 3. Individual size inequality of P. massoniana stands after thinning diseased trees
-
由表4可知:马尾松人工纯林聚集指数(R)>1,表现为均匀分布,疫木卫生伐后形成的马尾松阔叶林的R
$= $ 0.95<1,呈现聚集分布。马尾松人工纯林的平均竞争指数(Ci)为8.16,疫木卫生伐后形成的马尾松阔叶林的平均竞争指数增加至10.12。处理 聚集指数(R) 竞争指数(Ci) 对照 1.27±0.21 a 8.16±3.52 a 卫生伐 0.95±0.14 b 10.12±5.60 b 说明:同列不同字母表示同一指标不同处理间差异显著 (P<0.05) Table 4. Aggregation index and competition index of P. massoniana stands after sanitation thinning infected trees
由图2可知:马尾松人工纯林各径阶处受到的竞争压力基本一致,未发生很大的变化。疫木卫生伐后形成的马尾松阔叶林于6 cm径阶处林木受到的竞争压力最大,其次是8 cm径阶林木,各径阶处受到的竞争压力差异较大。
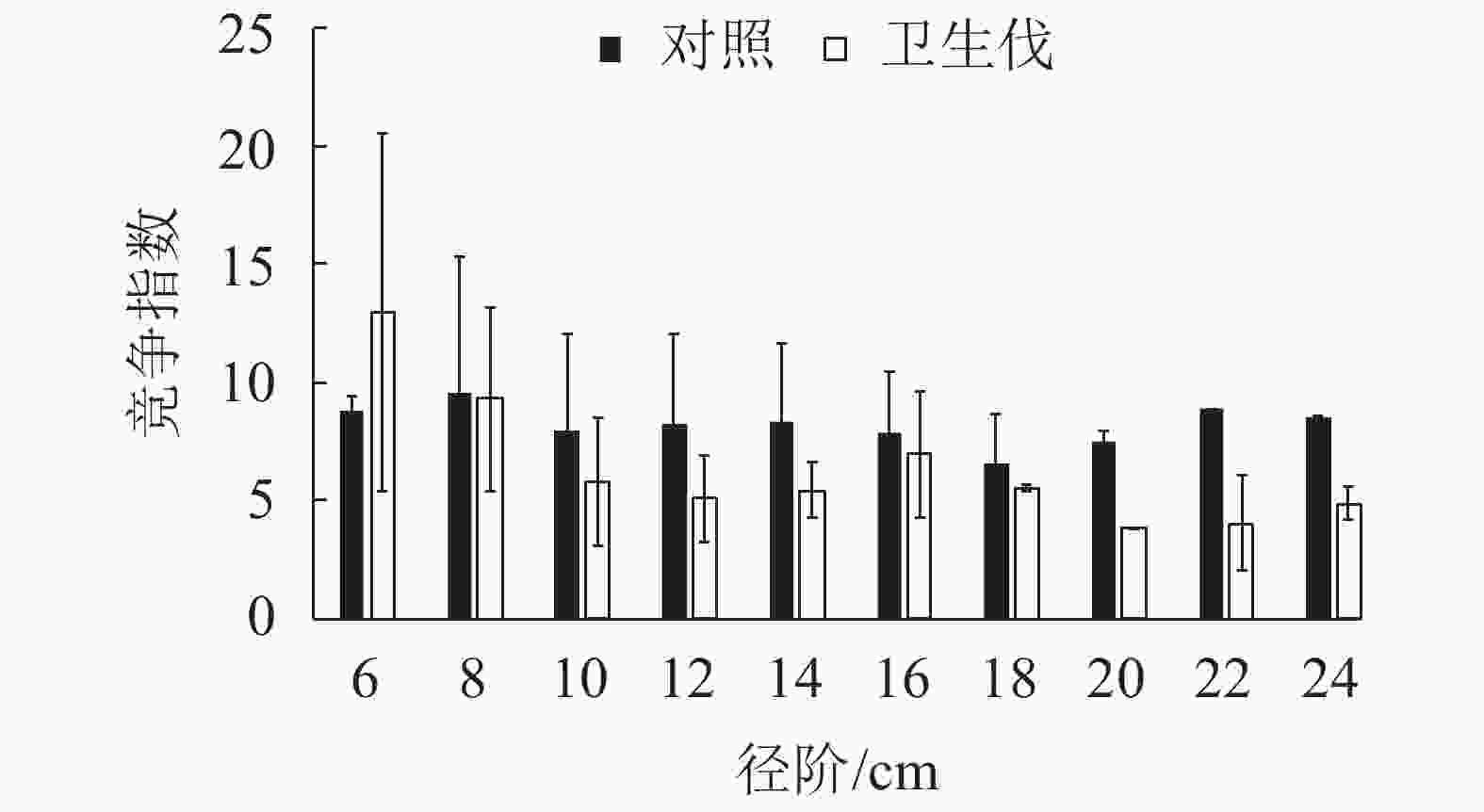
Figure 2. Distribution of DBH-competition index of P. massoniana stands after sanitation thinning infected trees
3.1. 疫木卫生伐对树种组成的影响
3.2. 疫木卫生伐对胸径和树高结构的影响
3.3. 疫木卫生伐对林木个体大小差异的影响
3.4. 疫木卫生伐对聚集指数和竞争指数的影响


-
本研究对入侵过的马尾松林连续几年开展疫木卫生伐,腾出林分生长空间,促进阔叶树种的天然更新。疫木采伐十几年后,受害马尾松林未再发现疫木,但是松木损失严重,只剩300株·hm−2马尾松。说明林业部门规定的及时卫生伐的处理措施在实践中有一定效果,能在一定程度上抵御林分尺度的松材线虫病的自然扩散。
中国营造的人工林大多为纯林,马尾松林也不例外。马尾松人工纯林的林分结构极其简单,树种单一,乔冠草各层次的植物多样性低[15-16],这为松材线虫病的入侵扩散提供了便利条件。松材线虫入侵马尾松林后,如果不加以人为干预,松林必然全部死亡[17]。本研究采取的疫木卫生伐对原马尾松纯林的林分结构产生了重要影响,经过十几年的恢复,使得马尾松人工纯林显著具有天然林的特点,具体表现在:树种组成由马尾松单一树种变为马尾松和落叶阔叶树种短柄枹为主的马尾松落叶阔叶混交林,进入了马尾松纯林的下一个演替阶段;胸径结构由正态分布变为反J型曲线,异龄林特征明显;垂直结构由单层林变为复层林;林木分布从均匀分布变为随机分布;林木大小个体差异变大,两级分化加剧。上述变化是松材线虫入侵松林后人为间伐干扰的结果;也与马尾松林是亚热带的先锋群落,群落本身并不稳定紧密相关[1]。
疫木卫生伐后形成的林窗会加快群落天然更新速度,短柄枹、白栎等演替早期的落叶乔木阔叶树种迅速占领了松木采伐移除后的生态位(表2),促使受害马尾松纯林向混交林演替的速度加快。这与王懿祥等[8]的研究结果相同,疫木卫生伐提高了林分林木个体大小的不一致性(表3)。这是由于疫木卫生伐后出现了大量天然更新的阔叶树,这部分林木个体较小;另外一方面,疫木卫生伐促进了保留木(马尾松)的生长,特别是加快了原优势木的生长,使其成长为较大的林木个体。因而也使得林木个体大小不一致性变为由少数较大的林木个体产生(LAC<1);林分的胸径结构变为反J型曲线(图1)。疫木卫生伐后形成的混交林竞争指数显著增加,林木之间的竞争压力变动加大(表4),这是由于大量天然更新的幼树其竞争指数较大的缘故,径阶-竞争指数关系图(图2)也反映了林木所受的竞争压力随着径阶的增大而减小,林木个体大小不一致性的增加也使得林木之间的竞争指数变动增加。在经营中应选出阔叶目标树,间伐邻近竞争木,减小竞争压力,促使阔叶目标树生长[18]。
马尾松人工纯林树种单一,结构简单,生态系统极其不稳定,对松材线虫的抵抗能力极差,应避免营造马尾松纯林[4]。松材线虫作为一个外来的选择因子,虽然会毁掉松林,但也能促使整个生态系统朝向更为稳定的方向发展。对于疫木卫生伐后形成的马尾松与以短柄枹和白栎为主的混交林而言,也正处于演替早期阶段,本身并不稳定。在森林经营条件许可的情况下,可以在林分内选择目标阔叶树,间伐邻近林木以减少竞争压力促使目标树生长;同时参考当地地带性植被常绿阔叶林的树种组成,及早补植地带性植被的建群种和优势种以及伴生种,如青冈、苦槠、木荷Schima superba等,加快群落演替速度,提高抗病害能力。
 DownLoad:
DownLoad: