







-
氮是植物生长必需的营养元素之一。大气氮沉降属于酸沉降的一种[1],是指陆地生态系统产生的含氮化合物排放到大气中,经过一系列复杂的化学反应后再次降落回陆地生态系统的过程[2]。20世纪50年代之后,随着工业全球化的推进,化石燃料的燃烧以及人口急速膨胀,大气中的活性氮也随之不断升高[3-4],并不断趋于全球化[5]。在长期氮沉降的环境下,进入陆地生态系统的活性氮已经远远超出了陆地生态系统本身的氮需求[6]。中国氮沉降现象日渐严重[7],已成为氮沉降世界三大高降区之一[8],中国农业和工业集约化地区的氮沉降问题尤其严峻[8-9],并且有不断增长的趋势[10]。在此背景下,氮沉降对森林土壤呼吸的影响已经成为近年来生态学关注的热点。土壤中磷元素的缺失是制约植物生产力的主要因素之一[11]。由于中国长江以南的大片亚热带丘陵山地以酸性土壤为主,土壤中原本含量较少的活性磷受强烈的吸附固定作用难以移动,从而导致了土壤能被植物体直接吸收的有效磷含量更低,呈现严重缺磷状态[12]。土壤中磷元素的缺乏,限制了植物体的生长发育从而抑制了土壤呼吸作用。并且最近几年大气氮沉降所带来的负面效应不断加剧[13],使土壤受到低磷胁迫的形势变得更加严峻[14]。森林土壤与精耕细作的田间土壤相比,所受到的低磷胁迫形式及程度显得更加复杂[14]。南方林地的土壤一般为红壤[15],土壤中有效磷含量较低一直是限制南方林地生产力的重要因素之一[16]。近年来,由于二氧化碳(CO2)、甲烷(CH4)、含氮化合物(NxO)等温室气体的排放,大气中的CO2相比工业革命之前上升了近1倍[17-18],其中全球土壤中的碳排放量达6.8×1013 kg·a−1[19]。土壤是陆地生态系统的主要碳库,其中植物土壤呼吸所产生的CO2是组成陆地生态系统碳循环过程的重要部分[20-22]。杉木Cunninghamia lanceolata是中国南方常绿针叶速生树种,为中国南方造林面积最大的用材树种之一[23],栽培历史长达1 000多年[24-25]。庞丽等[15]模拟了氮沉降环境下土壤氮磷比的变化,但模拟氮沉降情况下杉木林土壤呼吸对低磷胁迫的响应还鲜有报道。本研究通过模拟氮沉降试验,分析了杉木林在不同土壤磷水平环境下土壤碳排放的动态机制,为分析处于氮沉降不断增加和磷胁迫日趋严峻环境下的南方人工林的科学经营提供相关依据。
HTML
-
研究区处于浙江省杭州市临安区高坎村(30°21′N,119°67′E),该区属中亚热带季风气候区,气候较为温和,四季分明,雨量充沛,年平均降水量为1 632.6 mm,年平均气温为16.4 ℃,全年日照时数1 847.3 h,年均无霜期约230 d[26]。土壤为黄壤,地形地貌为低山丘陵,森林覆盖率76.5%。
-
2018年12月,选择10年生杉木幼龄林。在样地内选取生长情况相似且高度约3 m的杉木,并以此为中心设立1个3 m×3 m的独立小样方。为了避免试验干扰,每个独立小样方之间设置不小于3 m的缓冲带,本研究共设置27个独立小样方。参照国际上氮沉降模拟方法,依据中国亚热带地区的实际氮沉降量及未来增加趋势[27-30],以当地氮沉降率30.5 kg·hm−2·a−1为基础[26],设置2个处理梯度:低氮(N30:30 kg·hm−2·a−1)和高氮(N60:60 kg·hm−2·a−1)。参考国内外相关研究[29-30],磷添加设置了2个处理梯度:低磷(P20:20 mg·kg−1)和高磷(P40:40 mg·kg−1)。另外,再设置4个氮磷复合处理[低氮高磷(N30+P40)、低氮低磷(N30+P20)、高氮高磷(N60+P40)、高氮低磷(N60+P20)]及对照(ck)。各处理重复3次。
土壤的速效磷水平将磷酸二氢钾(KH2PO4)均匀地洒在样方内,进行30 cm的翻耕,使土壤上层速效磷含量达P20和P40的2个供磷水平,此后不再对杉木林添加磷。同时,根据氮处理水平,从2019年1月开始,每月模拟氮沉降喷施1次。具体方法为:每月月初的晴天,将每个样方所需喷施的一定量的硝酸铵(NH4NO3)溶解在4 L自来水中,在杉木的树冠上方用背式喷雾器均匀喷洒[1]。对照喷洒同量的自来水,以减少处理间因外加自来水不同而造成的影响。
-
为了测定样地内的土壤呼吸速率,2018年12月,在每个样方内安装直径20 cm,高12 cm的PVC连接环,安装时使其露出地表5 cm。本研究采用动态封闭气室法,使用LI-8100(LI-COR Inc.)土壤碳通量自动测量系统,观测杉木林土壤呼吸速率。2019年1−10月,每月中旬选取1个连续3 d晴朗并且最接近当月天气状况的日子,测定杉木林的土壤呼吸速率,测量结果代表测定当月杉木林土壤呼吸速率的平均值,用来分析氮磷添加下杉木林土壤呼吸的季节性变化特征。研究表明:植物土壤呼吸速率在10:00左右最接近当天的平均值[30-31]。因此,本研究用10:00测量的值代表杉木林土壤呼吸速率的日平均值[32],测定时间段为当天的10:00前后。
为了减少安置土壤呼吸测定环对杉木林土壤呼吸速率的影响,土壤呼吸测定环埋好后固定永久放置,并且在每次测定前1 d,将样方内的土壤呼吸测定环内的表层植被在尽量不破坏土壤的情况下彻底去除,以减少根系损伤及土壤扰动对测量结果的影响。
在测定土壤呼吸的同时,用LI-8100所配备的TDR土壤水分速测仪土壤温度传感器和土壤水分速测仪分别测定5 cm土层深处的土壤温度和土壤含水量[33]。
-
杉木林土壤呼吸速率与土壤温度的关系模型为:RS=aebT。其中,RS表示土壤呼吸速率(μmol·m−2·s−1),T表示土壤温度(℃),a、b为待定参数。杉木林土壤呼吸的温度敏感性系数(Q10)的计算公式为:Q10=e10b。其中,b为公式RS=aebT计算中得到的常量。本研究采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较分析不同氮沉降和磷添加处理水平下杉木林土壤呼吸速率。利用SPSS 26.0软件分析数据,用Origin 2017软件制图。
1.1. 研究区概况
1.2. 样地设置与试验设计
1.3. 土壤呼吸速率、土壤温度和土壤湿度的测定
1.4. 数据分析
-
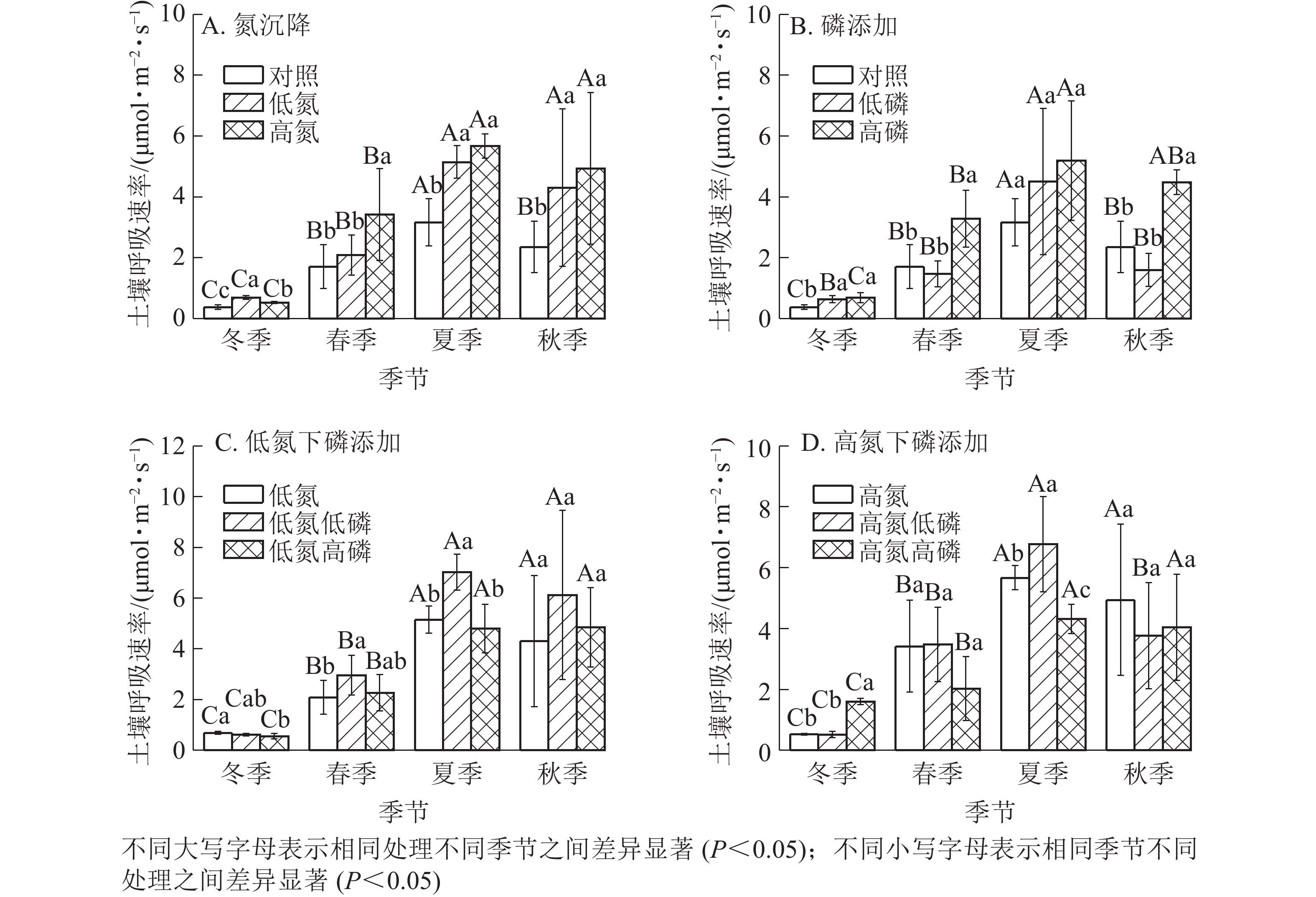
氮沉降、磷添加以及氮磷复合处理下杉木土壤呼吸速率都有明显的季节变化(图1),不同季节杉木林土壤呼吸作用差异显著(P<0.05),氮沉降和磷添加并没有改变杉木林土壤呼吸的季节性变化趋势。不同处理土壤呼吸均在夏季达到最高值,在冬季达到最低值。氮沉降显著促进了杉木林的土壤呼吸(P<0.05),其中在夏季促进作用最为显著(P<0.05);高氮处理对杉木林土壤呼吸的促进作用最为显著(P<0.05),在夏季与对照(ck)相比提高了1.78倍(图1A)。磷添加处理显著促进了杉木林的土壤呼吸(P<0.05),但低磷处理在春季和秋季对杉木土壤呼吸的影响不显著(P>0.05),甚至抑制了土壤呼吸作用(图1B)。磷添加处理对杉木林土壤呼吸的促进作用在夏季最为显著(P<0.05),高磷处理对杉木林土壤呼吸的促进作用最为显著(P<0.05),在夏季与对照(ck)相比提高了1.77倍(图1B)。在氮沉降下,磷添加对杉木林土壤呼吸产生了显著影响(P<0.05),其中高氮低磷处理对杉木林土壤呼吸的促进作用最为显著(P<0.05),在夏季与对照(ck)相比提高了2.12倍(图1C和图1D)。不同处理下的杉木林土壤呼吸均在冬季达到最低值,随着地表温度的上升杉木林土壤呼吸作用也不断增强,在夏季均达到最高值,随后不断降低,氮磷复合处理并没有改变杉木林土壤呼吸的季节性变化规律。
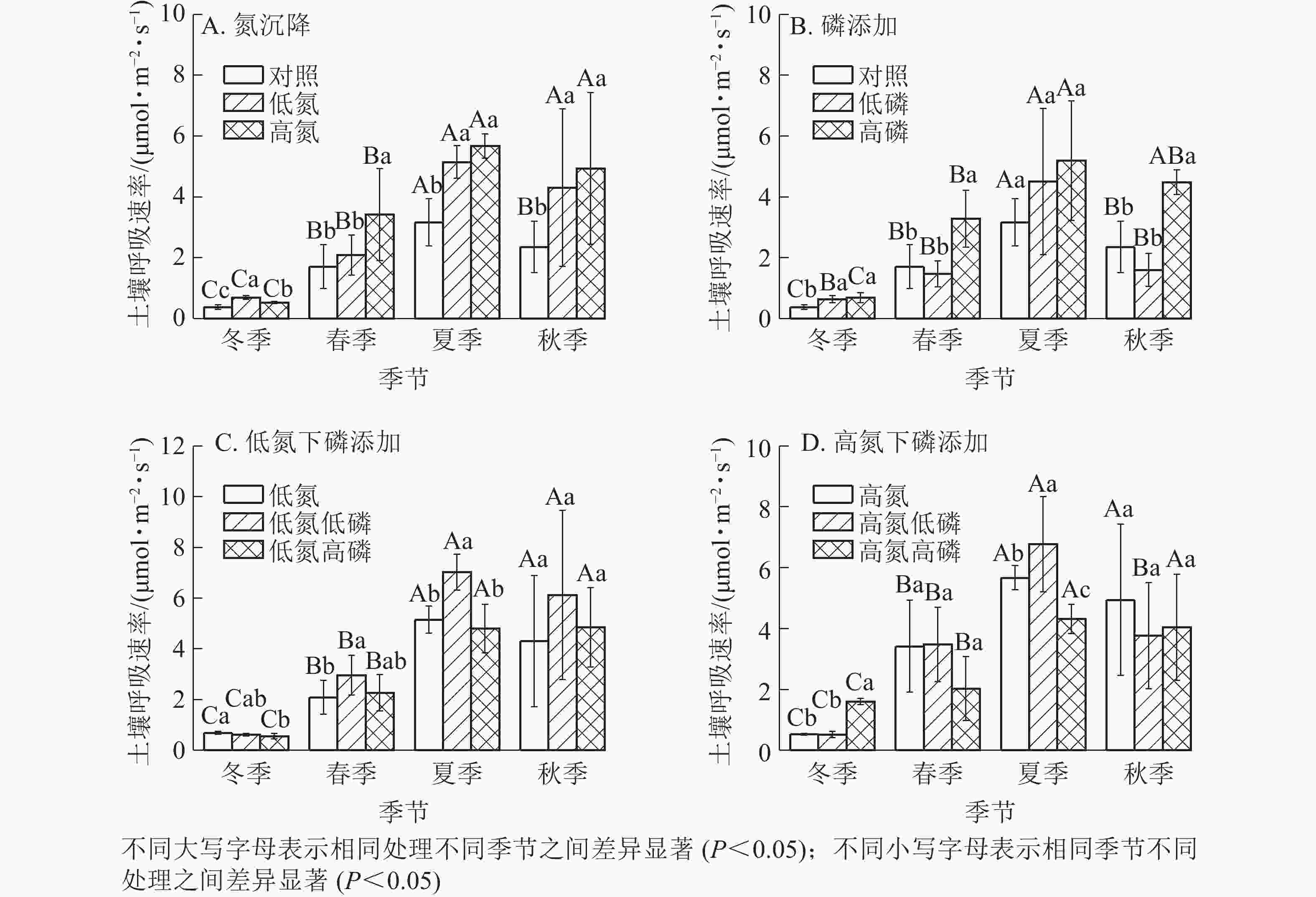
Figure 1. Seasonal change of soil respiration under different nitrogen and phosphorus treatments
-
杉木林土壤呼吸速率与土壤温度、土壤湿度存在着显著相关关系(P<0.05)(表1),杉木林土壤呼吸速率随着土壤温度的升高而升高,但随着土壤湿度的升高而降低。各处理下杉木林土壤呼吸速率和土壤温度均呈极显著正相关(P<0.01)(表2),除低磷处理外,杉木林土壤呼吸速率和土壤湿度也存在着显著(P<0.05)或者极显著负相关(P<0.01)(表2)。
环境因子 土壤呼吸速率 土壤温度 土壤湿度 土壤呼吸速率 1 土壤温度 0.834** 1 土壤湿度 −0.369** −0.449** 1 说明:双尾检验。**表示相关极显著(P<0.01) Table 1. Correlation between soil respiration and environmental factors
处理 土壤温度 土壤湿度 Q10 对照 0.930** −0.441* 3.318 低氮 0.964** −0.441* 3.087 高氮 0.930** −0.433* 3.318 低磷 0.751** −0.328 3.542 高磷 0.803** −0.520** 3.501 低氮低磷 0.972** −0.433* 3.626 低氮高磷 0.916** −0.433* 3.355 高氮低磷 0.908** −0.387* 3.393 高氮高磷 0.961** −0.581** 3.422 说明:双尾检验。*表示显著相关(P<0.05);**表示极显著相 关(P<0.01) Table 2. Correlation between soil respiration and environmental factors
土壤呼吸速率对土壤温度变化的敏感性可以通过Q10来反映。通过土壤呼吸速率和土壤温度拟合的指数方程可以得出:对照、低氮、高氮、低磷、高磷、低氮低磷、低氮高磷、高氮低磷、高氮高磷的Q10分别为3.318、3.087、3.318、3.542、3.501、3.626、3.355、3.393、3.422(表2)。可见,单独施加氮对杉木林土壤呼吸速率的敏感性几乎没有影响,甚至在一定程度上降低了杉木林土壤呼吸的敏感性;单独施加磷增强了杉木林土壤呼吸的敏感性;在氮磷复合作用下,杉木林土壤呼吸敏感性增强,其中低氮低磷处理对杉木林土壤呼吸的敏感性影响最大。
2.1. 杉木林土壤呼吸速率变化
2.2. 氮磷处理下杉木林土壤呼吸速率和土壤温度、湿度的Person相关分析
-
本研究发现:杉木林土壤呼吸速率均呈显著的季节性变化,氮沉降并没有改变杉木林土壤呼吸的季节性变化趋势,而季节使得土壤温湿度产生了变化,因此,土壤温湿度是影响土壤呼吸的主要因素。由于本研究区降水充沛,土壤环境相对湿润,土壤呼吸速率与土壤温度呈显著正相关,与土壤湿度呈负相关,说明过高的土壤湿度会使土壤的通透性变差,降低土壤中气体的交互,从而导致土壤呼吸作用受到抑制。模拟氮沉降处理对杉木林土壤呼吸敏感性变化的影响并不大,甚至降低了土壤呼吸敏感性,而单独磷添加以及氮磷复合处理均提高了杉木林土壤呼吸的敏感性,其中在低磷处理下,杉木林土壤呼吸的敏感度最大,可能是在模拟氮沉降处理初期对土壤有机物含量的促进作用并不显著,而在原本受到低磷胁迫的环境下施加了磷,从而提高了土壤中有机物含量,增加了土壤呼吸底物的含量以及土壤呼吸敏感性。
-
土壤呼吸分为2个过程:①微生物呼吸、根系呼吸和土壤中动物呼吸的生物呼吸;②含碳矿物质化学氧化作用的非生物学过程[34]。由于土壤中部分动物的呼吸作用和有机物质的化学氧化作用非常微弱,可以忽略不计[35-36],本研究便将土壤中微生物呼吸和植物根系呼吸定义为土壤呼吸的主要部分。同时,土壤呼吸也是陆地碳循环的重要组成部分,对全球气候变化和温室气体的排放都有着重大的影响。本研究表明:单一添加磷对杉木林土壤呼吸有显著影响。在低磷处理下,杉木林土壤呼吸先是稳定上升,到一定程度后呈逐渐下降的趋势。与对照相比,在低磷处理下,杉木土壤呼吸在春、秋季被抑制,可能是由于季节的更替导致地表植被以及凋落物增加,从而抑制了土壤呼吸;在高磷处理下,杉木林土壤呼吸显著提高,说明磷添加处理增加了土壤中有效磷的含量,改善了土壤所受到的低磷胁迫的影响,促进了杉木林土壤呼吸作用。这与薛美瑛等[37]发现的单独磷添加对植物土壤呼吸有促进作用的结果相似。
-
氮沉降对土壤呼吸的影响有3类:抑制作用、促进作用和无显著影响[37-40]。本研究发现:模拟氮沉降的10个月里,与对照相比,氮沉降明显促进了杉木林土壤呼吸速率,这是因为模拟氮沉降处理增强了杉木生长以及根系的发育,从而增强了杉木的自养呼吸。这与王泽西等[41]、向元彬等[42]和MADRITCH等[43]的结果相似。这可能是在氮处理的初期,土壤中的可用性氮增加,提高了土壤中微生物的活性,从而促进了土壤呼吸作用。而周世兴等[38]、MO等[44]研究表明:氮沉降抑制了土壤呼吸作用,这可能是因为施氮处理减少了林分凋落物量和细根生物量,从而改变了土壤微环境,降低了土壤中的微生物碳氮量,抑制了微生物活性,进而抑制了土壤呼吸作用[43]。邓琦等[45]研究表明:模拟氮沉降对土壤呼吸的影响不显著,原因是在降水量较多的地区,土壤含水率较高,抑制了土壤微生物的呼吸作用,因此,氮沉降作用对土壤呼吸速率没有较大影响。本研究表明:模拟氮沉降促进了杉木林土壤呼吸作用,可能是不同植物的土壤呼吸受到了不同氮需求的影响[42]。
-
本研究中,低氮低磷和低氮高磷处理的土壤呼吸速率显著高于低氮处理,高氮低磷处理的土壤呼吸速率显著高于高氮处理,而与高氮处理相比,高氮高磷处理的土壤呼吸速率受到了抑制。与对照相比,低氮和高氮处理下添加磷后显著促进了杉木林的土壤呼吸。低氮低磷、低氮高磷和高氮低磷处理同时增加了土壤中有效氮和有效磷的含量,提高了土壤中有机质含量,增强了土壤中微生物的生命活动,从而促进了土壤呼吸作用[45]。张彦东等[46]和薛美瑛等[37]研究发现:适当提高土壤中氮磷含量可以改善土壤的理化性质,使得土壤中可以被植物体直接吸收利用的有效磷含量提升,从而增加了植物体的生物量,增强了植物的土壤呼吸作用。与高氮处理相比,高氮高磷处理抑制了土壤呼吸作用,可能是高氮高磷降低了土壤的pH,植物体向地下输送的有机质有所减少,从而降低了土壤微生物活性,使土壤呼吸受到抑制。综上所述,氮沉降对土壤呼吸的影响,受到了不同植物对氮元素的需求以及所受到氮沉降作用时间的长短影响[45]。在氮沉降初期,增强了植物的生命活动,刺激了根系呼吸,促进了土壤呼吸作用,当土壤中氮水平达到饱和后,氮沉降对土壤呼吸起到抑制作用。在受到低磷胁迫较为严重的地区,磷添加增加了土壤中有效磷含量,促进了植物的生长,使土壤有机质含量增加,从而促进了土壤呼吸作用。可见,在氮沉降初期进行磷添加对陆地生态系统有较为积极的意义。
 DownLoad:
DownLoad: