







-
AP2/ERF基因家族是植物最大的转录因子家族之一,广泛参与植物生长发育和逆境胁迫响应等生物学过程,在植物遗传改良与育种方面具有重要应用价值[1-2]。AP2/ERF转录因子家族成员通常含有1~2个高度保守的AP2结构域,AP2结构域由60~70个氨基酸组成,构成典型的螺旋-转角-螺旋结构,通过特异性结合DNA以调节靶基因表达[3-4]。根据AP2结构域数量和特征序列,AP2/ERF基因家族可分为AP2、ERF、DREB、RAV和Soloist等5个亚家族[5]。一般来讲,AP2亚家族成员具有2个AP2结构域,在调控植物发育方面具有重要功能[6]。ERF、DREB和RAV亚家族成员则仅具有1个AP2结构域,其中RAV亚家族成员还具有1个B3结构域[7]。此外,其他具有特殊基因结构和类似AP2结构域的成员则属于Soloist亚家族[8]。
随着越来越多植物基因组被测序,AP2/ERF基因家族成员已在拟南芥Arabidopsis thaliana[5]、葡萄Vitis vinifera[7]、萝卜Raphanus sativus[9]、生姜Zingiber officinale[10]、花生Arachis hypogaea[11]、水稻Oryza sativa[12]、甘蔗Saccharum officinarum[13]、玉米Zea mays[14]和大麦Hordeum vulgare[15]等单双子叶植物中得到鉴定。对不同植物的研究表明:不同亚家族的AP2/ERF转录因子在植物发育和胁迫响应中发挥不同的功能。通常,AP2亚家族的转录因子参与了植物不同的发育过程,如拟南芥AP2基因调节开花时间[16-17],决定种子质量及大小[18];BBM基因能够促进体胚发生[6, 19-21]。ERF和DREB亚家族基因则与生物胁迫和环境因子胁迫响应有关[22],如水稻OsDREB基因就与植株对干旱、高盐、低温胁迫的耐受性有关[23],AtERF6等4个ERF基因在响应强光方面起重要作用[24]。而RAV亚家族基因则被认为在响应生物胁迫和非生物胁迫中发挥关键作用[25-26]。
光皮桦Betula luminifera属桦木科Betulaceae桦木属Betula珍贵用材树种,广泛分布于云南、贵州、广西、福建和浙江等。由于具有适应性强、速生以及材质优良等特点,光皮桦是南方山地造林的优良树种,兼具较高的经济价值和生态价值,其遗传育种工作也逐渐受到重视。因此,本研究利用基因组序列,对光皮桦AP2/ERF基因家族进行鉴定,对其进行理化性质、系统进化等生物信息学分析,同时通过表达分析对光皮桦AP2/ERF基因家族的组织表达特异性及对高温胁迫的响应进行研究,旨在为光皮桦AP2/ERF基因家族的功能研究及在遗传育种中的应用提供基础。
-
从Pfam数据库中(http://pfam.xfam.org/)下载AP2/ERF家族基因结构域的隐马尔可夫文件(PF00847),利用HMMER v3.1b1 软件筛选光皮桦基因组蛋白序列(未发表)中含有AP2/ERF结构域的序列(E≤e−5)。利用美国国家生物技术信息中心(NCBI) CDD Tools (
https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi )[27]对获得的候选基因进行结构域鉴定,去除其中结构域不完整的序列。最后将获得的光皮桦AP2/ERF基因上传至Genbank获得登录号。通过ExPaSy (https://www.expasy.org/ )在线分析预测工具[28],分别对光皮桦AP2/ERF基因的氨基酸数目、开放阅读框(ORF)长度、相对分子质量、理论等电点等性质进行分析。 -
利用MEGA 7软件对获得的光皮桦和拟南芥AP2/ERF基因家族的蛋白序列进行比对,参数为默认设置。拟南芥AP2/ERF蛋白序列下载于拟南芥数据库TAIR 。基于AP2结构域序列,利用MEGA软件采用邻接法(neighbor-joining,NJ)构建系统进化树,Bootstrap值设置为1000[29]。借助在线工具EvolView 绘制进化树图[30]。
-
利用MEME在线工具(
https://meme-suite.org/meme/ )对光皮桦基因蛋白序列上的保守基序进行预测,保守基序数量设置为10,其他参数为默认设置。采用TBtools软件绘制基因结构及保守基序图[31]。 -
利用PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )数据库[32],取光皮桦基因起始密码子上游2 000 bp作为启动子区域进行顺式作用元件预测。采用TBtools软件对启动子元件的分布进行绘图[31]。 -
利用STRING数据库(
https://string-db.org/ )[33],对光皮桦AP2/ERF蛋白相互作用网络进行预测,以拟南芥为参照物种,选择最低互动分为0.4,其他参数默认,借助Cytoscape软件绘制蛋白互作网络图[34]。 -
利用13个2年生光皮桦植株不同组织器官的转录组数据进行AP2/ERF基因组织表达特异性分析。13个不同组织器官包括根(Root)、嫩叶(YL)、成熟叶(ML)、雌花序(FC)、雄花序(MC)、第1~6节茎(S1~S6)、木质部(Xylem)及枝皮(Bark)。高温胁迫实验以6月龄光皮桦植株为材料,42 ℃处理12和36 h后取植株中部成熟叶片,以25 ℃生长植株成熟叶为对照,通过转录组测序分析AP2/ERF基因表达。提取FPKM值(a),进行log2a标准化处理,再借助TBtools软件绘制表达热图[31]。
-
通过生物信息学方法在光皮桦基因组数据库中筛选得到77条具有完整AP2结构域的基因序列(表1)。对这77个基因进行序列分析,结果显示:光皮桦AP2/ERF基因的 ORF长度为444~2121 bp,其编码的氨基酸为147~706个。其中:分子质量最小的蛋白是BlDREB2,仅16.65 kDa;分子质量最大的蛋白是BlAP2-6,达77.37 kDa;理论等电点为4.73~10.06,约77.9%的蛋白理论等电点小于7.0,说明光皮桦AP2/ERF蛋白含有较多酸性氨基酸(表1)。
分类 基因
名称登陆号 ORF
长度/bp氨基酸
数目/个分子量/
kDa理论等
电点分类 基因
名称登陆号 ORF
长度/bp氨基酸
数目/个分子量/
kDa理论等
电点AP2 BlAP2-1 ON092428 1527 508 55.95 6.20 ERF (B-1) BlERF1 ON092417 1131 376 41.40 6.56 BlAP2-2 ON092430 1494 497 55.01 6.58 BlERF4 ON092422 783 260 27.62 9.85 BlAP2-3 ON092431 1644 547 60.45 6.17 BlERF9 ON092433 633 210 22.91 6.85 BlAP2-4 ON092437 1203 400 44.64 8.37 BlERF15 ON092447 510 169 18.52 10.06 BlAP2-5 ON092446 1458 485 52.91 7.83 BlERF18 ON092458 468 155 16.80 9.81 BlAP2-6 ON092453 2121 706 77.37 6.39 BlERF33 ON092489 636 211 22.85 9.74 BlAP2-7 ON092456 975 324 37.20 5.44 BlERF34 ON092491 900 299 32.64 5.10 BlAP2-8 ON092457 1428 475 51.86 6.43 BlAP2-9 ON092466 1929 642 71.38 6.50 ERF (B-2) BlERF2 ON092418 942 313 34.89 5.65 BlAP2-10 ON092474 1956 651 71.92 6.53 BlERF13 ON092443 1137 378 42.28 5.12 BlAP2-11 ON092483 1614 537 58.88 5.95 BlAP2-12 ON092485 1068 355 39.52 7.16 ERF (B-3) BlERF3 ON092419 729 242 27.18 6.03 BlAP2-13 ON092493 1539 512 56.17 6.41 BlERF10 ON092438 837 278 29.98 6.46 BlERF11 ON092439 828 275 29.76 8.72 DREB (A-1) BlDREB6 ON092434 741 246 27.06 4.77 BlERF21 ON092468 996 331 36.46 6.03 BlDREB7 ON092435 714 237 26.13 4.89 BlERF22 ON092469 996 331 36.36 7.68 BlDREB8 ON092436 900 299 33.24 4.94 BlERF23 ON092470 996 331 36.52 6.67 BlDREB15 ON092454 678 225 25.22 8.58 BlERF24 ON092471 996 331 36.49 8.57 BlDREB17 ON092461 564 187 20.58 6.12 BlERF25 ON092472 1011 336 37.03 6.47 BlDREB23 ON092484 657 218 23.93 8.90 BlERF28 ON092477 699 232 25.68 6.21 BlDREB24 ON092487 612 203 22.27 5.47 BlDREB25 ON092488 609 202 22.14 5.48 ERF(B-4) BlERF6 ON092426 1092 363 39.30 6.51 BlDREB26 ON092490 621 206 22.93 5.48 BlERF26 ON092475 612 203 22.47 4.86 BlERF27 ON092476 657 218 24.47 6.13 DREB (A-3) BlDREB19 ON092463 972 323 34.95 6.58 BlERF31 ON092481 1383 460 49.08 5.72 BlERF32 ON092482 612 203 22.48 4.87 DREB (A-4) BlDREB5 ON092429 717 238 25.85 4.92 BlDREB9 ON092440 642 213 23.46 4.99 ERF(B-5) BlERF7 ON092427 924 307 35.10 5.14 BlDREB10 ON092441 558 185 20.22 4.73 BlERF17 ON092455 969 322 36.12 5.50 BlDREB12 ON092448 546 181 20.42 5.18 BlERF20 ON092464 1053 350 39.54 4.88 BlDREB14 ON092452 531 176 18.89 5.20 BlDREB16 ON092459 540 179 19.95 6.90 ERF(B-6) BlERF5 ON092423 549 182 20.42 6.71 BlDREB18 ON092462 603 200 21.85 4.95 BlERF8 ON092432 609 202 22.49 6.83 BlDREB21 ON092473 606 201 20.88 4.80 BlERF12 ON092442 624 207 23.23 6.98 BlDREB22 ON092480 684 227 24.39 4.81 BlERF14 ON092444 756 251 28.52 4.94 BlERF16 ON092450 996 331 36.52 5.00 DREB (A-5) BlDREB1 ON092420 615 204 22.57 5.11 BlERF19 ON092460 777 258 28.97 5.39 BlDREB2 ON092421 444 147 16.65 7.83 BlERF29 ON092478 573 190 21.52 8.85 BlDREB3 ON092424 699 232 24.96 5.16 BlERF30 ON092479 570 189 21.29 7.80 BlDREB11 ON092445 525 174 19.43 6.64 BlDREB13 ON092449 486 161 17.59 4.77 RAV BlRAV1 ON092451 948 315 35.16 9.00 BlDREB20 ON092467 624 207 22.78 4.76 BlRAV2 ON092465 1203 400 44.15 8.08 DREB (A-6) BlDREB4 ON092425 1134 377 41.15 5.54 Soloist BlSoloist ON092486 957 318 36.27 6.01 BlDREB27 ON092492 1008 335 36.97 6.18 Table 1. Gene features of AP2 /ERF gene family from B. luminifera
-
为了分析光皮桦AP2/ERF基因家族的进化关系,基于拟南芥及光皮桦AP2/ERF转录因子的AP2结构域序列,利用MEGA7软件构建了系统进化树。结果显示:这些转录因子可分为AP2、ERF、DREB、RAV和Soloist 等5个亚家族,并可进一步分成15个进化枝(图1)。光皮桦AP2/ERF基因家族在各亚家族中的分布并不均匀,其中AP2亚家族包含13个基因,占基因总数的16.89%;DREB亚家族包含27个基因,占基因总数的35.06%;ERF亚家族包含34个基因,占光皮桦AP2/ERF基因家族总数的44.15%;RAV 亚家族包含2个基因,Soloist亚家族仅有1个基因(表1,图1)。参考拟南芥等的研究[2],DREB亚家族可进一步分为6个亚类,即A1、A2、A3、A4、A5和A6;ERF亚家族可进一步分为6个亚类,即B1、B2、B3、B4、B5和B6。在光皮桦中,DREB亚家族成员集中分布在A1、A4、A5亚类上,其中A1和A4亚类均包含9个基因,A5亚类包含6个基因;光皮桦ERF亚家族的6个亚类分别包括7、2、9、5、3、8个成员(表1,图1)。

Figure 1. Phylogenetic tree of AP2/ERF transcription factors from B. luminifera and A. thaliana
-
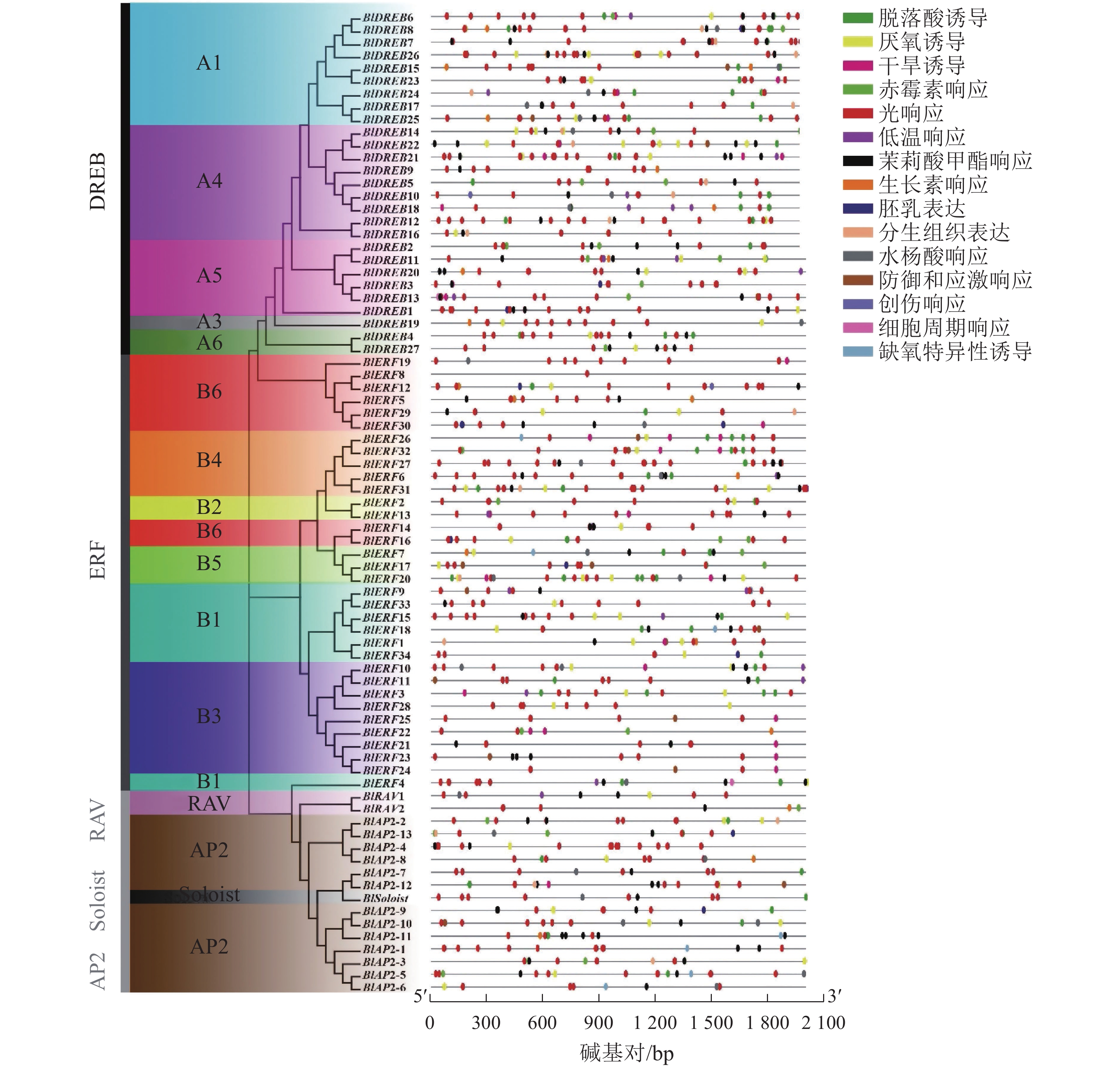
基因结构也是基因的保守性特征之一。对光皮桦AP2/ERF基因家族结构的分析显示:光皮桦AP2/ERF基因各亚家族间的基因结构存在较大差异。所有的AP2亚家族基因均具有多个内含子,其内含子数量为6~9个;除了BlDREB10具有1个内含子外,其余DREB亚家族成员均没有内含子(图2A)。此外,9个ERF和1个RAV亚家族成员具有1个内含子,其他成员则没有内含子;而BlSoloist则有5个内含子,与AP2亚家族成员比较相似(图2A)。
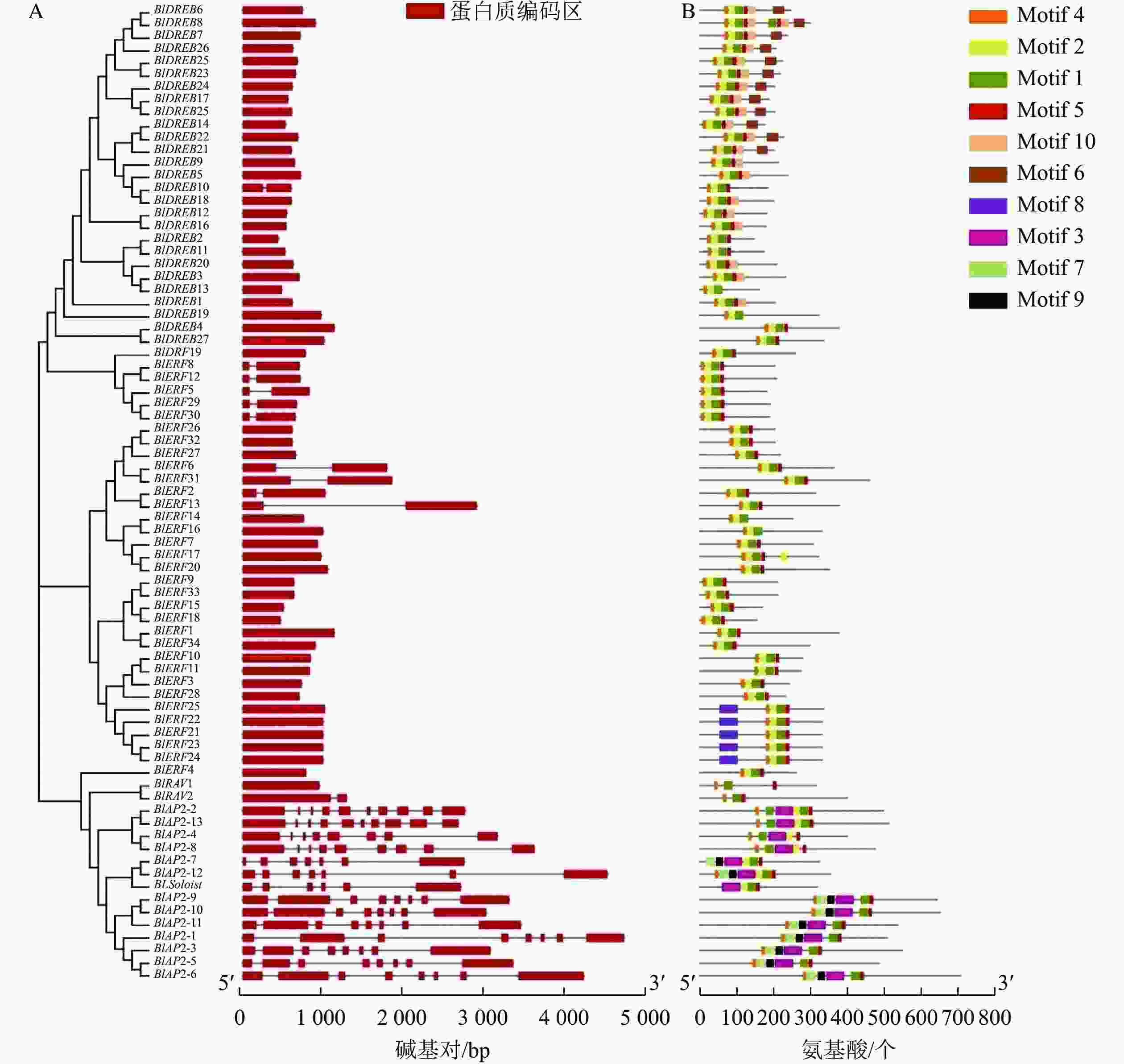
Figure 2. Gene structure (A) and conserved motif (B) of AP2/ERF family members in B. luminifera
进一步对光皮桦AP2/ERF转录因子的保守基序进行分析表明:绝大多数光皮桦AP2/ERF基因家族成员(71个)都同时拥有Motif 1、Motif 4和Motif 5这3个Motif,表明Motif 1、Motif 4和Motif 5是构成AP2结构域的主要基序(图2B)。同时,光皮桦同一亚家族基因的保守基序组成表现出高度的相似性,可能发挥相似的生物学功能。除了BlAP2-7外,其余AP2家族成员都具有Motif 4;2个RAV基因的保守基序较少,仅3个;大部分DREB亚家族成员特有Motif 10,少部分成员特有Motif 6。此外,ERF亚家族成员中仅BlERF21、BlERF22、BlERF23、BlERF24及BlERF25具有Motif 8,推测这5个基因发挥更特殊的生物学功能。
-
启动子区域的顺式作用元件在基因转录起始调控中起重要作用。利用PlantCARE数据库对77个光皮桦AP2/ERF基因家族启动子区域的顺式作用元件进行分析,共检测到1072个特异元件,大致可分成3类:激素响应、胁迫响应及生长发育相关元件。其中,激素响应元件有赤霉素、脱落酸、生长素等元件,胁迫响应元件包含缺氧、干旱、低温防御等相关元件,生长发育相关元件包含胚乳表达、分生组织表达等元件(图3)。光响应(492个)、脱落酸响应(180个)、茉莉酸甲酯响应(125个)及厌氧诱导(74个)元件是光皮桦AP2/ERF基因启动子序列所具有的主要顺式作用元件。其中,光响应元件在所有的基因启动子区域均有分布,除BlERF7和BlERF8仅包含1个光响应元件,其余基因包含多个光响应元件,如BlDREB8 (16个)、BlERF27 (15个)等。这提示光皮桦AP2/ERF基因可能参与了植株的光形态建成或光照相关的环境适应。而具有茉莉酸甲酯、脱落酸、赤霉素、生长素等激素响应元件的光皮桦AP2/ERF基因数量分别为61、63、20、18个,说明光皮桦AP2/ERF基因家族成员广泛参与了不同植物激素的信号途径。此外,仅BlDREB10和BlERF12具有创伤响应元件,仅BlDREB13、BlERF4和BlERF28具有细胞周期调控元件,这部分具有特殊元件的基因可能具有更特异的功能。
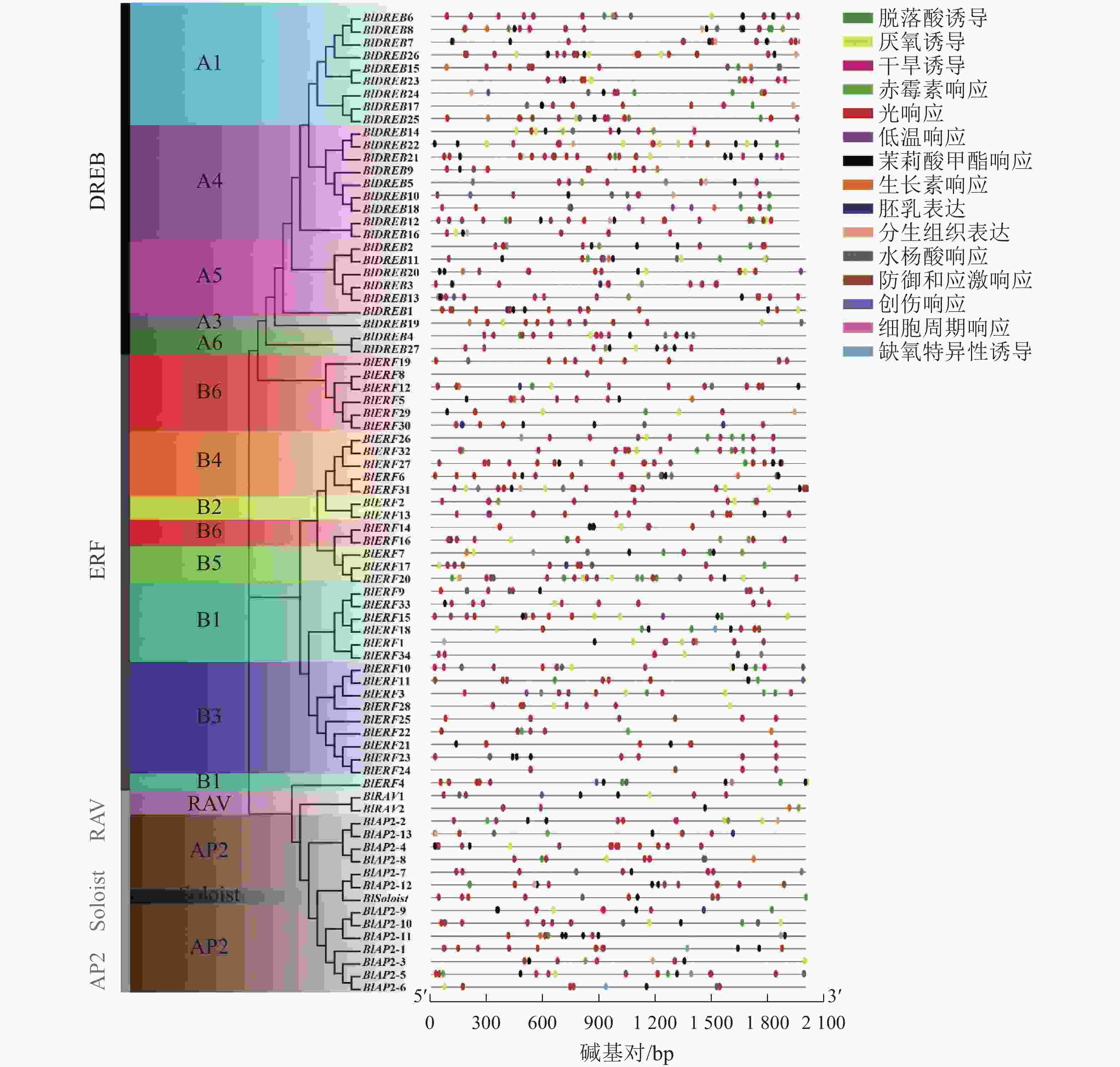
Figure 3. The cis-elements in promoter of AP2/ERF genes in B. luminifera
-
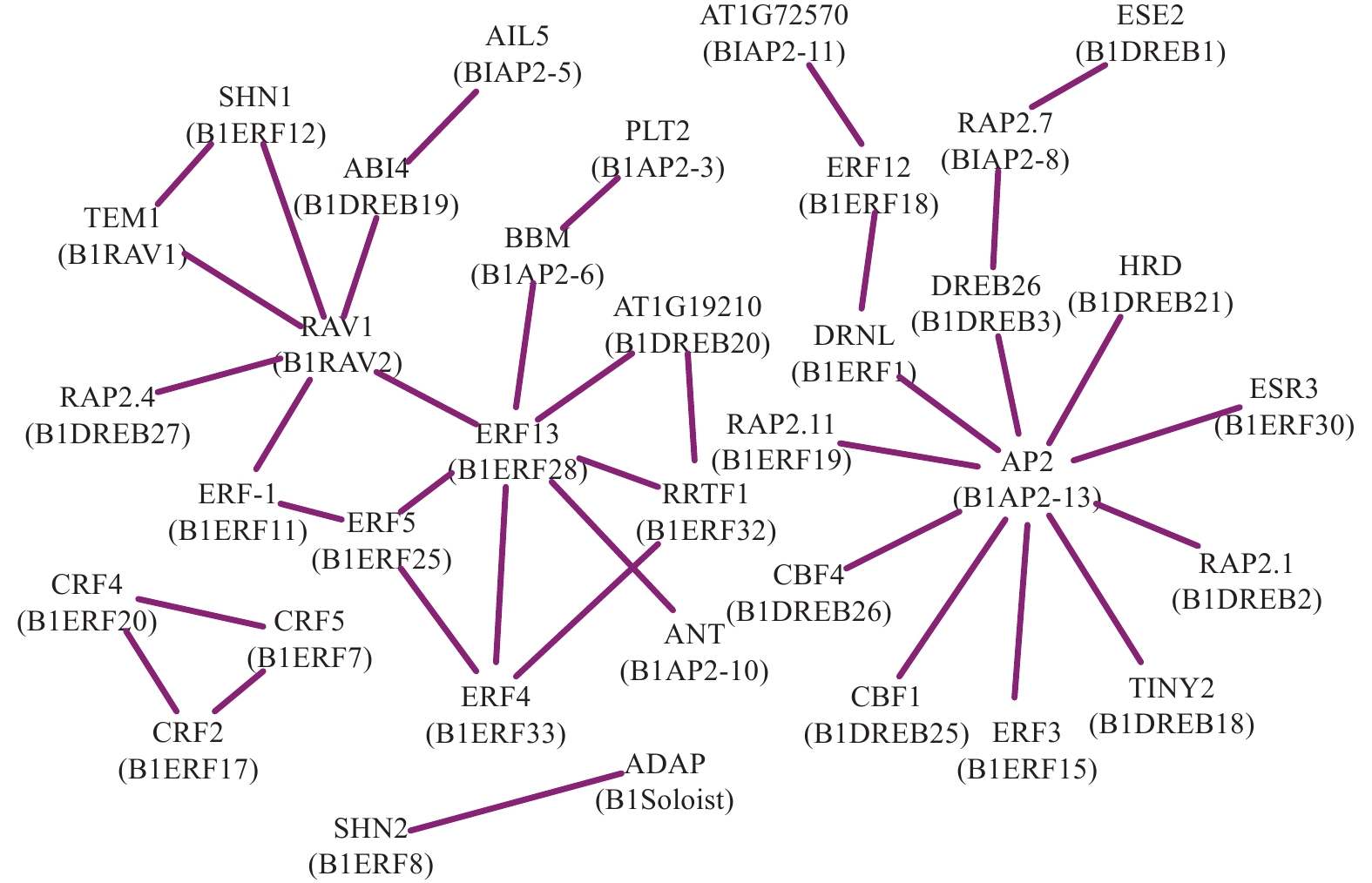
为了分析光皮桦AP2/ERF转录因子在调控过程中的协同作用,利用STRING数据库匹配到的49个拟南芥同源蛋白构建了相互作用网络,结果如图4所示。BlRAV2 (RAV1)、BlERF28 (ERF13)及BlAP2-13 (AP2)具有较多的节点(互作蛋白)数量,分别为6、7、10个,表明这3个基因在光皮桦AP2/ERF基因家族中可能占据核心地位并发挥着重要的生物学功能。此外,各亚家族成员内部及之间也存在复杂的相互作用,如BlERF7、BlERF17及BlERF20构成三角相互作用网络,而核心成员BlAP2-13与多个ERF及DREB成员分别存在互作关系,BlRAV2也与多个ERF和DREB成员互作(图4)。这种不同亚家族成员间复杂的互作关系,暗示这些基因可能在功能上存在非常复杂的交互关系。
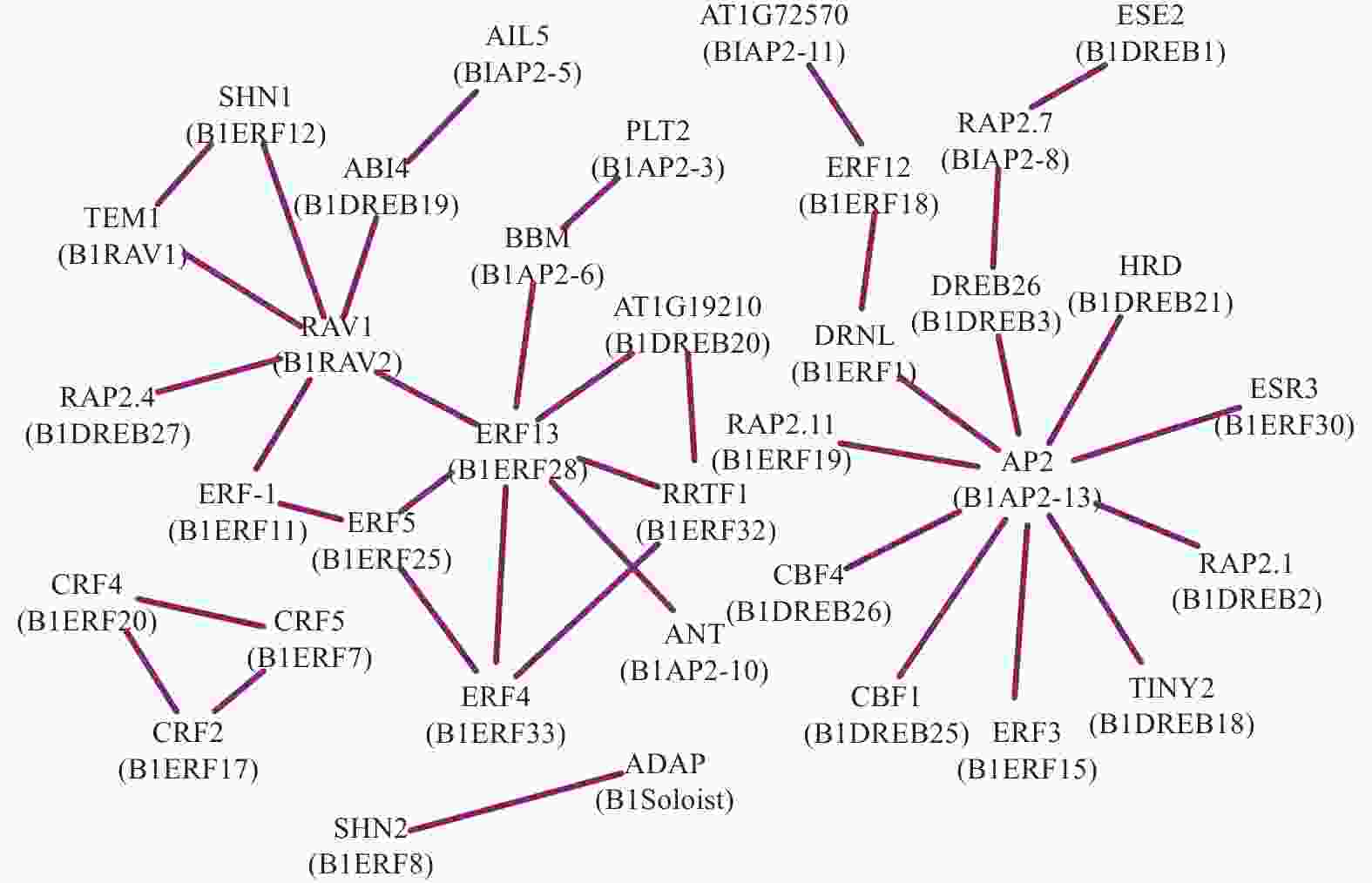
Figure 4. Prediction of interactions of AP2/ERF proteins in B. luminifera
-
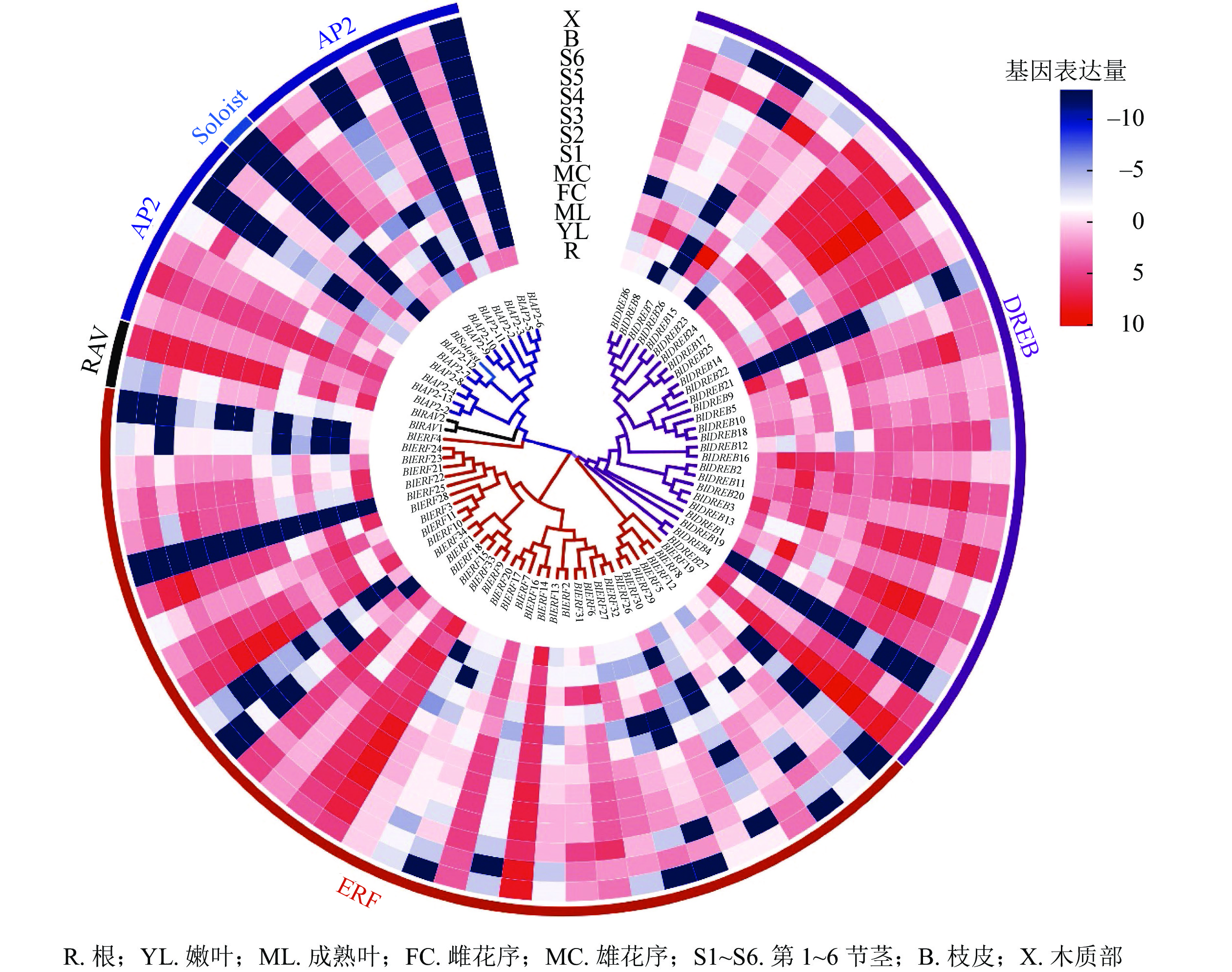
对77个光皮桦AP2/ERF家族成员在根、嫩叶、成熟叶、雌花序、雄花序、不同木质化的茎、皮、木质部等8个不同组织器官的表达情况进行分析。结果显示:多数(71个)家族基因的表达具有较强的组织特异性(图5)。其中,只在特定组织或器官中表达或表达量较强的基因有4个,如BlAP2-6只在根中表达、BlSoloist仅在嫩叶及第1节茎中表达;在所有组织器官中均有表达且表达量较高的基因有5个,分别为BlAP2-13、BlERF9、BlERF33、BlERF13、BlDREB27;而BlERF25在这13个组织器官中的表达量均极低(图5)。
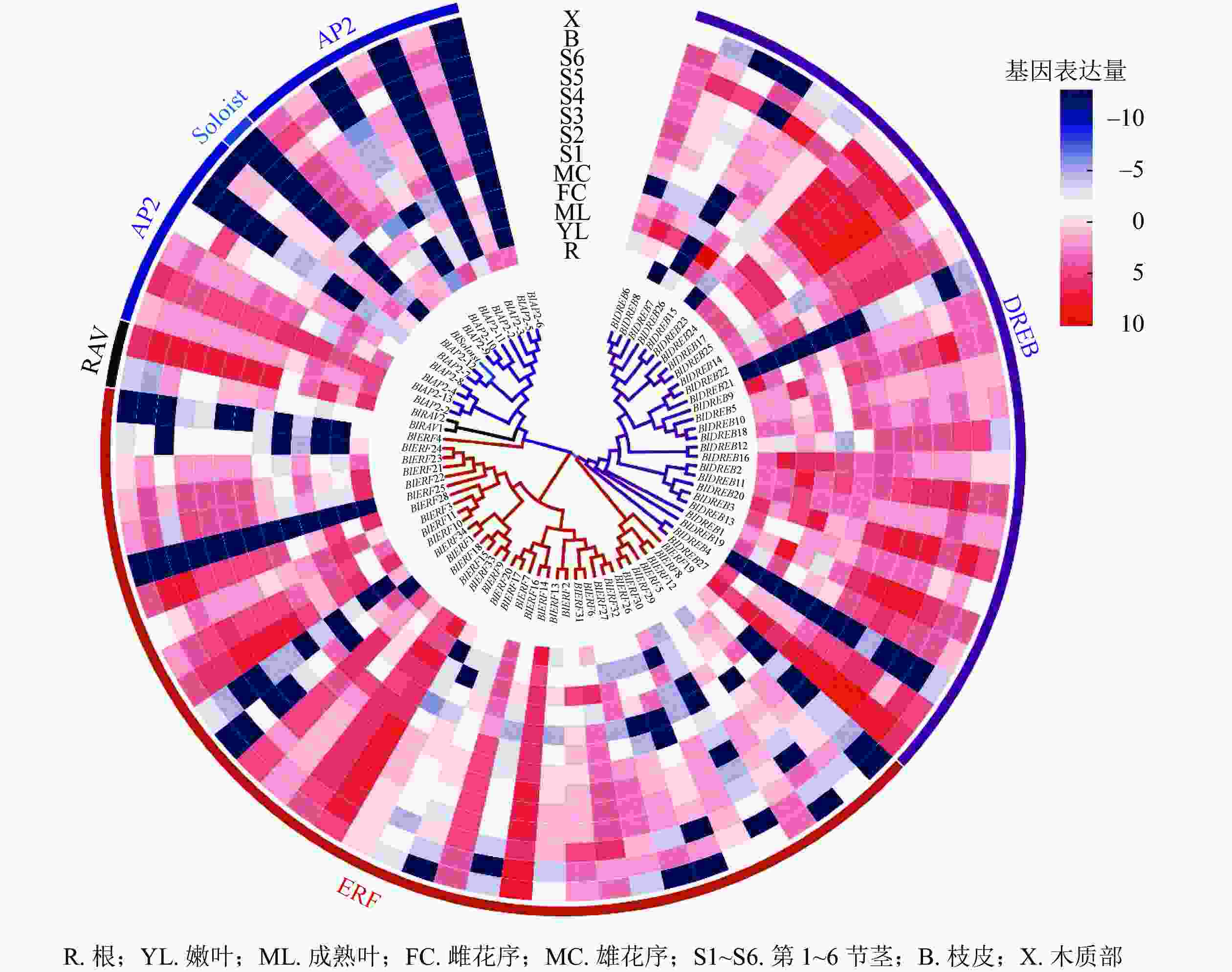
Figure 5. Expression profiles of AP2/ERF family genes in different tissues and organs of B. luminifera
-
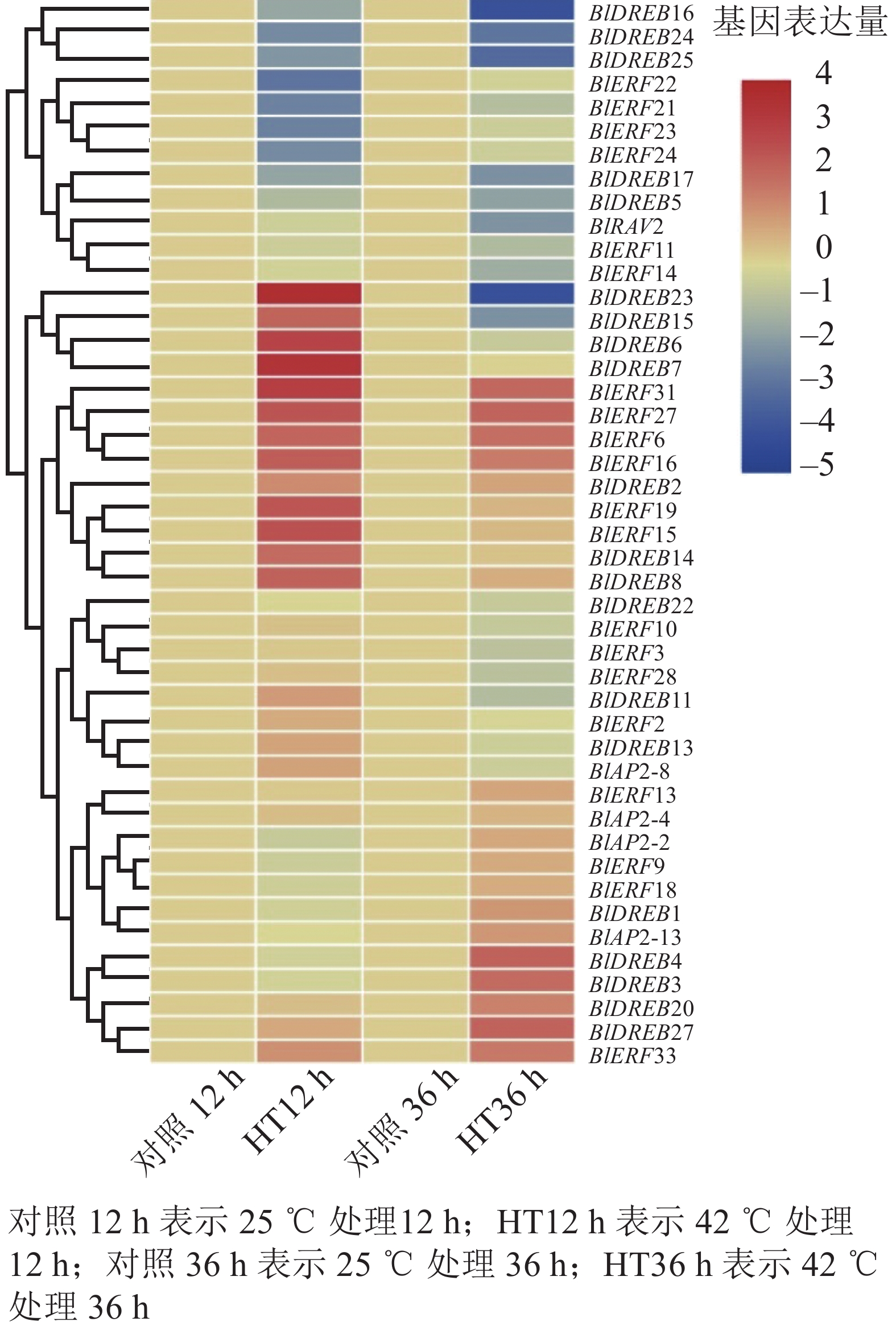
分析高温胁迫转录组数据,结果检测到45个基因在叶片中能够表达,包括4个AP2亚家族基因,20个DREB亚家族基因,20个ERF亚家族基因和1个RAV亚家族基因(图6)。进一步差异表达分析显示:多数ERF或DREB基因对高温胁迫产生了明显的响应,如BlDREB23在高温胁迫12 h后表达上调了12倍,BlERF6在高温胁迫12和36 h后分别被上调了3倍以上;而BlDREB24、BlDREB25和BlERF21等基因的表达则明显受到了高温胁迫的抑制(图6)。这些结果说明:ERF和DREB亚家族基因可能对光皮桦高温胁迫响应有重要的调控作用。
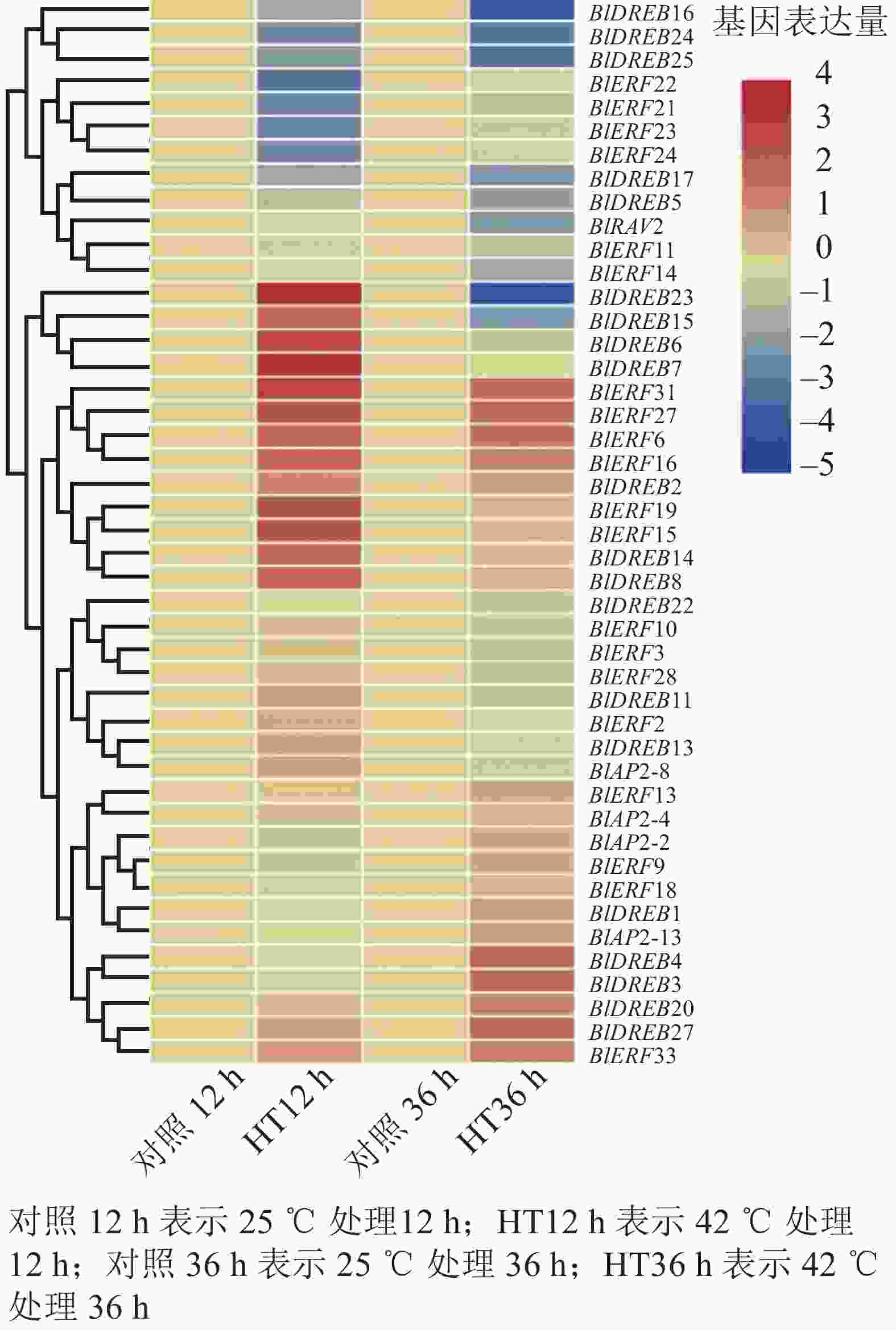
Figure 6. Expression analysis of AP2/ERF genes under heat stress in B. luminifera
-
AP2/ERF基因家族是植物最大的转录因子家族之一,在生长发育和逆境胁迫响应中扮演重要的角色[2, 22]。具有60~70个氨基酸的保守AP2结构域是该基因家族成员的重要特征。本研究在光皮桦基因组鉴定中获得了77个至少具有1个AP2结构域的AP2/ERF基因。与其他植物类似,这些光皮桦AP2/ERF基因也可被分为AP2、ERF、DREB、RAV和Soloist 等5个亚家族,包括13个AP2基因、27个DREB基因、34个ERF基因、2个RAV基因及1个Soloist基因。通常,在植物中ERF亚家族具有最多的成员,然后依次是DREB、AP2、RAV,Soloist亚家族成员最少。光皮桦与拟南芥[22]、水稻[22]、玉米[14]和杨树Populus trichocarpa[35]等物种的AP2/ERF基因各亚家族成员在数量上也具有相似的规律,这说明AP2/ERF基因在进化上可能有共同的起源。
转录因子的调控功能通常与一些重要的保守结构域或基序有关[36]。在AP2/ERF转录因子中,除了高度保守的AP2结构域,不同亚类的基因还具有特异的保守基序,如拟南芥AP2/ERF转录因子除了保守的AP2结构域,还具有50个保守基序[5],萝卜的AP2/ERF转录因子具有20个特异的保守基序[9]。光皮桦的AP2/ERF转录因子也具有7个特异的保守基序。进一步分析发现:同一亚家族基因具有相同或相似的保守基序,如绝大多数BlDREB基因具有特异基序Motif 10,同属ERF(B3)亚类的BlERF21~BlERF25具有1个特异基序Motif 8。在其他植物中,AP2/ERF基因的保守基序和结构域组成在亚家族内也具有类似的高度相似性,暗示同一亚家族基因可能具有相似的生物学功能和调控途径。此外,基因结构分析也表明:不同亚家族光皮桦AP2/ERF基因具有特异的基因结构特征,如AP2亚家族基因具有多个内含子(6~9个),而其他亚家族中仅少数基因具有单个内含子,其余基因则没有内含子。这种不同亚家族AP2/ERF基因具有的基因结构特征,在拟南芥、龙眼Dimocarpus longan、萝卜和玉米等不同植物中均存在类似现象,进一步说明该家族基因在进化上有共同的起源。
启动子顺式作用元件是基因功能的重要组分,能够反映基因潜在的功能和调控途径[11]。本研究显示:光皮桦AP2/ERF基因启动子区域的顺式作用元件主要可分为激素响应、胁迫响应和生长发育相关等3类,其中光响应元件、脱落酸响应元件和茉莉酸甲酯响应元件是最多的3种元件,这与美国山核桃Carya illinoinensis的AP2/ERF家族基因的顺式作用元件的分布模式类似[37]。有意思的是,不同AP2/ERF基因启动子的响应元件数量及类型不同,但不同亚家族基因启动子可能存在同类的响应元件,如脱落酸响应元件和茉莉酸甲酯响应元件在不同亚家族基因启动子中均存在。这说明不同亚家族基因在功能上产生了分化,但可能共同参与了相同的调控途径。同时,蛋白互作分析的结果显示:包括光皮桦在内的不同植物的AP2/ERF亚家族蛋白间存在广泛的互作关系[9, 11, 14, 38],进一步说明不同亚家族的AP2/ERF转录因子可能参与同一调控途径。
研究表明:不同AP2/ERF亚家族基因在植物生长发育和逆境胁迫响应中发挥不同作用。不同组织器官的表达分析显示:光皮桦AP2/ERF各家族成员在根、茎、叶等不同组织器官的表达存在差异,如BlAP2-6特异地在根中表达,而BlAP2-13、BlERF9、BlERF33、BlERF13和BlDREB27等5个基因则在所有组织器官中均有较高水平的表达。有意思的是,进化关系相近的基因,其表达模式也存在明显分化,如BlRAV1和BlRAV2的表达模式并不相同,而BlERF10和BlERF11、BlERF9和BlERF3的表达模式则非常相似。这说明,在这些进化关系较近的光皮桦AP2/ERF基因中既存在功能上的分化,也存在一定的功能冗余。已有研究表明:植物ERF和DREB基因主要在响应生物和非生物胁迫中发挥作用,它们的表达往往会受到逆境胁迫的诱导。如在萝卜中,RsERF003和RsERF039的表达明显受高温胁迫的诱导[9];玉米ZmERF135、水稻OsDREB2A和拟南芥AtDREB2A等基因的表达在被高温处理后明显上调[14, 39]。本研究的高温胁迫表达分析也表明:相比AP2亚家族基因,光皮桦ERF和DREB基因对高温胁迫更敏感,呈现出显著的表达差异,如BlDREB23、BlERF6等基因明显受到高温胁迫的诱导。这些结果显示:ERF和DREB基因在高温胁迫应答中发挥了重要作用。
在光皮桦77个AP2/ERF家族成员中,值得注意的是ERF(B3)亚类中的BlERF21~BlERF25分支。这5个基因具有1个特异基序Motif 8,且基因启动子区域中均含1~2个干旱诱导顺式作用元件,同时在高温42 ℃胁迫处理12 h后,除了BlERF25基因外,其他4个基因表达水平均明显下调,推测这一分支的5个基因可能在抵御高温及干旱胁迫中发挥着重要的作用,但具体基因功能仍需开展后续实验进行验证。
Identification and expression analysis of AP2/ERF gene family in Betula luminifera
doi: 10.11833/j.issn.2095-0756.20220331
- Received Date: 2022-04-29
- Accepted Date: 2022-07-17
- Rev Recd Date: 2022-07-04
- Available Online: 2022-11-21
- Publish Date: 2022-12-20
-
Key words:
- Betula luminifera /
- AP2/ERF family /
- expression pattern /
- heat stress
Abstract:
| Citation: | HUANG Yizi, QIAN Wang, QIU Shan, et al. Identification and expression analysis of AP2/ERF gene family in Betula luminifera[J]. Journal of Zhejiang A&F University, 2022, 39(6): 1183-1193. DOI: 10.11833/j.issn.2095-0756.20220331

|

 DownLoad:
DownLoad: