







-
闽楠Phoebe bournei被列为国家二级保护濒危树种[1−2],是“金丝楠木”的原植物种之一,天然分布于福建、浙江、江西、湖南、云南和广西等省[3]。闽楠木材纹理细致、结构紧密、香气浓郁、防虫耐腐、坚硬不易开裂,是制作高档家具、建筑和雕塑的优质木材。闽楠树体高大挺拔、冠型优美,现已成为中国南方地区珍贵造林与园林绿化树种。然而,干旱、低温等非生物胁迫显著影响闽楠的生长发育,极大限制了林分生产力。
转录因子作为一类与启动子顺式作用元件特异结合、调控功能基因表达的DNA结合蛋白,对植物多种生理生化过程起重要调控作用[4−5]。其中,碱性亮氨酸拉链(bZIP)转录因子是目前已知普遍存在、成员较多、功能复杂且高度保守的基因家族之一[6−7]。bZIP转录因子是按照其高度保守的结构域来命名的,保守结构域包括富含亮氨酸残基的拉链区域和N-X7-R/K碱性区域,由60~80个碱基组成,C端N-X7-R/K碱性区域由20个氨基酸构成,富含精氨酸和赖氨酸,可与DNA特异性结合;N端亮氨酸拉链区域富含亮氨酸,相邻7个氨基酸构成一段连续片段,且第7位碱基是亮氨酸,但亮氨酸拉链区域不完全保守,个别位点的亮氨酸可被甲硫氨酸、缬氨酸及异亮氨酸等疏水性氨基酸所取代[6]。
bZIP转录因子功能多样性,广泛参与调控脱落酸(ABA)、干旱、高盐及高温等多种非生物胁迫。例如,ABA、高盐和干旱胁迫下,ZmbZIP72在玉米Zea mays幼苗各器官上调表达,其异源过表达使拟南芥Arabidopsis thaliana显著改善叶片生理状态,通过减少失水和电解质渗漏来提高转基因株系的抗旱性和耐盐性[8]。高盐、干旱、高温、低温和ABA胁迫下,ZmbZIP4在玉米幼苗各器官被诱导表达,其异源过表达不仅增加拟南芥主根长和侧根数,还显著提高ABA合成,增强植株抵御非生物胁迫能力[9]。水稻Oryza sativa的OsbZIP42可正向调控ABA介导的信号通路,增强水稻过表达植株对干旱胁迫应答的敏感性[10]。bZIP还和ANAC096等转录因子互作,协同提高转基因植株抗旱性[11]。茶树Camellia sinensis的CsbZIP18是ABA信号和冷胁迫的负调控因子,异源过表达导致拟南芥对ABA信号敏感性下降、电解质渗漏升高以及光合效率下降,降低植株耐冻性[12]。bZIP还参与调控其他生物学过程,如组织器官发育[13−14]、盐胁迫[15]、激素和糖信号传递[16]、病虫害防御[17]、光反应和次生代谢物合成调控等[18−19]。
bZIP基因家族成员有进化多样性,成员数量在物种间存在显著差异,例如,拟南芥75个[20]、番茄Solanum lycopersicum 70个[21]、水稻89个[22]、玉米125个[23]、毛果杨Populus trichocarpa 214个[24],而闽楠bZIP成员数量不详,极大限制了闽楠bZIP基因功能研究。因此,本研究对闽楠bZIP转录因子家族成员进行全基因组鉴定,分析其蛋白理化性质,比较进化关系以及ABA处理下的表达水平,为解析bZIP家族成员的基因功能及其在逆境胁迫的调控机制提供理论基础。
-
从PFAM数据库获取包含保守结构域(PF00170、PF07716)的隐马尔科夫蛋白模型。运用HMMER软件在闽楠基因组[25]筛选bZIP同源序列。将初步获得的基因序列提交SMART数据库和美国国家生物技术信息中心(NCBI)网站(
https://www.ncbi.nlm.nih.gov/ )确认结构域,手动去除冗余,获得PbbZIP基因成员。利用Protparam数据库(https://www.expasy.org/ )进一步分析PbbZIP蛋白理化性质,通过植物亚细胞定位(Plant-mPLoc)数据库(http://www.csbio.sjtu.edu.cn )进行亚细胞定位分析,通过Prabi数据库(https://npsa-prabi.ibcp.fr/ )分析PbbZIP蛋白的二级结构。 -
运用Jaview软件对ClustalW输出的多序列比对结果进行美化,基因保守结构域用Tbtools[26]进行可视化,MEME suite数据库(
https://meme-suite.org/ )分析蛋白序列的保守基序,利用GSDS工具(http://gsds.gao-lab.org/ )在线绘制PbbZIP基因结构。 -
从Ensembl数据库(
http://plants.ensembl.org/species.html )获取拟南芥、番茄、毛果杨基因组数据,通过HMMER软件挑选出bZIP同源基因,构建最大似然进化树(Bootstrap: 1 000次)。通过Evolview网站(http://www.evolgenius.info/evolview )美化系统进化树。用TBtools的MCScanX工具分析PbbZIP基因的共线性关系。 -
在浙江农林大学苗圃地挑选长势健康的2年生闽楠半同胞家系WY8苗,选用2 mmol·L−1ABA均匀喷洒整株苗,使土壤表面湿润,吐温80 (Tween 80)为对照组(ck)。采用取样时间倒推法,分别在处理1、3、12、24、48和72 h取叶片和根,均在11:00取样,每个时间段包含15株闽楠苗,每5株混合作为1个生物学重复,3次重复,液氮速冻后储存于−80 ℃冰箱。
-
从闽楠基因组[25]提取PbbZIP基因编码区(CDS)上游2 000 bp序列,提交至PlantCARE数据库分析启动子顺式作用元件,统计与逆境胁迫相关的作用元件,对ABA响应元件较多的PbbZIP基因进行实时荧光定量,并分析其响应表达模式。
-
用CTAB法提取高质量总RNA,经质量分数为1%的琼脂糖凝胶电泳和NanoDrop检测合格后用于后续研究。每份RNA取1 μg,按照Vazyme公司提供的HiScript® II Reverse Transcriptase试剂盒反转录合成高质量cDNA,用ddH2O稀释5倍待用。使用NCBI 设计PbbZIP基因特异性引物用于实时荧光定量PCR (RT-qPCR)反应。使用RT-qPCR反应酶配成10.00 μL标准反应体系,包含1.00 μL cDNA、0.25 μL上下游引物、5.00 μL反应液和3.50 μL ddH2O。两步法RT-qPCR反应程序:95 ℃ 30 s,95 ℃ 10 s,60 ℃ 30 s,40个循环。闽楠ef1α基因(登录号:KX682032)作为内参基因,用2−ΔΔCt法计算各基因相对表达量[27],并通过SPSS进行方差分析及多重比较。
-
利用HMMER软件从闽楠基因组初步获得78条蛋白序列,经SMART和NCBI数据库验证结构域完整性,最终鉴定出63个PbbZIP成员,按染色体位置命名为PbbZIP01~PbbZIP63,并分析其蛋白特征和理化性质。PbbZIP蛋白长度介于110 (PbbZIP60)~835 (PbbZIP12)个氨基酸,相对分子量为13.05~88.65 kDa,平均为39.76 kDa。蛋白等电点为4.48 (PbbZIP02)~11.95 (PbbZIP32),疏水性为−1.19 (PbbZIP58)~−0.19 (PbbZIP16)。亚细胞定位分析结果表明:59个成员定位于细胞核,而PbbZIP02定位于细胞质,PbbZIP05和PbbZIP12定位于内质网,PbbZIP16定位于线粒体。
-
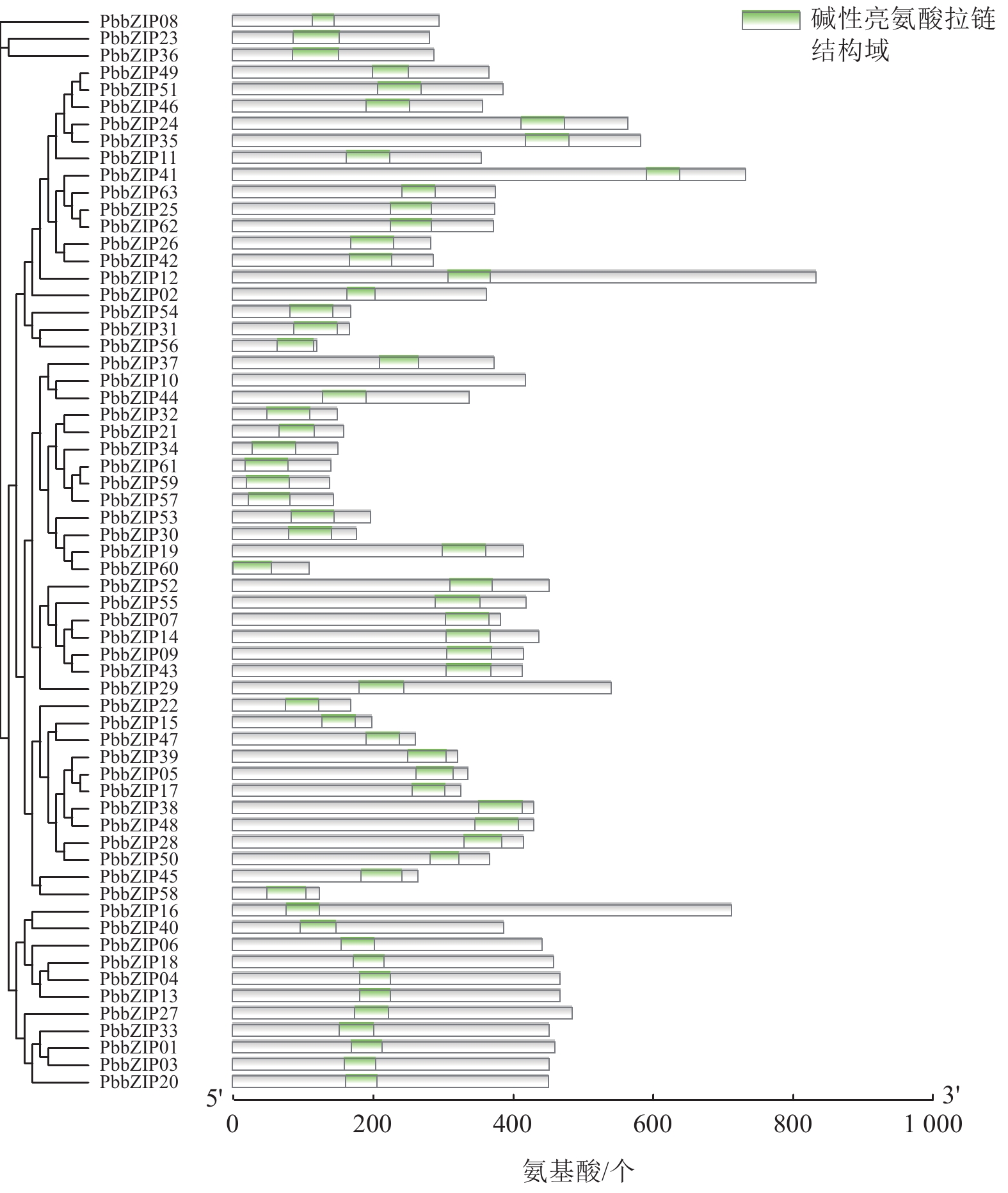
Motif分析结果表明:PbbZIP基因保守基序较丰富,且不同亚族差异明显。Motif 5和Motif 1在PbbZIP基因家族广泛存在(图1),分别为碱性结构区域和亮氨酸拉链结构区域,碱性区域可以和DNA特异性结合,发挥调控活性(图2)。A亚族特有Motif 6、Motif 10和Motif 13;C亚族和S亚族特有Motif 11;D亚族的保守基序较丰富,特有基序包括Motif 2、Motif 3、Motif 4、Motif 7、Motif 9和Motif 12;F亚族特有Motif 15;I亚族特有Motif 6;G亚家族特有Motif 8和Motif 14。其中,A、E、F、G、J亚族含有重复基序。
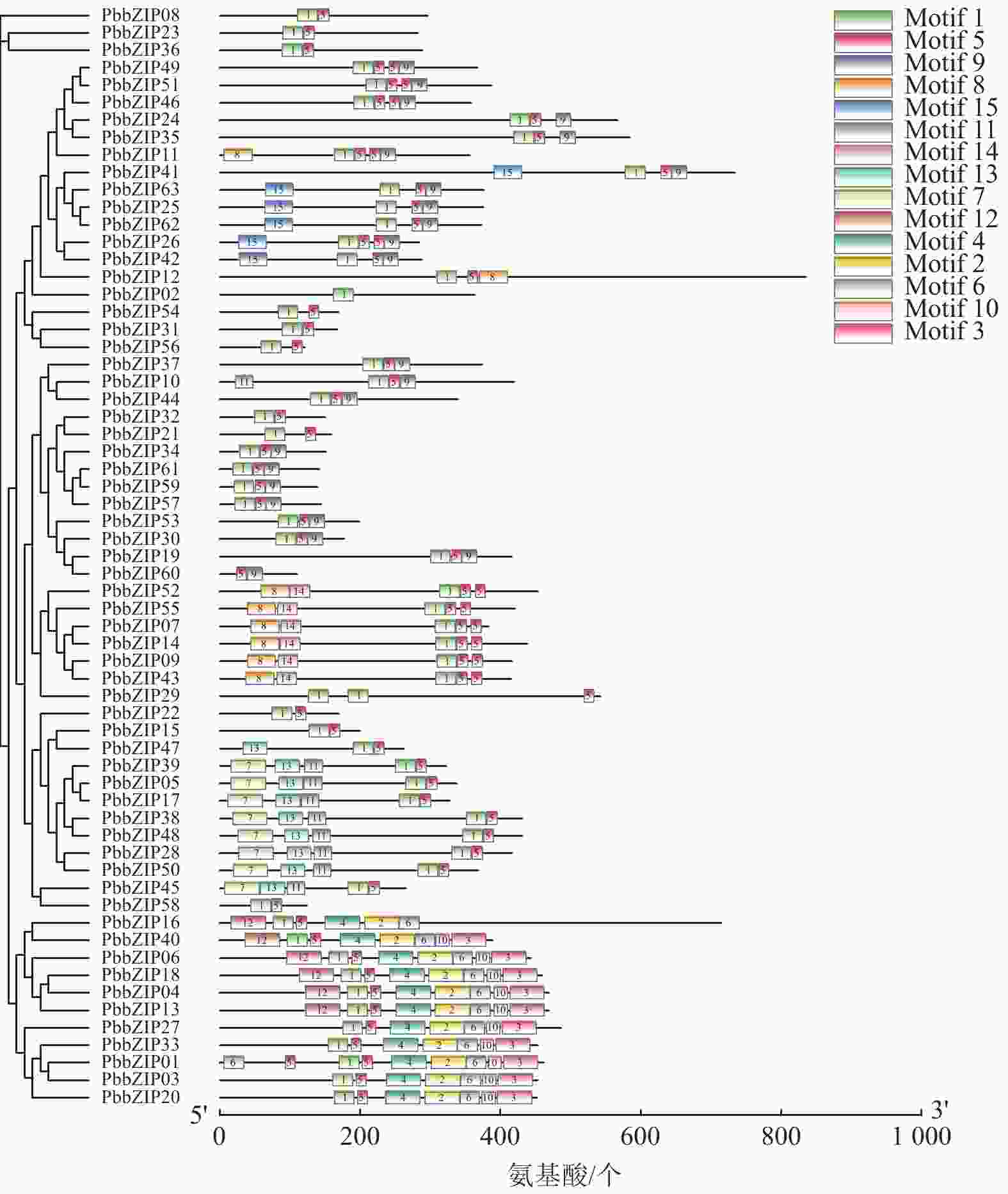
Figure 1. Conserved motifs of PbbZIP family members
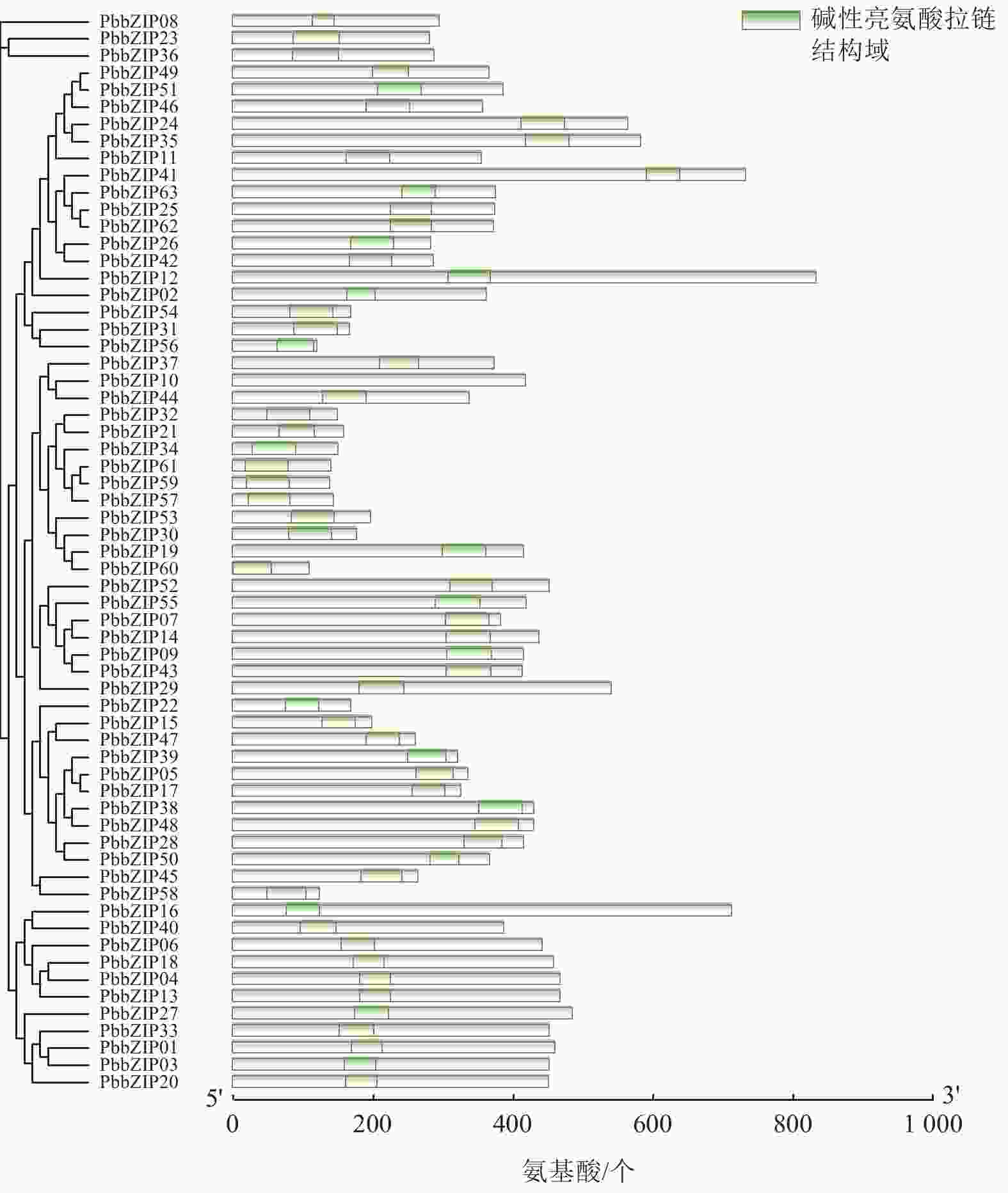
Figure 2. Domains of PbbZIP family members
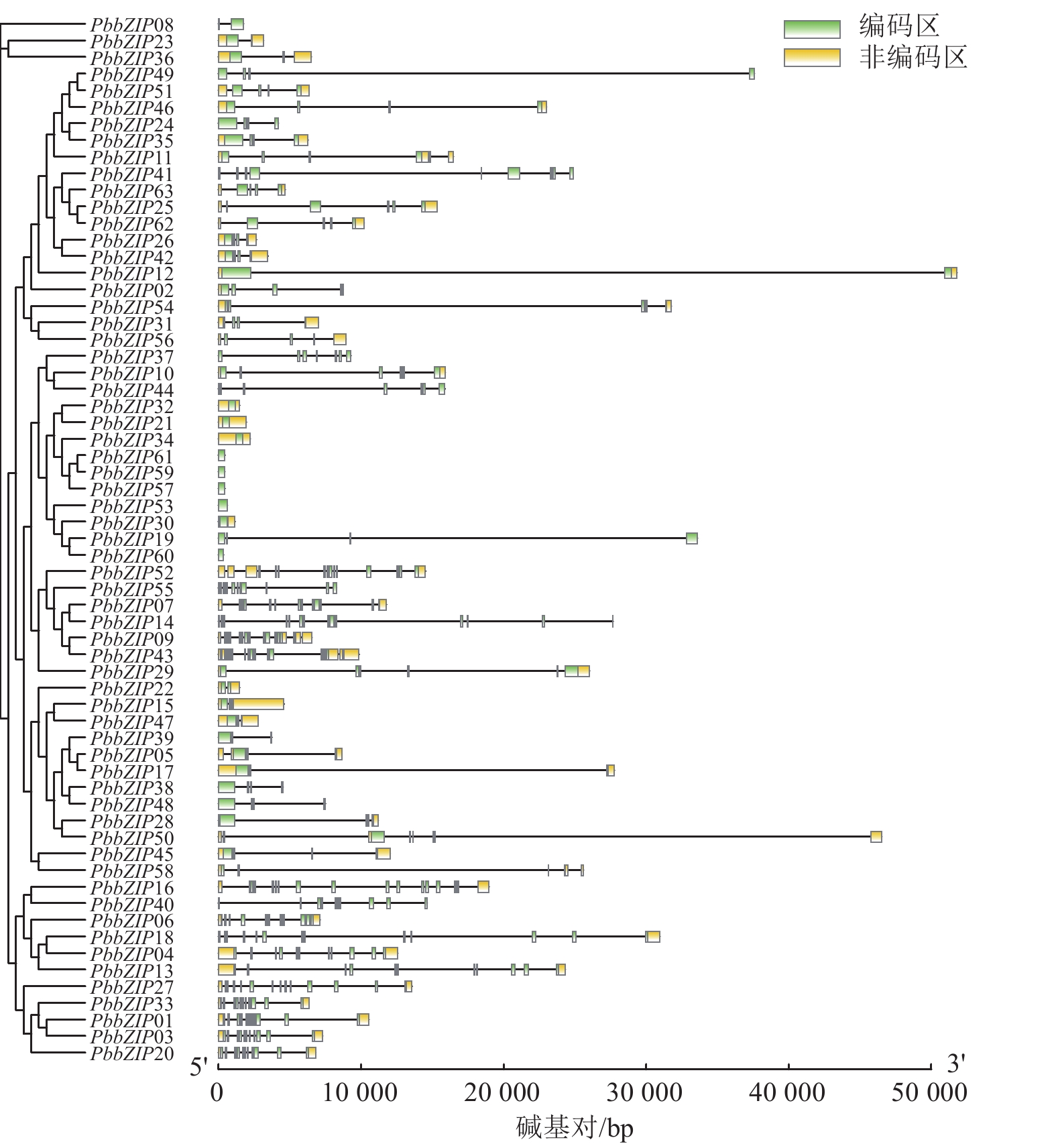
PbbZIP家族的基因结构在内含子、外显子的数量和位置存在显著多样性和差异性(图3)。例如,S亚族外显子数量最少(1~4个),且多数无内含子,这与拟南芥bZIP-S亚族结构相似;A亚族含有3~4个外显子,C亚族含有6~7个外显子,D亚族含有11~12个外显子;E亚族含有5~6个外显子,F亚族含有2~3个外显子,而G亚族内含子和外显子最多,为13~14个。
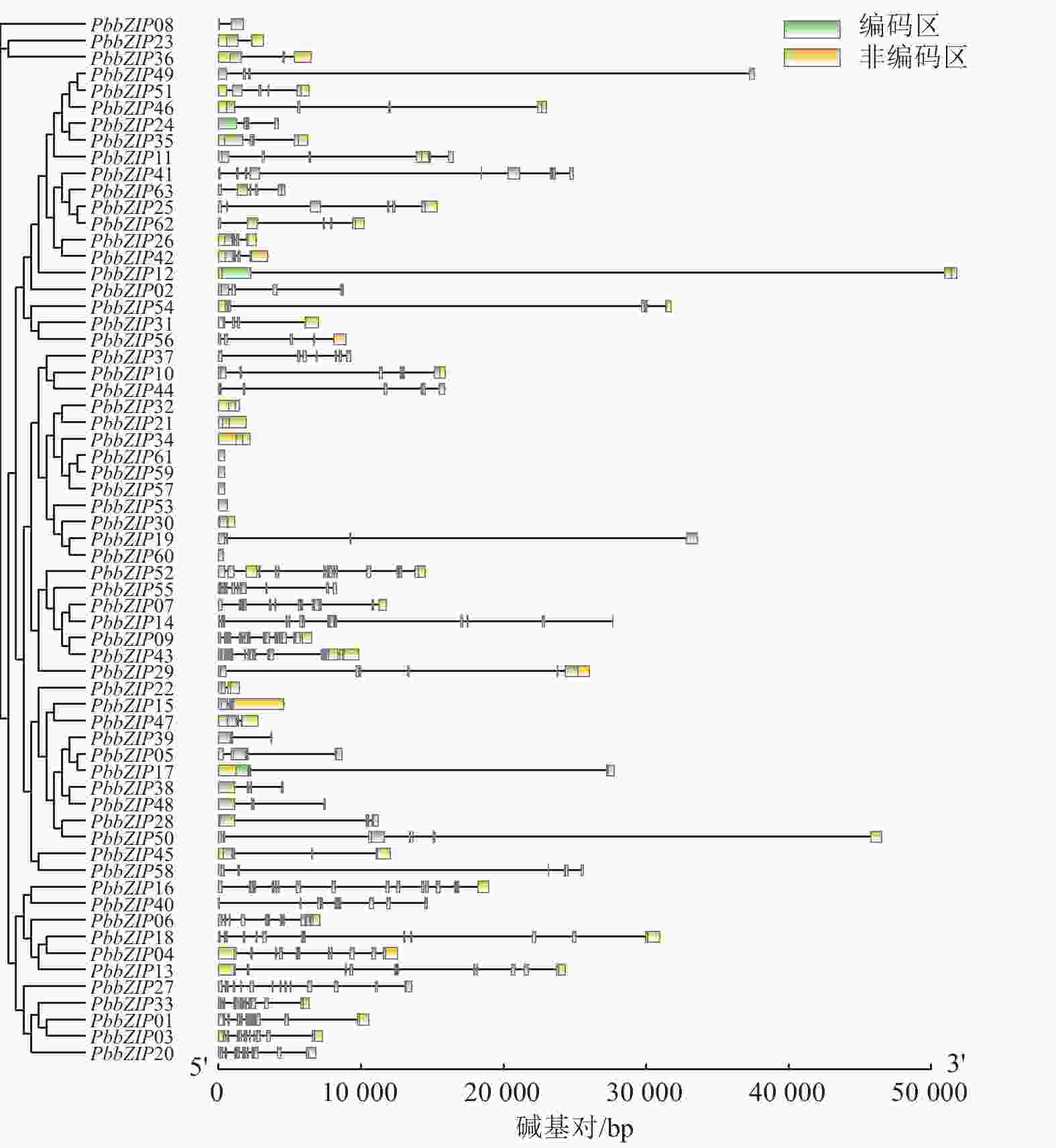
Figure 3. Gene structures of PbbZIP family members
-
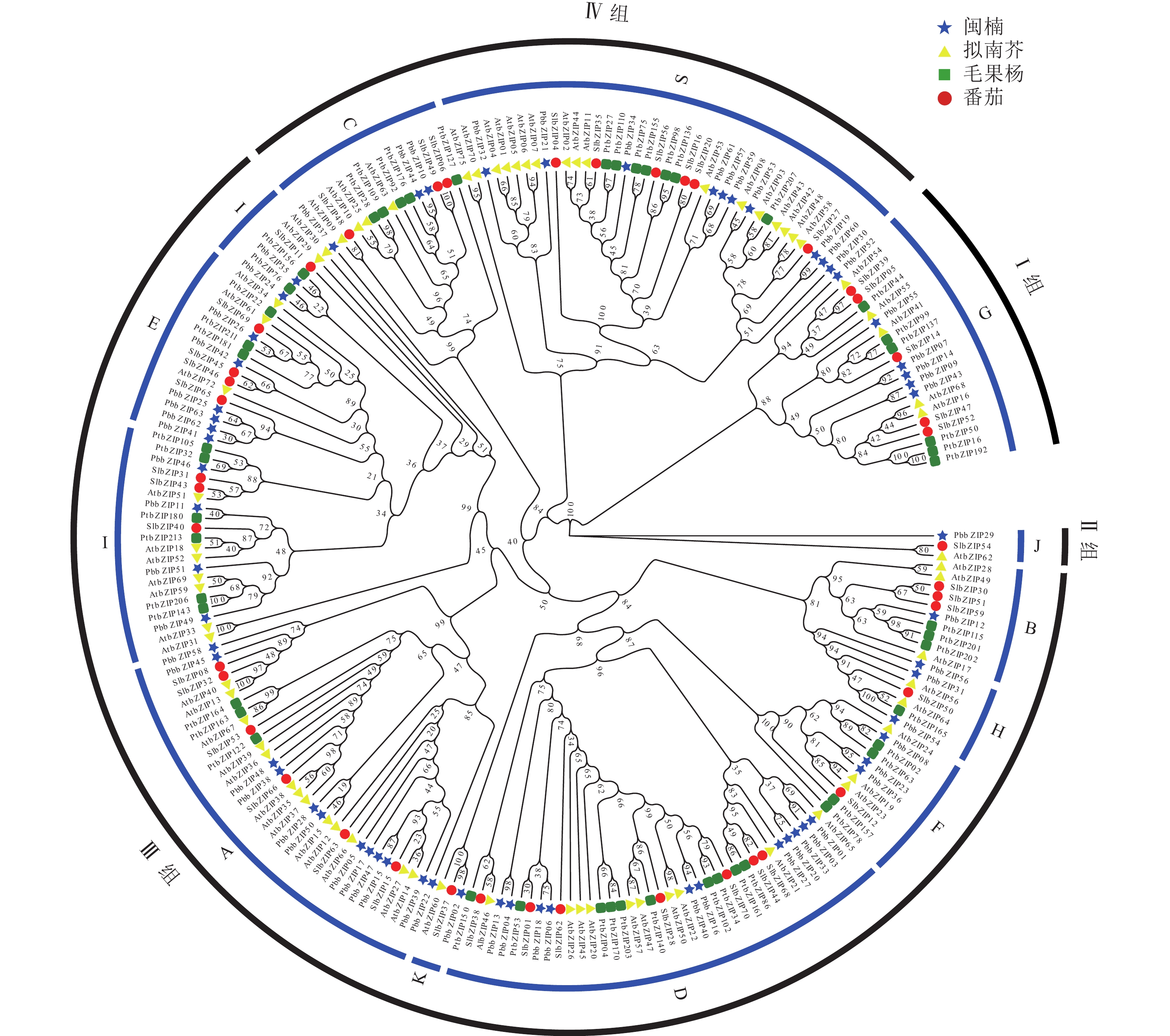
为研究bZIP基因的进化关系,选取拟南芥、番茄、毛果杨bZIP氨基酸序列与PbbZIP基因构建最大似然进化树(图4)。系统进化分析表明:63个PbbZIPs基因按序列同源性划分为4组12亚族,Ⅰ 组(G)、Ⅱ 组(J)、Ⅲ 组(B、H、E、I、F、K、A、D)和Ⅳ 组(C、S),其中A亚族12个、B亚族1个、C亚族3个、D亚族11个、E亚族5个、F亚族3个、G亚族6个、H亚族4个、I亚族6个、J亚族1个、K亚族1个和S亚族10个。
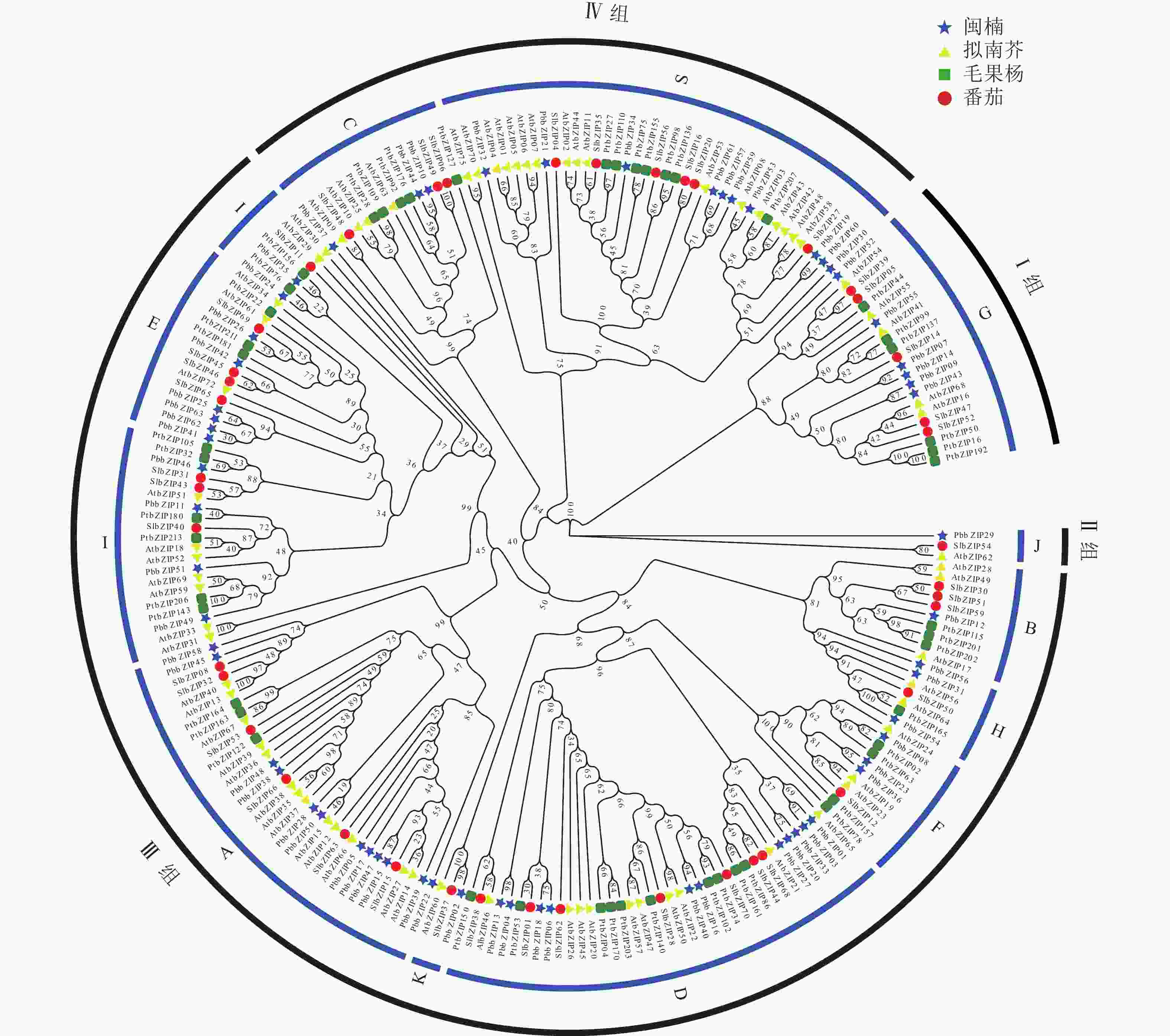
Figure 4. Phylogenetic analysis of bZIP family from P. bournei, P. trichocarpa, A. thaliana, and S. lycopersicum
-
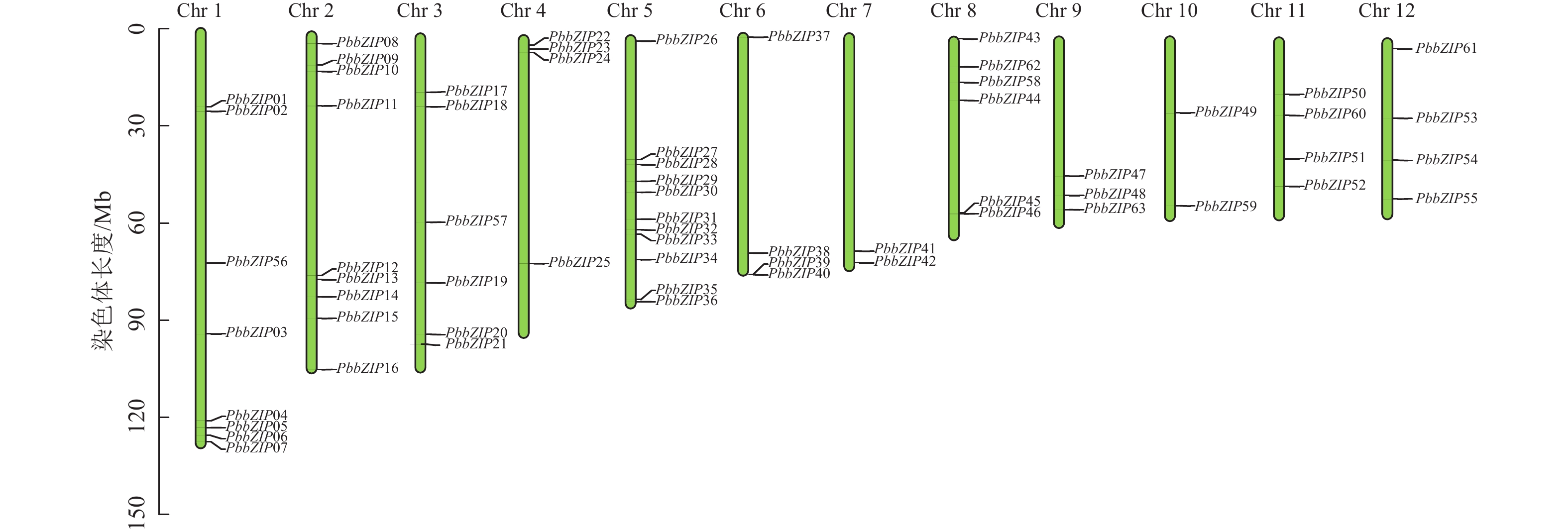
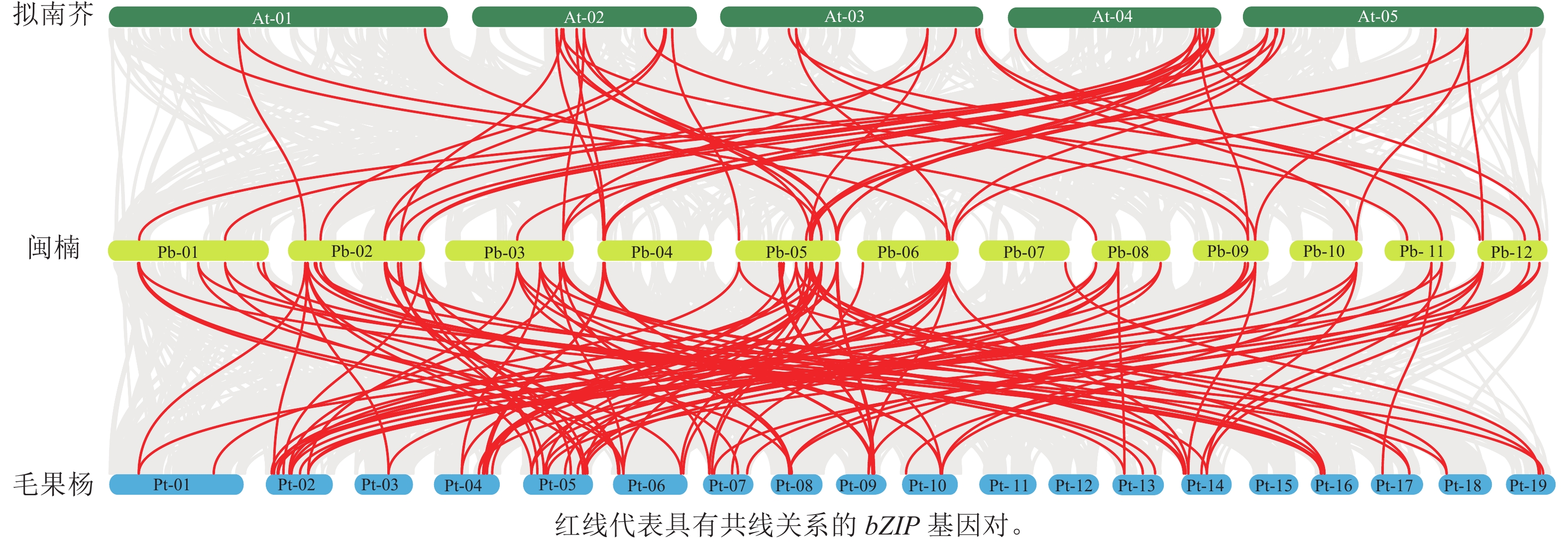
染色体定位结果表明:63个PbbZIPs基因不均匀分布于12条染色体(图5),其中Chr 5基因数量最多,含11个PbbZIPs成员,占17.46%,而Chr 7和Chr 10最少,仅有2个PbbZIPs成员。为深入了解PbbZIP基因家族的扩张模式,对63个PbbZIPs基因进行共线性分析,结果表明:分布于12条染色体上的27对PbbZIPs基因存在片段复制,其中D亚族共线基因对最多(10对),未检测到串联复制,表明片段复制是PbbZIP基因家族扩张的主要模式。不同物种间共线性结果表明(图6):闽楠与拟南芥47个、毛果杨110个bZIPs基因对存在共线性关系,闽楠和毛果杨bZIP基因更保守,亲缘关系较近。
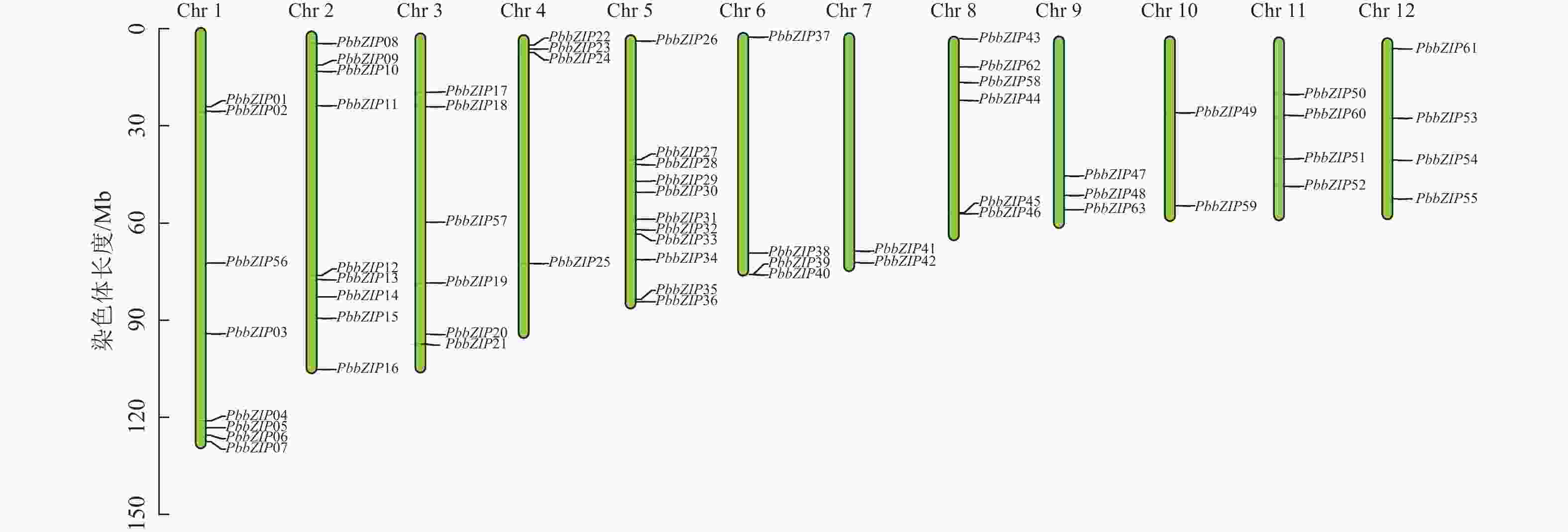
Figure 5. Chromosomal localization of PbbZIP family members

Figure 6. Interspecific collinear relationships of PbbZIP family members
-
PbbZIP基因CDS区上游启动子顺式作用元件分析结果表明(图7):共发现18种(745个)与胁迫相关的顺式作用元件,其中激素响应元件428个,包括ABA响应元件(ABRE) 129个,茉莉酸甲酯响应元件(TGACG-motif、CGTCA-motif) 146个,赤霉素响应元件(P-box、GARE-motif、TATC-box) 71个,水杨酸响应元件(SARE、TCA-element) 54个,生长素响应元件(TGA-element、TGA-box、AuxRR-core) 28个。此外,还发现多种非生物胁迫响应元件,如低温响应元件(LTR、DRE) 45个,干旱响应元件(MBS) 53个,缺氧厌氧元件(ARE、GC-motif) 182个,防御和胁迫响应元件(TC-rich repeats) 36个,伤害响应元件(WUN-motif) 1个。
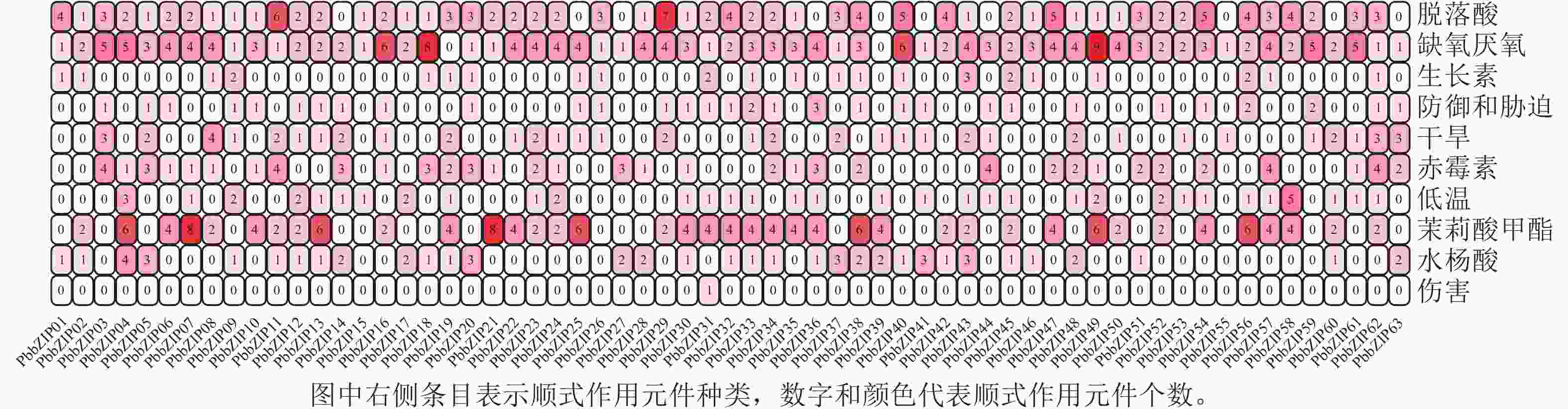
Figure 7. Cis-acting elements associated with adversity stress in promoters of PbbZIPs
-
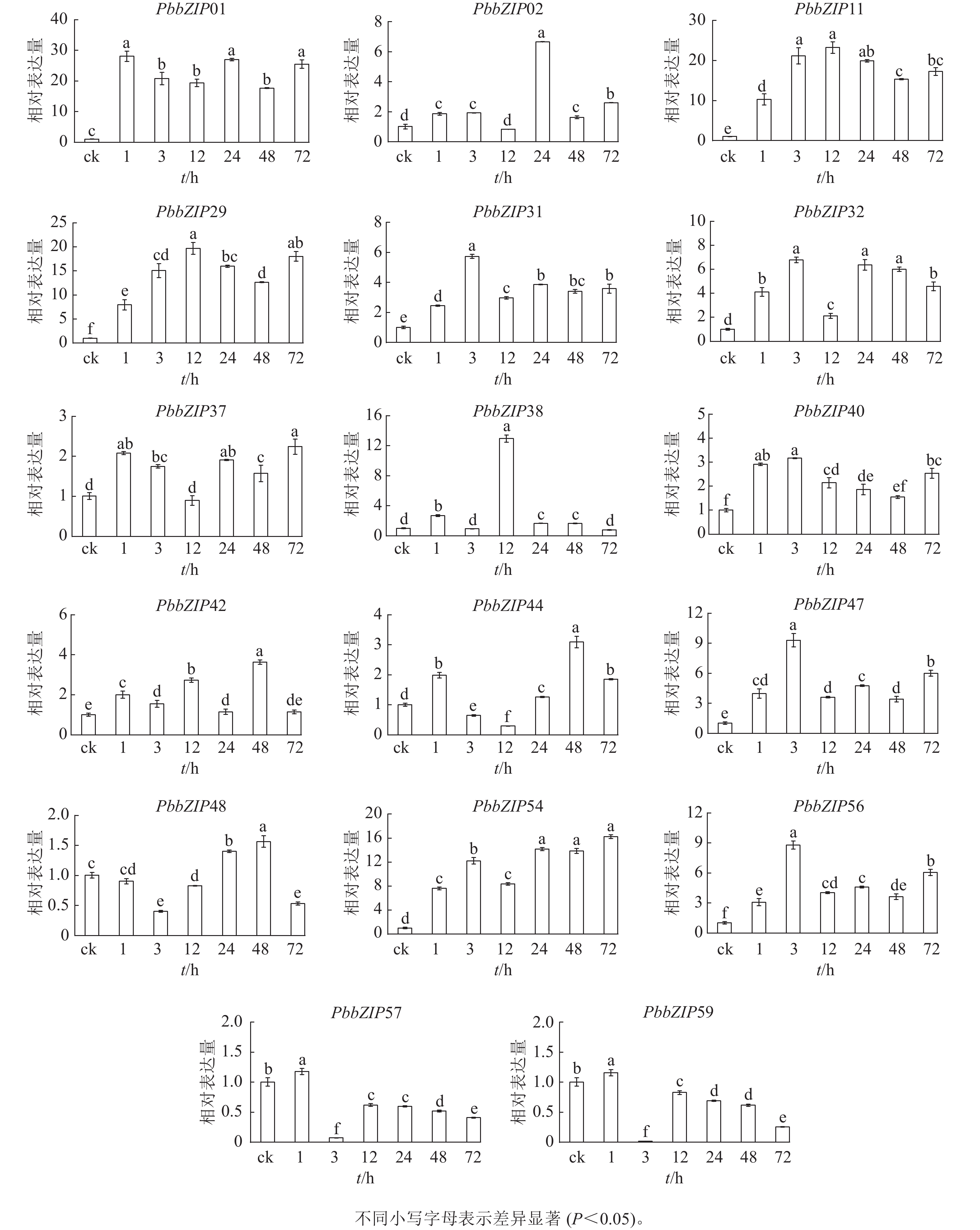
PbbZIP基因启动子区域富含ABA响应元件,挑选含较多ABA响应元件的17个PbbZIPs成员进行ABA处理下的表达模式分析。首先,应用RT-qPCR分析ABA处理后叶中17个PbbZIPs的响应程度,表明叶中17个PbbZIPs对ABA信号有着不同程度的诱导响应(图8)。例如,PbbZIP01、PbbZIP11、PbbZIP29和PbbZIP54等4个基因在叶中不同处理时间均被显著诱导(P<0.05),相对表达倍数高于8倍;PbbZIP31、PbbZIP32、PbbZIP37、PbbZIP40、PbbZIP42、PbbZIP47和PbbZIP56等7个成员均被ABA诱导表达,但表达倍数增加幅度不高;PbbZIP44、PbbZIP48、PbbZIP57和PbbZIP59在ABA处理3 h或12 h时被显著抑制表达(P<0.05);PbbZIP02和PbbZIP38分别仅在ABA处理24 h和12 h强烈诱导表达,而其他处理时间段基本不被诱导。
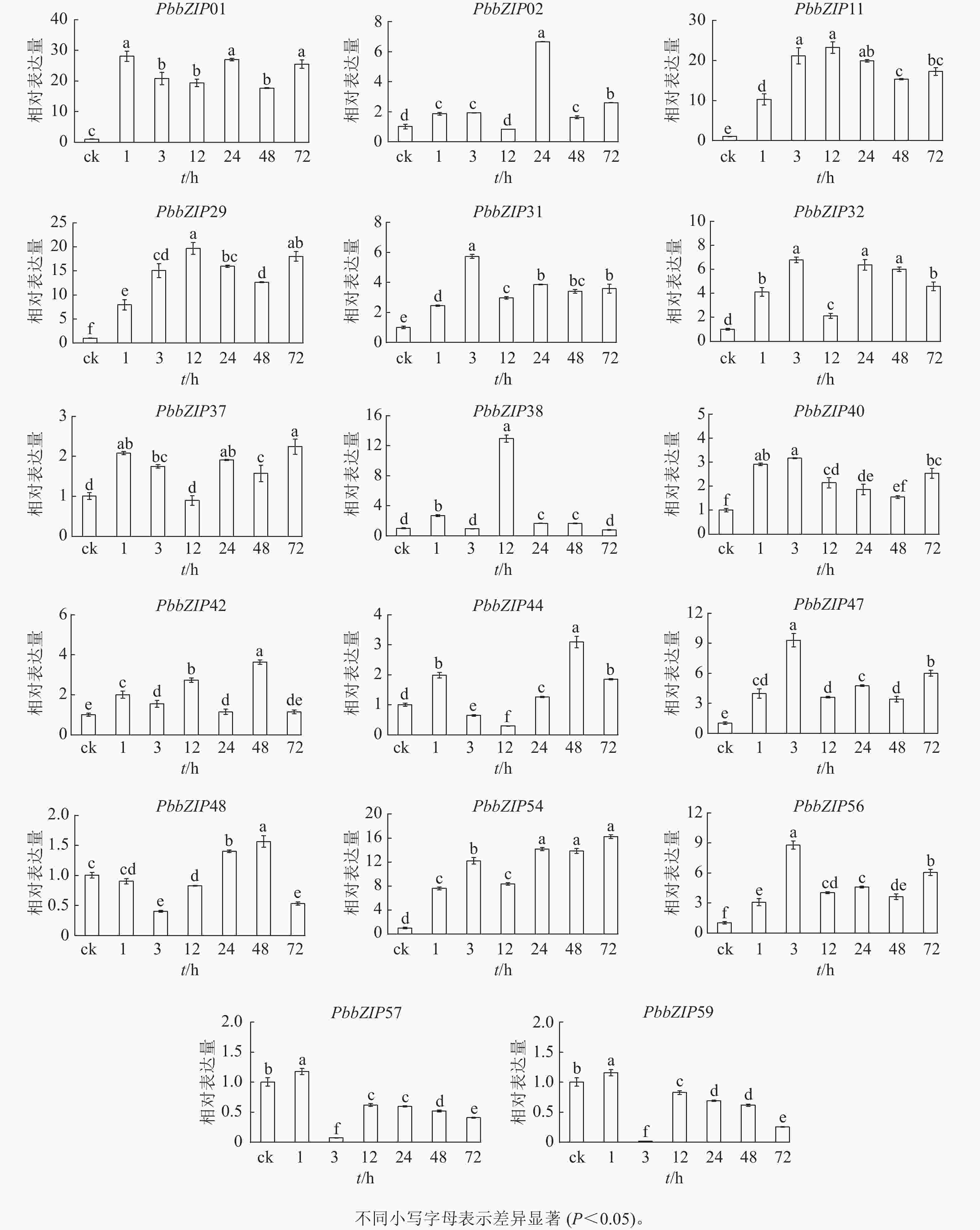
Figure 8. Response of PbbZIP genes to ABA treatment in leaves
-
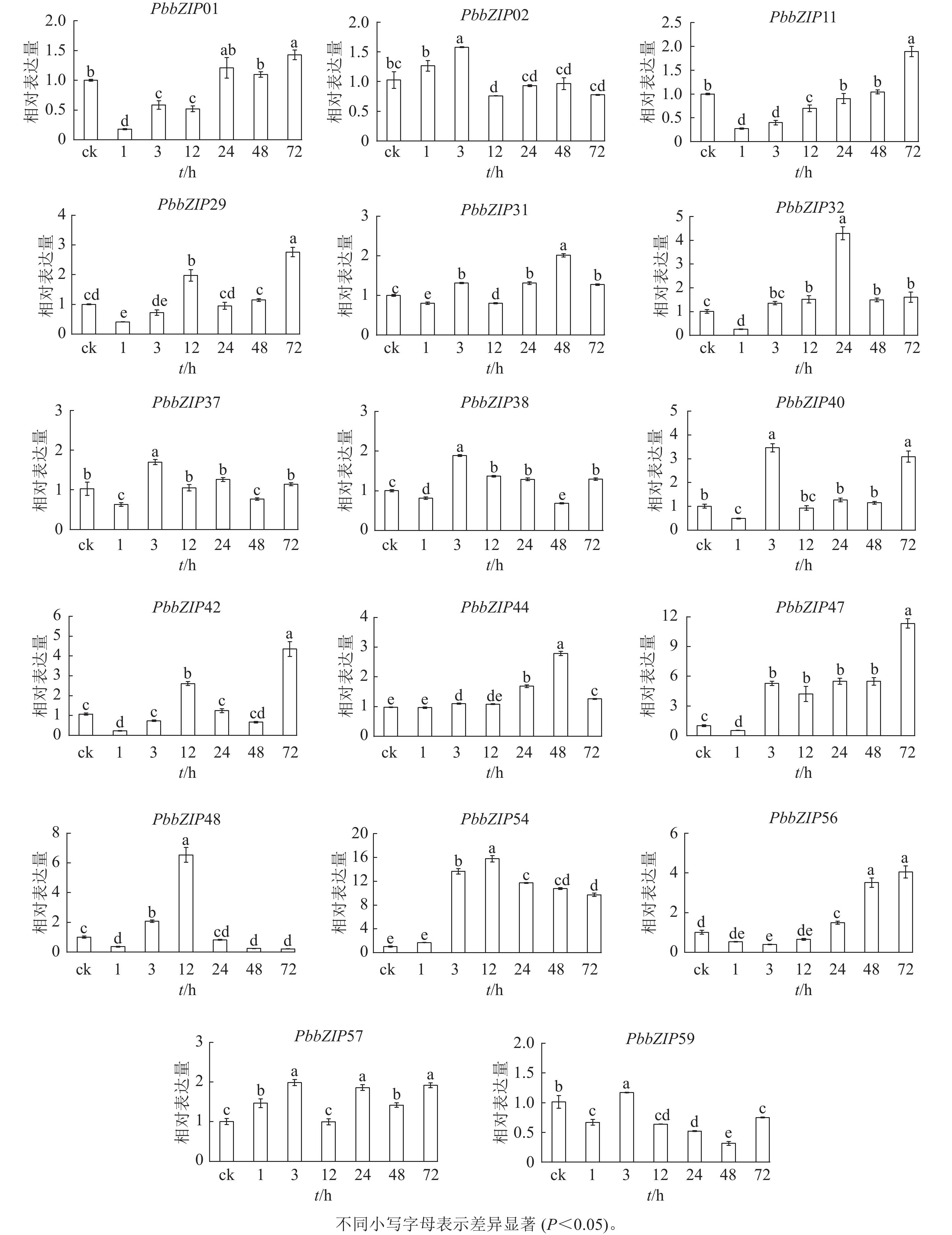
ABA处理1~72 h,17个PbbZIP基因在根中表现出不同趋势,且表达倍数较叶片低(图9)。PbbZIP01、PbbZIP11、PbbZIP29、PbbZIP31、PbbZIP32、PbbZIP37、PbbZIP38、PbbZIP40、PbbZIP42、PbbZIP47、PbbZIP48、PbbZIP56和PbbZIP59等表达水平先降低后升高,其中PbbZIP32、PbbZIP37、PbbZIP38、PbbZIP40、PbbZIP48和PbbZIP59等基因的表达量在ABA处理48 h后降低,而PbbZIP37、PbbZIP38、PbbZIP40和PbbZIP59表达水平在ABA处理72 h后显著升高(P<0.05)。PbbZIP02、PbbZIP44和PbbZIP57表达水平受ABA影响较小;PbbZIP54在ABA处理3 h后显著诱导表达(P<0.05),且保持较高水平,可能参与ABA信号响应发挥重要调控功能。
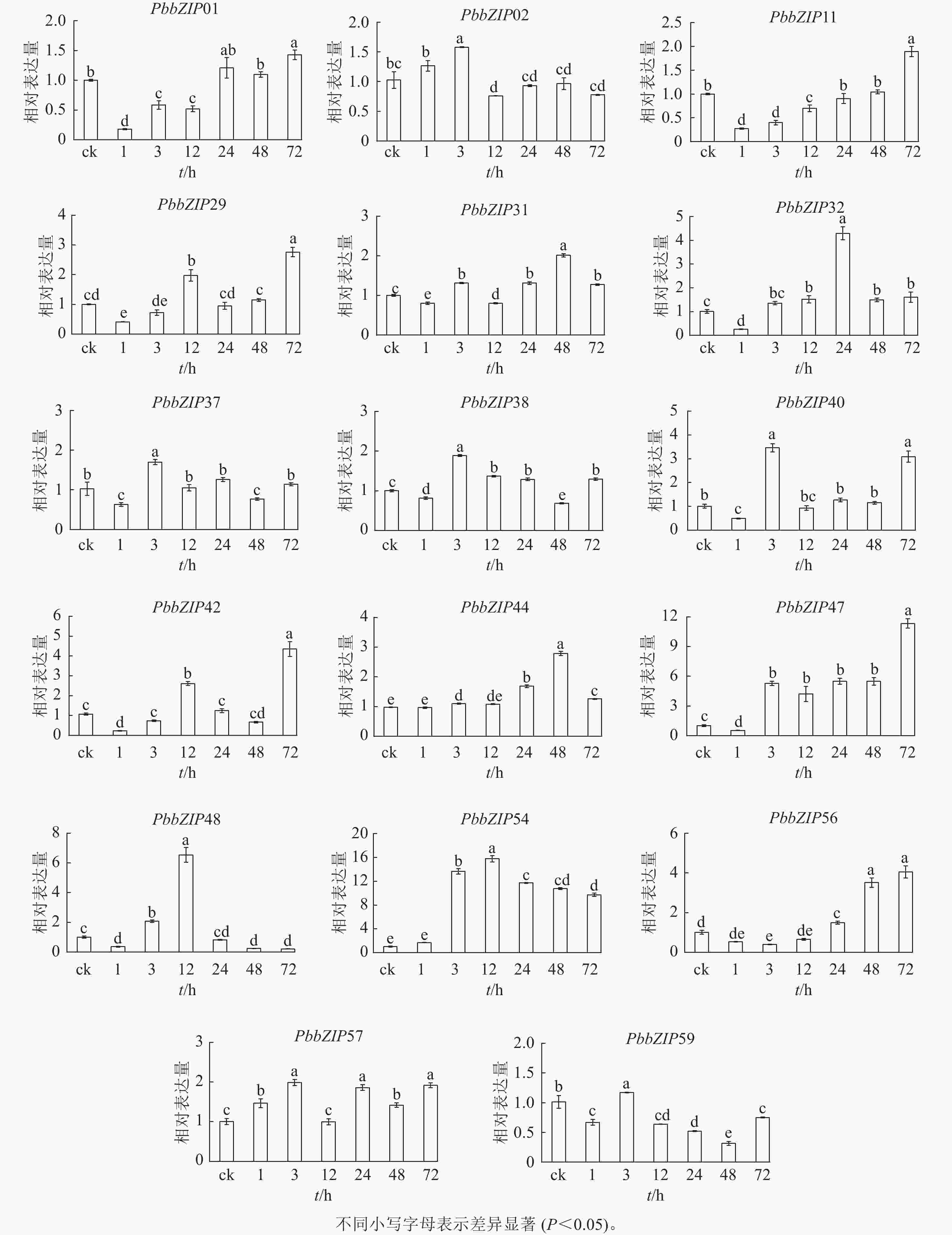
Figure 9. Response of PbbZIP genes to ABA treatment in roots
-
bZIP基因家族是成员和功能最复杂的转录因子家族之一,参与多种代谢过程,目前功能研究集中于拟南芥、毛果杨、水稻和番茄等物种。本研究从闽楠基因组鉴定出63个PbbZIP转录因子,该家族成员在蛋白理化性质、基因结构和保守结构域等方面与已报道物种(拟南芥[20]、毛果杨[24]、番茄[21]等)较相似,说明bZIP基因结构较保守。进一步将PbbZIP与拟南芥、番茄和毛果杨构建最大似然进化树,发现63个PbbZIP基因可进一步划分为12亚族,可能由4个以上不同功能的原始bZIP基因进化扩张产生[28],而在其他物种中,bZIP转录因子被划分为不同亚族,如拟南芥10个[20]、毛果杨12个[29]、烟草Nicotiana tabacum 11个[30]。其中PbbZIP29基因结构高度分化,单独聚类在一个进化分支上,其功能是否分化有待后续深入研究。
通过启动子顺式作用元件分析发现:PbbZIP基因CDS上游启动子区域存在多种响应元件,推测PbbZIP基因可能受生物及非生物胁迫诱导表达。例如ABA、茉莉酸甲酯和生长素等激素响应元件在启动子区域数量较多,推测PbbZIP基因可能受到激素诱导。低温、干旱和缺氧厌氧诱导等非生物胁迫响应元件数目也较多,推测PbbZIP表达可能受非生物胁迫诱导,正向或负向调控次生代谢途径而参与植物生长发育进程。例如,猕猴桃Actinidia deliciosa bZIP12正向调控ABA介导的AchnKCS酶基因,促进果实创伤愈合,免受病原体侵害[31]。在ABA、干旱、盐胁迫下,辣椒Capsicum annuum叶片bZIP-D亚族中的CaDILZ1显著上调,它通过调节ABA含量、气孔开合度及抗旱基因表达水平来增强转基因植株的耐旱性[32]。
17个PbbZIP基因在叶和根有不同程度的诱导表达,随ABA诱导表现出不同表达趋势。处理1~72 h,叶中PbbZIP11等基因和根中PbbZIP44等基因表达量先升高后降低,而叶中PbbZIP37等基因以及根中PbbZIP56基因表达趋势与之相反,先下降后上升,烟草bZIP-A亚族基因经1 μmol·L−1ABA处理24 h后也有相同趋势[33];武立伟等[34]研究发现:25和50 mg·L−1 ABA处理甘草Glycyrrhiza uralensis根,GubZIP1~GubZIP56响应趋势与本研究结果相似,表明PbbZIP基因可能参与ABA等非生物胁迫的响应;孙晓丽等[35]研究发现:拟南芥AtbZIP1与ABRE元件结合负向调节植株对ABA敏感性,AtbZIP1缺失提高了种子萌发率,增加了主根长和ABA响应基因的表达,推测PbbZIP基因可能有相似功能。叶中PbbZIP01、PbbZIP29、PbbZIP32、PbbZIP38和PbbZIP54等被ABA显著诱导,根中PbbZIP47和PbbZIP54被显著诱导,表达量较高,后续需进一步开展功能验证,阐明其生物学功能与调控机制。
-
本研究通过对闽楠bZIP转录因子家族鉴定和表达分析,发现63个基因家族成员高度保守,不同亚家族在蛋白理化性质、基因结构、染色体定位、保守基序等方面表现出进化多样性和差异性。PbbZIP基因的表达随ABA诱导时间增加呈非同步变化的趋势,具有一定组织特异性,叶响应程度普遍高于根,这种表达特征与闽楠胁迫耐受性高度关联,存在复杂的调控网络,需进一步探索PbbZIP基因功能。
Identification and expression analysis of bZIP gene family under ABA treatment in Phoebe bournei
doi: 10.11833/j.issn.2095-0756.20230342
- Received Date: 2023-06-01
- Accepted Date: 2023-12-12
- Rev Recd Date: 2023-10-17
- Available Online: 2024-01-24
- Publish Date: 2024-04-01
-
Key words:
- Phoebe bournei /
- bZIP family /
- systematic evolution /
- ABA /
- expression analysis
Abstract:
| Citation: | YANG Yong, ZHANG Junhong, HAN Xiao, et al. Identification and expression analysis of bZIP gene family under ABA treatment in Phoebe bournei[J]. Journal of Zhejiang A&F University, 2024, 41(2): 275-285. DOI: 10.11833/j.issn.2095-0756.20230342

|
 DownLoad:
DownLoad: