







-
生态公益林在水土保持、涵养水源、碳汇和生物多样性保护等方面具有重要作用。截至2018年,中国人工生态公益林面积已达2 649万hm2,其中南方林区的人工生态公益林占比约1/3[1]。马尾松Pinus massoniana和桉树Eucalyptus spp.是中国南方地区主要的速生丰产造林树种。截至2018年,马尾松和桉树人工林面积分别为252万hm2和547万hm2,分别占全国森林面积的1.15%和2.49%[1]。随着林业战略转变,部分近熟马尾松和桉树人工林由用材林转为生态公益林,营林方式也从短轮伐、多代连栽转为封山育林。然而,用材林的不当经营导致了病虫害暴发、生物多样性丧失及土壤退化等问题[2−4]。因此,人工生态公益林在转为封山育林后生态功能的恢复与提升,成为亟待解决的问题。目前,关于马尾松和桉树人工公益林生态功能的研究主要集中在碳汇、土壤养分、水源涵养和动植物多样性等方面[5−7]。土壤微生物作为连接地上与地下生态过程的桥梁,在维持生物地球化学循环、森林生物多样性以及生态系统功能中发挥着至关重要的作用[8]。同时,土壤微生物对环境变化反应敏感,是评估土壤质量和生态系统功能的重要指标[9]。然而,关于马尾松和桉树人工生态公益林土壤微生物多样性的研究尚未见报道,现有研究主要关注人工用材林的土壤微生物多样性,尤其是不同营林措施的影响[10−11]。部分研究通过对比桉树用材林与天然林的土壤微生物多样性评估人工林的多样性,但结论不一[12−13],而针对马尾松人工林与天然林土壤微生物多样性的对比研究仍较为匮乏。
为探讨马尾松和桉树人工生态公益林土壤细菌与真菌多样性及其影响因素,本研究以广西雅长兰科植物国家级自然保护区的2种地带性天然林(包括喀斯特林、土山天然林,均为公益林)和2种人工生态公益林(包括马尾松、尾叶桉E. urophylla人工林)为研究对象,比较天然林与人工林的土壤微生物多样性,评估人工林的土壤细菌和真菌多样性状况,并结合植物群落和土壤理化性质,分析微生物群落组成与多样性差异的影响机制,为人工生态公益林的科学经营及生物多样性保护提供依据。
-
研究区位于广西雅长兰科植物国家级自然保护区(24°44′16′′~24°53′58′′N,106°11′31′′~106°27′04′′E),属南亚热带季风气候,年平均气温为16.8 ℃,年平均降水量为1 058.0 mm。研究区位于云贵高原东南边缘,喀斯特与非喀斯特地貌交织。非喀斯特地貌以褐红土、红壤和黄壤为主,喀斯特地貌则以石灰土为主。研究区的天然林主要为由细叶云南松Pinus yunnanensis var. tenuifolia原始林被严重破坏后自我恢复形成的天然次生林[14]。
-
2022年7—9月,在保护区内设置喀斯特林(38 a)和土山天然林(40 a)样地,保护区外围设置马尾松人工林(20年生)和尾叶桉人工林(14年生)样地。每个林型内设置5个20 m × 20 m的样方,样方间距至少500 m,每个样方进一步划分为4个10 m × 10 m的小样方。记录每个小样方内所有木本植物的物种名称及胸径或基径。喀斯特林以壳斗科Fagaceae、金缕梅科Hamamelidaceae、鼠李科Rhamnaceae、豆科Fabaceae植物种为主(占比>5%),其中以滇青冈Cyclobalanopsis glaucoides、水锦树Wendlandia uvariifolia为优势种。土山天然林以壳斗科、大戟科Euphorbiaceae、楝科Meliaceae、漆树科Anacardiaceae、桑科Moraceae、豆科植物为主,其中以栓皮栎Quercus variabilis、白栎Quercus fabri为优势种。马尾松林以松科Pinaceae、楝科、报春花科Primulaceae、大麻科Cannabaceae为主,其中以马尾松为优势种。尾叶桉林以桃金娘科Myrtaceae、叶下珠科Phyllanthaceae、大戟科、夹竹桃科Apocynaceae植物为主,其中以尾叶桉为优势种。
-
在4个小样方中随机选择3个小样方,每个小样方设4个土壤采样点,采用环刀法采集表层(0~10 cm)土壤,并测定土壤温度、含水率和pH。将同一小样方内4个采样点的土壤混合成1个样本。最终,每个林型采集15个土壤样本,4个林型共采集60个样本,每个样本分别测定土壤养分和微生物群落。具体地,每个土样分成2份:一份风干用于测定土壤养分,另一份于−80 ℃冰箱保存,用于提取土壤DNA。土壤养分测定方法均参考《土壤农化分析》[15]。对每个土壤因子进行数据标准化,使每个因子均值为0,方差为1,用于后续分析。
使用MagaBio Soil/Feces Genomic DNA提取试剂盒(Bioer,杭州)提取DNA,并送至广东美格基因科技有限公司,在Illumina Novaseq 6000平台进行2×250 bp双端测序。细菌16S rRNA基因V3-V4区采用338F(5'-ACTCCTACGGGAGGCAGCA-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')引物扩增;真菌ITS2区采用ITS3F(5'-GCATCGATGAAGAACGCAGC-3')和ITS4R(5'-TCCTCCGCTTATTGATATGC-3')引物扩增。测序数据通过QIIME2平台处理,按97%相似度聚类操作分类单元(OTU),并与SILVA(细菌)和UNITE(真菌)数据库比对进行物种分类。采用PICRUSt2软件基于KEGG数据库对细菌进行功能预测,使用R语言FUNGuildR包基于FUNGuild数据库对真菌进行功能类群注释。为排除丰度或采样强度的影响,每个细菌和真菌样本分别稀释至38 868条和22 153条序列。
-
计算每个样本土壤细菌和真菌的物种丰富度指数、Shannon-Wiener指数、Simpson指数、均匀度指数,计算公式均参考文献[16]。对每个样本,计算各OTU (科或门)的相对丰度,即各OTU (科或门)的序列数占该样本总序列数的百分比。
通过单因素方差分析比较不同林型之间土壤微生物多样性、优势类群、功能类群和土壤理化性质的差异。使用基于Bray-Curtis距离的置换多元方差分析(PERMANOVA)和非度量多维尺度分析(NMDS)方法,分析土壤微生物群落组成的差异。通过指示物种分析确定各林型的指示OTU。使用Mantel检验分析微生物群落与植物群落组成(基于物种丰度)、土壤因子之间的相关性;同时,基于土壤微生物群落组成的Bray-Curtis距离和各土壤因子的欧氏距离、植物群落组成的Bray-Curtis距离,使用逐步线性回归模型解析各土壤因子、植物群落组成对微生物群落组成差异的解释度[17]。所有分析在R软件中进行,PERMANOVA分析使用vegan包的adonis2函数,Bray-Curtis距离和欧氏距离由vegdist函数计算;Mantel检验使用vegan包的mantel函数;影响因素的解释度通过relimpo包的calc.relimp函数确定;指示物种分析采用labdsv包中的indval函数。图表数据以平均值±标准误表示。
-
表1显示:除速效镁外,各土壤理化因子在不同林型间均存在显著差异。喀斯特林的总碳和总氮质量分数最高,土山天然林次之,人工林最低。天然林的总磷质量分数均显著高于人工林(P<0.05)。速效磷质量分数在土山天然林和尾叶桉林中较高,马尾松林次之,喀斯特林最低。土山天然林的速效钾质量分数最高,喀斯特林速效钾质量分数显著高于尾叶桉林(P<0.05),而与马尾松林无显著差异。喀斯特林的速效钙质量分数显著高于其他林型(P<0.05),土山天然林显著高于尾叶桉林(P<0.05),而与马尾松林无显著差异。喀斯特林的pH显著高于而温度显著低于其他林型(P<0.05),其余三者间无显著差异。尾叶桉林的含水率显著低于其他林型(P<0.05),其余三者间无显著差异。综上,除速效磷外,人工林的土壤养分水平低于天然林或与天然林相当。
林型 总碳/(g·kg−1) 总氮/(g·kg−1) 总磷/(g·kg−1) 速效磷/(mg·kg−1) 速效钾/(mg·kg−1) 喀斯特林 58.38±2.20 a 4.53±0.17 a 0.77±0.08 a 0.90±0.12 c 90.11±10.67 b 土山天然林 28.73±3.62 b 2.71±0.32 b 0.71±0.04 a 5.45±0.63 a 174.85±12.17 a 马尾松人工林 19.17±1.21 c 1.51±0.08 c 0.51±0.03 b 2.58±0.29 b 77.15±5.30 bc 尾叶桉人工林 14.06±1.39 c 1.03±0.10 d 0.45±0.04 b 3.68±1.19 a 60.54±2.52 c 林型 速效钙/(mg·kg−1) 速效镁/(mg·kg−1) pH 温度/℃ 含水率/% 喀斯特林 6 394.51±260.01 a 437.57±26.29 a 6.25±0.14 a 23.34±0.16 b 13.12±0.77 a 土山天然林 942.85±176.42 b 422.35±20.07 a 5.01±0.04 b 25.92±0.27 a 12.40±0.85 a 马尾松人工林 631.85±54.39 bc 360.43±22.45 a 5.15±0.06 b 26.45±0.06 a 14.36±0.46 a 尾叶桉人工林 484.56±57.32 c 371.09±16.05 a 4.94±0.03 b 26.42±0.07 a 9.38±0.25 b 说明:同列不同小写字母表示不同林型间差异显著(P<0.05)。 Table 1. Soil physical and chemical properties in different forest types
-
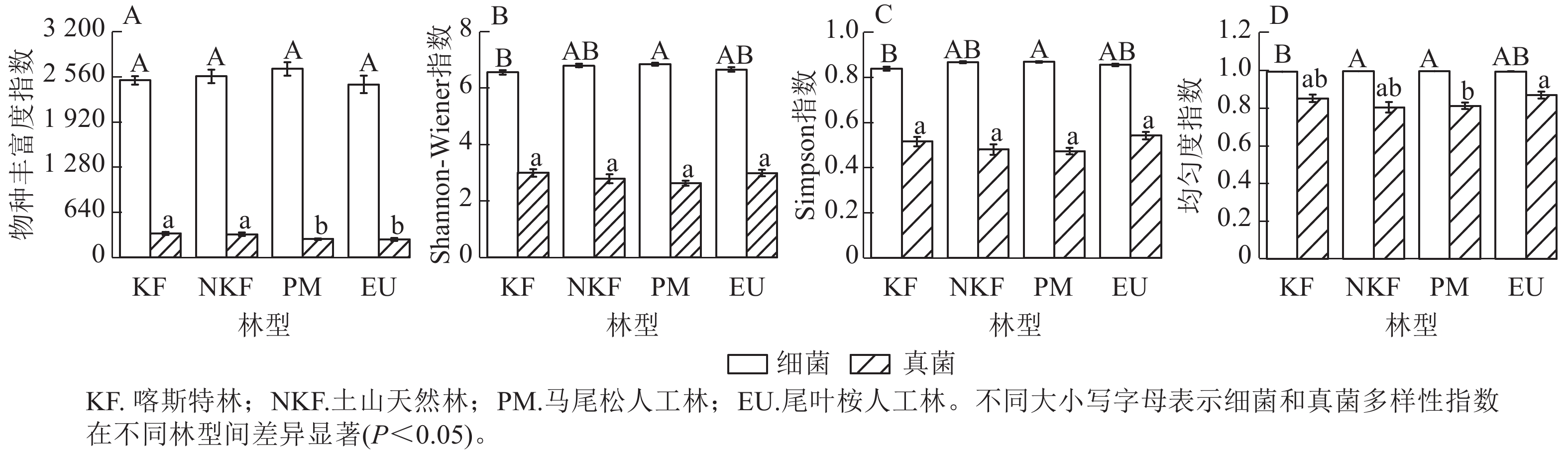
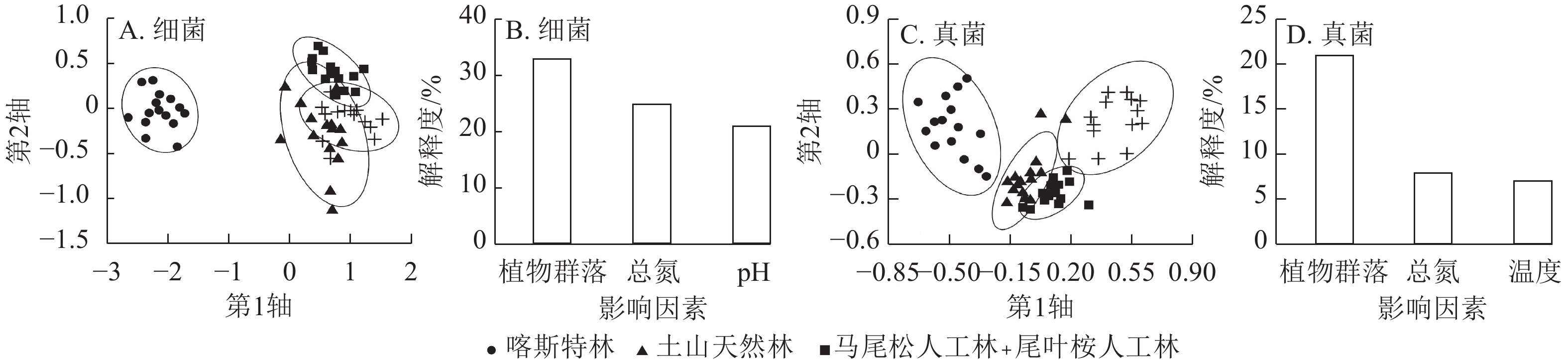
土壤细菌的物种丰富度指数在不同林型间无显著差异(图1A)。马尾松林的Shannon-Wiener和Simpson指数显著高于喀斯特林(P<0.05),但其余三者间无显著差异(图1B、图1C)。马尾松林和土山天然林的均匀度指数显著高于喀斯特林(P<0.05),但与尾叶桉林无显著差异,土山天然林、马尾松林、尾叶桉林间无显著差异(图1D)。PERMANOVA和NMDS分析表明:不同林型的细菌群落组成存在显著差异(F=12.23,R2=0.40,P<0.001,图2),任意2种林型之间的细菌群落组成均显著不同。

Figure 1. Diversity indices of soil bacteria and fungi in different forest types
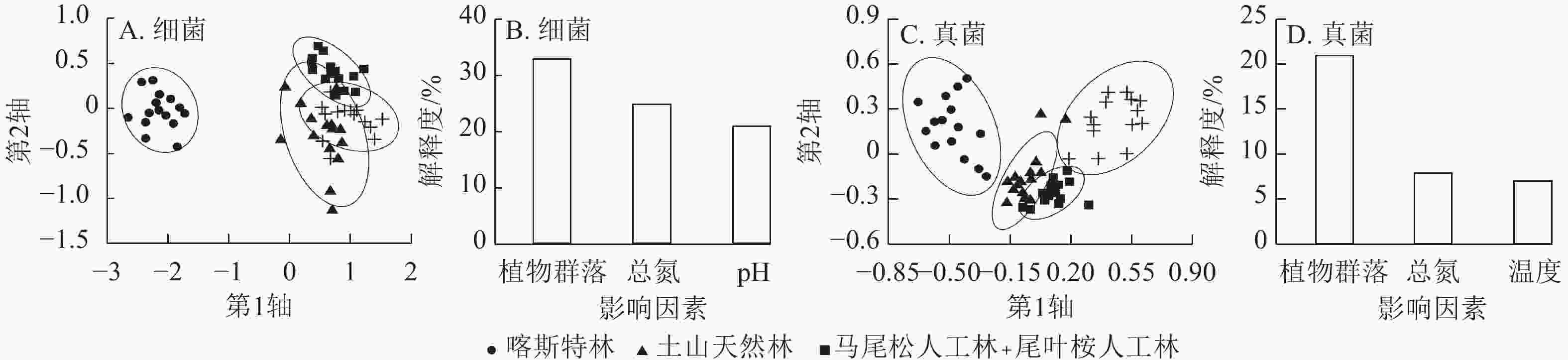
Figure 2. NMDS analysis of soil bacterial and fungal community composition across different forest types and the influencing factors of their compositional differences
在门水平上,尽管4种林型的细菌优势类群基本相同,但相对丰度在不同林型间存在显著差异(表2)。主要优势类群酸杆菌门Acidobacteria相对丰度在马尾松林中显著高于喀斯特林(P<0.05),但与土山天然林和尾叶桉林无显著差异,其余三者间也无显著差异。变形菌门Proteobacteria相对丰度在人工林中显著低于天然林(P<0.05)。绿弯菌门Chloroflexi相对丰度在尾叶桉林中最高,马尾松林次之,天然林最低。放线菌门Actinobacteria相对丰度在喀斯特林和尾叶桉林中显著高于土山天然林和马尾松林(P<0.05),但喀斯特林与尾叶桉林、土山天然林与马尾松林之间无显著差异。拟杆菌门Bacteroidetes相对丰度在喀斯特林中最高,土山天然林次之,人工林最低。疣微菌门Verrucomicrobia相对丰度在4种林型间无显著差异。
类群 门 相对丰度/% 喀斯特林 土山天然林 马尾松人工林 尾叶桉人工林 细菌 酸杆菌门Acidobacteria 29.23±1.60 b 34.99±3.30 ab 40.15±3.02 a 36.13±0.98 ab 变形菌门Proteobacteria 26.91±1.32 a 27.19±2.01 a 21.44±1.44 b 22.32±1.17 b 疣微菌门Verrucomicrobia 14.10±2.08 a 13.15±1.21 a 12.90±0.98 a 12.32±1.65 a 绿弯菌门Chloroflexi 5.45±0.42 c 5.58±0.36 c 8.52±0.58 b 11.13±0.67 a 放线菌门Actinobacteria 9.58±0.49 a 5.45±0.51 b 6.13±0.59 b 8.08±0.96 a 拟杆菌门Bacteroidetes 9.45±1.04 a 6.07±1.04 b 3.77±0.69 c 2.65±0.24 c 真菌 担子菌门Basidiomycota 49.95±6.46 a 48.79±8.99 a 53.37±3.53 a 48.79±8.99 a 被孢霉门Mortierellomycota 33.84±7.45 a 32.03±7.38 a 27.96±35.01 a 7.35±4.22 b 子囊菌门Ascomycota 12.70±2.53 b 16.35±1.61 b 15.99±2.44 b 41.50±6.22 a 说明:不同小写字母表示相同真菌门和相同细菌门不同林型间差异显著(P<0.05)。 Table 2. Major groups (relative abundance >5%) of soil bacterial and fungal communities at the phylum level in different forest types and their relative abundances
在科水平上,不同林型的细菌优势科存在差异(表3)。尾叶桉林和喀斯特林各自具有特有的优势科,而马尾松林和土山天然林则无特有优势科。其中,纤线杆菌科Ktedonobacteraceae为尾叶桉林的特有优势科,尾叶桉林的相对丰度最高,土山天然林和马尾松林次之,喀斯特林最低。梭菌科Pyrinomonadaceae和噬几丁质菌科Chitinophagaceae是喀斯特林的特有优势科,喀斯特林中梭菌科相对丰度显著高于其他林型(P<0.05),而其他林型间无显著差异。噬几丁质菌科相对丰度在喀斯特林中最高,土山天然林次之,人工林最低。大部分优势科为4种林型共有,除西索恩氏菌科Chthoniobacteraceae外,这些优势科的相对丰度在不同林型间均存在显著差异。其中,索力氏菌科Solibacteraceae在土山天然林和尾叶桉林中显著高于喀斯特林和马尾松林(P<0.05)。未知科2(酸杆菌门)在马尾松林中显著高于其他林型(P<0.05)。黄杆菌科Xanthobacteraceae在人工林中显著低于天然林(P<0.05)。未知科1(酸杆菌门)在喀斯特林中显著低于其他林型(P<0.05),而其他林型间无显著差异。
类群 门 科 相对丰度/% 喀斯特林 土山天然林 马尾松人工林 尾叶桉人工林 细菌 酸杆菌门Acidobacteria 未知科1 1.51±0.37 b 10.70±1.77 a 11.90±1.50 a 13.14±1.13 a 索力氏菌科Solibacteraceae 5.12±0.23 b 6.90±0.80 a 5.28±0.25 b 6.48±0.38 a 未知科2 6.33±1.35 b 6.2±1.21 b 10.04±0.86 a 6.06±0.54 b 梭菌科Pyrinomonadaceae 8.83±0.78 a 0.60±0.13 b 0.51±0.15 b 0.40±0.18 b 变形菌门Proteobacteria 黄杆菌科Xanthobacteraceae 10.08±0.44 a 11.15±0.65 a 7.45±0.77 b 7.60±0.50 b 疣微菌门Verrucomicrobia 西索恩氏菌科Chthoniobacteraceae 9.39±2.15 a 9.20±1.33 a 8.14±0.93 a 8.83±1.54 a 绿弯菌门Chloroflexi 纤线杆菌科Ktedonobacteraceae 0.37±0.18 c 3.28±0.66 b 3.47±0.86 b 8.12±0.85 a 拟杆菌门Bacteroidetes 噬几丁质菌科Chitinophagaceae 6.09±0.43 a 3.80±0.66 b 2.02±0.41 c 1.44±0.23 c 真菌 被孢霉门Mortierellomycota 被孢霉科Mortierellaceae 33.81±7.48 a 32.03±7.38 a 27.95±5.01 a 7.35±4.22 b 担子菌门Basidiomycota 红菇科Russulaceae 13.28±3.55 ab 27.42±8.69 a 5.96±3.45 b 9.31±4.29 b 蜡壳耳科Sebacinaceae 7.72±3.97 b 2.19±0.61 b 30.82±5.94 a 11.16±1.93b 革菌科Thelephoraceae 3.31±1.06 ab 3.78±1.44 ab 0.18±0.03 b 8.05±2.67 a 鹅膏菌科Amanitaceae 6.01±3.32 a 1.39±0.65 a 1.73±0.74 a 0.80±0.56 a 层腹菌科Hymenogastraceae 6.25±2.48 a 0.10±0.05 b 0.00±0.00 b 0.04±0.01 b 子囊菌门Ascomycota 船壳菌科Gloniaceae 0.07±0.03 b 1.21±0.74 b 5.19±2.61 b 24.58±5.71 a 说明:不同小写字母表示相同真菌科、细菌科不同林型间差异显著(P<0.05)。 Table 3. Major groups (relative abundance>5%) of soil bacterial and fungal communities at the family level in different forest types and their relative abundances
指示物种分析发现:喀斯特林细菌的指示OTU有314个(
https://zlxb.zafu.edu.cn/fileZJNLDXXB/journal/article/file/d5ea9075-6056-4245-a017-66c5a061609c.pdf ),集中(占比>5%)分布于梭菌科、西索恩氏菌科、噬几丁质菌科、黄杆菌科和索力氏菌科,占比为5.4%~10.2%。土山天然林有指示OTU 12个,分布较为零散。马尾松林有82个,集中于未知科2、土圈菌科Pedosphaeraceae、未知科1、索力氏菌科和未知科3(变形菌门),占比为6.1%~15.9%。尾叶桉林有75个,集中于纤线杆菌科、索力氏菌科、未知科1、分枝杆菌科Mycobacteriaceae和西索恩氏菌科,占比为5.3%~26.7%。这些指示OTU,除2个OTU的相对丰度为1.10%外,其余均<1.00%。Mantel检验显示:不同林型细菌群落组成与植物群落组成(r=0.76,P<0.001)、土壤因子(r=0.81,P<0.001)均呈极显著相关。逐步回归分析显示:不同林型细菌群落组成的差异主要受植物群落组成、土壤总氮质量分数和pH异质性的影响,解释度分别为33.4%、24.8%和20.9%,合计解释了群落组成79.1%的异质性(图2B)。
-
土壤真菌物种丰富度指数在人工林中显著低于天然林(P<0.05),但在天然林之间、人工林之间无显著差异(图1A)。Shannon-Wiener和Simpson指数在不同林型间无显著差异(图1B、C)。马尾松林的均匀度指数显著低于尾叶桉林(P<0.05),但与2种天然林无显著差异,其余三者间亦无显著差异(图1D)。PERMANOVA和NMDS分析显示:不同林型的真菌群落组成存在显著差异(F=7.33,R2=0.29,P<0.001,图2C),任意2种林型间的群落组成均显著不同。
在门水平上,4种林型真菌的优势类群相同(表2)。尾叶桉林中被孢霉门相对丰度显著低于其他林型(P<0.05),而子囊菌门则显著高于其他林型(P<0.05),其他林型间无显著差异。在科水平上,不同林型的优势科不完全一致(表3)。尾叶桉林和喀斯特林各有特有优势科,而马尾松林和土山天然林无特有优势科。革菌科为尾叶桉林特有优势科,其相对丰度显著高于马尾松林(P<0.05),但与2种天然林无显著差异。鹅膏菌科和层腹菌科为喀斯特林特有优势科,其中鹅膏菌科相对丰度在不同林型间无显著差异,层腹菌科相对丰度在喀斯特林中显著高于其他林型(P<0.05),其余三者间无显著差异。但大多数优势科为多个林型共有,这些科的相对丰度在不同林型之间存在显著差异。其中,被孢霉科相对丰度在尾叶桉林中显著低于其他林型(P<0.05),而船壳菌科相对丰度则显著高于其他林型(P<0.05),其他林型间无显著差异。红菇科相对丰度在土山天然林中显著高于人工林(P<0.05),而与喀斯特林无显著差异,其余三者间无显著差异。蜡壳耳科相对丰度在马尾松林中显著高于其他林型(P<0.05),其他林型间无显著差异。
指示物种分析显示:喀斯特林真菌指示OTU有24个(相对丰度为0.02%~9.30%),集中分布在被孢霉科(占33.3%)和链格孢科Herpotrichiellaceae (占8.3%)。土山天然林有19个(相对丰度为0.03%~3.60%),集中在红菇科(占15.8%)。马尾松林有25个(相对丰度为0.01%~26.90%),集中在被孢霉科、蜡壳耳科、丝盖伞科Inocybaceae和红菇科,占比为8.0%~16.0%。尾叶桉林有19个(相对丰度为0.03%~9.20%),集中在船壳菌科(占31.6%)和口蘑科Tricholomataceae (占10.5%)。
Mantel检验显示:不同林型真菌群落组成与植物群落组成(r=0.56,P<0.001)、土壤因子(r=0.41,P<0.001)均极显著相关。逐步回归分析显示:植物群落组成的差异是导致不同林型真菌群落组成差异的主要因素,单独解释了21.2%的异质性,其次为土壤总氮质量分数(7.8%)和温度(7.2%)的差异,三者合计解释了36.2%的异质性(图2D)。
-
细菌获得了6类生物代谢通路功能类群(表4)。结果显示:各林型均以代谢功能类群(包括碳水化合物代谢、氨基酸代谢、萜类与酮类化合物代谢、脂质代谢等)占绝对优势,其次为遗传信息处理,其他功能类群所占比例较低。尽管部分功能类群在不同林型之间存在统计学差异,但占比差异较小(<2%)。总体而言,不同细菌功能类群在各林型之间的差异不显著。
类群 功能类群 相对丰度/% 喀斯特林 土山天然林 马尾松人工林 尾叶桉人工林 细菌 代谢 82.47±0.12 a 81.76±0.14 b 81.17±0.20 b 81.45±0.15 b 遗传信息处理 11.09±0.15 b 11.23±0.14 b 11.85±0.14 a 11.55±0.09 ab 细胞过程 3.98±0.12 b 4.55±0.16 a 4.71±0.14 a 4.64±0.13 a 环境信息处理 1.93±0.03 a 1.91±0.02 a 1.86±0.01 a 1.91±0.02 a 人类疾病 0.25±0.02 a 0.21±0.04 a 0.14±0.03 a 0.18±0.03 a 有机系统 0.26±0.01 a 0.31±0.07 a 0.27±0.01 a 0.26±0.01 a 真菌 共生型 24.07±4.20 c 39.59±5.85 bc 44.03±5.00 b 64.06±4.83 a 腐生-共生型 31.69±4.81 a 34.38±5.14 a 20.32±2.76 ab 8.43±3.92 b 病理-腐生-共生型 2.68±1.02 a 1.24±0.27 a 1.48±0.20 a 4.74±3.08 a 腐生型 2.09±0.48 a 4.57±1.00 a 4.74±1.51 a 3.42±0.52 a 病理-腐生型 2.32±0.38 a 2.55±0.44 a 2.58±0.75 a 1.69±0.37 a 病理-共生型 0.13±0.05 a 2.39±2.20 a 0.13±0.08 a 0.13±0.04 a 病理型 0.61±0.20 a 1.44±0.87 a 0.50±0.10 a 0.14±0.02 a 说明:不同小写字母表示相同功能类群不同林型间差异显著(P<0.05)。 Table 4. Functional groups of soil bacterial and fungal communities in different forest types and their relative abundances
真菌确定了7种真菌营养类型(表4)。结果显示:各林型以共生型真菌和腐生-共生型真菌为优势,且不同林型相对丰度之间存在显著差异。其他营养类型相对丰度在不同林型中的比例较低,且无显著差异。其中,共生型真菌相对丰度在尾叶桉林中最高,且在马尾松林中显著高于喀斯特林(P<0.05),而与土山天然林无显著差异,2种天然林间无显著差异。腐生-共生型相对丰度在尾叶桉林中显著低于2种天然林(P<0.05),而与马尾松林无显著差异,其余三者间无显著差异。
-
为精准评估人工生态公益林土壤微生物多样性,本研究选取2种典型地带性天然林作为参照:一是代表稳定成熟生态系统的土山天然林,二是生态系统相对脆弱的喀斯特天然林(其土壤结构断裂、土层浅薄且肥力低)[18]。对比研究发现:马尾松和尾叶桉人工生态公益林的土壤肥力呈下降趋势。马尾松人工生态公益林的土壤细菌多样性显著高于喀斯特林,而与土山天然林无显著差异;尾叶桉人工生态公益林的细菌多样性与2种地带性天然林均无显著差异。2种人工生态公益林的土壤真菌多样性显著低于天然林。此外,土壤细菌群落组成在不同林型间存在显著差异,而功能类群在各林型间无显著差异;真菌群落组成和功能类群在不同林型间均存在显著差异。
本研究表明:植物群落组成是影响细菌群落组成差异的主要因素,与此前研究一致[19−20]。本研究发现豆科植物在喀斯特林(占比6.8%)和土山天然林(5.4%)中的比例较马尾松(2.8%)、尾叶桉(3.3%)人工林高,这可能导致固氮菌(如黄杆菌科)在天然林中较为丰富。土壤pH和总氮质量分数也影响细菌群落的组成,与已有研究一致[21−23]。喜碱型、富营养型菌群(如变形菌门、放线菌门、拟杆菌门[21−22, 24−25])在偏碱性喀斯特林、含氮量较高的土山天然林中更为丰富;而喜酸型、寡营养型菌群(如酸杆菌门、绿弯菌门[21−22, 24−25]),在偏酸性、含氮量较低的人工林中更为丰富。在科水平上,本研究发现:梭菌科、黄杆菌科和噬几丁质菌科的相对丰度与土壤pH、总氮均显著正相关,表现为喜碱、富营养型,在天然林中显著增加。相反,未知科1、索力氏菌科、未知科2和纤线杆菌科的相对丰度与土壤pH、总氮均显著负相关,表现为喜酸、寡营养型,在人工林中显著增加。这也解释了不同林型中指示OTU的差异。例如,喀斯特林的指示OTU以固氮和喜碱性菌群为主,土山天然林以固氮菌群为主,而2种人工林则以喜酸性菌群为主。这些影响因素通过影响细菌群落组成,进一步影响细菌多样性。以往研究发现,土壤细菌多样性与植物多样性及土壤总氮质量分数正相关[26−27],且碱性环境中多样性较高[28]。然而,本研究发现:尽管喀斯特林的土壤总氮较高且偏碱性,细菌多样性却最低。可能原因是喀斯特林土壤断裂且土层薄,导致土壤结构受损[18],从而降低了细菌多样性。其余林型细菌多样性差异不显著,可能与其土壤pH相似有关,而pH是影响细菌多样性的关键因子[29]。与本研究结果不同,谭宏伟等[11]发现:桉树用材林土壤酶活性降低,导致细菌多样性下降;而LAN等[12]则发现:施肥显著提高了桉树用材林的土壤养分和pH,从而增加细菌多样性。可能由于人工公益林未施肥且处于封禁管理状态,因此其细菌多样性未显著下降,这表明人工林生态功能已有效恢复。细菌功能类群在各林型间差异不显著,进一步支持了这一结论。尽管此前尚未有关于马尾松人工林细菌功能类群与天然林的对比研究报道,但对桉树用材林与天然林的对比研究也观察到类似现象[30],即代谢功能类群占据主导地位,而且两者之间的细菌功能类群差异并不显著。因此,鉴于PICRUSt功能预测方法的局限性,未来的研究应结合宏基因组测序以及氮磷等元素循环相关功能基因的分析,以便更准确地评估人工公益林生态功能的恢复状况。
相比细菌,土壤真菌通常与植物根系形成共生关系,因此植物群落在很大程度上影响真菌群落的组成[31]。2种天然林中,栎类为优势树种,导致被孢霉科[32]、红菇科[33]等与栎类共生的菌群显著增加;马尾松人工林中,与松树共生的被孢霉科[34]、蜡壳耳科[34]等菌群显著增加;尾叶桉人工林中,与尾叶桉共生的船壳菌科[35]显著增加。各林型的指示OTU与其优势植物形成共生关系。土壤总氮质量分数和温度也是影响真菌群落组成的主要因素,与已有研究结果一致[36−37]。本研究发现:土壤总氮质量分数较低且温度较高的尾叶桉林中,富营养型、喜冷的菌群(如被孢霉门[38])减少,而寡营养型、喜热的菌群(如子囊菌门[39])增加。在科水平上,被孢霉科和层腹菌科的相对丰度与土壤质量分数总氮呈正相关而与土壤温度呈负相关,表现为富营养型且喜冷,在土壤总氮质量分数较高、温度较低的喀斯特林中显著增加。相反,蜡壳耳科和船壳菌科的相对丰度与土壤总氮质量分数呈负相关,与土壤温度呈正相关,表现为寡营养型且喜热,在土壤总氮质量分数较低、温度较高的人工林中显著增加。与以往认为土壤真菌多样性与植物多样性、土壤总氮和温度呈正相关的研究不完全一致[27, 39]。本研究发现:尽管人工林土壤温度较高,但由于植物群落单一且土壤总氮质量分数较低,真菌多样性较低。相比之下,天然林土壤温度虽较低,但植物种类丰富且土壤总氮质量分数较高,真菌多样性较高。相反,ZHU等[13]发现:适度人为干扰促进了连作桉树用材林的土壤真菌多样性。综上,人工林植物群落的单一性是导致土壤真菌多样性下降的关键因素,真菌功能类群也随之发生变化。在人工林中,共生型类群显著高于天然林,可能是由于人工林以单一树种为主,长期进化关系促使形成单一共生菌群,同时单一环境有利于其繁殖和传播,进而形成优势类群,这可能导致人工林生态系统功能的单一化。在本研究的尾叶桉公益林中的发现与桉树用材林的研究结果吻合[30],均发现桉树纯林中的共生型真菌类群相对丰度显著高于天然林。然而,针对马尾松公益林,本研究结果与马尾松用材林存在差异。本研究发现马尾松公益林中共生型真菌类群丰度显著高于天然林,而WU等[40]及文翕等[41]的研究则表明两者差异不显著。造成这种差异的可能原因是,在WU等[40]和文翕等[41]研究的天然林中,尽管植物多样性较高,但共生型植物(如豆科植物、菌根植物)的占比可能也较高,从而提升了其共生型真菌类群的相对丰度。同样地,鉴于FUNGuild功能预测方法的局限性,未来应结合宏基因组测序等手段,以更准确地评估人工公益林生态功能的恢复状况。
-
本研究发现:与地带性天然林相比,马尾松和尾叶桉人工公益林植物群落组成单一、土壤养分下降,显著降低了土壤真菌多样性并改变了其功能类群,但土壤细菌多样性和功能类群均未发生显著的变化。因此,通过与其他乡土阔叶树种混交,定期施肥,提高土壤肥力,有利于恢复和维持马尾松、尾叶桉人工生态公益林的微生物多样性。
HTML
1.1. 研究区概况
1.2. 样地设置及植物群落调查
1.3. 土壤样品采集与处理
1.4. 数据分析
2.1. 不同林型土壤理化性质差异
2.2. 不同林型土壤细菌多样性和群落组成差异及其影响因素
2.3. 不同林型土壤真菌多样性和群落组成差异及其影响因素
2.4. 不同林型土壤细菌和真菌群落功能类群的差异
 2025-0305-附件材料.pdf
2025-0305-附件材料.pdf
|

|
 DownLoad:
DownLoad: