





-
森林可燃物是指在一定地段上的所有可燃物质的组成[1]。作为森林燃烧的3个要素(森林可燃物、火源和火环境)之一,森林可燃物是林火发生的物质基础和首要条件[2-3]。同时,与另外2个要素相比,可燃物更易于调控和管理[4-5]。因此,通过对森林可燃物进行管理可以有效降低林火的发生[6-7]。然而,随着可燃物调控措施的实施,森林的林内环境、植被生长及可燃物的分布会发生变化,如林分郁闭度的改变会影响林内光热条件,从而对林下植被产生影响[8-9]。因此,在可燃物调控中,不仅要达到降低火险的目标,还要考虑对林地环境的生态影响[10-11]。所以,调控强度的选择对于恢复和维持林地生态系统的多样性和稳定性具有十分重要的意义。目前,对森林可燃物的研究多集中在可燃物分布[7, 12]、可燃物调控措施[13]及可燃物与火行为[12, 14]等方面。而对调控措施实施后林下植被组成及生物量、生物多样性变化等研究很少,缺少不同调控措施的对比及对生态环境影响的评价。北京西山地区拥有丰富的森林资源,侧柏Platycladus orientalis作为华北地区典型的针叶树种和北京的乡土树种,在该地区分布广泛,其中优势树种为侧柏的小班129个,面积2 422.0 hm2,占44.0%;侧柏蓄积74 932.3 m3,占42.2%[15]。但由于该地区春季干旱多风、可燃物载量较高且人为活动频繁,火灾隐患较高[6-7]。因此,开展侧柏林可燃物调控并对调控措施进行评价,选择合适的调控方案,不仅可以有效降低该地区的森林火险,同时对森林群落多样性恢复和发展具有重要意义。本研究以北京西山林场侧柏林为对象,调查分析所选林分的可燃物垂直分布特征,并制定相应的调控技术,通过野外调查和相关实验,测定对比不同调控后侧柏林下植被的生物多样性和光合生理特性的变化,对可燃物调控的影响作出评价,以期为当地侧柏林科学管理提供参考。
-
北京西山试验林场位于北京西郊,连接海淀、石景山及门头沟区。经营总面积为5 931 hm2,森林覆被率为98%。有林地面积为5 457 hm2,其中灌木林地为383 hm2,疏林地为19.5 hm2,未成林造林地为62.7 hm2[15]。西山森林组成树种中,乔木主要包括侧柏、油松Pinus tabulaeformis、栓皮栎Quercus variabilis、刺槐Robinia pseudoacacis等。灌木主要包括孩儿拳头Grewia biloba var. parviflora、构树Broussonetia papyrifera、黄栌Cotinus coggygria、荆条Vitex negundo var. heterophylla、胡枝子Lespedeza bicolor、黑枣Diospyros lotus等。草本主要包括荩草Arthraxon hispidus、隐子草Cleistogenes squarrosa、紫花地丁Viola philippica、细叶薹草Carexs tenophylla、铁杆蒿Artemisia sacrorum等。西山森林虽然是人工林,但由于多年精心培育,林木生长良好,林相整齐,林分郁闭,森林资源颇具特色。
-
在北京西山林场,挑选有代表性的人工侧柏林,主要选择林相整齐、人为干扰小的地段,共4块区域(分别标号为Ⅰ、Ⅱ、Ⅲ和Ⅳ)。
-
2016年3月,在上述4块区域内分别设置3块20 m×20 m的标准地(a, b和c),进行样地基本信息和可燃物的调查。样地基本信息调查主要包括在样地内每木检尺,测胸径、树高、枝下高,运用对角线截距抽样法测林分郁闭度。同时记录标准地的海拔、坡度、坡向等地型因子(表1)。
表 1 侧柏样地的地形、林分因子基本情况
Table 1. Basic information of topography and stand factors in P. orientalis forest
样地号 林龄/a 海拔/m 经纬度 坡度/(°) 坡向 郁闭度 密度/(株·hm−2) 胸径/cm 树高/m 死枝/m 活枝高/m Ⅰa 29 263 40°00′47″N, 116°11′59″E 30 东北 0.85 1 425 13.4 7.6 1.6 2.8 Ⅰb 0.82 1 438 13.9 7.8 1.5 2.8 Ⅰc 0.84 1 429 13.6 7.7 1.6 2.9 Ⅱa 29 297 40°00′44″N, 116°11′56″E 26 北 0.87 1 570 11.2 7.5 1.9 2.5 Ⅱb 0.86 1 531 11.1 7.9 1.7 2.3 Ⅱc 0.86 1 562 11.7 7.2 1.9 2.6 Ⅲa 29 275 40°00′50″N, 116°11′59″E 25 东北 0.78 1 525 10.6 6.5 1.8 2.8 Ⅲb 0.79 1 529 10.4 6.1 1.9 2.9 Ⅲc 0.80 1 519 10.7 6.3 1.6 2.8 Ⅳa 29 270 40°00′51″N, 116°12′00″E 22 东北 0.85 1 125 16.2 8.1 1.8 3.3 Ⅳb 0.87 1 148 16.5 8.4 1.9 3.4 Ⅳc 0.85 1 137 16.2 8.2 1.9 3.6 -
可燃物调查主要包括冠层可燃物(乔木枝叶)、林下灌木和幼树(胸径小于3.0 cm)可燃物、地被凋落物层可燃物(调查时草本处于枯萎状态,归为地被凋落物层),具体调查方法参考王叁等[16]。在野外取样测可燃物湿质量,并带回实验室测定含水率,用于计算干可燃物的负荷量。依据侧柏林林分结构及可燃物垂直分布特征,并结合西山林场营林抚育的相关措施,制定不同强度的可燃物调控方案(表2)。
表 2 侧柏林可燃物调控方案
Table 2. Fuel management plan in the studied P. orientalis forest
样地号 调控方案 林分特征 可燃物分布特征 潜在林火类型 主要调控措施 辅助调控措施 Ⅰ ck 树高中等,密度大 地表凋落物和灌木可燃物负荷量大 易由地表火发展为树冠火 对照(不调控) 对照(不调控) Ⅱ P1 树高中等,密度很大 地表凋落物负荷量大,灌木可燃物
负荷量小易发生地表火 修枝高度到2.5 m 适当清理地表凋落物 Ⅲ P2 树高较低,密度很大
枝下高低地表凋落物和灌木可燃物负荷量大 易由地表火发展为树冠火 修枝高度到3.0 m
并割灌适当清理地表凋落物 Ⅳ P3 树高较高,密度低 地表凋落物和灌木可燃物负荷量大 易由地表火发展为树冠火 修枝高度到3.5 m
并割灌适当清理地表凋落物 -
实施调控1 a后,于2017年8月,在每个样地内,利用对角线法设置5个2 m×2 m的灌木样方,共计60个,调查灌木及小乔木(胸径小于3.0 cm)的种类、高度、盖度。同时在每个样地的灌木样方内设置2个l m×l m的草本样方,共计120个,记录草本的种类、高度、多度及盖度等[17-18]。
-
林下植物生长型主要包括灌木层和草本层2种,选择常用的α多样性特征对2种生长型进行计算。主要指标包括物种丰富指数(S)、Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)、Pielou均匀度指数(J)等[19-20]。
-
在调控1 a后的2017年8月,对林下植被进行光合特性的测定。这一时期降水丰沛,树木生理活动不受土壤水分胁迫。根据林下植被多样性调查结果,选择了优势灌木(孩儿拳头和构树)测定其光合作用。光响应曲线测定:在每块样地中,随机选取长势良好的孩儿拳头和构树各3株,利用LI-COR 6400在9:00−11:00(此时段环境因子适宜,植物生理活动活跃[21])测定其活体叶片的光响应曲线。测定时选择树冠外围光照充足,完全展开的无病虫害健康叶片,每株植物选择3~5个叶片测定。光合作用光响应曲线设定光合有效辐射(PAR)梯度为1 600、1 400、1 200、1 000、800、600、300、150、100、50、20、0 μmol·m−2·s−1。二氧化碳摩尔分数控制为400 μmol·mol−1,与大气二氧化碳摩尔分数相同。每种灌木叶片进行3~4次重复测定。利用LED红蓝光源测定梯度光照强度下,不同遮光处理和全光照的净光合速率,绘制光合-光响应(Pn-PAR)曲线[22]。根据光合-光响应曲线拟合的对数方程求得光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(DR)及最大净光合速率(Pmax),并比较其在不同光环境下的差异。将光合光响应曲线在低光量子通量密度下的净光合速率做直线回归,求得各遮光程度的表观量子效率(AQY)[23]。
-
可燃物负荷量及垂直分布特征与光合特性指标用Excel统计计算,并制图。生物多样性指标用Excel进行数据整理及各项指标的计算,并采用SPSS 19.0进行单因素方差分析(one-way ANOVA)差异性检验,显著水平为α=0.05。
-
由图1看出:侧柏林样地Ⅰ、Ⅱ、Ⅲ和Ⅳ在0~1 m层可燃物负荷量分布最大,分别为879.92、786.61、1 017.37、936.73 g·m−2,主要为地表凋落物(侧柏、灌木的枯枝落叶以及死地被物)和灌木可燃物(孩儿拳头、构树、黄栌、荆条、酸枣Ziziphus jujuba var. spinosa等)。其中样地Ⅰ、Ⅲ、Ⅳ中的灌木可燃物负荷量较大,火灾发生时容易由地表火蔓延为树冠火。乔木死枝可燃物负荷量较低,林火发生对乔木的影响比较小。大部分侧柏样地,2~9 m的可燃物量呈单峰分布,主要集中在乔木的中层位置。因此,减少乔木中下层可燃物负荷量,可以有效降低地表火蔓延至树冠火的可能。
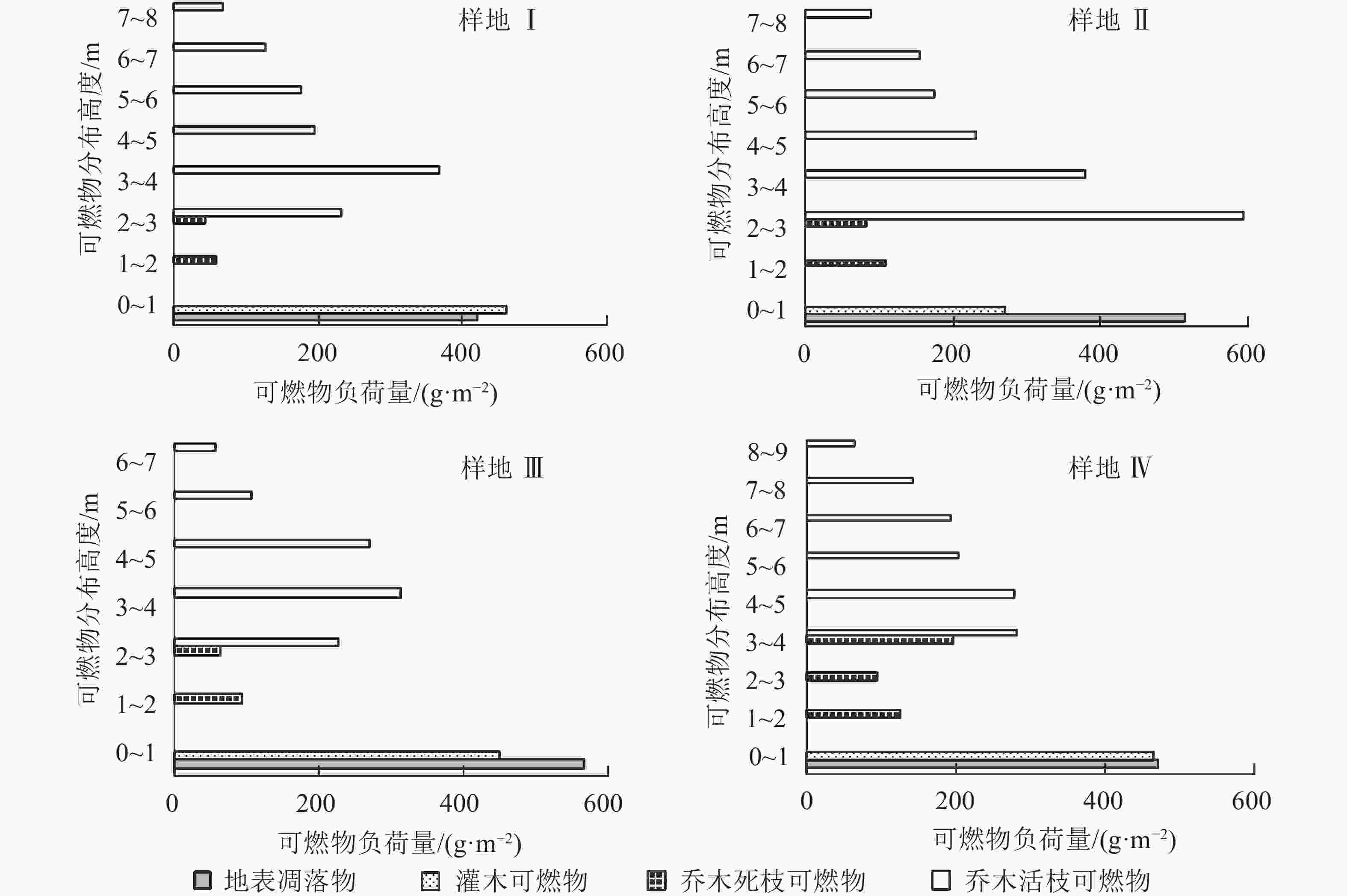
图 1 侧柏林样地易燃可燃物垂直分布
Figure 1. Vertical distribution of flammable fuel in four P. orientalis plots
-
由表3可以看出:实施调控后,林内灌木层物种丰富度、物种优势度、物种多样性及物种均匀度较对照林分均有增加。其中,P1和P2灌木层的物种优势度和均匀度显著高于对照林分(P<0.05),调控后P2灌木层的物种多样性较对照林分有显著增加(P<0.05),P3灌木层的物种优势度、物种多样性和物种均匀度均高于对照林分,但差异并不显著(P>0.05)。调控林分P1草本层的各项多样性指标均低于对照林分,P2和P3草本层的物种优势度、物种多样性以及物种均匀度均高于对照林分,其中,P2草本层的物种优势度和物种多样性指数显著高于对照林分(P<0.05),而P1、P2、P3的物种均匀度则与对照林分相比没有显著差异(P>0.05)。
表 3 侧柏林不同调控方案下灌木层和草本层生物多样性指数
Table 3. Biodiversity index of shrub layer and grass layer under different fuel treatments in the studied P. orientalis forest
调控方案 植物生长型 丰富度指数 Simpson指数 Shannon-Wiener指数 均匀度指数 ck 灌木层 8 0.57±0.04 B 1.12±0.03 B 0.67±0.06 B 草本层 15 0.56±0.09 b 1.33±0.14 b 0.57±0.13 a P1 灌木层 7 0.72±0.03 A 1.37±0.17 AB 0.83±0.03 A 草本层 10 0.48±0.14 b 1.02±0.39 b 0.51±0.15 a P2 灌木层 8 0.75±0.02 A 1.44±0.07 A 0.83±0.03 A 草本层 18 0.73±0.09 a 1.80±0.30 a 0.73±0.10 a P3 灌木层 10 0.69±0.13 AB 1.41±0.30 AB 0.76±0.09 AB 草本层 14 0.63±0.12 ab 1.38±0.35 ab 0.65±0.14 a 说明:数据为平均值±标准差。同列不同大写字母表示灌木层不同处理之间同一指标差异显著(P<0.05),同列不同小写字母表示草 本层不同处理之间同一指标差异显著(P<0.05) -
图2可以看出:P2和P3调控后孩儿拳头的净光合速率(Pn)的变化趋势要高于P1和对照,且随着光合有效辐射(PAR)梯度的增加,P2和P3的净光合速率拟合曲线趋于重叠。而构树的净光合速率变化趋势则表现为:调控组均高于对照,但随着光合有效辐射梯度的增加,这3种调控之间的净光合速率变化差异不大。
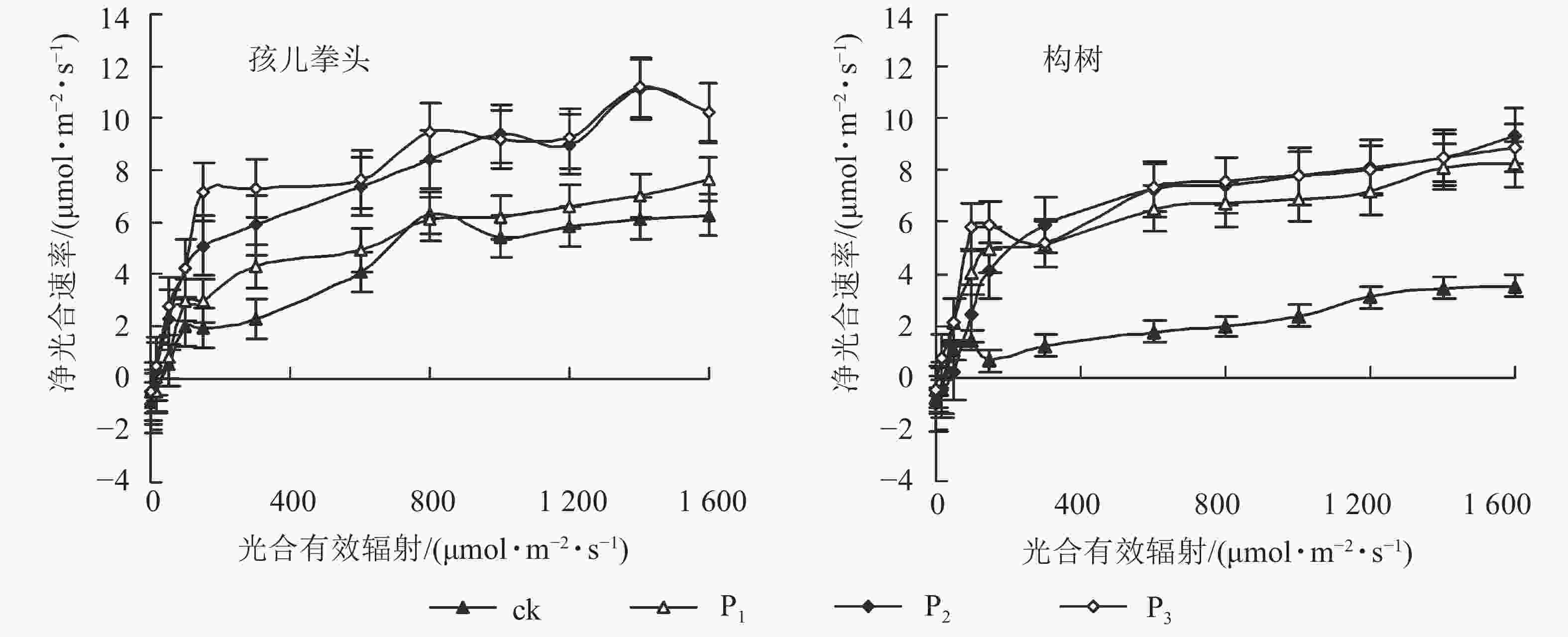
图 2 不同光合有效辐射下孩儿拳头和构树的净光合速率
Figure 2. Net photosynthetic rate of G. biloba var. parviflora and B. papyrifera under different photosynthetically active radiations
不同调控措施实施后,侧柏林下孩儿拳头和构树的光合生理指标发生变化。其中孩儿拳头和构树的表观量子效率均低于对照林分,而光补偿点、光饱和点、暗呼吸速率、最大净光合速率则高于对照林分(表4)。值得注意的是,P2调控后光补偿点和暗呼吸速率较对照林分增加幅度最小,但光饱和点和最大净光合速率较对照林分增加幅度最大。
表 4 不同处理林下优势灌木(孩儿拳头、构树)主要光合生理特征指标
Table 4. Photosynthetic physiological characteristics of main dominant shrub species(G. biloba var. parviflora, B. papyrifera) under different fuel treatments
调控方案 植物生长型 表观量子效率 光补偿点/
(mol·m−2·s−1)光饱和点/
(mol·m−2·s−1)暗呼吸速率/
(mol·m−2·s−1)最大净光合速率/
(μmol·m−2·s−1)ck 孩儿拳头 0.05±0.01 A 4.70±0.27 D 301.98±6.84 D 0.20±0.02 D 7.87±0.14 D 构树 0.05±0.01 a 11.90±0.02 d 161.54±4.69 d 0.46±0.01 d 3.52±0.07 d P1 孩儿拳头 0.03±0.01 B 7.77±0.13 B 353.73±5.47 B 0.50±0.01 B 8.77±0.06 C 构树 0.03±0.01 b 16.77±0.12 b 300.03±7.82 b 0.66±0.01 b 8.87±0.21 c P2 孩儿拳头 0.04±0.01 B 6.40±0.02 C 398.42±6.06 A 0.31±0.02 C 12.04±0.08 A 构树 0.03±0.01 b 14.43±0.39 c 338.40±6.49 a 0.55±0.01 c 10.62±0.22 a P3 孩儿拳头 0.04±0.01 B 12.89±0.01 A 331.73±5.77 C 0.67±0.01 A 11.80±0.05 B 构树 0.03±0.01 b 19.93±0.23 a 238.04±4.64 c 0.88±0.01 a 9.64±0.26 b 说明:数据为平均值±标准差。同列不同大写字母表示孩儿拳头不同处理之间同一指标差异显著(P<0.05),同列不同小写字母表示构树 不同处理之间同一指标差异显著(P<0.05) -
不同调控后林下植物多样性发生了改变。对林下灌木层而言,P1和P2调控后林下灌木的多样性显著增加,P3调控后林下灌木的多样性有增加的趋势,但与对照林分无显著差异。对林下草本层而言,P2调控后林下草本生物多样性显著增加,而P1和P3调控后林下草本生物多样性与对照林分没有显著差异。整体来看,P2方案更适宜调控后侧柏林下植被多样性的恢复和增加。这在降低森林火险的同时,也对维持森林群落结构的稳定性和多样性具有重要意义。
调控后,侧柏林下优势灌木的最大净光合速率、光饱和点、光补偿点、暗呼吸速率均高于对照林分,而表观量子效率与对照林分相比减小。SARIJEVA等[24]研究发现:生活在弱光中的植物的表观量子效率通常较高,而光补偿点和最大光合速率则比较低。VALLADARES等[25]对巴拿马热带雨林中灌丛的光适应性研究中发现同样的结果。这表明调控后林下灌草降低了对弱光的利用,但增强了对强光的利用和光合潜力。同时,本研究表明:P2处理后更利于林下灌草光合产物的积累。因此,对于研究地区的侧柏林,其可燃物调控措施应主要以中等强度修枝为主,并适当的进行割灌和清理地表凋落物。
可燃物调控作为一项干扰措施,对森林生态系统本身就有一定的“破坏”作用,然而可燃物调控措施的实施同时也改变了林分结构,造成林内光热条件发生变化,在一定程度上又提高了林下植被的多样性和光合能力[26]。本研究结合西山林场实际情况(西山侧柏林为人工纯林,林分组成单一,林下植被也比较简单),在实施调控措施降低了火灾发生概率的同时,增加林下植被多样性和光合产物积累,提高森林生态系统的稳定性,更符合其人工林的可持续经营管理。
本研究也发现了一些需要改变或优化的地方:①本研究是以林下植被的生物多样性和光合特性的改变作为对可燃物调控影响的评价,因此只选择了立地因子相似的单一林分类型,以尽量减少其他因素的干扰。所以,在以侧柏林为代表的森林可燃物调控评价的基础上,应增加对其他树种的研究。②可燃物空间分布主要包括垂直分布和水平分布,本研究只针对可燃物垂直分布特征进行了分析,这对于研究地表火向树冠火蔓延有一定的参考价值。而水平方向上可燃物的分布对林火蔓延的影响也至关重要,在接下来的研究中应扩大研究区域,增加可燃物水平分布调查,为西山林场可燃物管理提供更加全面、立体的参考依据。③可燃物调控除了对林下植被的生物多样性和光合特性产生影响,还可能对土壤、土壤微生物、水文等产生影响,增加对这些方面的研究可以对调控措施的生态影响做出更加全面的评价。
-
可燃物调控措施改变了林下植被的多样性,其中P2调控后林下灌木、草本的多样性均显著大于对照。可燃物调控措施改变了林下植被的光合特性,P2调控后林下灌木的光饱和点和最大净光合速率增幅最大,光合产物积累将更加明显。可燃物调控措施能有效降低森林火险,但同时会对森林群落结构产生一定的“破坏”作用,选择适当强度的调控方式,对森林群落多样性的恢复和稳定性的维持具有十分重要的意义。就本研究而言,P2调控方案最适于该研究区域。
Impact assessment of fuel regulation measures in Platycladus orientalis forest in Western Hills of Beijing
-
摘要:
目的 针对北京西山侧柏Platycladus orientalis林可燃物分布特征,实施不同强度可燃物调控措施,并依据调控后林下植被的生物多样性和光合特性的变化,对可燃物调控措施的影响进行评价。 方法 在北京西山试验林场,选取华北地区针叶林—侧柏林,通过对林分可燃物分布特征的调查和分析,制定以修枝、割灌为主的调控措施,分别为未调控(ck)、修枝高度2.5 m并适当清理地表凋落物(P1)、修枝高度3.0 m和割灌并适当清理地表凋落物(P2)、修枝高度3.5 m和割灌并适当清理地表凋落物(P3),每个处理设置3块20 m×20 m的标准地,对比调控与未调控林分的林下植物多样性和优势灌木的光合特性变化。 结果 不同强度可燃物调控措施均改变了林下植被的生物多样性,其中P2调控下灌木层和草本层的物种优势度和多样性及灌木层的物种均匀度显著高于对照;调控林分的林下优势灌木(主要是孩儿拳头Grewia biloba var. parviflora和构树Broussonetia papyrifera)的最大净光合速率、光饱和点、光补偿点、暗呼吸速率均比对照增加,而表观量子效率则减小,其中P2调控措施下的林下植物光合产物的积累更明显。 结论 调控措施改变了林分结构,使林下光照强度增加。不同调控强度对植被生物多样性和光合特性的影响存在差异,P2调控最有利于研究区域的林分恢复。图2表4参26 Abstract:Objective Based on the fuel distribution characteristics of the Platycladus orientalis forest in Western Hills of Beijing, fuel regulation measures of different intensities were implemented, and effects of the fuel regulation measures were evaluated according to the biodiversity and photosynthetic characteristics of understory vegetation. Method P. orientalis, a typical coniferous tree species in North China was selected as the research object. By investigating and analyzing distribution characteristics of P. orientalis forest fuel, regulation measures focusing on pruning, cutting and irrigation were formulated, including: unregulated stands (ck), pruning height 2.5 m and cleaning the surface litter appropriately (P1), pruning height 3.0 m and cutting shrubs and cleaning the surface litter appropriately (P2), pruning height 3.5 m and cutting shrubs and cleaning the surface litter appropriately (P3). Three sample plots of 20 m×20 m were established for each treatment, and changes of plant diversity and photosynthesis attributes of dominant shrubs were compared between regulated and unregulated stands. Result The biodiversity of understory vegetation was altered after different fuel managements, among which the species dominance (D) and diversity (H) of shrub layer and grass layer and the species evenness (J) of shrub layer after P2 management were significantly higher than those of ck. Compared with ck, the maximum net photosynthetic rate, light saturation point, light compensation point, dark respiration rate of the understory vegetation (Grewia biloba var. parviflora and Broussonetia papyrifera) all increased, while the apparent quantum efficiency decresed after P2 management. We observed that the accumulation of understory vegetation photosynthetic products was more obvious with P2 management. Conclusion The implementation of regulation measures changed the forest structure and increased the light intensity. The effects of different regulation intensity on biodiversity and photosynthetic characteristics of understory vegetation were different. P2 management was most conducive to stand recovery in the study area. [Ch, 2 fig. 4 tab. 26 ref.] -
Key words:
- forest ecology /
- Platycladus orientalis /
- fuel regulation /
- fuel load /
- biodiversity /
- photosynthetic characteristics /
- assessment
-
图 1 侧柏林样地易燃可燃物垂直分布
Figure 1 Vertical distribution of flammable fuel in four P. orientalis plots
图 2 不同光合有效辐射下孩儿拳头和构树的净光合速率
Figure 2 Net photosynthetic rate of G. biloba var. parviflora and B. papyrifera under different photosynthetically active radiations
表 1 侧柏样地的地形、林分因子基本情况
Table 1. Basic information of topography and stand factors in P. orientalis forest
样地号 林龄/a 海拔/m 经纬度 坡度/(°) 坡向 郁闭度 密度/(株·hm−2) 胸径/cm 树高/m 死枝/m 活枝高/m Ⅰa 29 263 40°00′47″N, 116°11′59″E 30 东北 0.85 1 425 13.4 7.6 1.6 2.8 Ⅰb 0.82 1 438 13.9 7.8 1.5 2.8 Ⅰc 0.84 1 429 13.6 7.7 1.6 2.9 Ⅱa 29 297 40°00′44″N, 116°11′56″E 26 北 0.87 1 570 11.2 7.5 1.9 2.5 Ⅱb 0.86 1 531 11.1 7.9 1.7 2.3 Ⅱc 0.86 1 562 11.7 7.2 1.9 2.6 Ⅲa 29 275 40°00′50″N, 116°11′59″E 25 东北 0.78 1 525 10.6 6.5 1.8 2.8 Ⅲb 0.79 1 529 10.4 6.1 1.9 2.9 Ⅲc 0.80 1 519 10.7 6.3 1.6 2.8 Ⅳa 29 270 40°00′51″N, 116°12′00″E 22 东北 0.85 1 125 16.2 8.1 1.8 3.3 Ⅳb 0.87 1 148 16.5 8.4 1.9 3.4 Ⅳc 0.85 1 137 16.2 8.2 1.9 3.6  下载: 导出CSV
下载: 导出CSV
表 2 侧柏林可燃物调控方案
Table 2. Fuel management plan in the studied P. orientalis forest
样地号 调控方案 林分特征 可燃物分布特征 潜在林火类型 主要调控措施 辅助调控措施 Ⅰ ck 树高中等,密度大 地表凋落物和灌木可燃物负荷量大 易由地表火发展为树冠火 对照(不调控) 对照(不调控) Ⅱ P1 树高中等,密度很大 地表凋落物负荷量大,灌木可燃物
负荷量小易发生地表火 修枝高度到2.5 m 适当清理地表凋落物 Ⅲ P2 树高较低,密度很大
枝下高低地表凋落物和灌木可燃物负荷量大 易由地表火发展为树冠火 修枝高度到3.0 m
并割灌适当清理地表凋落物 Ⅳ P3 树高较高,密度低 地表凋落物和灌木可燃物负荷量大 易由地表火发展为树冠火 修枝高度到3.5 m
并割灌适当清理地表凋落物
下载: 导出CSV
表 3 侧柏林不同调控方案下灌木层和草本层生物多样性指数
Table 3. Biodiversity index of shrub layer and grass layer under different fuel treatments in the studied P. orientalis forest
调控方案 植物生长型 丰富度指数 Simpson指数 Shannon-Wiener指数 均匀度指数 ck 灌木层 8 0.57±0.04 B 1.12±0.03 B 0.67±0.06 B 草本层 15 0.56±0.09 b 1.33±0.14 b 0.57±0.13 a P1 灌木层 7 0.72±0.03 A 1.37±0.17 AB 0.83±0.03 A 草本层 10 0.48±0.14 b 1.02±0.39 b 0.51±0.15 a P2 灌木层 8 0.75±0.02 A 1.44±0.07 A 0.83±0.03 A 草本层 18 0.73±0.09 a 1.80±0.30 a 0.73±0.10 a P3 灌木层 10 0.69±0.13 AB 1.41±0.30 AB 0.76±0.09 AB 草本层 14 0.63±0.12 ab 1.38±0.35 ab 0.65±0.14 a 说明:数据为平均值±标准差。同列不同大写字母表示灌木层不同处理之间同一指标差异显著(P<0.05),同列不同小写字母表示草 本层不同处理之间同一指标差异显著(P<0.05)
下载: 导出CSV
表 4 不同处理林下优势灌木(孩儿拳头、构树)主要光合生理特征指标
Table 4. Photosynthetic physiological characteristics of main dominant shrub species(G. biloba var. parviflora, B. papyrifera) under different fuel treatments
调控方案 植物生长型 表观量子效率 光补偿点/
(mol·m−2·s−1)光饱和点/
(mol·m−2·s−1)暗呼吸速率/
(mol·m−2·s−1)最大净光合速率/
(μmol·m−2·s−1)ck 孩儿拳头 0.05±0.01 A 4.70±0.27 D 301.98±6.84 D 0.20±0.02 D 7.87±0.14 D 构树 0.05±0.01 a 11.90±0.02 d 161.54±4.69 d 0.46±0.01 d 3.52±0.07 d P1 孩儿拳头 0.03±0.01 B 7.77±0.13 B 353.73±5.47 B 0.50±0.01 B 8.77±0.06 C 构树 0.03±0.01 b 16.77±0.12 b 300.03±7.82 b 0.66±0.01 b 8.87±0.21 c P2 孩儿拳头 0.04±0.01 B 6.40±0.02 C 398.42±6.06 A 0.31±0.02 C 12.04±0.08 A 构树 0.03±0.01 b 14.43±0.39 c 338.40±6.49 a 0.55±0.01 c 10.62±0.22 a P3 孩儿拳头 0.04±0.01 B 12.89±0.01 A 331.73±5.77 C 0.67±0.01 A 11.80±0.05 B 构树 0.03±0.01 b 19.93±0.23 a 238.04±4.64 c 0.88±0.01 a 9.64±0.26 b 说明:数据为平均值±标准差。同列不同大写字母表示孩儿拳头不同处理之间同一指标差异显著(P<0.05),同列不同小写字母表示构树 不同处理之间同一指标差异显著(P<0.05)
下载: 导出CSV
-
[1] 王明玉,舒立福,姚树人. 北京地区森林可燃物人工调控技术[J]. 森林防火, 2012, 9(3): 46 − 48. WANG Mingyu, SHU Lifu, YAO Shuren. The artificial control technology of forest fuel in Beijing [J]. For Fire Prev, 2012, 9(3): 46 − 48. [2] 王凯,牛树奎. 基于Rothermel模型的北京鹫峰国家森林公园潜在火行为[J]. 浙江农林大学学报, 2016, 33(1): 42 − 50. WANG Kai, NIU Shukui. Research on the potential fire behavior in Jiufeng National Forest Park of Beijing based on the Rothermel Model [J]. J Zhejiang A&F Univ, 2016, 33(1): 42 − 50. [3] 孙龙,王千雪,魏书精,等. 气候变化背景下我国森林火灾灾害的响应特征及展望[J]. 灾害学, 2014, 29(1): 12 − 17. SUN Long, WANG Qianxue, WEI Shujing, et al. Response characteristics and prospect of forest fire disasters in the context of climate change in China [J]. J Catastrophology, 2014, 29(1): 12 − 17. [4] MINAS J, HEARNE J, MARTELL D. Anintegrated optimization model for fuel management and fire suppression preparedness planning [J]. Ann Oper Res, 2015, 232(1): 201 − 215. [5] CALVO L, SANTALLA S, VALBUENA L, et al. Post-fire natural regeneration of a Pinus pinaster forest in NW Spain [J]. Plant Ecol, 2008, 197(1): 81 − 90. [6] 王明玉,周荣伍,赵凤君,等. 北京西山森林潜在火行为及防火林带有效宽度分布研究[J]. 火灾科学, 2008, 17(4): 209 − 215. WANG Mingyu, ZHOU Rongwu, ZHAO Fengjun, et al. Potential fire behavior and distribution of effective width of shaded fuel break in Xishan of Beijing [J]. Fire Saf Sci, 2008, 17(4): 209 − 215. [7] 王晓丽,牛树奎,马钦彦,等. 北京地区主要针叶林易燃可燃物垂直分布[J]. 北京林业大学学报, 2009, 31(2): 31 − 35. WANG Xiaoli, NIU Shukui, MA Qinyan, et al. Vertical distribution of flammable fuels in major conifer forests of Beijing area [J]. J Beijing For Univ, 2009, 31(2): 31 − 35. [8] 杨健,孔健健,刘波. 林火干扰对北方针叶林林下植被的影响[J]. 植物生态学报, 2013, 37(5): 474 − 480. YANG Jian, KONG Jianjian, LIU Bo. A review of effects of fire disturbance on understory vegetation in boreal coniferous forest [J]. Chin J Plant Ecol, 2013, 37(5): 474 − 480. [9] 李明泽,谢雨,邸雪颖,等. 大兴安岭林区地表可燃物载量遥感估测模型[J]. 东北林业大学学报, 2014, 42(5): 60 − 63. LI Mingze, XIE Yu, DI Xueying, et al. Remote sensing model for forest surface fuel loads in Daxing’an Mountains [J]. J Northeast For Univ, 2014, 42(5): 60 − 63. [10] 高仲亮,陈鹏宇,舒立福. 林下可燃物管理方式对森林碳储量的影响[J]. 安徽农业科学, 2013, 41(23): 9639 − 9641. GAO Zhongliang, CHEN Pengyu, SHU Lifu. Impacts of fuel management on forest carbon storage [J]. J Anhui Agric Sci, 2013, 41(23): 9639 − 9641. [11] 周涧青,刘晓东,郭怀文. 大兴安岭南部主要林分地表可燃物负荷量及其影响因子研究[J]. 西北农林科技大学学报, 2014, 42(6): 131 − 137. ZHOU Jianqing, LIU Xiaodong, GUO Huaiwen. Surface fuel loading and relevant influencing factors of main forest types in southern Daxing’an ling [J]. J Northwest A&F Univ, 2014, 42(6): 131 − 137. [12] 牛树奎,王叁,贺庆棠,等. 北京山区主要针叶林可燃物空间连续性研究——可燃物垂直连续性与树冠火发生[J]. 北京林业大学学报, 2012, 34(3): 1 − 7. NIU Shukui, WANG San, HE Qingtang, et al. Spatial continuity of fuels in major coniferous forests in Beijing mountainous area: fuel vertical continuity and crown fire spread [J]. J Beijing For Univ, 2012, 34(3): 1 − 7. [13] 金琳. 北京十三陵林场低山针叶林可燃物分布及调控技术研究[D]. 北京: 北京林业大学, 2012. JIN Lin. Fuel Distribution and Fuel Control Technology in the Low Mountain Coniferous Forest of Ming Tombs Forest Farm[D]. Beijing: Beijing Forestry University, 2012. [14] 王明玉,舒立福,赵凤君,等. 北京西山可燃物特点及潜在火行为[J]. 林业科学, 2010, 46(1): 84 − 90. WANG Mingyu, SHU Lifu, ZHAO Fengjun, et al. Characteristics of forest fuel and potential fire behavior in Xishan Mountain of Beijng [J]. Sci Silv Sin, 2010, 46(1): 84 − 90. [15] 申晋. 西山林场森林资源信息管理系统的研建[D]. 北京: 北京林业大学, 2008. SHEN Jin. Establishment of Forest Resources Information Management System for Beijing Xishan Trial Forest Farm[D]. Beijing: Beijing Forestry University, 2008. [16] 王叁,牛树奎,李德,等. 云南松林可燃物的垂直分布及影响因子[J]. 应用生态学报, 2013, 24(2): 331 − 337. WANG San, NIU Shukui, LI De, et al. Vertical distribution of fuels in Pinus yunnanensis forest and related affecting factors [J]. Chin J Appl Ecol, 2013, 24(2): 331 − 337. [17] 高贤明,马克平,陈灵芝. 暖温带若干落叶阔叶林群落物种多样性及其与群落动态的关系[J]. 植物生态学报, 2001, 25(3): 283 − 290. GAO Xianming, MA Keping, CHEN Lingzhi. Species diversity of some deciduous broad-leaved forests in the warm-temperate zone and its relations to community stability [J]. Chin J Plant Ecol, 2001, 25(3): 283 − 290. [18] 方精云,沈泽昊,唐志尧,等. “中国山地植物物种多样性调查计划”及若干技术规范[J]. 生物多样性, 2004, 12(1): 5 − 9. FANG Jingyun, SHEN Zehao, TANG Zhiyao, et al. The protocol for the survey plan for plant species diversity of China’s mountains [J]. Biodiversity Sci, 2004, 12(1): 5 − 9. [19] 孔凡洲,于仁成,徐子钧,等. 应用Excel软件计算生物多样性指数[J]. 海洋科学, 2012, 36(4): 57 − 62. KONG Fanzhou, YU Rencheng, XU Zijun, et al. Application of excel in calculation of biodiversity indices [J]. Mar Sci, 2012, 36(4): 57 − 62. [20] 王健铭,董芳宇,巴海•那斯拉,等. 中国黑戈壁植物多样性分布格局及其影响因素[J]. 生态学报, 2016, 36(12): 3488 − 3498. WANG Jianming, DONG Fangyu, Bahai Nasiya, et al. Plant distribution patterns and the factors influencing plant diversity in the Black Gobi Desert of China [J]. Acta Ecol Sin, 2016, 36(12): 3488 − 3498. [21] 潘瑞炽. 植物生理学[M]. 北京: 高等教育出版社, 2001. [22] 赵勋. 越南安息香不同种源苗期光合特性研究[D]. 杭州: 浙江农林大学, 2011. ZHAO Xun. Study on the Photosynthesis Characteristics of Different Provenances Styrax tonkinensis in Seedling[D]. Hangzhou: Zhejiang A&F University, 2011. [23] KNAPP E E, KEELEY J E, BALLENGER E A, et al. Fuel reduction and coarse woody debris dynamics with early season and late season prescribed fire in a Sierra Nevada mixed conifer forest [J]. For Ecol Manage, 2005, 208(1/3): 383 − 397. [24] SARIJEVA G, KNAPP M, Lichtenthaler H K. Differences in photosynthetic activity, chlorophyII and carotenoids levels and in chlorophyII fluorescence parameters in green sun and shade leaves of Ginkgo and Fagus [J]. J Plant Physiol, 2007, 164: 950 − 955. [25] VALLADARES F, WRIGHT S J, LASSO E, et al. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest [J]. Ecology, 2000, 81(7): 1925 − 1936. [26] 孙广贵,张命军. 不同强度间伐对长白山天然林林下植物多样性的影响[J]. 森林工程, 2011, 27(5): 13 − 19. SUN Guanggui, ZHANG Mingjun. Effect of thinning intensity on species diversity of different forest stands in Changbai Mountain [J]. For Eng, 2011, 27(5): 13 − 19. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190343
 点击查看大图
点击查看大图

计量
- 文章访问数: 2983
- HTML全文浏览量: 992
- PDF下载量: 44
- 被引次数: 0
