





-
矮化是一种重要的农艺性状, 在改善空间和土地利用率, 调整栽培密度, 提高抗倒伏能力等方面具有明显的形态特征优势[1]。竹类矮化措施在生产实践中主要应用于3个方面:①中国南方及长江流域冰冻雪灾给竹类生产带来严重的经济损失, 破坏了生态环境[2-4]。钩梢矮化是抵御冰雪风折灾害的有效措施。②笋用竹设施栽培受到常规温室高度的限制[5-6], 每年秋冬季降温前盖膜需要钩梢, 矮化植株方便日常经营管理。③园林景观中矮化竹植株构型具有较高观赏价值[7]。目前, 关于竹类矮化方法及矮化后笋产量[8-9]、光合生理[10]以及材性力学性质[11]等方面已有诸多研究。生产中常用的竹类矮化方法有钩梢[3-4]或利用植物生长调节剂[3, 5, 7, 12]抑制竹居间分生组织生长达到矮化目标。但钩梢会直接带走大量秆枝叶, 造成营养生长损耗。同时成竹株高较高、竹秆硬度强增加了钩梢难度。使用植物生长调节剂造成药剂残留且连续多次施药效果受到天气影响, 矮化成本较高。此外, 通过断鞭[13]、剥除笋箨[14]、修剪[15]限制营养供给来控制高度生长的竹类矮化方法也有研究报道, 但在生产实践中并不常见。绿竹Dendrocalamopsis oldhami是中国南方地区优良的笋材两用丛生竹种, 其材性优良、竹笋产量高, 具有较好的经济和生态价值[16]。但绿竹鲜笋不耐储存, 限制了绿竹笋的销售范围。绿竹笋在北方蔬菜市场尚属空白, 发展笋用绿竹具有较好的经济前景。随着"南竹北移"的实施, 受日光温室高度限制, 秋冬季需要钩梢。因此, 探索一种易操作、无药剂残留且不影响竹子正常生长的矮化方法尤为必要。光合作用是植物生长发育物质能源积累的基础[17]。光合能力与植物不同植株构型有着密切关系。习玉森等[18]指出矮化型桃Amygdalus persica在强光、高温胁迫下较正常植株光抑制程度轻, 物质积累能力强。罗静等[19]指出矮化苹果Malus pumila苗叶绿素含量增加具有较高的净光合速率而早产。本研究在借鉴成竹秋冬季钩梢实践基础上, 将矮化时间提前至笋期, 提出竹笋截梢的矮化方法, 比较不同高度竹笋截梢对绿竹生长的影响, 并从叶绿素荧光动力学角度分析矮化后绿竹的光合生理状况, 为绿竹矮化栽培提供参考。
-
研究地位于绿竹原产地福建省三明市尤溪县(25°58′08″N, 118°09′09″E)。该区属中亚热带季风性湿润气候, 1月平均气温为8.0~12.0 ℃, 7月平均气温为26.6~28.9 ℃。无霜期为312.0 d, 降水量为1 600.0~1 800.0 mm, 土壤类型为山地红壤。主要植被有马尾松Pinus massoniana、杉木Cunninghamia lanceolata、青冈Cyclobalanopsis glauca、甜槠Castanopsis eyrei、油茶Camellia oleifera、山杜英Elaeocarpus sylvestris、石楠Photinia serratifolia等。绿竹林地原由水稻Oryza sativa田改造而成, 存在的主要自然灾害为低温冻害。竹林密度为825丛·hm-2, 竹林年龄结构:2年生:1年生为1:2, 每丛竹株数4~6株。当年不挖笋, 全部留养母竹, 按照绿竹丰产栽培经验进行日常经营管理。
-
在2017年7月下旬至8月上旬绿竹出笋盛期进行试验处理, 共置5个截梢处理(表 1), 分别记作H1、H2、H3、H4、H5, 以不截梢处理为对照(ck), 每个处理选择7丛绿竹, 共计42丛。每丛选择基径为4.0~5.0 cm, 长势良好、无病虫害、生长基本一致的绿竹笋3~4株(竹丛中其他笋不作处理, 自然生长)。测量笋体基径及高度, 按照竹笋高度的20%截除笋梢幼嫩部分并挂标签牌。
表 1 绿竹不同高度竹笋截梢处理概况
Table 1. General situation among different height bamboo shoot truncation treatments of D.oldhami
处理 截梢前笋高/cm 截梢长度/cm 截梢后笋高/cm H1 60 12 48 H2 90 18 72 H3 120 24 96 H4 150 30 120 H5 180 36 144 -
2018年1月上旬绿竹高生长结束后, 以挂标签牌的绿竹为测定对象。每个处理随机择20株绿竹测量生长指标, 选择5株绿竹测定叶绿素质量分数及叶绿素荧光参数, 取中部生长基本一致的健康、成熟叶作为测定样本。
-
调查绿竹株高、成竹率、枝下高、节数、分枝率、第一盘主枝长度。枝下高为竹秆最下端第1盘分枝到地面垂直高度; 主枝长度为竹秆最下端第1盘最长枝长度。分枝率=分枝节数/(枝下节数+分枝节数)×100%;成竹率=成竹数/处理笋数×100%。
-
采用混合液浸取-分光光度计法测定[20]。将采集的鲜叶洗净、擦干、去除中脉、剪碎混合均匀后, 天平秤取0.100 g叶片放入盛有10 mL提取液(纯丙酮和无水乙醇1:1配成)的具塞试管中, 置于黑暗环境中叶片失绿直至完全变白。分别测定波长为645和663 nm下的光密度(D), 并根据Arnon公式计算叶绿素质量分数。wchla=[12.72D(663)-2.59D(645)]×V/(103×W), wchlb=[22.88D(645)-4.67D(663)]×V/(103×W), wchl=[20.29D(645)+8.05D(663)]×V/(103×W), 其中:wchla、wchlb和wchl分别表示叶绿素a、叶绿素b和总叶绿素质量分数(mg·g-1), D(645)为波长645 nm处的光密度, D(663)为波长663 nm处的光密度, V为提取液总量(mL), W为样品质量(g)。
-
测定方法参考宋莉英等[21]的方法。采用Imaging PAM-2100(德国WALZ公司)便携式脉冲调制式叶绿素荧光仪测定绿竹叶片的叶绿素荧光参数。测定时间为晴天无风的9:00-11:00, 测量前使叶片暗适应30 min, 选定5个圆形测试目标区域, 然后打开测量光(0.5 μmol·m-2·s-1)测定初始荧光(Fo), 饱和光脉冲2 700 μmol·m-2·s-1(脉冲时间0.8 s)诱导最大荧光(Fm)、可变荧光(Fv=Fm-Fo)、PSⅡ最大光化学效率(Fv/Fm)。待荧光曲线基本稳定, 打开单饱和白光脉冲1次, 此后测得PSⅡ实际光量子效率(Yield)、电子传递速率(ETR)、光化学猝灭系数(qP)及非光化学猝灭系数(qNP)。
-
数据统计和作图由Excel 2013完成。用SPSS 21.0对不同竹笋截梢处理下绿竹生长指标、叶绿素质量分数以及荧光参数进行单因素方差分析(one-way ANOVA)和Pearson相关性分析, Duncan多重比较法进行显著性差异分析。
-
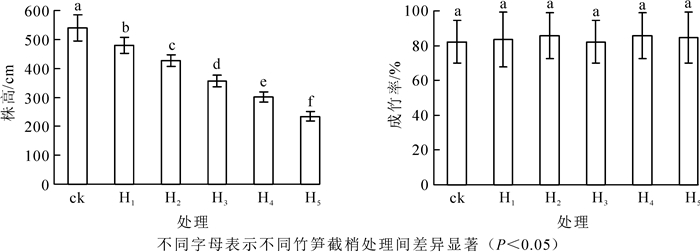
由图 1可知:竹笋截梢可以有效控制绿竹株高生长, 各竹笋截梢处理之间绿竹株高达极显著差异(P<0.01)。截去笋梢长度越长, 即截去笋梢部位笋节越多, 成竹后株高越矮。其中H1(60 cm)笋截梢后, 株高继续生长431.60 cm, H5(180 cm)笋截梢后, 株高继续生长90.70 cm。对株高y(cm)和竹笋截梢前绿竹笋高度x(cm)建立函数表达式为:y=599.49-1.95x(R2=0.90, P<0.01)。H5处理株高最低, 绿竹株高由对照539.40 cm降至234.70 cm, 较对照降低了56.49%, 达到了矮化栽培高度要求。竹笋不同截梢处理与对照的绿竹成竹率均为82.14%~85.71%, 成竹率差异未达到显著水平(P>0.05), 表明竹笋截梢处理不影响绿竹正常成活。
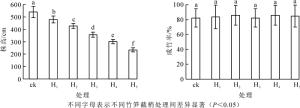
图 1 不同竹笋截梢处理下绿竹的株高和成竹率
Figure 1. Plant height and survival rate of D.oldhami under different bamboo shoot truncation treatments
-
从表 2可见:随着绿竹株高降低, 枝下高、节数、分枝率及主枝长度均达显著差异(P<0.05)。竹笋截梢后枝下高呈不断降低趋势, H5处理枝下高最低, 较对照显著下降36.69%, 与其他组差异均显著; 竹笋截梢后笋梢部分笋节被截去, 因此竹节相应减少, 节数与株高有相同的变化趋势。H5节数较对照降低45.59%, 除与H4处理无显著差异外, 与其他各组均有显著差异; 在分枝率方面, H3、H4和H5竹笋截梢处理较对照分别降低了14.10%、19.02%和12.13%。竹笋截梢后节数降低, 节上的分枝盘数减少, 因此, 分枝率变小; H4和H5主枝长度与对照均达到显著差异, 分别增长了10.00%和8.45%, 竹笋截梢促进了主枝长度生长。竹笋截梢后绿竹形态指标变化系数从大到小为株高(56.49%)、节数(36.69%)、枝下高(36.69%)、分枝率(19.02%)、主枝长度(11.06%)。表明竹笋截梢对绿竹株高影响最大, 其次为节数, 主枝长度影响最小。
表 2 不同竹笋截梢处理绿竹其他形态变化
Table 2. Morphological indexes changes of D.oldhami under different bamboo shoot truncation treatments
处理 枝下高/cm 节数 分枝率/% 主枝长度/cm ck 123.75±23.34 a 19.85±1.78 a 67.86±4.24 a 216.05±17.60 bc H1 119.05±20.75 ab 16.95±1.82 b 70.09±4.58 a 209.05±30.27 c H2 107.50±14.61 ab 15.95±0.83 b 67.95±4.74 a 219.25±20.82 bc H3 104.25±39.99 b 12.40±1.56 c 58.29±9.17 b 219.15±42.87 bc H4 110.45±18.65 ab 11.60±1.53 cd 54.95±7.50 b 239.95±9.83 a H5 78.35±21.03 c 10.80±3.20 d 59.63±11.71 b 234.30±42.06 ab 说明:同列不同小写字母表示不同竹笋截梢处理间差异显著(P<0.05) -
由表 3可知:竹笋截梢处理与对照绿竹叶绿素a、叶绿素b、总叶绿素及叶绿素a/b均差异显著(P<0.05)。H4和H5处理的叶绿素a较高, 显著高于H1和对照; 处理H2、H3、H4和H5的叶绿素b较高且处理间差异不显著; 总叶绿素从大到小为H5、H3、H4、H2、H1、ck, 不同竹笋截梢处理的总叶绿素变化有差异, 总体呈上升趋势, 绿竹总叶绿素在H5处理下最大, 较对照显著提高了65.34%, 与H2、H3、H4处理无显著差异; 对照叶绿素a/b最大, 显著高于竹笋截梢处理。H5处理较对照叶绿素a/b显著降低了29.11%。叶绿素a、叶绿素b及总叶绿素随着株高的降低而增加, 叶绿素a/b降低。
表 3 不同竹笋截梢处理绿竹叶绿素质量分数及组成变化
Table 3. Changes of chlorophyll content and composition ratio of D. oldhami under different bamboo shoot truncation treatments
处理 叶绿素a/(mg·g-1) 叶绿素b/(mg·g-1) 总叶绿素/(mg·g-1) 叶绿素a/b ck 1.95±0.20 c 0.76±0.02 b 2.71±0.21 b 2.56±0.27 a H1 1.92±0.010 c 1.01±0.11 b 2.93±0.19 b 1.91±0.18 b H2 2.15±0.23 bc 1.79±0.24 a 3.94+0.38 a 1.21±0.17 d H3 2.28±0.22 bc 1.82+0.18 a 4.10+0.13 a 1.27±0.23 cd H4 2.45±0.54 ab 1.59±0.42 a 4.04±0.62 a 1.69±0.75 bcd H5 2.81±0.33 a 1.66±0.45 a 4.47±0.63 a 1.81±0.54 bc 说明:同列不同小写字母表示不同竹笋截梢处理间差异显著(P<0.05) -
从图 2可知:竹笋截梢处理的初始荧光(Fo)与对照差异不显著(P>0.05);竹笋截梢处理提高了PSⅡ最大光化学效率(Fv/Fm), 其中:H4、H5较对照显著提高了15.86%和16.46%, 而竹笋截梢处理间未发现显著差异; ETR和PSⅡ实际光量子产量(Yield)随着株高降低, 有相同的变化趋势, H1、H2处理均与对照无显著差异, H5处理下最大, 较对照分别提高了48.63%和40.81%。不同竹笋截梢处理光化学猝灭系数(qP)变化有一定差异, 但总体呈不断上升趋势。H4、H5处理下化学猝灭系数较对照组差异均达到了显著水平, 化学猝灭系数最大的为H5处理, 较对照提高了74.35%。各竹笋截梢处理绿竹叶片的非光化学猝灭系数(qNP)均有显著降低, 对照处理的非光化学猝灭系数最大。H5处理的非光化学猝灭系数较对照降低了47.58%。非光化学猝灭系数与化学猝灭系数有着相反的变化趋势。不同竹笋截梢处理后绿竹PSⅡ最大光化学效率、PSⅡ实际光量子产量、电子传递速率及化学猝灭系数均高于对照, 而非光化学猝灭系数降低。叶绿素荧光参数表明竹笋截梢增加绿竹叶片的光能利用效率。
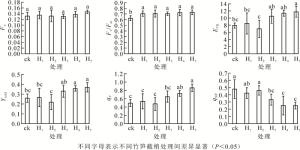
图 2 不同竹笋截梢处理下绿竹叶绿素荧光参数变化
Figure 2. Chlorophyll fluorescence parameters changes of D. oldhami under different bamboo shoot truncation treatments
-
相关分析(表 4)显示:株高与总叶绿素以及PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光量子产量(Yield)、电子传递速率(ETR)、光化学猝灭系数(qP)呈负相关, 与叶绿素a/b及非光化学猝灭系数(qNP)呈显著正相关。总叶绿素与叶绿素a/b呈显著负相关。PSⅡ实际光量子产量与电子传递速率、光化学猝灭系数呈显著正相关, 光化学猝灭系数与非光化学猝灭系数呈显著负相关。Pearson相关性分析表明:绿竹植株构型与叶绿素质量分数及叶绿素荧光特性有紧密关系。
表 4 绿竹株高与叶绿素质量分数及叶绿素荧光参数相关性分析
Table 4. Correlation analysis of plant height, chlorophyll content, chlorophyll fluorescence parameters of D. oldhami
指标 株高 总叶绿素 叶绿素a/b Fo Fv/Fm Yield ETR qP 总叶绿素 -0.809** 叶绿素a/b 0.368* -0.563** Fo -0.339 0.197 -0.048 FV/Fm -0.536* 0.410* -0.284 0.140 Yield -0.574* 0.372* -0.021 -0.150 0.169 ETR -0.607* 0.398* -0.068 -0.141 0.192 0.990** qP -0.697* 0.435* -0.030 0.044 0.177 0.895** 0.895** qNP 0.704** -0.391* -0.281 -0.206 -0.268 -0.637** -0.640** -0.719** 说明:*表示显著相关(P<0.05), **表示极显著相关(P<0.01).株高、叶绿素质量分数和叶绿素荧光参数样本n=25 -
竹高度生长依靠笋节居间分生组织的分生细胞分裂、伸长生长来增加节间的长度[22]。笋梢部分笋节密集, 笋节发育成为竹节。竹笋截梢按照竹笋高度20%截去梢头部分, 随着竹笋高度增加, 竹笋截梢强度增加, 截去的笋节越多, 竹节相应减少, 因此, 成竹矮化效果越明显。本研究发现:随株高降低, 枝下高、节数及分枝率减少, 主枝长度增长, 地上部分营养生长重新分配, 植株形态相应发生明显变化。其中, 绿竹竹笋高H5(180 cm)截梢处理后, 株高由对照的539.40 cm降低至234.70 cm, 较对照降低了56.49%。官凤英等[5]对绿竹喷施0.8 g·L-1多效唑后发现株高、枝下高分别降低了45.30%和46.70%, 周建革等[3]对毛竹Phyllostachys edulis喷施2.0%矮壮素后株高和枝下高分别降低了28.12%和30.37%。绿竹和毛竹存在共同特点即株高降低后, 枝下高降低。枝下高降低不利于挖笋施肥等经营活动, 需要加强相应剪除靠近地面枝条等竹林抚育措施来克服不利影响。竹笋截梢会使成竹高度降低, 竹材产量相应会降低, 不宜用在材用林上。
竹笋在成竹过程不同高度时期均有可能退笋, 退笋与笋体高度密切相关[23]。一般竹笋高于40 cm, 退笋现象较少。竹笋截梢各处理后绿竹成竹率在82.14%~85.71%, 与对照无显著差异, 表明竹笋截梢不影响绿竹成活。这可能是由于竹笋截梢处理选择高度为60~180 cm竹笋, 生长旺盛, 具有一定的抵抗力, 所以退笋率低, 这与郑郁善等[24]和岳祥华等[25]研究毛竹及紫竹Phyllostachys nigra的退笋规律基本一致。
-
植物体构件之间存在协调反馈机制, 即当某一构件部分抑制生长或缺失时, 剩余构件表现一个资源再分配, 某些功能增强的现象。这种补偿机制是植物应对外界扰动的生长策略[26-28]。郑士光等[29]研究发现:柠条Caragana microphylla在平茬后根系提高了对水分和养分的吸收, 促进地下根系生长。尚富华等[30]指出:毛白杨Populus tomentosa修枝后会提高剩余枝叶的光合速率等途径补偿。本研究表明:不同竹笋截梢处理后, 绿竹叶绿素a、叶绿素b及总叶绿素随着株高的降低而增加, 而叶绿素a/b降低。叶绿素a有利于吸收长波光, 叶绿素b促进吸收短波光。当叶绿素a/b减少时, 植物对蓝紫光的利用[31]效率增加。总叶绿素增加, 使得叶片叶肉细胞光合活性增强[32]。叶绿素荧光技术可以间接无损伤地研究光合作用过程中能量吸收传递与转化等特征[33]。当叶片内囊体破坏时, PSⅡ光系统反应中心失活, 初始荧光(Fo)增加[34]。本研究发现:不同竹笋截梢处理间的初始荧光(Fo)差异不显著, 表明竹笋截梢处理未对绿竹叶片内囊体造成不利影响。竹笋截梢处理提高了PSⅡ反应过程潜在活性, 促进了光合电子从PSⅡ反应中心到库源的传递速率, 使得PSⅡ最大光化学效率(Fv/Fm)增加。当光能过剩时, 非光化学猝灭系数(qNP)增加[35]。竹笋截梢处理非光化学猝灭系数较对照显著降低, 降低了叶片热耗散。随着株高降低, PSⅡ实际光量子效率(Yield)、电子传递速率(ETR)增加, 电子传递的量子产额增加, 促进暗反应的光合碳同化和有机物积累[36]。这与陈洪国[37]和魏亚娟等[38]对菊花Chrysanthemum morifolium及榆叶梅Prunus triloba通过使用植物生长调节剂获得矮化植株构型后光合特性变化规律类似, 表明矮化植株一定程度上光合色素含量增加, 光合效率提高。Pearson相关性分析表明:PSⅡ光化学功能、叶绿素质量分数与绿竹株高显著相关。竹笋截梢处理后绿竹叶绿素质量分数提高及叶绿素荧光参数表现更高的光能利用效率。本研究认为可能的原因有:一方面竹笋截梢后顶端优势去除后, 作为株高降低的补偿, 促进了枝叶萌生。枝叶生长有助于空间拓宽能力增加对光能的获取; 另一方面竹笋截梢绿竹株高降低后, 竹林冠层光照条件发生改变。改善光环境, 提高光能利用效率, 以获得更多的光合同化产物积累。
-
根据栽培目标选择适合的绿竹笋高度截梢可以有效控制株高生长, 而且矮化绿竹可使叶片叶绿素质量分数增加, 光能利用效率提高, 达到矮化栽培要求。竹笋截梢这种物理矮化方法避免直接带走大量秆枝叶, 操作相对简单, 且可以消除植物生长调节剂矮化药剂残留隐患, 在其他竹种矮化上具有借鉴意义。本研究对绿竹高生长结束后成竹形态特征与叶绿素质量分数及叶绿素荧光参数进行初步研究, 其更深层次光合机制还需进一步完善。此外, 竹笋截梢后对绿竹笋产量、出笋时期及竹材力学性质等影响还有待深入研究。
Effects of bamboo shoot truncation on growth and chlorophyll fluorescence characteristics of Dendrocalamopsis oldhami
-
摘要:
目的 探讨绿竹Dendrocalamopsis oldhami矮化方法及矮化后绿竹生长规律及光合机制具有重要意义。 方法 以福建省尤溪县出笋盛期绿竹笋为研究对象,不截梢为对照(ck),比较不同高度H1(60 cm)、H2(90 cm)、H3(120 cm)、H4(150 cm)、H5(180 cm)绿竹笋截梢对绿竹形态指标、成竹率、叶绿素质量分数及叶绿素荧光参数的影响。 结果 ① 竹笋截梢后绿竹形态发生显著变化,株高与截梢前绿竹笋高度呈线性极显著负相关(R2=0.90,P < 0.01),其中:H5处理绿竹株高由对照的539.40 cm降低至234.70 cm,较对照降低了56.49%。随着绿竹株高降低,枝下高、节数及分枝率减少,主枝长度增加。竹笋截梢后绿竹成竹率为82.14%~85.71%,成竹率未达显著差异(P>0.05)。②竹笋截梢显著提高绿竹叶绿素a、叶绿素b及总叶绿素,降低了叶绿素a/b。③竹笋截梢处理初始荧光(Fo)与对照无显著差异。随着绿竹株高降低,PSⅡ最大光化学效率(Fv/Fm)和光化学猝灭系数(qP)增加,而非光化学猝灭系数(qNP)呈下降趋势。电子传递速率(ETR)和PSⅡ实际光量子效率(Yield)增加,在H5时达最大值,较对照分别提高了48.63%和42.17%。④Pearson相关性分析表明:绿竹株高与总叶绿素总量及叶绿素荧光参数Fv/Fm、Yield、ETR显著负相关,与叶绿素a/b及qNP显著正相关。 结论 绿竹笋高度H5(180 cm)时截梢处理可有效控制高度生长,矮化后光化学功能进一步提高。 Abstract:Objective The aim is to explore the dwarf method and clarify preliminarily the growth and photosynthetic mechanism of Dendrocalamus oldhami, effects of bamboo shoot truncation on morphology, survival rate, chlorophyll content and chlorophyll fluorescence parameters of D. oldhami. Method Different heights H1(60 cm), H2(90 cm), H3(120 cm), H4(150 cm), H5(180 cm) of bamboo shoots at peak period in Youxi County, Fujian Province were used as test materials and bamboo shoots without truncation as the control group. Result (1) Morphological characteristics changed significantly after bamboo shoot truncation treatment, and there was a significant negative correlation between plant height and bamboo shoot height before truncation (R2=0.90, P < 0.01). Plant height of H5 after treatment was 539.40 cm and that of the control group was 234.70 cm, reducing by 56.49%. With bamboo shoot truncation treatment, the under branch height, internode number and branch rate decreased significantly while the length of the main branch increased. Survival rate of D. oldhami ranged from 82.14% to 85.71%, and no significant difference in survival rate was observed (P>0.05).(2) The chlorophyll a, chlorophyll b and total chlorophyll of D. oldhami increased while chlorophyll a/b decreased. (3) There was no significant difference in initial fluorescence (Fo) between the experiment groups and the control group. With the decrease of plant height, the PS Ⅱ maximal photochemical efficiency(Fv/Fm) and photochemical quenching coefficient (qP) enhanced, while the nonphotochemical quenching coefficient (qNP) reduced. The maximum values of electron transport rate (ETR) and PS Ⅱ actual photochemical efficiency(Yield) were achieved under H5 treatment, increasing by 48.63% and 42.17% respectively compared with the control group. (4) Pearson correlation analysis showed that plant height was negatively correlated with total chlorophyll and chlorophyll fluorescence parameters Fv/Fm, Yield and ETR, and positively correlated with chlorophyll a/b and qNP. Conclusion It could be concluded that 180 cm was the optimal bamboo shoot truncation treatment, which helped control plant height and improve photochemical effect. -
Key words:
- botany /
- Dendrocalamopsis oldhami /
- dwarf /
- bamboo shoot truncation /
- morphology /
- chlorophyll fluorescence
-
图 1 不同竹笋截梢处理下绿竹的株高和成竹率
Figure 1 Plant height and survival rate of D.oldhami under different bamboo shoot truncation treatments
图 2 不同竹笋截梢处理下绿竹叶绿素荧光参数变化
Figure 2 Chlorophyll fluorescence parameters changes of D. oldhami under different bamboo shoot truncation treatments
表 1 绿竹不同高度竹笋截梢处理概况
Table 1. General situation among different height bamboo shoot truncation treatments of D.oldhami
处理 截梢前笋高/cm 截梢长度/cm 截梢后笋高/cm H1 60 12 48 H2 90 18 72 H3 120 24 96 H4 150 30 120 H5 180 36 144  下载: 导出CSV
下载: 导出CSV
表 2 不同竹笋截梢处理绿竹其他形态变化
Table 2. Morphological indexes changes of D.oldhami under different bamboo shoot truncation treatments
处理 枝下高/cm 节数 分枝率/% 主枝长度/cm ck 123.75±23.34 a 19.85±1.78 a 67.86±4.24 a 216.05±17.60 bc H1 119.05±20.75 ab 16.95±1.82 b 70.09±4.58 a 209.05±30.27 c H2 107.50±14.61 ab 15.95±0.83 b 67.95±4.74 a 219.25±20.82 bc H3 104.25±39.99 b 12.40±1.56 c 58.29±9.17 b 219.15±42.87 bc H4 110.45±18.65 ab 11.60±1.53 cd 54.95±7.50 b 239.95±9.83 a H5 78.35±21.03 c 10.80±3.20 d 59.63±11.71 b 234.30±42.06 ab 说明:同列不同小写字母表示不同竹笋截梢处理间差异显著(P<0.05)
下载: 导出CSV
表 3 不同竹笋截梢处理绿竹叶绿素质量分数及组成变化
Table 3. Changes of chlorophyll content and composition ratio of D. oldhami under different bamboo shoot truncation treatments
处理 叶绿素a/(mg·g-1) 叶绿素b/(mg·g-1) 总叶绿素/(mg·g-1) 叶绿素a/b ck 1.95±0.20 c 0.76±0.02 b 2.71±0.21 b 2.56±0.27 a H1 1.92±0.010 c 1.01±0.11 b 2.93±0.19 b 1.91±0.18 b H2 2.15±0.23 bc 1.79±0.24 a 3.94+0.38 a 1.21±0.17 d H3 2.28±0.22 bc 1.82+0.18 a 4.10+0.13 a 1.27±0.23 cd H4 2.45±0.54 ab 1.59±0.42 a 4.04±0.62 a 1.69±0.75 bcd H5 2.81±0.33 a 1.66±0.45 a 4.47±0.63 a 1.81±0.54 bc 说明:同列不同小写字母表示不同竹笋截梢处理间差异显著(P<0.05)
下载: 导出CSV
表 4 绿竹株高与叶绿素质量分数及叶绿素荧光参数相关性分析
Table 4. Correlation analysis of plant height, chlorophyll content, chlorophyll fluorescence parameters of D. oldhami
指标 株高 总叶绿素 叶绿素a/b Fo Fv/Fm Yield ETR qP 总叶绿素 -0.809** 叶绿素a/b 0.368* -0.563** Fo -0.339 0.197 -0.048 FV/Fm -0.536* 0.410* -0.284 0.140 Yield -0.574* 0.372* -0.021 -0.150 0.169 ETR -0.607* 0.398* -0.068 -0.141 0.192 0.990** qP -0.697* 0.435* -0.030 0.044 0.177 0.895** 0.895** qNP 0.704** -0.391* -0.281 -0.206 -0.268 -0.637** -0.640** -0.719** 说明:*表示显著相关(P<0.05), **表示极显著相关(P<0.01).株高、叶绿素质量分数和叶绿素荧光参数样本n=25
下载: 导出CSV
-
[1] 刘平, 杨慧, 孟雪, 等.植物矮化研究进展[J].安徽农业科学, 2010, 38(27):15442-15443. LIU Ping, YANG Hui, MENG Xue, et al. Research advance of plant dwarfing[J]. J Anhui Agric Sci, 2010, 38(27):15442-15443. [2] 张帆, 万雪琴, 朱小琼, 等.自然低温对慈竹和撑绿杂交竹生理特性的影响[J].浙江农林大学学报, 2012, 29(1):17-22. ZHANG Fan, WAN Xueqin, ZHU Xiaoqiong, et al. Physiological characteristics of Neosinocalamus affinis and Bambusa pervariabilis×Dendrocalamopsis grandis with natural low temperature[J]. J Zhejiang A&F Univ, 2012, 29(1):17-22. [3] 周建革, 范明香, 余水生, 等.矮壮素对毛竹的矮化效应研究[J].世界竹藤通讯, 2012, 10(4):20-22. ZHOU Jiange, FAN Mingxiang, YU Shuisheng, et al. A study of dwarfing effect of chlormequat on moso[J]. World Bamboo Rattan, 2012, 10(4):20-22. [4] 罗龙发.新罗区毛竹不同强度钩梢试验[J].福建林业科技, 2016, 43(3):165-169. LUO Longfa. Obtruncation experiment of Phyllostachys edulis by different intensity in Xinluo[J]. J Fujian For Sci Technol, 2016, 43(3):165-169. [5] 官凤英, 范少辉, 刘碧桃.笋用绿竹设施栽培高生长控制技术研究[J].湖北农业科学, 2010, 49(12):3095-3098. GUAN Fengying, FAN Shaohui, LIU Bitao. Study on the control techniques of height growth for shoot-producing Dendrocalamopsis oldhami under facility cultivation[J]. Hubei Agric Sci, 2010, 49(12):3095-3098. [6] 高贵宾, 顾小平, 吴晓丽, 等.设施栽培绿竹笋用林的出笋规律[J].南京林业大学学报(自然科学版), 2009, 33(2):27-30. GAO Guibin, GU Xiaoping, WU Xiaoli, et al. Study on shoot emergence of Bambusa oldhamii in greenhouse[J]. J Nanjing For Univ Nat Sci Ed, 2009, 33(2):27-30. [7] 赵海燕, 刘桂华, 岳祥华.两种植物生长延缓剂对紫竹的矮化效应[J].江西农业大学学报, 2013, 35(4):786-790. ZHAO Haiyan, LIU Guihua, YUE Xianghua. Dwarfing effect of two retardants on Phyllostachys nigra[J]. Acta Agric Univ Jiangxi, 2013, 35(4):786-790. [8] 李冬林, 孙戴妍, 孙威, 等.笋用竹林丰产经营关键技术与发展建议[J].世界竹藤通讯, 2018, 16(5):25-28. LI Donglin, SUN Daiyan, SUN Wei, et al. Key techniques for high yield cultivation of shoot bamboo forest and their development suggestions[J]. World Bamboo Rattan, 2018, 16(5):25-28. [9] 唐大超, 唐晓东.高效培育麻竹笋用林技术研究[J].世界竹藤通讯, 2008, 6(6):21-24. TANG Dachao, TANG Xiaodong. A study of high effective cultivation technology for Dendrocalamus latiflorus shoot stand[J]. World Bamboo Rattan, 2008, 6(6):21-24. [10] 童龙, 李彬, 陈丽洁, 等.钩梢强度对麻竹生物量分配和生理特性的影响[J].应用生态学报, 2016, 27(10):3137-3144. TONG Long, LI Bin, CHEN Lijie, et al. Effects of truncation intensity on the biomass allocation and physiological characteristics of Dendrocalamus latiflorus[J]. Chin J Appl Ecol, 2016, 27(10):3137-3144. [11] 董敦义, 李子川, 桂仁意, 等.夏季与秋季钩梢对5年生毛竹竹材物理力学性质的影响[J].江西农业大学学报, 2015, 37(2):225-230. DONG Dunyi, LI Zichuan, GUI Renyi, et al. Influence of obtruncation in summer and autumn on physical and mechanical properties of 5 years old culms of Phyllostachys pubescens[J]. Acta Agric Univ Jiangxi, 2015, 37(2):225-230. [12] 刘碧桃, 官凤英, 范少辉, 等.两种生长延缓剂对绿竹矮化作用的研究[J].西北林学院学报, 2011, 26(4):142-147. LIU Bitao, GUAN Fengying, FAN Shaohui, et al. Dwarf effect of two retardants on Dendrocalamopsis oldhami[J]. J Northwest For Univ, 2011, 26(4):142-147. [13] 叶淑贤, 陆媛媛, 朱文强, 等.断鞭对毛竹竹笋-幼竹高生长的影响[J].西北林学院学报, 2013, 28(1):100-103. YE Shuxian, LU Yuanyuan, ZHU Wenqiang, et al. Effects of cutting rhizome on the height growth of shoots and seedings of Phyllostachys edulis[J]. J Northwest For Univ, 2013, 28(1):100-103. [14] 商继东, 李川, 孙杰, 等.竹箨剥除对毛竹竹笋及幼竹高生长的影响[J].东北林业大学学报, 2013, 41(11):10-12. SHANG Jidong, LI Chuan, SUN Jie, et al. Effects of stripping sheath on bamboo shoot and height growth of Phyllostachys edulis[J]. J Northeast For Univ, 2013, 41(11):10-12. [15] 刘国华, 王福升.修剪对6种地被竹高生长的影响[J].南京林业大学学报(自然科学版), 2008, 32(2):145-147. LIU Guohua, WANG Fusheng. Height growth regularity of six dwarf bamboo's culm[J]. J Nanjing For Univ Nat Sci Ed, 2008, 32(2):145-147. [16] 郑蓉.绿竹遗传多样性与优良地理种源选择的研究[D].北京: 北京林业大学, 2008. ZHENG Rong. Genetic Diversity and Selection of Super Geographical Provenances of Dendrocalamopsis oldhami[D]. Beijing: Beijing Forestry University, 2008. [17] 黄华宏, 陈奋学, 童再康.矮生杉木光合特性及叶绿素荧光参数研究[J].北京林业大学学报, 2009, 31(2):69-73. HUANG Huahong, CHEN Fenxue, TONG Zaikang. Photosynthetic properties and chlorophyll florescence parameters of dwarf Chinese fir[J]. J Beijing For Univ, 2009, 31(2):69-73. [18] 习玉森, 姜卫兵, 文杨, 等.不同生长型桃树光合效能及其季节变化特征初探[J].西北植物学报, 2016, 36(9):1836-1845. XI Yusen, JIANG Weibing, WEN Yang, et al. Photosynthetic efficiency and its seasonal change of peach trees with different growth types[J]. Acta Bot Boreali-Occident Sin, 2016, 36(9):1836-1845. [19] 罗静, 易盼盼, 王飞, 等.不同矮化中间砧对苹果苗光合特性的影响[J].西北农林科技大学学报(自然科学版), 2016, 44(4):177-184. LUO Jing, YI Panpan, WANG Fei, et al. Effects of different dwarf interstocks on photosynthetic characteristics of grafted apple trees[J]. J Northwest A&F Univ Nat Sci Ed, 2016, 44(4):177-184. [20] 张嘉桐, 关颖慧, 司莉青, 等. Pb2+、Cd2+复合胁迫对桑树光合作用的影响[J].北京林业大学学报, 2018, 40(4):16-23. ZHANG Jiatong, GUAN Yinghui, SI Liqing, et al. Effects of Pb2+ and Cd2+ combined stress on photosynthesis of Morus alba[J]. J Beijing For Univ, 2018, 40(4):16-23. [21] 宋莉英, 孙兰兰, 舒展, 等.干旱和复水对入侵植物三裂叶蟛蜞菊叶片叶绿素荧光特性的影响[J].生态学报, 2009, 29(7):3713-3721. SONG Liying, SUN Lanlan, SHU Zhan, et al. Effects of drought stress and rehydration on chlorophyll fluorescence characteristics in leaves of invasive Wedelia trilobata[J]. Acta Ecol Sin, 2009, 29(7):3713-3721. [22] 李霞, 郭振华.麻竹节间伸长过程的初步研究[J].植物分类与资源学报, 2014, 36(1):22-28. LI Xia, GUO Zhenhua. A pilot study on internode elongation in a paleotropical bamboo, Dendrocalamus latiflorus(Poaceae:Bambusoideae)[J]. Plant Diversity Resour, 2014, 36(1):22-28. [23] 曲良谱.金明竹在泰州地区发笋退笋规律研究[J].竹子研究汇刊, 2017, 36(4):37-40. QU Liangpu. The shooting and decay of Phyllostachys bambusoides f. castillonis in Taizhou area[J]. J Bamboo Res, 2017, 36(4):37-40. [24] 郑郁善, 洪伟, 邱尔发.毛竹出笋退笋规律的研究[J].林业科学, 1998, 34(专刊1):73-77. ZHENG Yushan, HONG Wei, QIU Erfa. Study on the regulation of shoot emergence and degradation of Phyllostachys heterocycla cv. pubescens shoot[J]. Sci Silv Sin, 1998, 34(spec 1):73-77. [25] 岳祥华, 赵海燕, 何小定, 等.紫竹在黄山地区发笋成竹规律的研究[J].安徽农业大学学报, 2012, 39(3):377-380. YUE Xianghua, ZHAO Haiyan, HE Xiaoding, et al. Study on shooting and the height growth rhythm of Phyllostachys nigra in Huangshan area[J]. J Anhui Agric Univ, 2012, 39(3):377-380. [26] BOND W J, MIDGLEY J J. Ecology of sprouting in woody plants:the persistence niche[J]. Trends Ecol Evol, 2001, 16(1):45-51. [27] 王震, 张利文, 虞毅, 等.平茬高度对四合木生长及生理特性的影响[J].生态学报, 2013, 33(22):7078-7087. WANG Zhen, ZHANG Liwen, YU Yi, et al. Effect of different stubble height treatments on the annual growth index and physiological characteristics of Tetraena mongolica in two growing seasons[J]. Acta Ecol Sin, 2013, 33(22):7078-7087. [28] ALCORN P J, FORRESTER D I, THOMAS D S, et al. Changes in whole-tree water use following live-crown pruning in young plantation-grown Eucalyptus pilularis and Eucalyptus cloeziana[J]. Forests, 2013, 4(1):106-121. [29] 郑士光, 贾黎明, 庞琪伟, 等.平茬对柠条林地根系数量和分布的影响[J].北京林业大学学报, 2010, 32(3):64-69. ZHENG Shiguang, JIA Liming, PANG Qiwei, et al. Stumping effects on number and distribution of roots of Caragana microphylla Lam. plantations[J]. J Beijing For Univ, 2010, 32(3):64-69. [30] 尚富华, 李吉跃, 胡磊, 等.修枝对毛白杨无性系生长、净光合速率和蒸腾速率的影响[J].中国农学通报, 2010, 26(23):134-139. SHANG Fuhua, LI Jiyue, HU Lei, et al. Effects of pruning on growth, net photosynthesis rate and transpiration rate of Populus tomentosa in plantation[J]. Chin Agric Sci Bull, 2010, 26(23):134-139. [31] 张杰, 邹学忠, 杨传平, 等.不同蒙古栎种源的叶绿素荧光特性[J].东北林业大学学报, 2005, 33(3):20-21. ZHANG Jie, ZOU Xuezhong, YANG Chuanping, et al. The chlorophy fluorescence characteristic of Quercusm ongolica from different provenances[J]. J Northeast For Univ, 2005, 33(3):20-21. [32] 向芬, 李维, 刘红艳, 等.氮素水平对不同品种茶树光合及叶绿素荧光特性的影响[J].西北植物学报, 2018, 38(6):1138-1145. XIANG Fen, LI Wei, LIU Hongyan, et al. Characteristics of photosynthetic and chlorophyll fluorescence of tea varieties under different nitrogen application levels[J]. Acta Bot Boreali-Occident Sin, 2018, 38(6):1138-1145. [33] ZHAO Xinhua, DU Qi, ZHAO Yue, et al. Effects of different potassium stress on leaf photosynthesis and chlorophyll fluorescence in maize (Zea mays L.) at seedling stage[J]. Agric Sci, 2016, 7(1):44-53. [34] 魏晓东, 陈国祥, 施大伟, 等.干旱胁迫对银杏叶片光合系统Ⅱ荧光特性的影响[J].生态学报, 2012, 32(23):7492-7500. WEI Xiaodong, CHEN Guoxiang, SHI Dawei, et al. Effects of drought on fluorescence characteristics of photosystemⅡ in leaves of Ginkgo biloba[J]. Acta Ecol Sin, 2012, 32(23):7492-7500. [35] 周哲宇, 徐超, 胡策, 等.毛竹快速生长期的叶绿素荧光参数特征[J].浙江农林大学学报, 2018, 35(1):75-80. ZHOU Zheyu, XU Chao, HU Ce, et al. Chlorophyll fluorescence characteristics of Phyllostachys edulis during its fast growth period[J]. J Zhejiang A&F Univ, 2018, 35(1):75-80. [36] 冯露, 吴际洋, 鞠易倩, 等.紫叶紫薇呈色生理及光合特性研究[J].北京林业大学学报, 2017, 39(12):93-101. FENG Lu, WU Jiyang, JU Yiqian, et al. Leaf color characteristics and photosynthetic characteristics of purple-leafed Lagerstroemia indica 'Ebony Ember'[J]. J Beijing For Univ, 2017, 39(12):93-101. [37] 陈洪国.植物生长调节剂对菊花幼苗生长及光合作用的影响[J].安徽农业科学, 2006, 34(9):1852-1854. CHEN Hongguo. Effect of plant growth regulator on growth and photosynthesis of young Chrysanthemum morifolium[J]. J Anhui Agric Sci, 2006, 34(9):1852-1854. [38] 魏亚娟, 刘宗奇, 汪季, 等.植物生长调节剂对榆叶梅生长及叶绿素荧光参数的调控效应[J].西北农林科技大学学报(自然科学版), 2019, 47(3):1-9. WEI Yajuan, LIU Zongqi, WANG Ji, et al. Effect of plant growth regulator on growth and chlorophyll fluorescence parameters of Prunus triloba[J]. J Northwest A&F Univ Nat Sci Ed, 2019, 47(3):1-9. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.01.007
 点击查看大图
点击查看大图
计量
- 文章访问数: 2697
- HTML全文浏览量: 761
- PDF下载量: 50
- 被引次数: 0
