




-
林分结构综合反映了林分发展的过程,也是森林未来发展的驱动源泉,已成为分析和管理森林生态系统的重要因子[1]。林分结构包括非空间结构和空间结构,非空间结构用来描述林分平均特征,不受林分相对位置的约束,而空间结构是林木在林地上的分布格局及其属性在空间上的排列方式[2],空间结构在很大程度上决定了林分的稳定性、发展的可能性和经营空间的大小[3],对森林的演替趋势具有决定性作用[4],因此,空间结构已成为森林结构研究的热点[5]。分析森林群落的空间结构特征,有助于科学制定合理的经营措施。芦芽山是汾河和桑干河的发源地,是中国暖温带残存的天然次生林分布区中保存较完整的地区之一,存在大面积完好、典型的以华北落叶松Larix principis-rupprechtii和云杉Picea spp.为建群种组成的属于演替后期相对稳定的群落[6]。前人针对该林分已做了许多研究,其内容主要集中在群落的种间联结性[7]、林内土壤温度及含水量的季节差异[8]、树木径向生长[9-10]及其对气候变化的响应[11-12]等,对其林分空间结构的研究仍鲜见报道,因此,本研究以该地华北落叶松-云杉天然次生林为对象,通过计算其混交度、角尺度、大小比数,以及林分空间结构指数研究不同海拔该林分的结构特征,旨在为该地华北落叶松-云杉天然次生林的科学经营提供依据,同时为相似立地条件下人工林空间结构的优化经营奠定理论基础。
-
研究地位于山西省忻州市宁武县境内的芦芽山国家级自然保护区内(38°36′~39°02′N,111°46′~112°54′E),保护区面积21 453 hm2,海拔1 346~2 787 m。该地属暖温带半湿润区,具明显的大陆性气候特点,夏季凉爽多雨,冬季寒冷干燥。据距其最近的五寨气象站1957−2007年资料,其年平均气温为4.9 ℃,1月平均气温为−10.2 ℃,7月平均气温为22.4 ℃,年降水量472.7 mm,年蒸发量1 800 mm,年均相对湿度55%,无霜期130 d。土壤主要是在残积和坡积母质上发育起来的山地褐土和山地淋溶褐土等。海拔1 850~2 650 m是以华北落叶松和云杉属Picea植物白扦Picea meyeri、青扦Picea wilsonii为建群种组成的寒温性针叶林占优势的稳定森林群落。
-
在芦芽山国家级自然保护区阴坡海拔1950~2550 m范围内选取起源相同、林龄相近,未经人为扰动的华北落叶松-云杉天然次生林,按1 950、2 050、2 150、2 350和2 550 m海拔梯度布设样地,每个海拔4个样地,样地面积为20 m×20 m。每个样地内,对胸径5 cm以上的活树进行每木检尺,记录树种、胸径、树高、枝下高、冠幅,并对每株树进行定位。同时在每个海拔的样地内选取主要优势树种的3株优势木,采用生长锥在树干基部钻取髓心,根据3株优势木年龄的均值确定林龄。样地基本概况见表1。
表 1 样地基本概况
Table 1. Survey of sample plots
海拔/m 坡度/(°) 坡位 林龄/a 林分密度/(株·hm−2) 平均树高/m 平均胸径/cm 郁闭度 1 950 29 中下部 105 950 13.0 22.91 0.70 2 050 23 中下部 115 1200 17.5 25.27 0.65 2 150 30 中部 100 1600 12.2 19.53 0.70 2 350 22 中下部 120 1425 16.5 21.98 0.75 2 550 15 中部 115 1952 13.4 21.45 0.75 -
角尺度
${W_i} = \dfrac{{\sum\limits_{j = 1}^n {{Z_{i\!j}}} }}{n}$ 。其中:Wi为第i株参照树的角尺度;α为参照树与2个最近相邻木夹角的较小角,αj为第j个α角,α0为标准角(72°),当αj<α0时,Zij=1,当αj>α0时,Zij=0;n为参照树的相邻木株数,本研究n=4。Wi反映了林木的水平分布格局,取值为0、0.25、0.50、0.75或1.00,代表林木处于绝对均匀、均匀、随机、不均匀或绝对不均匀状态。林分随机分布的角尺度为[0.475,0.517],角尺度小于0.475的分布为均匀分布,大于0.517的分布则为聚集分布。大小比数
${U_i} = \dfrac{{\sum\limits_{j = 1}^n {{K_{i\!j}}} }}{n}$ 。其中:Ui为第i株参照树的大小比数;参照树i胸径小于第j株相邻木胸径时,Kij=1,参照树i胸径大于第j株相邻木胸径时,Kij=0。Ui反映了林木个体间的大小分化程度,取值为0、0.25、0.50、0.75或1.00,代表林木大小分化程度分别为优势、亚优势、中庸、劣势或绝对劣势。混交度
${{M_i}} = \dfrac{{\sum\limits_{j = 1}^n {{m_{i\!j}}} }}{n}$ 。其中:Mi为第i株参照树的混交度;参照树i与第j株相邻木为同种时,mij=0,参照树i与第j株相邻木非同种时,mij=1。Mi反映了树种间隔离程度,取值为0、0.25、0.50、0.75或1.00,代表混交程度分别为零度、弱度、中度、强度或极强度混交。林分空间结构指数
${I_{{\rm{FSS}}}} = \left\{\! \!{\begin{array}{*{20}{l}}{{{[\overline M(100 - \overline U) \times 2\overline W]}^{0.333\;3}},\overline W \leqslant50}\\{{{[\overline M(100 - \overline U) \times 2(100 - \overline W)]}^{0.333\;3}},\quad \overline W > 50}\end{array}} \right.$ 。其中:0≤IFSS≤100,0≤$\overline M $ ≤100,0≤$\overline U $ ≤100,0≤$\overline W $ ≤100。$\overline M = 100\left( {\dfrac{{\sum\limits_{i=1}^{N} {{M_i}} }}{N}} \right)$ ,$\overline W = 100\left( {\dfrac{{\sum\limits_{i=1}^{N} {{W_i}} }}{N}} \right)$ ,$\overline U = 100\left( {\dfrac{{\sum\limits_{i=1}^{N} {{U_i}} }}{N}} \right)$ ,其中:N为样地树木株数。 -
芦芽山阴坡不同海拔华北落叶松-云杉天然次生林按相对多度筛选出群落优势度较高的物种仅为华北落叶松和云杉,因此在针对树种进行空间结构计算时,仅针对优势树种进行。群落及各树种平均角尺度及其参数分布见表2,仅海拔2 150 m处群落的平均角尺度为0.51,其余海拔处林分的平均角尺度为0.52~0.53,可见,华北落叶松-云杉天然次生林几乎均为聚集分布型,但聚集程度较低。华北落叶松-云杉天然次生林群落的角尺度整体近似服从正态分布,不同海拔间未表现出明显差异。5个海拔群落中Wi=0的频率很低,甚至不存在,说明该林分中林木很均匀分布的情况极少,而Wi=0.5的频率均为最高,占比均在40%以上,说明该天然林分中林木大多处于随机分布状态,Wi=0.75与Wi=1.00角尺度的频率之和稍大于Wi=0与Wi=0.25角尺度的频率之和,说明不均匀分布的状态稍多于均匀分布的状态。
表 2 不同海拔树种和群落角尺度特征
Table 2. Species angle index of L. principis-rupprechtii-Picea spp. secondary forests
海拔/m 树种 不同角尺度Wi下的分布频率 平均值 0 0.25 0.50 0.75 1.00 1 950 华北落叶松 0 0 0.60 0.2 0.20 0.65 云杉 0 0.36 0.36 0.21 0.07 0.50 群落 0 0.28 0.41 0.21 0.10 0.53 2 050 华北落叶松 0 0.12 0.56 0.20 0.12 0.58 云杉 0 0.25 0.75 0 0 0.44 群落 0 0.17 0.61 0.14 0.08 0.53 2 150 华北落叶松 0.02 0.20 0.53 0.23 0.02 0.51 云杉 0 0.17 0.50 0.28 0.06 0.56 群落 0.02 0.21 0.51 0.24 0.02 0.51 2 350 华北落叶松 0 0.29 0.57 0.14 0 0.46 云杉 0 0.19 0.51 0.26 0.04 0.54 群落 0 0.22 0.50 0.24 0.04 0.53 2 550 华北落叶松 0 0 1.00 0 0 0.50 云杉 0.02 0.18 0.51 0.27 0.02 0.52 群落 0.02 0.17 0.53 0.26 0.02 0.52 不同海拔的2种优势树种的角尺度特征表现不尽相同,华北落叶松的角尺度为0.46~0.65,云杉的角尺度为0.44~0.56,华北落叶松在2 350 m处表现为均匀分布,低海拔以聚集分布为主,高海拔以随机分布为主;云杉则在2 050 m处呈均匀分布,低海拔以随机分布为主,高海拔以聚集分布为主。随海拔的升高,华北落叶松分布株数呈先增加后减少的趋势,云杉则呈先减少后增加的趋势,且华北落叶松和云杉的株数分别在海拔2 150和2 550 m达到最大,在其株数较多的海拔均呈聚集分布。2个树种的角尺度均在Wi=0.50时的分布频率最大,优势树种在林分中都属于随机分布。
-
芦芽山阴坡华北落叶松-云杉天然次生林各海拔群落平均大小比数为0.47~0.52,整体均接近中庸状态,说明林内林木胸径的差异不明显,大小分化不严重,且不同海拔群落各大小比数所占的频率相差不大,约20%(表3)。
表 3 不同海拔树种的大小比数特征
Table 3. Species neighborhood comparison of L. principis-rupprechtii-Picea spp. secondary forests
海拔/m 树种 不同大小比数Ui下的分布频率 Ui平均值 0 0.25 0.50 0.75 1.00 1 950 华北落叶松 0.60 0.20 0 0.20 0 0.20 云杉 0.14 0.43 0.29 0.14 0 0.36 群落 0.17 0.28 0.17 0.24 0.14 0.47 2 050 华北落叶松 0.32 0.20 0.20 0.24 0.04 0.37 云杉 0 0.13 0.25 0.25 0.38 0.72 群落 0.22 0.17 0.19 0.25 0.17 0.49 2 150 华北落叶松 0.20 0.28 0.12 0.22 0.18 0.48 云杉 0.06 0.22 0.22 0.22 0.28 0.61 群落 0.15 0.26 0.15 0.21 0.22 0.52 2 350 华北落叶松 0.57 0.29 0 0 0.14 0.21 云杉 0.13 0.21 0.17 0.28 0.21 0.56 群落 0.19 0.22 0.15 0.24 0.20 0.51 2 550 华北落叶松 1.00 0 0 0 0 0 云杉 0.20 0.20 0.17 0.26 0.18 0.50 群落 0.23 0.19 0.16 0.24 0.17 0.48 2种优势树种大小比数在不同海拔表现不同。随着海拔的升高,华北落叶松先由亚优势状态发展为中庸状态,后由中庸状态发展为优势状态;云杉则先由近亚优势状态发展为劣势状态,后发展为中庸状态,且2种优势树种均在其株数达到最大的海拔处表现为中庸状态。不同海拔华北落叶松在大小比数Ui=0或Ui=0.25时的分布频率均最大,云杉除在1 950 m时大小比数在Ui=0.25时的分布频率最大外,其余海拔均为Ui=0.75或Ui=1.00时分布频率最大。可见,华北落叶松在不同海拔的林分中均处于优势或亚优势状态,云杉处于劣势状态,华北落叶松的竞争力明显大于云杉。
-
随着海拔高度的上升,华北落叶松-云杉天然次生林群落的平均混交度呈下降趋势,可见随海拔升高混交程度由强变弱,树种的组成趋于单一化。群落的混交度频率分布规律在不同海拔间差异较大(表4),海拔1 950 m时,以中强度混交为主,中强度混交比例之和达到86%以上,海拔 2 050 m以上,以零度和弱度混交为主。零度和弱度混交比例之和达到60%以上,且混交度Mi=0的频率随海拔升高逐渐增加,可见优势树种随海拔增加单种聚集现象加剧,进而导致了高海拔处林分整体处于弱度混交。
表 4 不同海拔树种和群落的混交度特征
Table 4. Species mingling degree of L. principis-rupprechtii-Picea spp. secondary forests
海拔/m 树种 不同混交度Mi下的分布频率 Mi平均值 0 0.25 0.50 0.75 1.00 1 950 华北落叶松 0 0 0 0.40 0.60 0.90 云杉 0 0.29 0.50 0.14 0.07 0.50 群落 0 0.14 0.34 0.24 0.28 0.66 2 050 华北落叶松 0.20 0.52 0.20 0 0.08 0.31 云杉 0 0.63 0 0 0.38 0.53 群落 0.14 0.50 0.14 0 0.22 0.42 2 150 华北落叶松 0.55 0.22 0.12 0.10 0.02 0.20 云杉 0 0.11 0.22 0.61 0.06 0.65 群落 0.39 0.21 0.15 0.16 0.09 0.35 2 350 华北落叶松 0 0 0 0.43 0.57 0.89 云杉 0.45 0.45 0.11 0 0 0.16 群落 0.39 0.39 0.09 0.06 0.07 0.26 2 550 华北落叶松 0 0 0 0 1.00 1.00 云杉 0.82 0.17 0.01 0 0 0.05 群落 0.79 0.16 0.01 0 0.04 0.09 随着海拔的升高,华北落叶松的混交状态由强变弱,后又变强,云杉则除2 150 m海拔处为强度混交为主外,其余海拔均为中弱度混交为主。可能是随着海拔的升高,华北落叶松的数量先增加后减少,云杉则表现为先减少后增加,进而其数量在增加时的海拔处均表现出了明显的单种聚集现象,因而呈弱度混交状态。
-
林分空间结构指数能够很好地反映不同林分所处的状态。图1可看出:随着海拔的升高,芦芽山华北落叶松-云杉天然次生林的林分空间结构指数逐渐减小,可见低海拔林分的空间结构较好。
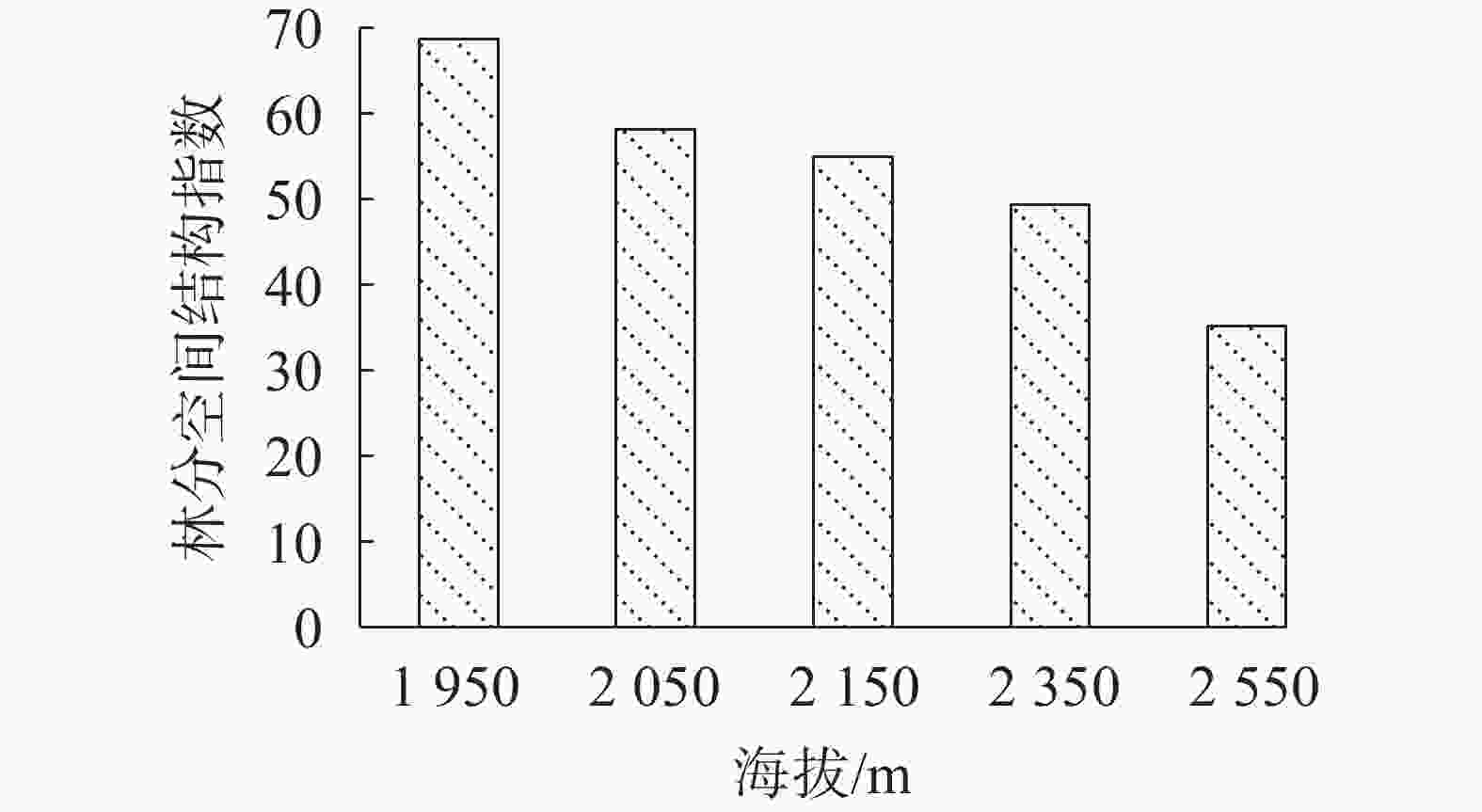
图 1 不同海拔的林分空间结构指数
Figure 1. Stand spatial structure index of L. principis-rupprechtii-Picea spp. secondary forests
-
芦芽山阴坡华北落叶松-云杉天然次生林群落的空间分布格局总体上以聚集分布为主,但聚集程度较低,接近随机分布。一般情况下,群落不受严重干扰,经过演替后形成的顶级群落的水平分布格局为随机分布[13-14],可见,芦芽山阴坡华北落叶松-云杉天然次生林正处在演替后期向顶级群落演替的阶段。各海拔的林木均以随机分布的占比最大,比例均大于40%,不均匀分布的比例大于均匀分布的比例,这与柏云龙等[15]及夏富才等[16]研究结果相同。本研究中优势树种华北落叶松和云杉虽在各海拔的空间分布格局不甚相同,角尺度范围为0.44~0.65,但均接近随机分布,进一步证实了该林分正处于向顶级群落类型演替的后期阶段,与赵鸣飞等[6]对芦芽山寒温性针叶林的研究结果一致。
各海拔群落大小分化程度差异不大,且整体均接近中庸状态。林木大小分化程度不大,这与肖欣等[17]和韩金等[18]研究结果相同。2个优势树种的角尺度均在Wi=0.50时分布频率最大,说明优势树种个体大多数呈随机分布的情况。华北落叶松的大小比数在所有海拔均小于云杉,表明该群落中华北落叶松的竞争力大于云杉,但在高海拔华北落叶松的取值仅在0上分布,原因是由于华北落叶松数量较少但胸径都很大,在结构单元中处于明显的优势地位。
华北落叶松-云杉天然次生林群落的平均混交度随海拔的升高呈下降趋势,结果与实际相符。该群落以华北落叶松和云杉为主要优势树种,低海拔时混生有少量的白桦Betula platyphylla、红桦B. albo-sinensis、辽东栎Quercus wutaishanica等树种,随着海拔的升高,混生树种逐渐减少,甚至消失,进而导致群落的混交度下降。
芦芽山阴坡华北落叶松-云杉天然次生林群落的角尺度、林木大小分化程度在不同海拔间均无显著性差异,而混交度随海拔升高呈明显的下降趋势,可见,该林分不同海拔空间结构的差异主要表现在混交度指数上。陶观护等[19]对不同密度油松Pinus tabulaeformis飞播林的林分空间结构的研究中表明,林分密度对混交度有着显著影响。芦芽山阴坡华北落叶松-云杉天然次生林不同海拔间林分空间结构的差异可能源于其林分密度的差异,2个优势树种在不同海拔间的密度变化导致其空间结构指数的差异也证实了这一观点。但是否存在其他因素(如坡度等)导致不同海拔间林分空间结构的差异仍需进行深入研究。
空间结构指数是反映林分空间结构综合情况的指标,数值越大,说明该林分的空间结构越优化。芦芽山阴坡华北落叶松-云杉天然次生林群落以海拔1 950 m时林分整体结构最好。一般认为,林分空间结构越优,林分的功能越强,稳定性越高[20]。故海拔1 950 m时的华北落叶松-云杉天然次生林的稳定性也最高。因此,在林分经营过程中,针对以混交度为明显差异造成的空间结构指数不同的华北落叶松-云杉天然次生林,可选择以1 950 m处的林分结构为借鉴对高海拔的林分进行结构调整,以提高林分稳定性。高海拔华北落叶松的胸径优势非常明显,占据着群落的最高层,但其株数比例较小,林下更新困难,因此在高海拔,需采取生态抚育措施对混交度为0和0.25的林木进行调整,改善混交比例,降低单种聚集现象,同时促进华北落叶松的更新,以保证物种多样性和群落的持续稳定发展。
Spatial structure of Larix principis-rupprechtii-Picea spp. secondary forests on shady slope of Luyashan National Nature Reserve
-
摘要:
目的 分析芦芽山国家级自然保护区阴坡典型天然次生林的林分空间结构,为当地天然次生林的科学经营和相似立地条件下人工林的培育提供参考依据。 方法 以芦芽山自然保护区阴坡5个不同海拔的华北落叶松Larix principis-rupprechtii-云杉Picea spp.天然次生林群落调查数据为基础,通过计算群落的角尺度、大小比数、混交度和林分空间结构指数,分析不同海拔群落的空间结构特征。 结果 华北落叶松-云杉天然次生林平均角尺度为0.51~0.53,林木空间分布格局以聚集分布为主,但聚集程度较低,优势树种在林分中属于随机分布;各海拔林木的大小分化程度整体均接近中庸状态,林木大小分化程度较小,华北落叶松的大小比数小于云杉,表明华北落叶松的竞争力较大;随海拔高度的上升,林木空间隔离程度呈下降趋势;林分空间结构指数随海拔升高而下降,低海拔林分的整体结构较其他海拔更好。 结论 研究区华北落叶松-云杉天然次生林群落均处于演替后期阶段,海拔1 950 m处林分的空间结构最好,稳定性最高。图1表4参20 -
关键词:
- 角尺度 /
- 混交度 /
- 大小比数 /
- 林分空间结构指数 /
- 华北落叶松-云杉天然次生林
Abstract:Objective The objective is to analyze the spatial structure of typical secondary forests on shady slope of Luyashan National Nature Reserve, and provide reference for scientific management of natural secondary forest and artificial forest in similar habitats. Method Based on the survey data of Larix principis-rupprechtii-Picea spp. secondary forest communities at five different altitudes on the shady slope of Luyashan National Nature Reserve, the spatial structures of the communities at different altitudes were analyzed by calculating the angle scale, size ratio, mingling degree and forest spatial structure index. Result The average angular scale was 0.51−0.53. The spatial distribution pattern of the forests was dominated by aggregation distributions, but the degree of aggregation was low. The dominant tree species were randomly distributed in the stand. The size differentiation degree of the trees at all altitudes was close to the mean state, and the size differentiation degree of L. principis-rupprechtii was smaller than that of Picea spp., indicating that L. principis-rupprechtii was more competitive. With the increase of altitude, the spatial isolation degree and the spatial structure index decreased. The overall structure of lower altitude stand was better than that of other altitudes. Conclusion The communities of L. principis-rupprechtii-Picea spp. secondary forests in the study area are in late state of succession. The stand at 1 950 m above sea level has the best spatial structure and stability. [Ch, 1 fig. 4 tab. 20 ref.] -
图 1 不同海拔的林分空间结构指数
Figure 1 Stand spatial structure index of L. principis-rupprechtii-Picea spp. secondary forests
表 1 样地基本概况
Table 1. Survey of sample plots
海拔/m 坡度/(°) 坡位 林龄/a 林分密度/(株·hm−2) 平均树高/m 平均胸径/cm 郁闭度 1 950 29 中下部 105 950 13.0 22.91 0.70 2 050 23 中下部 115 1200 17.5 25.27 0.65 2 150 30 中部 100 1600 12.2 19.53 0.70 2 350 22 中下部 120 1425 16.5 21.98 0.75 2 550 15 中部 115 1952 13.4 21.45 0.75  下载: 导出CSV
下载: 导出CSV
表 2 不同海拔树种和群落角尺度特征
Table 2. Species angle index of L. principis-rupprechtii-Picea spp. secondary forests
海拔/m 树种 不同角尺度Wi下的分布频率 平均值 0 0.25 0.50 0.75 1.00 1 950 华北落叶松 0 0 0.60 0.2 0.20 0.65 云杉 0 0.36 0.36 0.21 0.07 0.50 群落 0 0.28 0.41 0.21 0.10 0.53 2 050 华北落叶松 0 0.12 0.56 0.20 0.12 0.58 云杉 0 0.25 0.75 0 0 0.44 群落 0 0.17 0.61 0.14 0.08 0.53 2 150 华北落叶松 0.02 0.20 0.53 0.23 0.02 0.51 云杉 0 0.17 0.50 0.28 0.06 0.56 群落 0.02 0.21 0.51 0.24 0.02 0.51 2 350 华北落叶松 0 0.29 0.57 0.14 0 0.46 云杉 0 0.19 0.51 0.26 0.04 0.54 群落 0 0.22 0.50 0.24 0.04 0.53 2 550 华北落叶松 0 0 1.00 0 0 0.50 云杉 0.02 0.18 0.51 0.27 0.02 0.52 群落 0.02 0.17 0.53 0.26 0.02 0.52
下载: 导出CSV
表 3 不同海拔树种的大小比数特征
Table 3. Species neighborhood comparison of L. principis-rupprechtii-Picea spp. secondary forests
海拔/m 树种 不同大小比数Ui下的分布频率 Ui平均值 0 0.25 0.50 0.75 1.00 1 950 华北落叶松 0.60 0.20 0 0.20 0 0.20 云杉 0.14 0.43 0.29 0.14 0 0.36 群落 0.17 0.28 0.17 0.24 0.14 0.47 2 050 华北落叶松 0.32 0.20 0.20 0.24 0.04 0.37 云杉 0 0.13 0.25 0.25 0.38 0.72 群落 0.22 0.17 0.19 0.25 0.17 0.49 2 150 华北落叶松 0.20 0.28 0.12 0.22 0.18 0.48 云杉 0.06 0.22 0.22 0.22 0.28 0.61 群落 0.15 0.26 0.15 0.21 0.22 0.52 2 350 华北落叶松 0.57 0.29 0 0 0.14 0.21 云杉 0.13 0.21 0.17 0.28 0.21 0.56 群落 0.19 0.22 0.15 0.24 0.20 0.51 2 550 华北落叶松 1.00 0 0 0 0 0 云杉 0.20 0.20 0.17 0.26 0.18 0.50 群落 0.23 0.19 0.16 0.24 0.17 0.48
下载: 导出CSV
表 4 不同海拔树种和群落的混交度特征
Table 4. Species mingling degree of L. principis-rupprechtii-Picea spp. secondary forests
海拔/m 树种 不同混交度Mi下的分布频率 Mi平均值 0 0.25 0.50 0.75 1.00 1 950 华北落叶松 0 0 0 0.40 0.60 0.90 云杉 0 0.29 0.50 0.14 0.07 0.50 群落 0 0.14 0.34 0.24 0.28 0.66 2 050 华北落叶松 0.20 0.52 0.20 0 0.08 0.31 云杉 0 0.63 0 0 0.38 0.53 群落 0.14 0.50 0.14 0 0.22 0.42 2 150 华北落叶松 0.55 0.22 0.12 0.10 0.02 0.20 云杉 0 0.11 0.22 0.61 0.06 0.65 群落 0.39 0.21 0.15 0.16 0.09 0.35 2 350 华北落叶松 0 0 0 0.43 0.57 0.89 云杉 0.45 0.45 0.11 0 0 0.16 群落 0.39 0.39 0.09 0.06 0.07 0.26 2 550 华北落叶松 0 0 0 0 1.00 1.00 云杉 0.82 0.17 0.01 0 0 0.05 群落 0.79 0.16 0.01 0 0.04 0.09
下载: 导出CSV
-
[1] 张连金, 赖光辉, 封焕英, 等. 京西油松生态公益林空间结构评价[J]. 生态学杂志, 2018, 37(5): 1316 − 1325. ZHANG Lianjin, LAI Guanghui, FENG Huanying, et al. Evaluation of the stand spatial structure of Pinus tabuliformis ecological forest in West Beijing [J]. Chin J Ecol, 2018, 37(5): 1316 − 1325. [2] POMMERENING A. Approaches to quantifying forest structures [J]. Forestry, 2002, 75(3): 305 − 324. [3] 李语晨, 程金花, 李明峰, 等. 湖北省九华山林场不同树种配置杉阔混交林林分空间结构特征分析[J]. 生态学报, 2019, 39(6): 1908 − 1916. LI Yuchen, CHENG Jinhua, LI Mingfeng, et al. Analysis of the stand spatial structure of Cunninghamia lanceolata-broadleaved mixed plantation on a Jiuhua Mountain farm, Hubei Province [J]. Acta Ecol Sin, 2019, 39(6): 1908 − 1916. [4] 湛斌, 赵家豪, 陈斌, 等. 江西武夷山南方铁杉原始林空间结构特征分析[J]. 西北林学院学报, 2019, 34(4): 192 − 198. ZHAN Bin, ZHAO Jiahao, CHEN Bin, et al. Analysis of the spatial structure of Tsuga chinensis var. tchekiangensis virgin forest in Jiangxi Wuyishan National Nature Reserve [J]. J Northwest For Univ, 2019, 34(4): 192 − 198. [5] AGUIRRE O, HUI Gangying, GADOW K V, et al. An analysis of spatial forest structure using neighbourhood-based variables [J]. For Ecol Manage, 2003, 183(1/3): 137 − 145. [6] 赵鸣飞, 薛峰, 王宇航, 等. 山西芦芽山针叶林草本层群落谱系结构与多样性的海拔格局[J]. 植物生态学报, 2017, 41(7): 707 − 715. ZHAO Mingfei, XUE Feng, WANG Yuhang, et al. Phylogenetic structure and diversity of herbaceous communities in the conifer forests along an elevational gradient in Luya Mountain, Shanxi, China [J]. Chin J Plant Ecol, 2017, 41(7): 707 − 715. [7] 武秀娟. 芦芽山阴坡典型天然次生林群落的种间联结性[J]. 西北林学院学报, 2020, 35(1): 54 − 61. WU Xiujuan. Interspecific association of the plant communities in natural secondary forests on north slope of Luya Mountain [J]. J Northwest For Univ, 2020, 35(1): 54 − 61. [8] 薛峰, 江源, 王明昌, 等. 芦芽山针叶林分布上下限土壤温度及含水量的季节差异[J]. 生态学报, 2020, 40(1): 141 − 150. XUE Feng, JIANG Yuan, WANG Mingchang, et al. Seasonal changes in soil temperature and water content at the upper and lower limits of coniferous forest on Luya Mountain, China [J]. Acta Ecol Sin, 2020, 40(1): 141 − 150. [9] 董满宇, 江源, 王明昌, 等. 芦芽山林线华北落叶松径向变化季节特征[J]. 生态学报, 2012, 32(23): 7430 − 7439. DONG Manyu, JIANG Yuan, WANG Mingchang, et al. Seasonal variations in the stems of Larix principis-rupprechtii at the treeline of the Luya Mountain [J]. Acta Ecol Sin, 2012, 32(23): 7430 − 7439. [10] 董满宇, 江源, 杨浩春, 等. 芦芽山林线白杄生长季径向生长动态[J]. 植物生态学报, 2012, 36(9): 956 − 964. DONG Manyu, JIANG Yuan, YANG Haochun, et al. Dynamic of stem radial growth of Pica meyeri during the growing season at the treeline of Luya Mountain, China [J]. Chin J Plant Ecol, 2012, 36(9): 956 − 964. [11] 张文涛, 江源, 王明昌, 等. 芦芽山阳坡不同海拔白杄径向生长对气候变暖的响应[J]. 植物生态学报, 2013, 37(12): 1142 − 1152. ZHANG Wentao, JIANG Yuan, WANG Mingchang, et al. Responses of radial growth to climate warming in Picea meyeri trees growing at different elevations on the southern slope of Luya Mountain [J]. Chin J Plant Ecol, 2013, 37(12): 1142 − 1152. [12] 张文涛, 江源, 王明昌, 等. 芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应[J]. 生态学报, 2015, 35(19): 6481 − 6488. ZHANG Wentao, JIANG Yuan, WANG Mingchang, et al. Responses of radial growth in Larix principis-rupprechtii to climate change along an elevation gradient on the southern slope of Luya Mountain [J]. Acta Ecol Sin, 2015, 35(19): 6481 − 6488. [13] 王群, 张金池, 田月亮, 等. 浙江丹阳山天然混交林林分空间结构分析[J]. 浙江农林大学学报, 2012, 29(6): 875 − 882. WANG Qun, ZHANG Jinchi, TIAN Yueliang, et al. Stand spatial structure of a natural mixed forest in the Fengyang Mountains of Zhejiang [J]. J Zhejiang A&F Univ, 2012, 29(6): 875 − 882. [14] 张家城, 陈力, 郭泉水, 等. 演替顶级阶段森林群落优势树种分布的变动趋势研究[J]. 植物生态学报, 1999, 23(3): 256 − 268. ZHANG Jiacheng, CHEN Li, GUO Quanshui, et al. Research on the change trend of dominant tree population distribution patterns during development process of climax forest communities [J]. Acta Phytoecol Sin, 1999, 23(3): 256 − 268. [15] 柏云龙, 刘华, 白志强, 等. 天山云杉天然林群落空间结构异质性分析[J]. 植物科学学报, 2012, 30(6): 552 − 557. BAI Yunlong, LIU Hua, BAI Zhiqiang, et al. Spatial pattern heterogeneity of Picea schrenkiana var. tianschanica natural forest in the Tianshan Mountains, northwest China [J]. Plant Sci J, 2012, 30(6): 552 − 557. [16] 夏富才, 姚大地, 赵秀海, 等. 长白山北坡阔叶红松林空间结构[J]. 东北林业大学学报, 2009, 37(10): 5 − 7. XIA Fucai, YAO Dadi, ZHAO Xiuhai, et al. Horizontal spatial structure of Korean pine broad leaved forest in northern slope of Changbai Mountains [J]. J Northeast For Univ, 2009, 37(10): 5 − 7. [17] 肖欣, 丁松, 欧阳勋志. 井冈山国家级自然保护区典型常绿阔叶林林分空间结构分析[J]. 西北农林科技大学学报(自然科学版), 2016, 44(3): 133 − 140. XIAO Xin, DING Song, OUYANG Xunzhi. Stand spatial structure of typical evergreen broad-leaved forests in Jinggang Mountain National Nature Reserve [J]. J Northwest A&F Univ Nat Sci Ed, 2016, 44(3): 133 − 140. [18] 韩金, 王新杰, 段皓赟. 金沟岭云冷杉针阔混交林林分结构研究[J]. 中南林业科技大学学报, 2019, 39(1): 58 − 63. HAN Jin, WANG Xinjie, DUAN Haoyun. Study on forest structure of coniferous and broad-leaved mixed forest in Jingouling Forest Farm of Wangqing County of Jilin Province [J]. J Cent South Univ For Technol, 2019, 39(1): 58 − 63. [19] 陶观护, 卜元坤, 薛卫鹏, 等. 不同密度油松飞播林灌草多样性与林分空间结构关系[J]. 森林与环境学报, 2020, 40(2): 171 − 177. TAO Guanhu, BU Yuankun, XUE Weipeng, et al. Relationship between understory diversity and stand spatial structure in air-drilled Pinus tabulaeformis forests of different densities [J]. J For Environ, 2020, 40(2): 171 − 177. [20] 王志鸣, 郭秋菊, 艾训儒, 等. 恩施州不同海拔马尾松天然次生林林分结构特征研究[J]. 西南林业大学学报, 2019, 39(1): 114 − 122. WANG Zhiming, GUO Qiuju, AI Xunru, et al. Structure characteristics of Pinus massoniana natural secondary forest at different altitude of Enshi [J]. J Southwest For Univ, 2019, 39(1): 114 − 122. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200261
 点击查看大图
点击查看大图

计量
- 文章访问数: 2276
- HTML全文浏览量: 610
- PDF下载量: 46
- 被引次数: 0










