




-
氮、磷、钾是植物生长发育的必需营养元素。植物体内的氮、磷、钾含量分布及其生态化学计量特征能够反映植物对各器官营养元素的分配及相互作用关系,对判断限制性营养元素、养分利用情况等具有重要作用[1−3],是当前生态学研究的主要内容和热点[4−7]。桦木科Betulaceae铁木属Ostrya植物在全世界共有7种,其中中国自然分布的有4种,而浙江省境内仅见2种,即天目铁木Ostrya rehderiana和多脉铁木O. multinervis。天目铁木为国家二级保护树种,现有野生大树仅5株[8],生长于杭州市临安区天目山镇,多脉铁木仅分布于文成县石垟林场,野生株数118株[9],是浙江省重点保护野生植物。铁木属植物具有树体高大,干形通直圆满,材质优良的特点。国内学者已对天目铁木和多脉铁木的生育、繁殖、遗传、栽培等方面进行了一定的研究[10−17]。目前,国内外缺少有关天目铁木和多脉铁木这2种珍稀植物不同器官营养元素含量及生态化学计量差异的研究。掌握天目铁木与多脉铁木生长对元素的需求情况,可以提供针对性的施肥方案,有助于天目铁木与多脉铁木的正常生长与繁育。本研究对2种铁木属植物根系、枝条、叶片、树皮的氮磷钾质量分数以及根区土壤的基本理化性质进行研究,探究2种铁木属植物的生态化学计量特征与差异,旨在为这2种植物的生存繁衍提供适合的营养管理方案,为扩大其种群数量提出合理的措施。
-
天目铁木研究区位于杭州市临安区西天目山,中心位置为30°17′N,119°27′E。属北亚热带季风气候,年平均气温为14.8 ℃;最冷月为1月,月平均气温3.4 ℃;最热月为7月,月平均气温为28.1 ℃。年降水量1390.0 mm,年日照时数1550.0 h,无霜期235.0 d[18]。土壤类型属于凝灰岩发育的红壤土类。
多脉铁木研究区位于文成县石垟林场,分布中心地理位置为27°50′N,119°50′E。属中亚热带季风气候,年平均气温为12.8 ℃;最冷月为1月,月平均气温为4.0 ℃;最热月为7月,月平均气温为23.6 ℃。年降水量1604.0 mm,年均相对湿度84.0%,年日照时数1755.0 h,全年无霜期288.0 d[19]。该区域属原始次生常绿阔叶林,森林覆盖率达90%以上。多脉铁木群落分布区域的海拔为500~700 m,坡度30°~35°,南坡和坡中部地表有石块层覆盖,厚度20~30 cm,石块大小20~40 cm[20]。土壤类型为发育于花岗岩的红壤土类。2种铁木属植物根区土壤均为酸性土壤,其理化性质如表1。
表 1 天目铁木和多脉铁木根区土壤理化性质
Table 1. Basic physical and chemical properties of soil in O. rehderiana and O. multinervis woodlands
树种 pH 碱解氮/(mg·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) 有机碳/(g·kg−1) 天目铁木 6.26±0.03 a 166.4±2.67 a 5.25±0.38 a 158.5±1.48 a 24.1±0.23 A 多脉铁木 5.55±0.18 b 137.3±9.65 b 4.45±0.61 a 152.3±1.69 b 16.1±0.14 B 说明:表中同列小写字母表示不同树种土壤间差异显著(P<0.05),同列大写字母表示不同树种土壤间差异极显著(P<0.01) -
在研究区对天目铁木和多脉铁木群落展开调查,测量胸径和株高。选取地径和株高均为平均值的植株各3株[21],分别采集2种铁木属植物东、南、西、北带叶枝条各1根,包括当年生和多年生枝条,长度为(50.0±5.0) cm,枝径为(0.5±0.1) cm。随机均匀采集距离地面1.0 m高的2种植株树皮样品,采集后均匀混合。根系的取样以样本植株为中心,分别以0.5 m为半径,挖取深度为30.0 cm,宽度为50.0 cm的土壤剖面,采集剖面中的所有根系。
-
植物样品用去离子水洗净,置于烘箱中105 ℃杀青30 min,然后在80 ℃条件下烘干至恒量[22−23],取出,用高速粉碎机将样品粉碎,过0.149 mm筛后,分别装袋编号,待用。土壤有机质的测定采用重铬酸钾容量法,土壤碱解氮的测定采用碱解扩散法,土壤有效磷的测定采用钼蓝比色法,土壤速效钾的测定采用火焰光度计法;植物样品经过H2SO4-H2O2处理,全氮采用半微量开氏法,全磷采用钼蓝比色法,全钾采用火焰光度法。
-
数据均采用Excel 2016及SPSS 22.0软件整理,采用单因素方差分析的最小显著差异(LSD)法进行差异显著性检验及相关性分析。图表绘制采用Origin 2018软件处理。
-
由图1可见:氮、磷、钾在2种铁木属植株不同器官之间的差异较为显著。叶片中氮质量分数均为最高,显著高于根、枝和树皮(P<0.05)。2种铁木的枝、叶、树皮相同器官之间相比,氮质量分数并无显著性差异(P>0.05),仅多脉铁木的根氮质量分数显著高于天目铁木(P<0.05)。2种铁木的根、枝中磷质量分数最高,树皮中磷质量分数显著低于根、枝和叶(P<0.05)。但是2种铁木的根、枝、叶和树皮相同器官之间相比,磷质量分数并无显著性差异(P>0.05)。2种铁木的叶中钾质量分数最高,树皮中钾质量分数显著低于叶(P<0.05)。其中多脉铁木的根、枝、叶中钾质量分数都高于天目铁木,特别是枝和叶,钾质量分数都显著高于天目铁木的枝、叶(P<0.05)。但是多脉铁木的树皮中钾质量分数略低于天目铁木,但无显著性差异(P>0.05)。总体看来,多脉铁木植株中钾质量分数高于天目铁木。
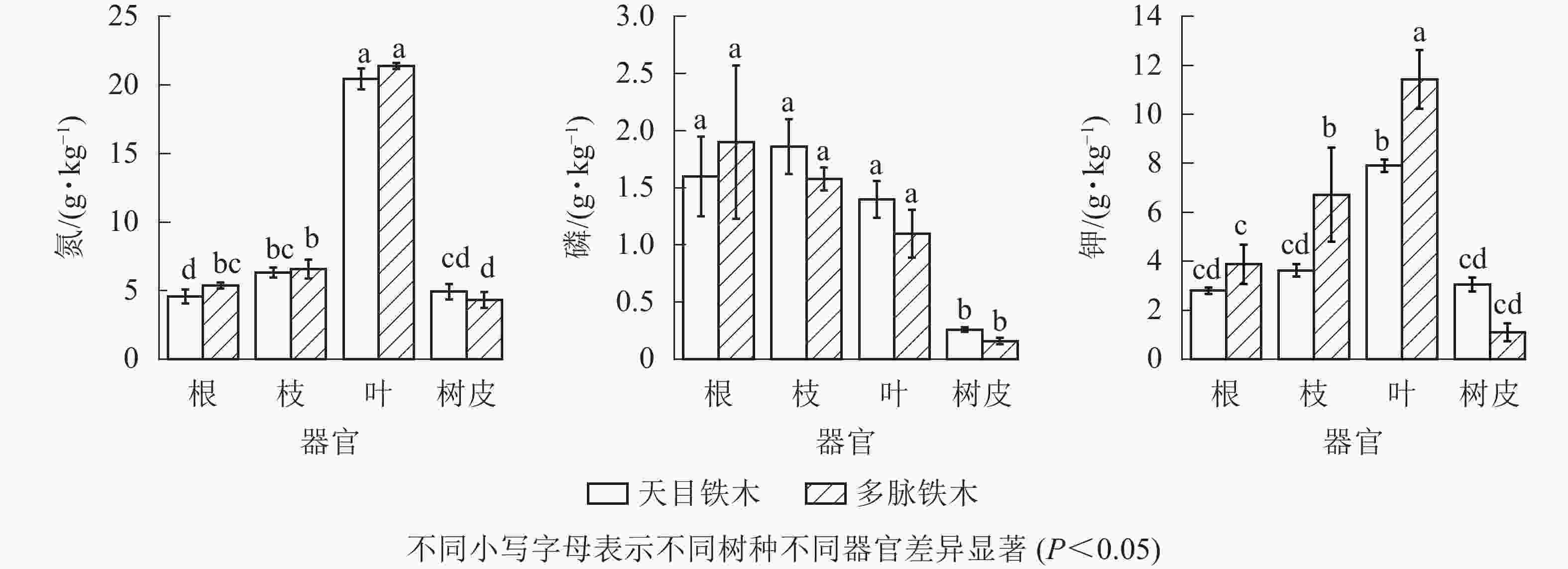
图 1 不同器官氮、磷、钾质量分数
Figure 1. Mass fractions of N, P and K in different organs
树皮中的氮、磷、钾元素的质量分数较低,而叶片的氮和钾质量分数远大于其他器官,根和枝的各元素质量分数较为接近。2种铁木相同器官之间相比,仅多脉铁木根中的氮和叶、枝中的钾显著高于天目铁木(P<0.05)。
-
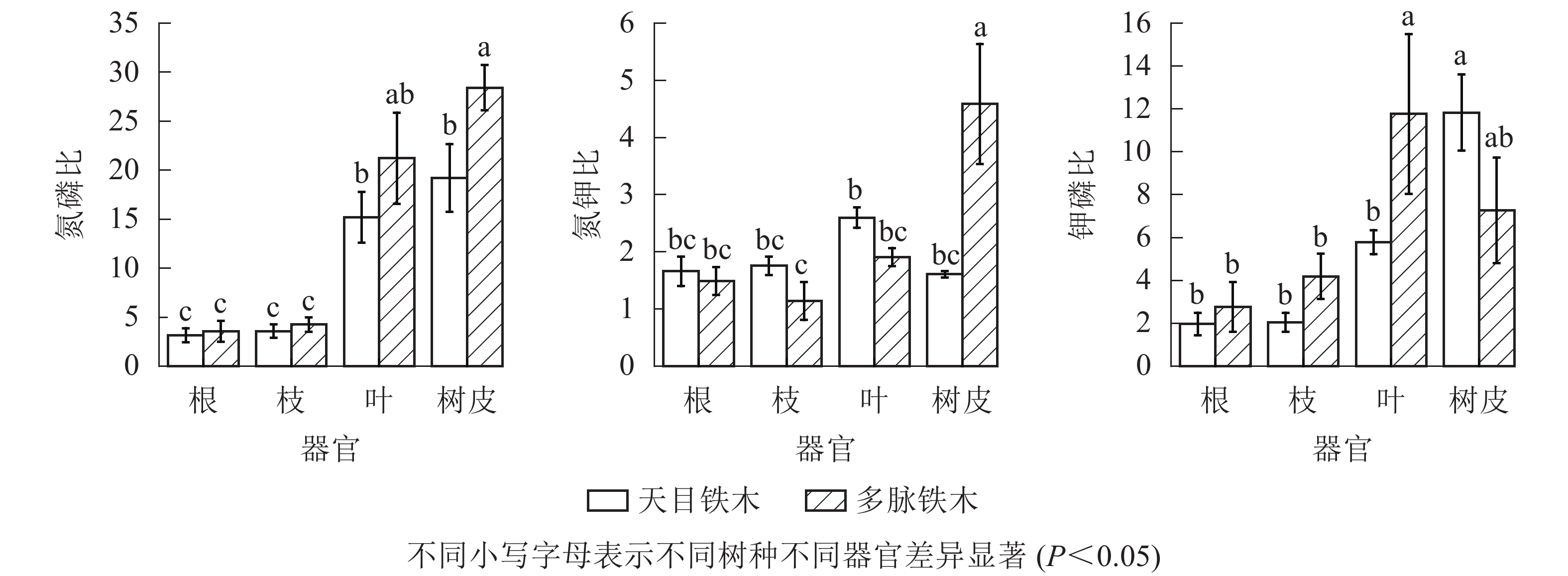
从图2中可知:2种铁木不同器官氮磷比差异变化较大,叶和树皮氮磷比显著高于根和枝(P<0.05),根和枝之间无显著差异(P>0.05)。多脉铁木的树皮氮磷比显著高于天目铁木的树皮(P<0.05)。2树种根、枝、叶相同器官之间相比,多脉铁木氮磷比高于天目铁木,但无显著差异(P>0.05)。总体上多脉铁木各不同器官的氮磷比都高于天目铁木。2种铁木不同器官氮钾比之间差异变化较小,仅有多脉铁木树皮氮钾比显著高于其他器官(P<0.05)以及天目铁木叶显著高于多脉铁木枝(P<0.05)。2树种的根、枝、叶相同器官之间相比,天目铁木略高于多脉铁木,但无显著性差异(P>0.05),多脉铁木树皮氮钾比显著高于天目铁木树皮。2种铁木不同器官钾磷比之间差异变化较大,多脉铁木叶和天目铁木树皮钾磷比显著高于其他器官(P<0.05),且多脉铁木叶也显著高于天目铁木叶(P<0.05)。2树种的根、枝相同器官之间相比,多脉铁木略高于天目铁木,但无显著性差异(P>0.05)。总体上除天目铁木树皮钾磷比高于多脉铁木,其他根、枝、叶器官都低于多脉铁木。
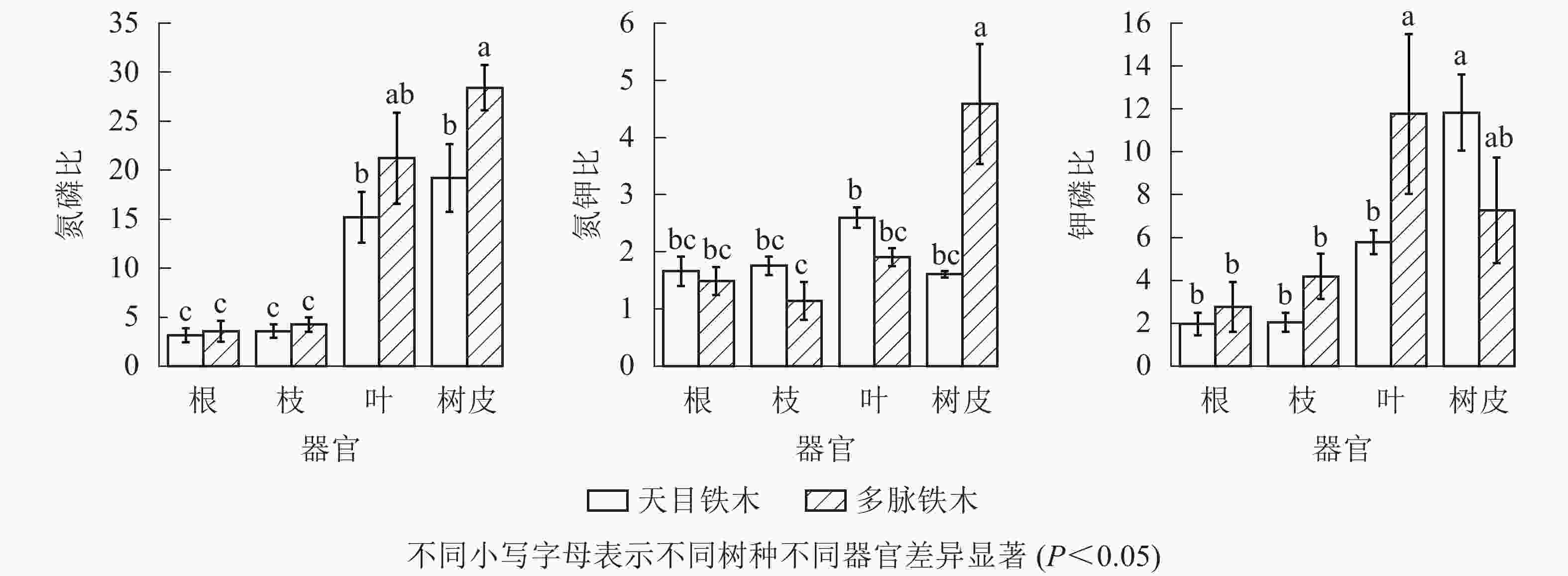
图 2 不同器官氮磷比、氮钾比、钾磷比
Figure 2. Ratio N∶P, N∶K, P∶K in different organs
-
通过植物的氮、磷、钾质量分数及其特征可以判断植物的营养状况与养分利用情况,判断植物在生长过程中应对外界环境变化的能力,也可与同种或其他物种的植物营养情况进行差异比较。本研究结果得出:天目铁木叶的氮、磷、钾质量分数平均值分别为20.441、1.400、7.914 g·kg−1,氮质量分数高于全国陆生植物叶片平均质量分数(19.090 g·kg−1),磷、钾质量分数平均值低于全国陆生植物叶片磷(1.560 g·kg−1)、钾(15.090 g·kg−1)质量分数的平均水平[24],钾质量分数平均值低于全国阔叶树(8.950 g·kg−1)和灌丛(8.380 g·kg−1)质量分数的平均水平[25],磷的质量分数高于全国常绿木本植物质量分数的平均水平(0.960 g·kg−1)。多脉铁木叶的氮、磷、钾质量分数平均值分别为21.376、1.097、11.425 g·kg−1,氮的质量分数高于全国陆生植物叶片氮的质量分数(19.090 g·kg−1),磷、钾的质量分数平均值低于全国陆生植物叶片磷(1.560 g·kg−1)、钾(15.090 g·kg−1)质量分数的平均水平[24],钾质量分数平均值高于全国阔叶树(8.950 g·kg−1)和灌丛(8.380 g·kg−1)质量分数的平均水平[25],磷的质量分数高于全国常绿木本植物磷质量分数(0.960 g·kg−1)平均水平。
2种植物叶片中氮质量分数均高于全国陆生植物叶片,这可能与研究区样地土壤氮质量分数背景值相对较高有关;2种植物叶片中磷质量分数均低于全国陆生植物叶片,一定程度上与2种植物体对磷的分配规律不同及利用能力较弱有关。叶片中的磷质量分数低可能是导致2种植物种群数量小的原因之一。
-
植物内元素的化学计量比可以客观反映植物的限制性元素及生长过程中所遵循的营养调控方案。氮、磷、钾是影响植物生长的限制性元素,氮磷比、氮钾比、钾磷比可作为植物营养元素限制的判断性指标[21,23,26]。研究表明:植物叶片的氮磷比可作为森林植物营养元素限制的判断性指标,当氮磷比<14时,植物生长主要受氮的限制;当氮磷比>16时,植物生长主要受磷的限制;当氮磷比为14~16时,受氮和磷的共同限制[27−29]。
天目铁木叶的氮磷比为14~16,说明天目铁木的生长受到氮、磷共同限制。当氮钾比>2.100,钾磷比<3.400时,植物的生长主要受钾的限制[30−32]。天目铁木氮钾比的平均值为1.607~2.597,钾磷比平均值为1.970~11.837,说明天目铁木的生长没有受到钾元素的限制。
多脉铁木叶的氮磷比远大于16,初步判断多脉铁木的生长主要受磷的限制。当氮钾比>2.100并且钾磷比<3.400时,植物的生长主要受钾的限制[30-32]。多脉铁木氮钾比的平均值为1.143~4.590,钾磷比平均值为2.770~11.770,说明多脉铁木的生长没有受到钾元素的限制。因此,在林地土壤管理中,天目铁木可适当增施氮肥和磷肥,而多脉铁可适当增施磷肥。
-
植株氮、磷、钾在不同器官中的质量分数反映了植物的生理活动和对不同环境的适应能力[32],叶片是植物光合作用的主要场所,氮是细胞合成叶绿素和蛋白质的主要元素,因此植物的叶片中氮的质量分数较高。天目铁木和多脉铁木叶片氮的平均比例达到56.3%和56.7%,这也进一步表明营养元素在不同器官与不同组分之间有显著差异,叶片氮质量分数显著高于其他器官。
磷不仅是核酸和核蛋白的主要成分,构成生物膜的重要组成部分,也对细胞分裂和植物各器官的分化发育具有不同程度的作用[33]。植物体不同器官中磷质量分数明显受到外界供磷水平的影响,当植株缺磷时,根系会保留从土壤中吸收的大部分磷,地上部生长发育所需的磷则主要依靠枝、叶中的磷再利用;而供磷适宜的植株根系仅会保留吸收的部分磷,大多数则运往地上部。根据本研究结果可以推测,2种铁木属植物生长受到了磷的限制。
根、树皮在植物中的作用是主要负责水分和养料的吸收与运输,对外界环境的响应不够敏感,因此根、树皮中的营养元素质量分数相对来说较为稳定。根是吸收外界营养物质和水分的器官,它将营养物质及水分由下往上传导至地上部分,以此完成植物正常的生长发育。由于天目铁木和多脉铁木生长的环境处于种群密度较高的森林群落中,为了获取更多的阳光照射,树木必须向上生长,增加叶片的数量及面积并且提升细胞内叶绿素的含量。植物选择优先满足叶片的各项生理活动及代谢的需求,对森林植物的生长来说,叶片中增加氮质量分数是合理且非常必要的调控手段。
Ecological stoichiometric characteristics of nitrogen, phosphorus and potassium in different organs of Ostrya rehderiana and O. multinervis
-
摘要:
目的 研究天目铁木Ostrya rehderiana和多脉铁木O. multinervis等2种极小种群植物不同器官的化学计量特征,推测2种植物生长的限制性元素,为林地土壤管理提供科学依据。 方法 以天目铁木和多脉铁木为研究对象,对植物根、枝、叶、树皮全氮、全磷、全钾的质量分数以及土壤基本理化性质进行了测定。采用SPSS软件进行差异显著性检验,分析2种植物不同器官之间的差异。 结果 天目铁木的氮、钾质量分数从大到小依次为叶、枝、皮、根,磷质量分数从大到小依次为枝、根、叶、皮;多脉铁木的氮、钾质量分数从大到小依次为叶、枝、根、皮,磷质量分数从大到小依次为根、枝、叶、皮。天目铁木的氮磷比平均值为3.18~19.22,氮钾比平均值为1.60~2.56,钾磷比平均值为1.97~11.80。多脉铁木氮磷比平均值为3.57~28.43,氮钾比平均值为1.14~4.59,钾磷比平均值为2.77~11.77。2种树种不同器官之间氮、磷、钾生态化学计量特征较为相似,仅多脉铁木根中的氮和叶、枝中的钾显著高于天目铁木的对应器官(P<0.05)。 结论 影响天目铁木生长的限制性元素是氮和磷,影响多脉铁木生长的限制性元素是磷。因此,在林地土壤管理中,天目铁木应适当增施氮肥和磷肥,而多脉铁木则应适当增施磷肥。图2表1参33 Abstract:Object This study aims to explore the stoichiometric characteristics of different organs of two minimal population plants, Ostrya rehderiana and O. multinervis, and to predict the limiting elements of the growth of the two plants, so as to provide scientific basis and suggestions for forest soil management. Method The mass fractions of total nitrogen, total phosphorus and total potassium in roots, branches, leaves and bark of plants as well as the basic physical and chemical properties of soil were determined. SPSS software was used to test the significance of differences and analyze the differences among different organs of the two plants. Result The N and K fractions of O. rehderiana in order from large to small were leaf, branch, bark and root, and the P fraction was branch, root, leaf and bark. The N and K fractions of O. multinervis in order from large to small were leaf, branch, root and bark, and the P mass was root, branch, leaf and bark. The mean values of N∶P, N∶K and K∶P in O. rehderiana were 3.18−19.22, 1.60−2.56, and 1.97−11.80, respectively.The mean values of N∶P, N∶K and K∶P in O. multinervis were 3.57−28.43, 1.14−4.59, and 2.77−11.77, respectively. The ecological stoichiometric features of N, P and K among different organs of the two species were similar, and only N in roots and K in leaves and branches of O. multinervis were significantly higher than those in the corresponding organs of O. rehderiana(P<0.05). Conclusion The limiting elements affecting the growth of O. rehderiana are N and P, and the one affecting the growth of O. multinervis is P. Therefore, in woodland soil management, N and P fertilizer should be properly applied to O. rehderiana, while P fertilizer should be properly applied to O. multinervis.[Ch, 2 fig. 1 tab. 33 ref.] -
Key words:
- forest ecology /
- Ostrya rehderiana /
- Ostrya multinervis /
- organ /
- ecological stoichiometry
-
表 1 天目铁木和多脉铁木根区土壤理化性质
Table 1. Basic physical and chemical properties of soil in O. rehderiana and O. multinervis woodlands
树种 pH 碱解氮/(mg·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) 有机碳/(g·kg−1) 天目铁木 6.26±0.03 a 166.4±2.67 a 5.25±0.38 a 158.5±1.48 a 24.1±0.23 A 多脉铁木 5.55±0.18 b 137.3±9.65 b 4.45±0.61 a 152.3±1.69 b 16.1±0.14 B 说明:表中同列小写字母表示不同树种土壤间差异显著(P<0.05),同列大写字母表示不同树种土壤间差异极显著(P<0.01)  下载: 导出CSV
下载: 导出CSV
-
[1] 孙雪娇, 常顺利, 宋成程, 等. 雪岭云杉不同器官N、P、K化学计量特征随生长阶段的变化[J]. 生态学杂志, 2018, 37(5): 1291 − 1298. SUN Xuejiao, CHANG Shunli, SONG Chengcheng, et al. Age-related N, P, and K stoichiometry in different organs of Picea schrenkiana [J]. Chin J Ecol, 2018, 37(5): 1291 − 1298. [2] 牛得草, 李茜, 江世高, 等. 阿拉善荒漠区6种主要灌木植物叶片C∶N∶P化学计量比的季节变化[J]. 植物生态学报, 2013, 37(4): 317 − 325. NIU Decao, LI Qian, JIANG Shigao, et al. Seasonal variations of leaf C∶N∶P stoichiometry of six shrubs in desert of China’s Alxa Plateau [J]. Chin J Plant Ecol, 2013, 37(4): 317 − 325. [3] 周鹏, 耿燕, 马文红, 等. 温带草地主要优势植物不同器官间功能性状的关联[J]. 植物生态学报, 2010, 34(1): 7 − 16. ZHOU Peng, GENG Yan, MA Wenhong, et al. Linkages of functional traits among plant organs in the dominant species of the Inner Mongolia grassland, China [J]. Chin J Plant Ecol, 2010, 34(1): 7 − 16. [4] 李红林, 贡璐, 洪毅. 克里雅绿洲旱生芦苇根茎叶C、N、P化学计量特征的季节变化[J]. 生态学报, 2016, 36(20): 6547 − 6555. LI Honglin, GONG Lu, HONG Yi. Seasonal variations in C, N, and P stoichiometry of roots, stems, and leaves of Phragmites australis in the Keriya Oasis, Xinjiang, China [J]. Acta Ecol Sin, 2016, 36(20): 6547 − 6555. [5] 李月芬, 王冬艳, LASOUKANH V, 等. 基于土壤化学性质与神经网络的羊草碳氮磷含量预测[J]. 农业工程学报, 2014, 30(3): 104 − 111. LI Yuefen, WANG Dongyan, LASOUKANH V, et al. Prediction of carbon, nitrogen and phosphorus contents of Leymus chinensis based on soil chemical properties using artificial neural networks [J]. Trans Chin Soc Agric Eng, 2014, 30(3): 104 − 111. [6] 曾冬萍, 蒋利玲, 曾从盛, 等. 生态化学计量学特征及其应用研究进展[J]. 生态学报, 2013, 33(18): 5484 − 5492. ZENG Dongping, JIANG Liling, ZENG Congsheng, et al. Reviews on the ecological stoichiometry characteristics and its applications [J]. Acta Ecol Sin, 2013, 33(18): 5484 − 5492. [7] AGREN G I, WEIH M. Plant stoichiometry at different scales: element concentration patterns reflect environment more than genotype [J]. New Phytol, 2012, 194(4): 944 − 952. [8] 赵明水, 张华峰. 天目铁木物理力学性质初步分析[J]. 浙江林业科技, 2006, 26(1): 52 − 55. ZHAO Mingshui, ZHANG Huafeng. Analysis on physical mechanics property of Ostrya rehderiana wood [J]. J Zhejiang For Sci Technol, 2006, 26(1): 52 − 55. [9] 吴世斌, 库伟鹏, 周小荣, 等. 浙江省极小种群多脉铁木年龄结构与动态[J]. 浙江农业科学, 2018, 59(8): 1381 − 1384. WU Shibin, KU Weipeng, ZHOU Xiaorong, et al. Age structure and dynamics of extremely small populations of Ostrya multinervis in Zhejiang Province [J]. J Zhejiang Agric Sci, 2018, 59(8): 1381 − 1384. [10] 张若蕙, 龚关文, 沈锡康, 等. 天目铁木花粉、种子及幼苗的研究[J]. 浙江林业科技, 1988, 8(4): 7 − 11, 30. ZHANG Ruohui, GONG Guanwen, SHEN Xikang, et al. A study of pollen, seed and seedling of Ostrya rehderiana Chun [J]. J Zhejiang For Sci Technol, 1988, 8(4): 7 − 11, 30. [11] 王祖良, 陆海根, 黄珊珊, 等. 天目铁木雄花序内源多胺的HPLC分析及其动态变化[J]. 安徽农业大学学报, 2012, 39(1): 79 − 83. WANG Zuliang, LU Haigen, HUANG Shanshan, et al. Dynamic changes of endogenous polyamines in male anthotaxy of Ostrya rehderiana by HPLC [J]. J Anhui Agric Univ, 2012, 39(1): 79 − 83. [12] 管康林, 陶银周. 濒危树种天目铁木的现状和繁殖[J]. 浙江林学院学报, 1988, 5(1): 90 − 92. GUAN Kanglin, TAO Yinzhou. Current situation and propagation of rare tree species: Ostrya rederiana [J]. J Zhejiang For Coll, 1988, 5(1): 90 − 92. [13] 乐笑玮, 崔敏燕, 杨淑贞, 等. 濒危植物天目铁木种子休眠及萌发特征研究[J]. 华东师范大学学报(自然科学版), 2013(6): 150 − 158. LE Xiaowei, CUI Minyan, YANG Shuzhen, et al. Characters on the seed dormancy and germination of an endangered species, Ostrya rehderiana, in Tianmu Mountain, China [J]. J East China Norm Univ Nat Sci, 2013(6): 150 − 158. [14] 孟爱平, 何子灿, 李建强, 等. 桦木科2种濒危植物的染色体数目[J]. 武汉植物学研究, 2004, 22(2): 171 − 173. MENG Aiping, HE Zican, LI Jianqiang, et al. Chromosome numbers of two threatened species of Betulaceae [J]. J Wuhan Bot Res, 2004, 22(2): 171 − 173. [15] 王祖良, 丁丽霞, 赵明水, 等. 濒危植物天目铁木遗传多样性的RAPD分析[J]. 浙江林学院学报, 2008, 25(3): 304 − 308. WANG Zuliang, DING Lixia, ZHAO Mingshui, et al. Genetic diversity of Ostrya rehderiana revealed by RAPD markers [J]. J Zhejiang For Coll, 2008, 25(3): 304 − 308. [16] 顾地周, 陆爽, 巴春影, 等. 天目铁木愈伤组织和芽苗诱导技术[J]. 浙江大学学报(理学版), 2013, 40(2): 216 − 220. GU Dizhou, LU Shuang, BA Chunying, et al. Technique of callus induction and bud seedling of Ostrya rehderiana Chun [J]. J Zhejiang Univ Sci Ed, 2013, 40(2): 216 − 220. [17] 王晓燕, 杨淑贞, 赵明水, 等. 濒危植物天目铁木和羊角槭的光合及蒸腾特性日动态比较[J]. 华东师范大学学报(自然科学版), 2015(2): 113 − 121. WANG Xiaoyan, YANG Shuzhen, ZHAO Mingshui, et al. Comparative diurnal variations in photosynthesis and transpiration ofendangered plant species, Ostrya rehderiana and Acer yangjuechi [J]. J East China Norm Univ Nat Sci, 2015(2): 113 − 121. [18] 罗远, 吴世斌, 库伟鹏, 等. 珍稀濒危植物天目铁木群落结构及物种多样性[J]. 浙江农业科学, 2018, 59(11): 2061 − 2064. LUO Yuan, WU Shibin, KU Weipeng, et al. Community structure characteristics and species diversity of rare and endangered plants of Ostrya rehderiana [J]. J Zhejiang Agric Sci, 2018, 59(11): 2061 − 2064. [19] 《石垟林场志》编篡委员会. 石垟林场志[M]. 北京: 北京艺术与科学电子出版社, 2012. [20] 吴世斌, 库伟鹏, 周小荣, 等. 浙江文成珍稀植物多脉铁木群落结构及物种多样性[J]. 浙江农林大学学报, 2019, 36(1): 31 − 37. WU Shibin, KU Weipeng, ZHOU Xiaorong, et al. Structural characteristics and species diversity for survival of the rare plant Ostrya multinervis [J]. J Zhejiang A&F Univ, 2019, 36(1): 31 − 37. [21] 叶柳欣, 张勇, 蒋仲龙, 等. 不 同林龄杨梅叶片与土壤的碳、氮、磷生态化学计量特征[J]. 安徽农业大学学报, 2019, 46(3): 454 − 459. YE Liuxin, ZHANG Yong, JIANG Zhonglong, et al. The stoichiometic characteristics of carbon, nitrogen and phosphorus in soil and leaves of different ages of Myrica rubra [J]. J Anhui Agric Univ, 2019, 46(3): 454 − 459. [22] 王增, 蒋仲龙, 刘海英, 等. 油茶不同器官氮、磷、钾化学计量特征随年龄的变化[J]. 浙江农林大学学报, 2019, 36(2): 264 − 270. WANG Zeng, JIANG Zhonglong, LIU Haiying, et al. Ecological stoichiometry of N, P, and K with age in Camellia oleifera organs [J]. J Zhejiang A&F Univ, 2019, 36(2): 264 − 270. [23] 吴家森, 张勇, 吕爱华, 等. 不同林龄油茶叶片与土壤的碳氮磷生态化学计量特征研究[J]. 西南林业大学学报(自然科学), 2019, 39(3): 86 − 92. WU Jiasen, ZHANG YONG, LÜ Aihua, et al. Eco-stoichiometric characteristics of carbon, nitrogen and phosphorusin leaves and soil of Camellia oleifera at different ages [J]. J Southwest For Univ Nat Sci, 2019, 39(3): 86 − 92. [24] 任书杰, 于贵瑞, 陶波, 等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 2007, 28(12): 2665 − 2673. REN Shujie, YU Guirui, TAO Bo, et al. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC [J]. Environ Sci, 2007, 28(12): 2665 − 2673. [25] 秦海, 李俊祥, 高三平, 等. 中国660种陆生植物叶片8种元素含量特征[J]. 生态学报, 2010, 30(5): 1247 − 1257. QIN Hai, LI Junxiang, GAO Sanping, et al. Characteristics of leaf element contents for eight nutrients across 660 terrestrial plant species in China [J]. Acta Ecol Sin, 2010, 30(5): 1247 − 1257. [26] 郭素娟, 谢明明, 张丽, 等. 板栗细根碳、氮、磷化学计量时间变异特征[J]. 植物营养与肥料学报, 2018, 24(3): 825 − 832. GUO Sujuan, XIE Mingming, ZHANG Li, et al. Temporal variation of C, N, P stoichiometric in fine roots of Castanea mollissima [J]. J Plant Nutr Fert, 2018, 24(3): 825 − 832. [27] 皮发剑, 袁丛军, 喻理飞, 等. 黔中天然次生林主要优势树种叶片生态化学计量特征[J]. 生态环境学报, 2016, 25(5): 801 − 807. PI Fajian, YUAN Congjun, YU Lifei, et al. Ecological stoichiometry characteristics of plant leaves from the maindominant species of natural secondary forest in the central of Guizhou [J]. Ecol Environ Sci, 2016, 25(5): 801 − 807. [28] 姜沛沛, 曹扬, 陈云明, 等. 不同林龄油松(Pinus tabulaeformis)人工林植物、凋落物与土壤C、N、P化学计量特征[J]. 生态学报, 2016, 36(19): 6188 − 6197. JIANG Peipei, CAO Yang, CHEN Yunming, et al. Variation of C, N, and P stoichiometry in plant tissue, litter, and soil during stand development in Pinus tabulaeformis plantation [J]. Acta Ecol Sin, 2016, 36(19): 6188 − 6197. [29] VENTERINK H G M O, WASSEN M J, VERKROOST A W M, et al. Species richness-productivity patterns differ between N-, P-, and K-limited wetlands [J]. Ecology, 2003, 84(8): 2191 − 2199. [30] 吴家森, 蒋仲龙, 吕爱华, 等. 不同年龄杨梅各器官氮、磷、钾化学计量特征[J]. 江西农业大学学报, 2019, 41(3): 447 − 453. WU Jiasen, JIANG Zhonglong, LÜ Aihua, et al. The ecological stoichiometry of N, P and K in organs of Myrica rubra of different ages [J]. Acta Agric Univ Jiangxi, 2019, 41(3): 447 − 453. [31] KOCRSCLMAN W, MEULEMAN AF M. The vegetation N∶P ratio: anew tool to detect the nature of nutrient limitation [J]. J Appl Ecol, 1996, 33(6): 1441 − 1450. [32] WRIGHT I J, REICH P B, WESTOBY M, et al. The worldwide leaf economics spectrum [J]. Nature, 2004, 428(6985): 821 − 827. [33] ZHANG Sheng, ZHOU Rong, ZHAO Hongxia, et al. iTRAQ-based quantitative proteomic analysis gives insight into sexually different metabolic processes of poplars under nitrogen and phosphorus deficiencies [J]. Proteomics, 2016, 16(4): 614 − 628. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200470
 点击查看大图
点击查看大图

图(2) / 表(1)
计量
- 文章访问数: 2391
- HTML全文浏览量: 649
- PDF下载量: 47
- 被引次数: 0
