




-
间伐是优化林分密度,调整林分空间分布格局的重要经营措施[1-2],能通过增加林冠开度、降低冠层叶面积指数、增加冠层枝叶反射的散射光和透过林层的直射光来提高林下植被的光合作用速率,促进林下植物有机质的积累[3-4]。林下灌木在稳定森林群落结构和维持物种多样性以及种子萌发和幼苗更新等方面发挥重要作用[5-6],在一定程度上影响着林下更新幼苗定居及早期生长状况[7]。植物功能性状是植物在漫长进化过程中为适应或降低环境干扰、充分利用环境资源所形成的生物特征,能揭示植物适应环境的能力和策略[8-9]。环境引起的植物功能性状的变化主要表现在器官(根、茎、叶等)的结构性状和化学性状的权衡关系[10],其中,叶片作为光能捕获及有机质转化器,与植物的资源获取和生物量密切相关[11];茎是植物地上部分的支撑组织,具有储存和运输养分功能[12];根系的生长和发达程度决定了土壤资源的获取和利用状况[13]。三者功能性状均对植物生长状况具有一定指示作用。目前,关于环境变化对植物功能性状影响的研究主要集中于叶片[14],对整个植株各器官功能性状的研究未见报道。间伐对林冠结构的调整使林下光照增加,植物结构性状和化学性状的调节会影响植物光合作用[15],进而促进植物有机质的积累。研究间伐对林下灌木功能性状的影响,为有效改善林分结构,促进栓皮栎Quercus variabilis人工林更新提供理论和技术支撑。荆条Vitex negundo var. heterophylla作为研究区灌层的建群种,具有较强的耐旱性,是干旱半干旱地区生态恢复的重要指示性物种[16-17]。栓皮栎人工林自然整枝能力较弱,林相差,林下更新萌蘖多,实生苗少且未得到有效保护,急需开展以人工促进天然更新为主要措施的森林抚育。有研究表明:弱度间伐最有利于栎类幼苗更新[18],林下灌木层盖度对4~10年生幼苗数量具有决定性作用[19]。荆条的“肥岛”效应为林下幼苗生长提供相对充足的生长资源[20]。因此,有必要研究弱度间伐对林下荆条功能性状的影响,以较准确地反映间伐后荆条应对环境资源变化的生存策略,为干旱半干旱地区栓皮栎人工林可持续经营管理提供科学依据。
-
研究区位于河南省国有登封林场清凉寺林区(34°26′~34°33′N,112°44′~113°05′E),海拔为518~755 m,坡度为5°~30°。该地区属暖温带大陆性季风气候,年平均气温14.2 ℃,年平均降水量614.0 mm,降水主要集中在夏季,占全年总降水量的33.8%左右。土壤贫瘠,以褐土和棕壤为主,表现为富钾、少氮、极缺磷。乔木以栓皮栎、侧柏Platycladus orientalis、刺槐Robinia pseudoacacia、槲栎Q. aliena、黄栌Cotinus coggygria为主,灌木以荆条Vitex negundo var. heterophylla、扁担杆Grewia biloba、酸枣Ziziphus jujuba为主。
-
2016年在登封国有林场选取2个林分立地条件、坡度、坡向、坡位相同,林龄、林分密度、林木生长状况基本一致的林班,样地栓皮栎间伐及未间伐林分年龄均为40 a,郁闭度约0.90,坡度分别为9°和7°,坡向均为南坡,平均胸径分别为10.01和10.10 cm,平均树高分别为7.98和8.06 m,林分密度分别为2 200和2 211株·hm−2。分别设置1个100 m×100 m(1 hm2)间伐样地和未间伐样地。间伐方式以修枝、非全面割灌、伐除病腐木及没有培育前途的林木为主的生产性间伐,基本不降低林分蓄积量(间伐株数强度31.55%,蓄积强度为3.26%)。2019年调查间伐样地林分密度为1 532株·hm−2,林木平均胸径12.80 cm,平均树高11.45 m,未间伐样地林分密度2 110株·hm−2,林木平均胸径10.64 cm,平均树高9.05 m。间伐3 a后,间伐后林木生长加快,平均胸径及树高均有所提高,一方面是因为大径级林木被保留下来,另一方面是保留木生长空间及资源充足,加速林木生长。2019年6月在间伐样地和未间伐样地分别均匀设置6个20 m×20 m的方形森林动态监测样方,调查每个样方内荆条平均高度及冠幅,选取具有平均高度和平均叶量的荆条3株,共36株。采集东西南北4个方向完全展开、没有病虫害的叶片。用BY-N4型植物营养测定仪测定叶片叶绿素和氮质量分数,用Yaxin-1241叶面积仪测定荆条叶面积、叶长、叶宽等指标,采用全根挖掘法采集植株根系,带回实验室于2019年6−7月进行地上功能性状和根系结构功能指标测定。样地设置及样地基本情况见闫东锋等[1]。
-
选取荆条根、茎、叶共32个植物功能指标,调查及测定的植物功能性状指标见表1。表1中,叶长宽比=叶长/叶宽,比叶面积=叶面积/叶片质量,叶干物质质量分数=叶干质量/叶鲜质量,茎干物质质量分数=茎干质量/茎鲜质量,根干物质质量分数=根干质量/根鲜质量,比根长=总根长/根干质量,基径采用Haglof游标卡尺测定。植株各器官全磷质量分数通过钼锑抗比色法测定,全钾采用火焰光度计测定,有机碳通过烘箱加热法测定。
表 1 所选植物功能性状
Table 1. Functional traits of selected plants
植物功能性状 单位符号 植物功能性状 单位符号 植物功能性状 单位符号 植物功能性状 单位符号 高度 m 基径 cm 根干物质质量分数 茎全氮 g·kg−1 叶周长 cm 茎生物量 g 根生物量比 茎全钾 mg·kg−1 叶面积 cm2 茎干物质质量分数 根冠比 茎全磷 g·kg−1 比叶面积 cm2·g−1 主根长 cm 比根长 cm·g−1 茎有机碳 g·kg−1 叶绿素 总根长 cm 叶全氮 g·kg−1 根全氮 g·kg−1 叶生物量 g 根表面积 cm2 叶全钾 mg·kg−1 根全钾 mg·kg−1 叶干物质质量分数 根体积 cm3 叶全磷 g·kg−1 根全磷 g·kg−1 叶长宽比 根生物量 g 叶有机碳 g·kg−1 根有机碳 g·kg−1 采用GLA 2.0软件获取林下光照条件(林下直射光和林下散射光)。用CI-110冠层分析仪测定荆条冠层的光合有效辐射强度、太阳辐射时间等指标。采用五点取样法对样地0~10和10~20 cm的土壤取样,并充分混匀带回实验室测定土壤营养元素质量分数,其中土壤全氮采用凯氏定氮法测定,全磷、全钾、有机碳的测定与荆条养分测定方法相同。采用环刀法测定土壤容重,并计算土壤毛管孔隙度、非毛管孔隙度等物理性质。
-
对各器官功能性状的差异进行显著性检验(t检验),在检验之前对数据的正态性进行检验;计算荆条地上功能性状与林下光环境和根系与土壤理化性质的Pearson相关系数;以荆条各器官功能性状指标为因变量,以与其相关关系显著的环境因子作为自变量进行线性逐步回归分析。
-
从表2可见:间伐样地荆条平均高为0.97 m,高于未间伐样地(0.92 m),2个样地荆条高为0.86~1.06 m,但无显著差异(P>0.05)。间伐样地荆条叶周长、叶面积、比叶面积和叶生物量(48.91 cm,17.27 cm2,31.97 cm2·g−1,90.53 g)均显著高于未间伐样地(33.16 cm,10.18 cm2,24.07 cm2·g−1,67.55 g,P<0.05),叶干物质质量分数(0.35)低于未间伐样地(0.44),但差异不显著(P>0.05)。基径(1.39 cm)高于未间伐样地(1.18 cm),但分布均为0.60~1.69 cm。
表 2 间伐对荆条地上部分功能性状的影响
Table 2. Effect of thinning on the functional traits of aboveground part of on V. negundo var. heterophylla
项目 措施 高度/m 叶周长/cm 叶面积/cm2 比叶面积/(cm2·g−1) 叶绿素 叶生物量/g 平均值±标准差 间伐 0.97±0.03 A 48.91±1.04 A 17.27±0.49 A 31.97±0.63 A 43.55±2.50 A 90.53±9.68 A 未间伐 0.92±0.03 A 33.16±1.77 B 10.18±1.71 B 24.07±1.85 B 34.15±3.60 A 67.55±4.52 B 数值范围 间伐 0.91~1.06 46.37~52.49 15.90~18.55 30.37~33.12 37.75~52.75 66.02~112.78 未间伐 0.86~0.98 28.31~37.33 5.53~15.67 18.15~28.46 20.50~40.05 53.01~76.94 项目 措施 叶干物质质量分数 叶长宽比 基径/cm 茎生物量/g 茎干物质质量分数 平均值±标准差 间伐 0.35±0.02 A 1.60±0.06 A 1.39±0.20 A 32.91±6.82 A 0.55±0.02 A 未间伐 0.44±0.05 A 2.43±0.88 A 1.18±0.15 A 25.02±6.13 A 0.52±0.03 A 数值范围 间伐 0.30~0.38 1.45~1.76 0.60~1.69 16.64~43.28 0.47~0.59 未间伐 0.35~0.47 1.20~5.91 0.80~1.69 12.13~47.25 0.41~0.58 说明:同列不同字母表示功能性状间存在显著差异(P<0.05),相同字母表示无显著差异(P>0.05) -
由表3可知:间伐后荆条主根长和根表面积(9.02 cm,701.032 cm2)大于未间伐样地(8.8 cm,516.52 cm2),但差异不显著(P>0.05);总根长、根生物量、比根长和根体积(207.3 cm,43.78 g,6.79 cm·g−1,31.79 cm3)均显著高于未间伐样地(133.96 cm,34.58 g,3.97 cm·g−1,18.50 cm3,P<0.05)。荆条根干物质质量分数和根生物量比小于未间伐样地,间伐样地和未间伐样地荆条根冠比差异不大。
表 3 间伐对荆条地下部分功能性状的影响
Table 3. Effect of thinning on the functional traits of underground part of V. negundo var. heterophylla
项目 措施 主根长/cm 根总长/cm 根表面积/cm2 根体积/cm3 根生物量/g 平均值±标准差 间伐 9.02±0.91 A 207.30±24.40 A 701.03±180.68 A 31.79±3.28 A 43.78±7.36 A 未间伐 8.80±0.61 A 133.96±12.61 B 516.52±251.68 A 18.50±4.45 B 34.58±3.32 B 数值范围 间伐 6.6~12.10 131.42~285.56 508.90~1060.67 21.99~42.34 14.88~56.32 未间伐 7.5~10.60 108.45~169.85 275.21~865.78 9.0~33.65 27.22~42.49 项目 措施 根干物质质量分数 根生物量比 根冠比 比根长/(cm·g−1) 平均值±标准差 间伐 0.62±0.01 B 0.40±0.08 B 1.16±0.42 A 6.79±0.31 A 未间伐 0.68±0.05 A 0.52±0.05 A 1.16±0.31 A 3.97±0.41 B 数值范围 间伐 0.52~0.75 0.21~0.53 0.27~2.15 2.64~19.19 未间伐 0.56~0.82 0.39~0.69 0.44~2.28 2.69~5.22 说明:同列不同字母表示功能性状间存在显著差异(P<0.05),相同字母表示无显著差异(P>0.05) -
对栓皮栎人工林间伐和未间伐林下荆条各器官养分分配比较分析(图1)发现:荆条营养物质植株各器官中的分配规律一致:氮元素主要分配给叶,其次为根;磷元素质量分数较低且主要分布在植株根中;钾元素分配规律从大到小依次为叶、茎、根;有机碳质量分数从大到小依次为茎、根、叶。
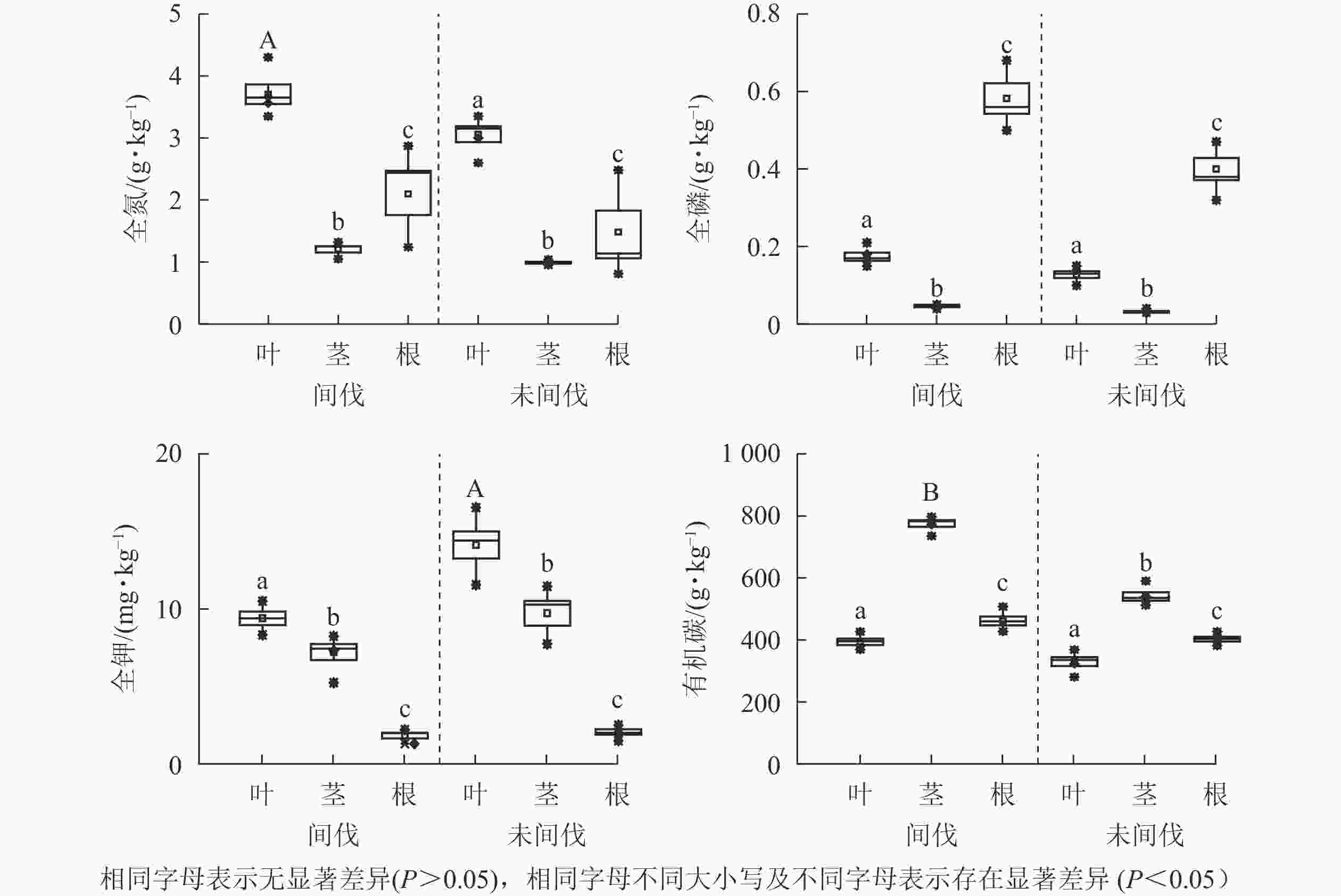
图 1 间伐对荆条各器官养分分配影响
Figure 1. Effect of thinning on nutrient distribution of various organs of V. negundo var. heterophylla
间伐与未间伐样地荆条同一器官化学性状分配规律表现为:间伐样地荆条叶片氮质量分数以及根和茎的有机碳质量分数(3.72、775.33、461.30 g·kg−1),显著高于未间伐样地(3.06、540.16、402.49 g·kg−1,P<0.05)。这说明间伐不改变矿质养分在荆条各器官的分配规律,但在一定程度上促进叶和茎功能性状的优化。间伐样地荆条叶全钾质量分数(9.08 mg·kg−1)显著低于未间伐样地(11.63 mg·kg−1,P<0.05),茎全钾质量分数(5.69 mg·kg−1)低于未间伐样地(4.43 mg·kg−1),但差异不显著(P>0.05)。这与未间伐样地土壤钾质量分数较高有关。
-
由表4可知:荆条叶面积与林下散射辐射通量、总辐射通量和太阳辐射时间呈极显著正相关关系(P<0.01),比叶面积与林下直射辐射通量和散射辐射通量呈极显著正相关(P<0.01),与太阳辐射时间呈显著正相关(P<0.05)。叶全氮质量分数和有机碳质量分数与光合有效辐射强度和总辐射通量呈显著正相关(P<0.05),与太阳辐射时间呈极显著正相关(P<0.01)。茎干物质质量分数与光合有效辐射呈显著正相关(P<0.05),茎全钾与林下直射辐射通量和散射辐射通量呈显著正相关(P<0.05),与太阳辐射时间呈极显著正相关(P<0.01)。茎有机碳质量分数与林下直射辐射通量和总辐射通量呈显著正相关(P<0.05)。
表 4 荆条地上部分功能性状与林下光环境的相关性分析
Table 4. Correlation analysis of aboveground part functional traits of V. negundo var. heterophylla and understory light environment
地上部分功能性状 光合有效辐射强度 直射辐射通量 散射辐射通量 总辐射通量 太阳辐射时间 叶面积 0.534 0.442 0.823** 0.837** 0.882** 叶绿素 0.263 0.578 0.484 0.274 0.528 叶干物质质量分数 0.861 0.215 0.521 0.215 0.779 比叶面积 0.547 0.797** 0.837** 0.768** 0.661* 叶全氮 0.706* 0.304 0.492 0.902* 0.946* 叶全磷 0.180 0.071 0.186 0.552 0.620 叶全钾 0.664* 0.831** 0.311 0.752* 0.520 叶有机碳 0.663* 0.821** 0.752* 0.761* 0.940** 茎干物质质量分数 0.841** 0.018 0.488 0.831** 0.783** 茎生物量比 −0.400 0.171 −0.316 0.067 −0.141 茎全氮 0.415 0.296 0.447 0.111 0.569 茎全磷 0.209 0.160 0.244 0.031 0.435 茎全钾 0.640* 0.710* 0.769** 0.786** 0.649* 茎有机碳 0.313 0.862** 0.686 0.801** 0.669 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
表5表明:栓皮栎人工林林下荆条总根长与土壤容重和毛管孔隙度、总孔隙度呈显著负相关(P<0.01),与土壤非毛管孔隙度、土壤有机碳呈极显著正相关(P<0.01),根表面积与土壤容重呈显著负相关(P<0.05),与土壤非毛管孔隙度、有机质和全氮呈极显著正相关(P<0.05),根体积与土壤容重呈极显著负相关(P<0.01),与非毛管孔隙度和土壤有机质呈极显著正相关(P<0.01)。根全氮与土壤非毛管孔隙度呈显著正相关(P<0.05),根有机碳与土壤容重呈极显著负相关(P<0.01),与土壤非毛管孔隙度和土壤有机碳呈极显著正相关(P<0.01)。这说明间伐改善了土壤团粒结构,使土壤容重降低,有利于增大土壤非毛管孔隙度,促进荆条根系对土壤养分和水分的吸收,从而促进植物的生长以及土壤有机碳的积累。
表 5 荆条根功能性状与土壤因子的相关性分析
Table 5. Correlation analysis of the functional characters of V. negundo var. heterophylla root and soil factors
根功能性状 土壤容重 非毛管孔隙度 毛管孔隙度 总孔隙度 土壤有机碳 土壤全氮 土壤全磷 土壤全钾 主根长 −0.547 0.612 −0.541 −0.454 0.619 0.756* 0.432 0.357 总根长 −0.807** 0.829** −0.848** −0.791** 0.873** 0.451 −0.109 −0.155 根表面积 −0.739* 0.740* −0.672 −0.577 0.734* 0.690* 0.290 0.431 根体积 −0.914** 0.953** −0.876 −0.759 0.915** 0.604 0.175 0.139 根生物量 −0.479 0.460 0.212 0.228 0.233 0.009 −0.407 −0.120 根干物质质量分数 −0.629 0.650* −0.077 −0.016 0.378 0.634* 0.238 0.479 根生物量比 0.174 −0.158 −0.597 −0.578 −0.474 −0.031 −0.104 0.423 根冠比 −0.142 0.166 −0.663* −0.620 −0.016 0.217 0.260 0.481 比根长 −0.588 0.551 −0.834** −0.838** 0.752* 0.485 0.108 −0.074 根全氮 −0.634 0.647* −0.578 −0.540 0.586 0.505 −0.060 0.153 根全磷 −0.858** 0.717* −0.934** −0.871** 0.818** 0.265 −0.220 −0.221 根全钾 −0.813** 0.775** 0.212 0.228 0.750* 0.439 −0.234 −0.223 根有机碳 −0.916** 0.913** −0.077 −0.016 0.979** 0.433 −0.058 −0.197 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01) -
为进一步探究栓皮栎人工林间伐背景下,林下荆条地上部分功能性状与林下光环境和荆条根与土壤理化特性指标值的关系,以荆条功能性状为因变量(y),以与各功能性状密切相关的5个光环境因子和5个土壤理化因子为自变量进行线性逐步回归分析,结果见表6。
表 6 荆条各器官功能性状与环境因子的逐步回归分析模型
Table 6. Stepwise regression analysis of V. negundo var. heterophylla functional traits and environmental factors each organ
功能性状 回归方程 P R2 叶面积 y=−6.89+1.282x4+0.056x5 0.026 0.896 比叶面积 y=−7.385−0.13x1+10.229x3 0.014 0.910 叶全氮 y=1.143+0.013x4+0.089x5 0.048 0.911 叶有机碳 y=77.476+77.11x1−0.794x3 0.160 0.987 茎干物质质量分数 y=0.415+0.001x1 0.002 0.706 茎有机碳 y=74.329+121.525x2 0.038 0.620 总根长 y=−118.671−2.234 x7+3.555 x8+4.312 x9 0.001 0.783 根干物质 y=−230.074+10x7−0.013x9 0.017 0.753 根体积 y=−9.176−15.724 x6+3.499 x7+0.199 x9 0.001 0.923 比根长 y=−17.111−4.765 x7+1.124x8+0.840 x9 0.003 0.930 根有机碳 y=−338.69+4.253x9−26.676x10 0.001 0.978 说明:x1为光合有效辐射;x2为林下直射辐射通量;x3为林下散射辐射通量;x4为林下总辐射通量;x5为太阳辐射时间;x6为土壤容 重;x7为土壤非毛管孔隙度;x8为毛管总孔隙度;x9为土壤有机质;x10为土壤全氮。 由地上部分功能性状指标与光环境的线性逐步回归分析结果可以看出:荆条叶面积和叶全氮均与林下总辐射通量和太阳直射时间存在较好的线性回归关系,R2分别达0.896和0.911,P均小于0.05。比叶面积和叶有机碳质量分数与光合有效辐射和林下散射辐射线性回归拟合效果较好,R2分别达0.910和0.987,P分别小于0.01和0.05。茎干物质质量分数与光合有效辐射有较强的线性回归关系(P<0.01,R2=0.896),而茎有机碳质量分数与林下直射辐射通量存在较好的线性回归关系(P<0.05,R2=0.620)。
根功能性状与其相关性较强的土壤理化特性线性逐步回归分析结果表明:根总长与土壤非毛管孔隙度、总孔隙度和土壤有机碳,根体积和比根长与土壤容重土壤非毛管孔隙度和毛管孔隙度,根有机碳与土壤有机碳和土壤全磷的线性回归拟合效果较好,R2分别达0.783、0.923、0.930、0.978,P均小于0.01。根干物质质量分数与土壤非毛管孔隙度和土壤有机碳,根体积与土壤容重存在较好的线性回归关系(R2=0.753,P<0.05)。其中,对土壤理化特征反应敏感的指标为根有机碳质量分数、比根长、根体积,R2分别为0.978、0.930、0.923,对根系影响最大的土壤理化指标是土壤有机质。
-
生产性抚育间伐往往是采用清理病腐木、修枝、割灌等为主的低强度间伐为特征的作业方法,而本研究对象采用的是割除对影响抚育作业的少量灌木为主的非全面割灌作业方式。相对于酸枣Ziziphus jujuba等其他灌木,荆条对于开展作业活动影响不大,因此作业时大部分被保留下来。间伐能够改善林内环境,提高栎林林下幼苗天然更新密度[18],促进栎类树种幼苗的定居和生长,而荆条作为栓皮栎人工林林下的主要建群灌木种类,对于林下更新幼苗具有一定的保护和“肥岛”效应[20],可以为林下更新幼苗的早期定居和生长提供遮光环境,这不仅对于需要遮光的栓皮栎更新幼苗的生长发育具有一定的保护作用,且由于荆条灌丛“肥岛”现象对土壤持水保肥发挥着重要作用,可为幼苗生长提供相对充足的水肥资源,因此,林下荆条数量及生长状况直接或间接地影响着林下更新幼苗的存活与发育,而林下更新幼苗的生长状况决定着林分更新潜力和抗干扰能力。因此,间伐作为改善林分结构的主要措施,研究生产性间伐对林下主要灌木种类(如荆条)的影响,对于预测林下天然更新动态,制定科学合理的人工促进天然更新措施(如选择性割除或保留林下灌木),实现森林可持续经营具有一定的参考意义。
-
间伐后林内环境发生变化,植物叶片是对光环境变化最敏感的器官,而茎功能性状与植物固碳能力、养分水分传输和防御能力密切相关,对植物生长发育状况具有指示作用[10-11]。根系是与土壤密切接触的器官,其功能性状均能对环境变化做出响应。本研究发现:间伐样地荆条叶面积、比叶面积、叶全氮、茎全氮和茎有机碳显著高于未间伐样地。主要原因可能是,相对于未间伐样地,间伐样地林冠开度的增大使林内光照、温度、水分及土壤理化性质发生异质性[21],荆条能够获取更充足的资源,使地上部分功能性状均处于较高水平。这说明间伐后林下光环境充足,荆条采用资源获取性生长策略,通过优化自身的结构性状和化学性状,截获更多光能,促进荆条叶片光合作用和有机质的积累。未间伐样地荆条具有较低的比叶面积和较高的叶干物质,说明荆条采取保守型生存策略,通过提高养分利用率,来适应遮光较强的未间伐样地,这与路兴慧等[22]的研究结果一致,即较小的比叶面积和较高的叶干物质有利于林下苗木适应低光环境相一致。总根长、根体积、根生物量和根有机碳均显著高于未间伐样地,原因可能是间伐样地冠层开度的增大在一定程度上增加林内土壤温度,减少降水截留,增加土壤湿度,有利于土壤微生物的生存与繁殖,加速林下腐殖质的分解,有利于林下土壤矿质养分的增加,为荆条根系提供充足的养分,促进根系长度的增加和生物量的积累。但根干物质低于未间伐样地,差异不显著。这可能由于相对于间伐样地,未间伐样地根系的水分比例相对较低,保持较高的根干物质,有利于荆条根系的水分保持。
-
森林冠层是森林内部与外界环境相互作用最直接、最活跃的交界面[23]。林冠结构的垂直分量会影响穿过林冠的透光率[24],林冠开度的扩大能促进林下总辐射通量、林下直射光和散射光的增加[3]。间伐引起的林下光环境的优化将直接或间接的影响着植物地上部分功能性状。本研究发现:荆条比叶面积与光合有效辐射强度和散射辐射通量呈极显著的线性正相关,与CORNELISSEN等[25]认为:叶面积越大越有利于植物增强捕获资源能力的研究结果一致。HOGAN等[26]研究间伐对热带幼树功能性状的影响结果:林冠开度增大有利于比叶面积较大的先锋树种定居和生长也佐证这一结论。叶全氮与林下总辐射通量呈极显著的线性正相关关系。这说明间伐后,冠层结构的优化促进林下光环境改善,有利于荆条叶进行光合作用,而氮是光合蛋白质的重要组成成分,与植物的光合作用密切相关。这与高林浩等[27]的研究结果比叶面积与叶全氮含量呈显著正相关相互验证。茎有机碳均与直射辐射通量呈显著正相关。虽然茎光合作用较弱,但在直射辐射通量增加时,叶片光合作用合成更多有机碳,在运输过程中储存在茎中。
-
根系作为土壤和植物地上部分的枢纽,既可吸收土壤水分和矿质营养反馈植物生长,又能周转、分解土壤养分和改善土壤理化特性[28]。荆条具有浅根特性。研究区荆条根系在0~20 cm土层中所占比例最大,且由于荆条具有“肥岛”效应,其土壤养分表现为表层养分最丰富[20],因此,本研究分析荆条根系与0~20 cm土壤理化特性的关系。发达的根系能降低土壤容重,增加非毛管孔隙度,促进土壤有机质积累[29]。本研究发现:间伐样地荆条根系生物量显著高于未间伐样地,且总根长与土壤容重呈极显著负相关,与非毛管孔隙度和土壤矿质养分呈极显著正相关,这说明间伐有利于改善土壤孔隙结构,使土壤水分易于根系吸收。这与李建聪[30]研究的土壤非毛管孔隙度与根系的分布具有一致性的结果类似。荆条比根长与土壤有机碳极显著线性正相关,根有机碳与土壤有机质极显著线性正相关。这说明灌木倾向于通过比根长提高水分和养分的获取能力[13]。JACKSON等[31]认为:根系分布异质性的主要原因是土壤养分及空间异质性,根系对土壤空间异质性的基本响应是调整生物量,这说明间伐样地荆条根系趋向于资源获取效率提升。但是,由于森林经营的长周期性,随着时间的推移,冠层开度下降,林下光环境减弱,林内环境与未间伐样地的差异性缩小,因此想要探究间伐对林下荆条的长期影响,需进行进一步周期性系统研究。
-
栓皮栎人工林间伐后,荆条拥有较高叶周长、叶面积、比叶面积、叶全氮、茎有机碳、总根长、根体积、根生物量、根有机碳功能性状组合,其资源获取性生存策略突出。地上部分对光环境最敏感的功能性状有叶面积、比叶面积、叶氮质量分数和茎总有机碳质量分数。其中,林下散射辐射通量、总辐射通量和太阳直射时间是荆条地上部分功能性状最主要的影响因子。荆条根功能性状中总根长、比根长是对土壤理化特性最敏感的性状指标,而影响荆条根功能性状的关键因子是土壤有机质。
Effects of thinning on the functional traits of understory Vitex negundo var. heterophylla in Quercus variabilis plantation
-
摘要:
目的 研究间伐对栓皮栎Quercus variabilis人工林林下荆条Vitex negundo var. heterophylla根、茎、叶功能性状的影响,为人工林林分结构优化和林下幼苗更新保护提供理论基础。 方法 以间伐后3 a栓皮栎人工林灌木层荆条为研究对象,比较其叶面积、比叶面积、叶干物质质量分数、茎干物质质量分数、总根长、比根长等结构性状,以及植株各器官全氮、全磷、全钾和总有机碳质量分数等化学性状的差异性,研究植物功能性状与林下光环境及土壤理化特征的关系。 结果 间伐后荆条地上部分植物功能性状指标(叶总有机碳、叶面积、比叶面积、叶有机氮、茎总有机碳)和地下部分植物功能性状指标(总根长、根体积、根生物量、根总有机碳)均显著高于对照样地(P<0.05)。相关和逐步回归分析结果表明:林下散射辐射是影响荆条叶结构性状的主要因子,林下直射辐射通量、光合有效辐射和太阳总辐射时间是影响叶化学性状的关键因子,茎功能性状主要受林下直射辐射和散射辐射的共同作用。总根长、比根长和根有机碳与土壤非毛管孔隙度、土壤总孔隙度、土壤有机碳和土壤全氮存在显著相关(P<0.05),其中,土壤有机质是影响荆条根功能性状的关键因子。 结论 间伐能显著促进栓皮栎人工林下荆条叶和根功能性状的发育,对茎功能性状的影响较弱。图1表6参31 Abstract:Objective The objective of this study is to explore the effect of thinning on the functional traits of roots, stems and leaves of Vitex negundo var. heterophylla under Quercus variabilis plantation in order to provide theoretical basis for the optimization of plantation structure and the regeneration and protection of understory seedlings. Method The V. negundo var. heterophylla in the shrub layer of Q. variabilis plantation 3 years after thinning was taken as the research object, and the differences of leaf area, specific leaf area, leaf dry matter content, stem dry matter content, total root length, specific root length and other structural traits, as well as total nitrogen, total phosphorus, total potassium and total organic carbon content in various plant organs were compared. The relationship between plant functional traits and the understory light environment and soil physical and chemical characteristics was studied. Result After thinning, the aboveground functional traits of plants of V. negundo var. heterophylla such as total leaf organic carbon, leaf area, specific leaf area, leaf organic nitrogen, stem total organic carbon, and the underground functional traits of plants such as root length, root volume, root biomass, the total organic carbon content of roots were significantly higher than those of the control plot (P<0.05). Correlation and stepwise regression analysis results showed that scattered radiation was the main factor affecting the structural properties of V. negundo var. heterophylla leaves. Direct radiation flux, photosynthetically active radiation and total solar radiation time were the key factors affecting leaf chemical properties, while stem functional properties were mainly affected by the direct radiation and scattered radiation under the forest. Total root length, specific root length and root organic carbon content were significantly correlated with soil non-capillary porosity, soil total porosity, soil organic carbon and soil total nitrogen (P<0.05), among which soil organic matter content was the key factor affecting functional traits. Conclusion Thinning can significantly promote the development of the functional traits of leaves and roots of Q. variabilis plantation, and has a weak effect on stem functional traits. [Ch, 1 fig. 6 tab. 31 ref.] -
图 1 间伐对荆条各器官养分分配影响
Figure 1 Effect of thinning on nutrient distribution of various organs of V. negundo var. heterophylla
表 1 所选植物功能性状
Table 1. Functional traits of selected plants
植物功能性状 单位符号 植物功能性状 单位符号 植物功能性状 单位符号 植物功能性状 单位符号 高度 m 基径 cm 根干物质质量分数 茎全氮 g·kg−1 叶周长 cm 茎生物量 g 根生物量比 茎全钾 mg·kg−1 叶面积 cm2 茎干物质质量分数 根冠比 茎全磷 g·kg−1 比叶面积 cm2·g−1 主根长 cm 比根长 cm·g−1 茎有机碳 g·kg−1 叶绿素 总根长 cm 叶全氮 g·kg−1 根全氮 g·kg−1 叶生物量 g 根表面积 cm2 叶全钾 mg·kg−1 根全钾 mg·kg−1 叶干物质质量分数 根体积 cm3 叶全磷 g·kg−1 根全磷 g·kg−1 叶长宽比 根生物量 g 叶有机碳 g·kg−1 根有机碳 g·kg−1  下载: 导出CSV
下载: 导出CSV
表 2 间伐对荆条地上部分功能性状的影响
Table 2. Effect of thinning on the functional traits of aboveground part of on V. negundo var. heterophylla
项目 措施 高度/m 叶周长/cm 叶面积/cm2 比叶面积/(cm2·g−1) 叶绿素 叶生物量/g 平均值±标准差 间伐 0.97±0.03 A 48.91±1.04 A 17.27±0.49 A 31.97±0.63 A 43.55±2.50 A 90.53±9.68 A 未间伐 0.92±0.03 A 33.16±1.77 B 10.18±1.71 B 24.07±1.85 B 34.15±3.60 A 67.55±4.52 B 数值范围 间伐 0.91~1.06 46.37~52.49 15.90~18.55 30.37~33.12 37.75~52.75 66.02~112.78 未间伐 0.86~0.98 28.31~37.33 5.53~15.67 18.15~28.46 20.50~40.05 53.01~76.94 项目 措施 叶干物质质量分数 叶长宽比 基径/cm 茎生物量/g 茎干物质质量分数 平均值±标准差 间伐 0.35±0.02 A 1.60±0.06 A 1.39±0.20 A 32.91±6.82 A 0.55±0.02 A 未间伐 0.44±0.05 A 2.43±0.88 A 1.18±0.15 A 25.02±6.13 A 0.52±0.03 A 数值范围 间伐 0.30~0.38 1.45~1.76 0.60~1.69 16.64~43.28 0.47~0.59 未间伐 0.35~0.47 1.20~5.91 0.80~1.69 12.13~47.25 0.41~0.58 说明:同列不同字母表示功能性状间存在显著差异(P<0.05),相同字母表示无显著差异(P>0.05)
下载: 导出CSV
表 3 间伐对荆条地下部分功能性状的影响
Table 3. Effect of thinning on the functional traits of underground part of V. negundo var. heterophylla
项目 措施 主根长/cm 根总长/cm 根表面积/cm2 根体积/cm3 根生物量/g 平均值±标准差 间伐 9.02±0.91 A 207.30±24.40 A 701.03±180.68 A 31.79±3.28 A 43.78±7.36 A 未间伐 8.80±0.61 A 133.96±12.61 B 516.52±251.68 A 18.50±4.45 B 34.58±3.32 B 数值范围 间伐 6.6~12.10 131.42~285.56 508.90~1060.67 21.99~42.34 14.88~56.32 未间伐 7.5~10.60 108.45~169.85 275.21~865.78 9.0~33.65 27.22~42.49 项目 措施 根干物质质量分数 根生物量比 根冠比 比根长/(cm·g−1) 平均值±标准差 间伐 0.62±0.01 B 0.40±0.08 B 1.16±0.42 A 6.79±0.31 A 未间伐 0.68±0.05 A 0.52±0.05 A 1.16±0.31 A 3.97±0.41 B 数值范围 间伐 0.52~0.75 0.21~0.53 0.27~2.15 2.64~19.19 未间伐 0.56~0.82 0.39~0.69 0.44~2.28 2.69~5.22 说明:同列不同字母表示功能性状间存在显著差异(P<0.05),相同字母表示无显著差异(P>0.05)
下载: 导出CSV
表 4 荆条地上部分功能性状与林下光环境的相关性分析
Table 4. Correlation analysis of aboveground part functional traits of V. negundo var. heterophylla and understory light environment
地上部分功能性状 光合有效辐射强度 直射辐射通量 散射辐射通量 总辐射通量 太阳辐射时间 叶面积 0.534 0.442 0.823** 0.837** 0.882** 叶绿素 0.263 0.578 0.484 0.274 0.528 叶干物质质量分数 0.861 0.215 0.521 0.215 0.779 比叶面积 0.547 0.797** 0.837** 0.768** 0.661* 叶全氮 0.706* 0.304 0.492 0.902* 0.946* 叶全磷 0.180 0.071 0.186 0.552 0.620 叶全钾 0.664* 0.831** 0.311 0.752* 0.520 叶有机碳 0.663* 0.821** 0.752* 0.761* 0.940** 茎干物质质量分数 0.841** 0.018 0.488 0.831** 0.783** 茎生物量比 −0.400 0.171 −0.316 0.067 −0.141 茎全氮 0.415 0.296 0.447 0.111 0.569 茎全磷 0.209 0.160 0.244 0.031 0.435 茎全钾 0.640* 0.710* 0.769** 0.786** 0.649* 茎有机碳 0.313 0.862** 0.686 0.801** 0.669 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
表 5 荆条根功能性状与土壤因子的相关性分析
Table 5. Correlation analysis of the functional characters of V. negundo var. heterophylla root and soil factors
根功能性状 土壤容重 非毛管孔隙度 毛管孔隙度 总孔隙度 土壤有机碳 土壤全氮 土壤全磷 土壤全钾 主根长 −0.547 0.612 −0.541 −0.454 0.619 0.756* 0.432 0.357 总根长 −0.807** 0.829** −0.848** −0.791** 0.873** 0.451 −0.109 −0.155 根表面积 −0.739* 0.740* −0.672 −0.577 0.734* 0.690* 0.290 0.431 根体积 −0.914** 0.953** −0.876 −0.759 0.915** 0.604 0.175 0.139 根生物量 −0.479 0.460 0.212 0.228 0.233 0.009 −0.407 −0.120 根干物质质量分数 −0.629 0.650* −0.077 −0.016 0.378 0.634* 0.238 0.479 根生物量比 0.174 −0.158 −0.597 −0.578 −0.474 −0.031 −0.104 0.423 根冠比 −0.142 0.166 −0.663* −0.620 −0.016 0.217 0.260 0.481 比根长 −0.588 0.551 −0.834** −0.838** 0.752* 0.485 0.108 −0.074 根全氮 −0.634 0.647* −0.578 −0.540 0.586 0.505 −0.060 0.153 根全磷 −0.858** 0.717* −0.934** −0.871** 0.818** 0.265 −0.220 −0.221 根全钾 −0.813** 0.775** 0.212 0.228 0.750* 0.439 −0.234 −0.223 根有机碳 −0.916** 0.913** −0.077 −0.016 0.979** 0.433 −0.058 −0.197 说明:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)
下载: 导出CSV
表 6 荆条各器官功能性状与环境因子的逐步回归分析模型
Table 6. Stepwise regression analysis of V. negundo var. heterophylla functional traits and environmental factors each organ
功能性状 回归方程 P R2 叶面积 y=−6.89+1.282x4+0.056x5 0.026 0.896 比叶面积 y=−7.385−0.13x1+10.229x3 0.014 0.910 叶全氮 y=1.143+0.013x4+0.089x5 0.048 0.911 叶有机碳 y=77.476+77.11x1−0.794x3 0.160 0.987 茎干物质质量分数 y=0.415+0.001x1 0.002 0.706 茎有机碳 y=74.329+121.525x2 0.038 0.620 总根长 y=−118.671−2.234 x7+3.555 x8+4.312 x9 0.001 0.783 根干物质 y=−230.074+10x7−0.013x9 0.017 0.753 根体积 y=−9.176−15.724 x6+3.499 x7+0.199 x9 0.001 0.923 比根长 y=−17.111−4.765 x7+1.124x8+0.840 x9 0.003 0.930 根有机碳 y=−338.69+4.253x9−26.676x10 0.001 0.978 说明:x1为光合有效辐射;x2为林下直射辐射通量;x3为林下散射辐射通量;x4为林下总辐射通量;x5为太阳辐射时间;x6为土壤容 重;x7为土壤非毛管孔隙度;x8为毛管总孔隙度;x9为土壤有机质;x10为土壤全氮。
下载: 导出CSV
-
[1] 闫东锋, 贺文, 马瑞婷, 等. 抚育间伐对栓皮栎种群空间分布格局的影响[J]. 生态环境学报, 2020, 29(3): 429 − 437. YAN Dongfeng, HE Wen, MA Ruiting, et al. Effects of forest thinning on the spatial distribution patterns of Quercus variabilis population [J]. Ecol Environ Sci, 2020, 29(3): 429 − 437. [2] 段梦成, 王国梁, 史君怡, 等. 间伐对油松人工林优势种群结构与分布格局的影响[J]. 生态学杂志, 2019, 38(1): 1 − 10. DUAN Mengcheng, WANG Guoliang, SHI Junyi, et al. Effects of thinning on structure and spatial pattern of dominant populations in Pinus tabulifomis plantations [J]. Chin J Ecol, 2019, 38(1): 1 − 10. [3] 石君杰, 陈忠震, 王广海, 等. 间伐对杨桦次生林冠层结构及林下光照的影响[J]. 应用生态学报, 2019, 30(6): 1956 − 1964. SHI Junjie, CHEN Zhongzhen, WANG Guanghai, et al. Impacts of thinning on canopy structure and understory light in secondary poplar-birch forests [J]. Chin J Appl Ecol, 2019, 30(6): 1956 − 1964. [4] 张甜, 朱玉杰, 董希斌. 抚育间伐对大兴安岭天然用材林冠层结构及光环境特征的影响[J]. 东北林业大学学报, 2016, 44(10): 1 − 7. ZHANG Tian, ZHU Yujie, DONG Xibin. Canopy structure and light characters after tending felling in Daxing’an mountains [J]. J Northeast For Univ, 2016, 44(10): 1 − 7. [5] 李瑞霞, 马洪靖, 闵建刚, 等. 间伐对马尾松人工林林下植物多样性的短期和长期影响[J]. 生态环境学报, 2012, 21(5): 807 − 812. LI Ruixia, MA Hongjing, MIN Jiangang, et al. Short-term and long-term effects of thinning on the undergrowth diversity in the Pinus massoniana plantation [J]. Ecol Environ Sci, 2012, 21(5): 807 − 812. [6] 张象君, 王庆成, 郝龙飞, 等. 长白落叶松人工林林隙间伐对林下更新及植物多样性的影响[J]. 林业科学, 2011, 47(8): 7 − 13. ZHANG Xiangjun, WANG Qingcheng, HAO Longfei, et al. Effect of gap thinning on the regeneration and plant species diversity in Larix olgensis plantation [J]. Sci Silv Sin, 2011, 47(8): 7 − 13. [7] 刘晓东. 影响栎类苗木早期生长关键因子的试验研究[J]. 甘肃科技, 2012, 28(21): 155 − 156, 148. LIU Xiaodong. Experimental study on key factors affecting early growth of oak seedlings [J]. Gansu Sci Technol, 2012, 28(21): 155 − 156, 148. [8] 孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能[J]. 植物生态学报, 2007, 31(1): 150 − 165. MENG Tingting, NI Jian, WANG Guohong. Plant functional traits, environment and ecosystem functioning [J]. Chin J Plant Ecol, 2007, 31(1): 150 − 165. [9] 刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学: 生命科学, 2015, 45(4): 325 − 339. LIU Xiaojuan, MA Keping. Plant functional traits: concepts, applications and future directions [J]. Sci Sin Vitae, 2015, 45(4): 325 − 339. [10] 王进, 朱江, 艾训儒, 等. 湖北星斗山地形变化对不同生活型植物叶功能性状的影响[J]. 植物生态学报, 2019, 43(5): 447 − 457. WANG Jin, ZHU Jiang, AI Xunru, et al. Effects of topography on leaf functional traits across plant life forms in Xingdou Mountain, Hubei, China [J]. Chin J Plant Ecol, 2019, 43(5): 447 − 457. [11] 张秀芳, 穆振北, 林美娇, 等. 琅岐岛4种优势植物叶功能性状及其影响因子[J]. 应用与环境生物学报, 2020, 26(3): 667 − 673. ZHANG Xiufang, MU Zhenbei, LIN Meijiao, et al. Functional traits of leaves of four dominant plants on Langqi Island, Fuzhou, and factors influencing these traits [J]. Chin J Appl Environ Biol, 2020, 26(3): 667 − 673. [12] 陈延松, 李玲玲, 周守标, 等. 珍珠菜属植物功能性状的趋异分化[J]. 生态学杂志, 2019, 38(6): 1653 − 1661. CHEN Yansong, LI Lingling, ZHOU Zhoubiao, et al. The differentiation of plant functional traits in genus Lysimachia L. [J]. Chin J Ecol, 2019, 38(6): 1653 − 1661. [13] 闫东锋, 张振, 杨喜田. 豫南山区典型林分地表层根系结构与土壤特性的关系[J]. 东北林业大学学报, 2014, 42(12): 30 − 36. YAN Dongfeng, ZHANG Zhen, YANG Xitian. Root structures and its relation with top soil properties of typical forest in southern mountains in Henan Province [J]. J Northeast For Univ, 2014, 42(12): 30 − 36. [14] 孙梅, 田昆, 张贇, 等. 植物叶片功能性状及其环境适应研究[J]. 植物科学学报, 2017, 35(6): 940 − 949. SUN Mei, TIAN Kun, ZHANG Yun, et al. Research on leaf functional traits and their environmental adaptation [J]. Plant Sci J, 2017, 35(6): 940 − 949. [15] 郑芬, 李兆佳, 邱治军, 等. 广东南岭天然常绿阔叶林林下光环境对林下幼树功能性状的影响[J]. 生态学报, 2020, 40(13): 4516 − 4527. ZHENG Fen, LI Zhaojia, QIU Zhijun, et al. Effects of understory light on functional traits of evergreen broad-leaved forest saplings in Nanling Mountains, Guangdong Province [J]. Acta Ecol Sin, 2020, 40(13): 4516 − 4527. [16] 陈嘉欣, 张玲玲, 张国庆, 等. 6种园林植物耐旱性分析[J]. 热带亚热带植物学报, 2020, 28(3): 310 − 316. CHEN Jiaxin, ZHANG Lingling, ZHANG Guoqing, et al. Drought tolerance of six garden species [J]. J Trop Subtrop Bot, 2020, 28(3): 310 − 316. [17] 刘雅辰, 赵琛迪, 杨子, 等. 太行山南麓不同龄级荆条光合特性及光响应研究[J]. 河南农业大学学报, 2020, 54(2): 203 − 208, 230. LIU Yachen, ZHAO Chendi, YANG Zi, et al. Study on photosynthetic characteristics and light response of Vitex negundo of different age classesat the southern foot of Taihang Mountain [J]. J Henan Agric Univ, 2020, 54(2): 203 − 208, 230. [18] 闫东锋, 马瑞婷, 杨庆培, 等. 间伐强度对栎类天然次生林幼苗更新的影响[J]. 河南农业大学学报, 2019, 53(2): 187 − 192. YAN Dongfeng, MA Ruiting, YANG Qingpei, et al. Effect of thinning intensity on seedling regeneration characteristics in natural secondary oak forests [J]. J Henan Agric Univ, 2019, 53(2): 187 − 192. [19] 韩文娟, 袁晓青, 张文辉. 油松人工林林窗对幼苗天然更新的影响[J]. 应用生态学报, 2012, 23(11): 2940 − 2948. HAN Wenjuan, YUAN Xiaoqing, ZHANG Wenhui. Effects of gap size on seedling natural regeneration in artificial Pinus tabulaeformis plantation [J]. Chin J Appl Ecol, 2012, 23(11): 2940 − 2948. [20] 赵河, 张志铭, 赵勇, 等. 模拟氮沉降对荆条灌木‘肥岛’土壤养分的影响[J]. 生态学报, 2017, 37(18): 6014 − 6020. ZHAO He, ZHANG Zhiming, ZHAO Yong, et al. Effects of simulated nitrogen deposition on soil nutrients of Vitex negundo L. ‘fertile islands’ [J]. Acta Ecol Sin, 2017, 37(18): 6014 − 6020. [21] 商添雄, 韩海荣, 程小琴, 等. 华北落叶松人工林生长对抚育间伐的响应及其与土壤因子的关系[J]. 林业科学研究, 2019, 32(6): 40 − 47. SHANG Tianxiong, HAN Hairong, CHENG Xiaoqin, et al. Response of Larix principis-rupprechtii plantation growth to thinning and its relationship with soil factors [J]. For Res, 2019, 32(6): 40 − 47. [22] 路兴慧, 丁易, 臧润国, 等. 海南岛热带低地雨林老龄林木本植物幼苗的功能性状分析[J]. 植物生态学报, 2011, 35(12): 1300 − 1309. LU Xinghui, DING Yi, ZANG Runguo, et al. Analysis of functional traits of woody plant seedlings in an old-growth tropical lowland rainforest on Hainan Island, China [J]. Chin J Plant Ecol, 2011, 35(12): 1300 − 1309. [23] OSADA N, OKABE Y, HAYASHI D, et al. Differences between height- and light-dependent changes in shoot traits in five deciduous tree species [J]. Oecologia, 2013, 174(1): 1 − 12. [24] LHOTKA J M. LOEWENSTEIN E F. Indirect measures for characterizing light along a gradient of mixed-hardwood riparian forest canopy structures [J]. For Ecol Manage, 2006, 226(1/3): 310 − 318. [25] CORNELISSEN J H C, LAVOREL S, GARNIER E B, et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide [J]. Aust J Bot, 2003, 51(4): 335 − 380. [26] HOGAN J A, HERAULT B, BACHELOT B, et al. Understanding the recruitment response of juvenile neotropical trees to logging intensity using functional traits [J]. Ecol Appl, 2018, 28(8): 1998 − 2010. [27] 高林浩, 孙晗, 白雪卡, 等. 气候、系统发育对长白山乔灌木比叶面积与叶元素含量关系的影响[J]. 北京林业大学学报, 2020, 42(2): 19 − 30. GAO Linhao, SUN Han, BAI Xueka, et al. Effects of climate and phylogeny on the relationship between specific leaf area and leaf element concentration of trees and shrubs in Changbai Mountain of northeastern China [J]. J Beijing For Univ, 2020, 42(2): 19 − 30. [28] 欧延升, 汪霞, 李佳, 等. 不同恢复年限人工草地土壤碳氮磷含量及其生态化学计量特征[J]. 应用与环境生物学报, 2019, 25(1): 38 − 45. OU Yansheng, WANG Xia, LI Jia, et al. Content and ecological stoichiometry characteristics of soil carbon, nitrogen, and phosphorus in artificial grassland under different restoration years [J]. Chin J Appl Environ Biol, 2019, 25(1): 38 − 45. [29] 沈晶玉, 周心澄, 张伟华, 等. 祁连山南麓植物根系改善土壤抗冲性研究[J]. 中国水土保持科学, 2004, 2(4): 87 − 91. SHEN Jingyu, ZHOU Xincheng, ZHANG Weihua, et al. Effects of plant root system on the anti-scourability of soil in the south of Qilian Mountain [J]. Sci Soil Water Conserv, 2004, 2(4): 87 − 91. [30] 李建聪. 北京西山酸枣灌木林对土壤物理性质的影响[J]. 内蒙古林业科技, 2015, 41(3): 14 − 17. LI Jiancong. Effects of Ziziphus jujube on soil physical properties in West Mountains of Beijing [J]. J Inn Mongolia For Sci Technol, 2015, 41(3): 14 − 17. [31] JACKSON R B, CANADELL J R, MOONEY H A, et al. A global analysis of root distribution for terrestri biomass [J]. Oecologia, 1996, 108(3): 389 − 411. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200551
 点击查看大图
点击查看大图

计量
- 文章访问数: 1806
- HTML全文浏览量: 654
- PDF下载量: 50
- 被引次数: 0
