




-
大量栖息地的破坏导致了生态系统的退化,进而造成生物多样性损失和生态系统功能的降低[1]。中国许多地区均开展了公益林建设和森林管理项目,公益林面积达到了8500万hm2(占中国森林面积的41.6%)[2],以期降低生物群落受生态系统退化的影响[3]。浙江拥有阔叶林、针阔混交林以及针叶林3种主要森林类型的公益林群落,具体包括天然次生林及部分人工种植林[4]。次生林和人工种植林缓解了生物多样性和生态系统功能的衰退[5],但是了解维持生态系统功能所需要的生物多样性的数量和类型仍然是公益林建设和管理的关键问题[6-9]。近来有研究指出,相对于传统的生物多样性,基于进化信息和功能属性的谱系多样性能更好地推测生态系统过程及预测生态系统功能[10]。比如,谱系多样性能很好地解释热带森林早期演替阶段的生物量[11]。类似地,CADOTTE等[12]发现:群落中若是聚集亲缘关系较远的物种可通过生态位互补产生更多的生物量。因此,探索谱系多样性和生态系统服务(如生物量积累)之间的关系,对于缓解气候变化和理解生态系统的功能具有重要意义[13-15]。大多数关于公益林的研究集中于传统的植物多样性[16],极少有研究定量地测量公益林不同森林类型的生物多样性,尤其是谱系多样性和生物量之间的关系。本研究主要将进化历史(谱系多样性)、生态系统和功能与不同森林类型的公益林联系起来,首次探索公益林建设过程中谱系多样性和物种丰富度与树木地上生物量之间的关系。除了群落内的物种谱系和物种多样性,森林群落生态系统功能也被环境条件影响[17-18],其中,非生物因素(如土壤肥力、海拔等)是局部区域内物种多样性和生物量的关键驱动因子[19-21]。在群落演替早期,充足的土壤肥力可以促进生态位分化[22],进而增加物种多样性和加快种群更新。具体来说,速生、资源获取型和喜阳树种逐渐被生长缓慢、资源保守型和耐荫树种取代[11]。此外,不同的森林群落类型拥有不同的群落结构和物种组成,因此土壤的化学组成也会发生变化[23]。有研究发现:当杉木Cunninghamia lanceolata人工林转化成另一种阔叶树种人工林后,土壤养分状况得到了改善[24]。然而,环境因子和生物多样性是如何影响不同森林类型的生态系统功能仍需要深入研究。森林群落的恢复需要数十年的时间,尤其是在林分破坏严重的群落中,因此预测生产力格局更显得至关重要[25]。本研究选取浙江省部分公益林中主要的3种森林类型为研究对象,测定群落内的环境因子以及谱系多样性和物种丰富度,以期更好地理解生物多样性和环境因子对公益林不同林型生态系统生物量的影响,主要探讨:①公益林不同森林类型中物种丰富度和谱系多样性如何影响生物量?②在环境因子(海拔、坡度、坡向、土壤厚度、土壤吸湿水、土壤pH、土壤有机质、土壤速效氮、土壤速效磷和土壤速效钾)作用下,物种丰富度和谱系多样性对生物量的影响如何?通过研究上述问题,为浙江省不同森林类型公益林森林生态系统管理以及生态系统功能研究提供理论支撑。
-
研究区域为浙江省淳安县(29°11′~30°02′N, 118°20′~119°20′E)、建德市(29°12′~29°46′N, 118°53′~119°45′E)和磐安县(28°49′~29°19′N, 120°17′~120°47′E)。淳安县最高海拔为1523 m,建德市大部分地区海拔为1 000 m以下,磐安县的最高海拔为1314 m。淳安县、磐安县和建德市拥有相似的气候(亚热带季风气候),平均年降水量分别为1430.0、1 600.0和1 468.8 mm。淳安县年平均气温为17.0 ℃,1月平均气温为5.0 ℃,7月平均气温为28.9 ℃,年均日照时数为1 951.0 h。磐安县年平均气温为15.7 ℃,1月气温为2.0~4.3 ℃,7月气温为25.6~28.8 ℃,无霜期为200.0~243.0 d,该地气候性灾害多发,对植被干扰较大。建德市年平均气温为17.4 ℃,年均日照时数1760.0 h。浙江省物种类型丰富,主要优势种有木荷Schima superba、青冈Cyclobalanopsis glauca、润楠Machilus nanmu和杉木[3-4, 16],并存在多种森林类型,主要包括常绿及落叶阔叶林、常绿落叶阔叶混交林、针阔混交林和针叶林等[26]。
-
本研究依托2015−2018年浙江省公益林数据,根据样方调查记载,主要选择能涵盖不同环境条件和生物多样性水平的森林样方作为研究对象。选取197个20 m×20 m的固定监测样方,其中淳安县28个、建德市117个、磐安县53个。样地海拔均在1 000 m以下。淳安县的样方全坡均有分布,建德市和磐安县的样地分布坡度为0°~50°。利用样方群落恢复历史来代替林龄,具体根据样方所在林区斑块内的砍伐记录确定,且各个林龄段均有分布。调查并鉴定了所有胸径(DBH)≥5 cm的存活木本植物,并记录胸径、树高和坐标。每个样方所代表的群落类型利用重要值公式将其划分为3种类型[3]:针叶林、针阔混交林和阔叶林。针叶树种的重要值>66.7%,则定义为针叶林(n=49);若重要值介于33.3%和66.7%之间则为混交林(n=69);若重要值<33.3%(则阔叶树种的重要值>66.7%),则样方被定义为阔叶林(n=79,表1)。本研究不包括纯林种植地。
表 1 公益林3种森林类型样方概况
Table 1. Summary of three forest types of welfare forests
森林类型 样方数/个 优势树种 物种数/种 林龄范围/a 针叶林 49 杉木、马尾松Pinus massoniana 61 0~50 针阔混交林 69 杉木、马尾松、木荷和青冈 96 0~52 阔叶林 79 石栎Lithocarpus glaber、青冈、白栎Quercus fabri和苦槠Castanopsis sclerophylla 89 0~60 -
环境因素包括样方的4个地形因子,即海拔、坡度、坡向和土壤厚度,以及6个土壤因子,即土壤吸湿水、土壤pH、土壤有机质、土壤速效氮、土壤速效磷和土壤速效钾。在每个样方上使用三点法在土壤0~10 cm层收集土样,每个点使用容积为100 cm3的土壤环刀采集3个样品用于测定土壤物理性质,且在每个点用直径3.5 cm的10.0 cm土钻采集3份土样,混合均匀后放入密封袋在实验室测定土壤化学性质。具体是将风干土壤样品置于105 ℃烘箱中恒温6 h烘干,并计算土壤吸湿水;土壤pH采用1.0∶2.5土水比,利用pH计测定;土壤有机质质量分数采用水合热重铬酸钾氧化-比色法测定;土壤速效氮采用碱解扩散法测定;土壤速效钾采用火焰光度法测定;土壤速效磷采用钼锑抗比色法测定[27]。
-
参照The Plant List (
http://www.theplantlist.org/ )以及中国植物志网络版(http://www.iplant.cn/frps )确定物种名是否符合被子植被分类系统(Angiosperm Phylogeny Group Ⅲ)[28]。采用Phylomatic在线工具生成的Newick格式的谱系进化树,每个进化树基于ZANNE等[29]为骨架构建。 -
采用物种丰富度(species richness)代表样方的物种多样性,采用vegan包计算[30]。谱系多样性则使用Faith谱系多样性(Faith’s phylogenetic diversity, DP),其提供沿谱系树的进化历史量[31-33],使用picante包中的psd函数计算[34]。计算公式如下: $ D_{\rm{P}}=\sum\nolimits_{\left\{\boldsymbol{c}\in \boldsymbol{C}\right\}}L_c $。其中:Lc为物种谱系树的分支长度,c为连接谱系树节点的一段分支,C为连接谱系树上所有物种最短路径的所有分支长度总和。
-
使用每个样方中所有乔木个体的生物量来评估群落中的生态系统功能,通过袁位高等[35]提出的浙江省公益林生物量模型计算。首先将乔木树种划分为松类、杉类、硬阔(Ⅰ和Ⅱ)、软阔、毛竹Phyllostachys edulis和杂竹共7个树种组,其次利用树高、胸径和枝下高代入相应模型计算树木各组生物量并求和。折断和枯死木不包括在分析中。
-
为探究环境因子和生物多样性对生物量的影响,采用多元线性模型分析影响不同林型样方内生物量的主要因子。本研究利用MASS包中glm.nb函数来拟合广义线性模型。由于生物量数据存在显著过度离散,使用负二项分布族的对数(log)联结拟合,并利用vif函数对解释变量进行共线性分析,发现所有解释变量间的方差膨胀因子(variance inflation factor, VIF)均小于5,说明变量间无显著共线性[36]。使用MuMIN包的dredge函数构建所有不同解释变量组合的模型来决定何种解释变量对预测生物量最重要,随后根据赤池信息准则(Akaike’s information criterion,AIC)选择最低AIC值的模型[37]。最后,使用同样的模型分别观察每种林型内谱系多样性、物种丰富度和生物量的关系。以上分析均在R3.6.0中进行。
-
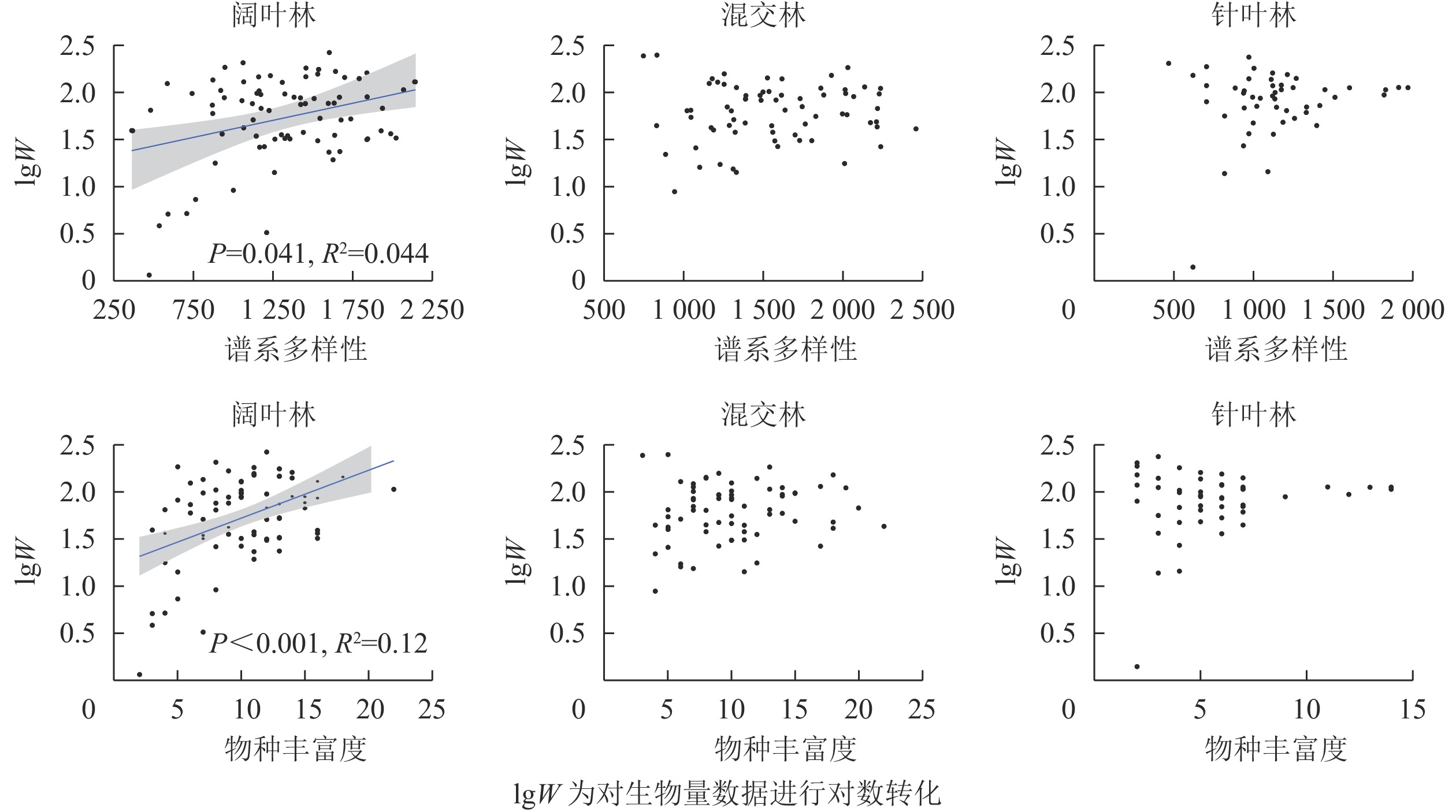
197个公益林不同森林类型样方中总计测量了13867株121种植物的谱系多样性和物种丰富度。混交林具有最高的谱系多样性,随后是阔叶林和针叶林(图1A);阔叶林和混交林拥有相近的物种丰富度且明显高于针叶林(图1B);3种森林类型的生物量无显著差异,其中针叶林的平均生物量最高,其次为阔叶林和混交林(图1C)。谱系多样性比物种丰富度能更好地区分不同的森林类型。
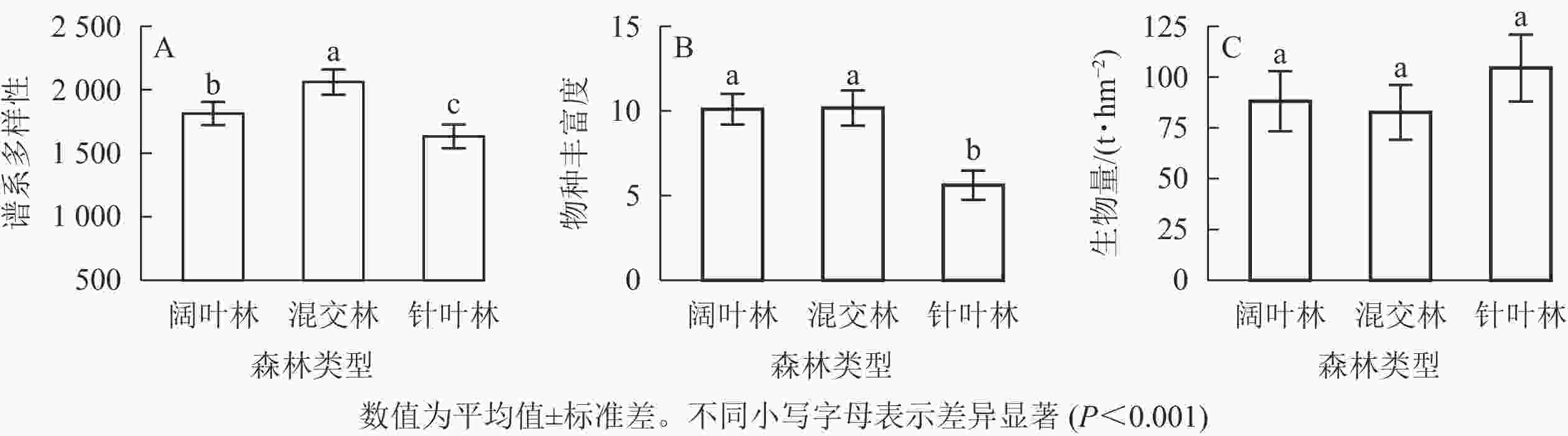
图 1 3种森林类型中谱系多样性、物种丰富度和生物量差异
Figure 1. Phylogenetic diversity, species richness and biomass per plot in three forest types
-
不考虑环境因子时,谱系多样性(R2=0.044,P=0.041)和物种丰富度(R2=0.12,P<0.001)仅显著影响阔叶林的生物量(图2),而在混交林和针叶林中,谱系多样性和物种丰富度对生物量的影响均不显著(P>0.05)。同时考虑环境因子和生物多样性对生物量的影响时,谱系多样性[Z=−2.413(Z值为用于检验相应估计值为0的假设的Wald统计量), P=0.016]、物种丰富度(Z=4.084, P<0.001)、土壤速效氮(Z=2.19, P=0.029)、土壤厚度(Z=2.366, P=0.018)和土壤吸湿水(Z=−3.16, P=0.002)在阔叶林中对生物量呈不同程度的显著影响(表2)。土壤厚度对混交林中生物量为不显著影响(Z=1.679, P=0.093)。在针叶林中,土壤厚度(Z=3.325, P<0.001)和土壤酸碱度(Z=−2.481, P=0.013)对生物量呈显著影响。
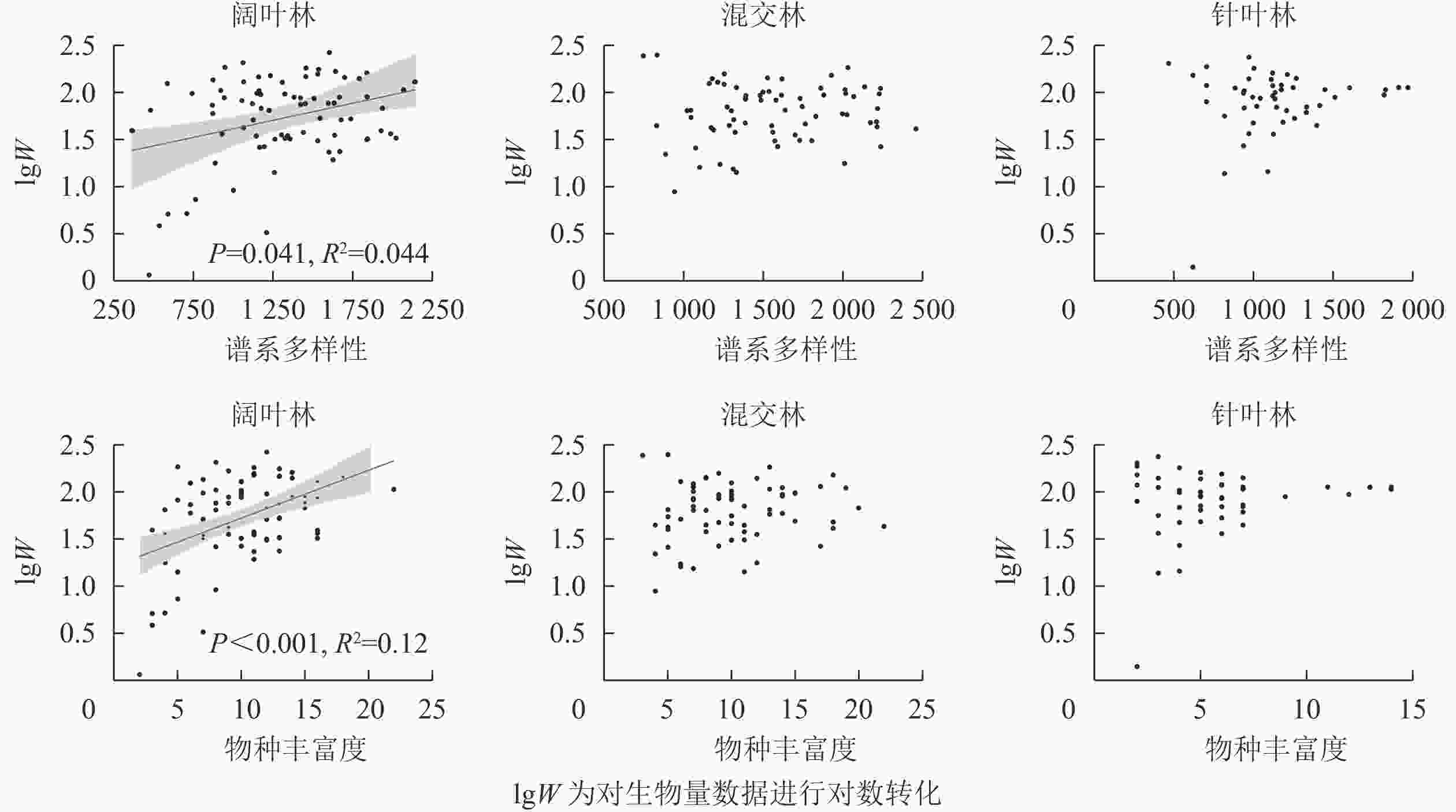
图 2 公益林3种森林类型谱系多样性、物种丰富度与生物量的关系
Figure 2. Relationship between phylogenetic diversity, species richness and above-ground biomass within each forest type
表 2 不同森林类型的生物量与生物多样性、环境因子之间的关系
Table 2. Relationship between total above-ground biomass (AGB) and biodiversity, environmental factors within each forest type
森林类型 解释因子 估计系数 标准误 Z 显著度 阔叶林 截距 1.849 8 0.426 5 4.337 <0.001*** 谱系多样性 −0.001 0 0.000 4 −2.413 0.016* 物种丰富度 0.164 8 0.040 3 4.084 <0.001*** 土壤速效氮 0.002 9 0.001 3 2.190 0.029* 土壤厚度 0.010 8 0.004 5 2.366 0.018* 土壤吸湿水 0.424 8 0.137 6 −3.160 0.002* 混交林 截距 1.701 2 0.253 9 6.700 <0.001*** 土壤厚度 0.008 5 0.005 1 1.679 0.093 针叶林 截距 3.965 4 0.940 7 4.216 <0.001*** 土壤厚度 0.014 9 0.004 5 3.325 <0.001*** 土壤酸碱度 −0.458 1 0.184 7 −2.481 0.013* 坡度 −0.011 2 0.006 3 −1.775 0.076 说明:*P<0.05, **P<0.01, ***P<0.001;Z值为用于检验相应估计值为0的假设的Wald统计量 -
本研究发现:公益林中的针阔混交林和阔叶林拥有更高的谱系多样性和更多亲缘关系较远的物种。在物种丰富的阔叶林中,谱系多样性和物种丰富度均能较好地预测生物量,验证了生态位互补假说对植物多样性和生态系统功能的重要影响,这与之前生物多样性和生物量的研究一致[38-39]。而在热带森林中,SATDICHANH等[38]发现谱系多样性仅在低龄林中与生物量相关。此外,OUYANG等[40]发现:不同演替阶段的群落生物量与谱系多样性无关,这或许与本研究选用的林分划分依据为物种组成有关。在部分生态系统研究中,谱系多样性对生态系统功能的预测要弱于物种丰富度,如VENAIL等[41]对草原的研究也得出了相似的结论。此外,车盈等[42]发现:谱系多样性仅在小尺度上对群落生产力的预测要大于物种多样性,这或许是因为其研究对象为典型阔叶红松Pinus koraiensis林,而本研究包括多种森林类型的次生林;另外,其对生态系统功能计算方式也与本研究不同。因此,我们推测,生物多样性与生物量之间的关系会因为森林群落的物种组成和划分标准而存在差异。
生物多样性和环境因子如何维持以及提升生态系统功能的问题,在全球范围内引起了广泛的关注。本研究发现:当同时考虑环境因子和生物多样性时,针阔混交林和针叶林中的生物量仅与环境因子有关,而在阔叶林中环境因子和生物多样性共同对生物量有显著的影响。该结果与干扰下的大山冲森林公园研究结果[40]不一致,该研究发现谱系多样性和物种丰富度均与生物量没有显著关系,但是在加入环境因子的混合模型后,物种丰富度与生物量存在显著相关。这可能是因为本研究所选的森林群落包含各林龄段的群落(0~60 a),随着物种丰富度的增加,群落中的生物量逐渐增加。意外的是,在环境因子和生物多样性的共同作用下,阔叶林中谱系多样性与生物量呈显著负相关,或许由于谱系多样性包含物种间的亲缘关系信息,谱系多样性上升也往往意味着部分高生物量的先锋树种被后期耐荫树种所代替,导致生物量降低[43]。本研究的针叶林以杉木为主要树种,对土壤养分有较大需求,因此土壤较厚时,杉木能更有效利用土壤资源。然而,生境筛选的重要性随着森林群落的改变而下降。本研究也印证了这点。之前的研究表明:海拔是影响森林生物量的主要环境因素之一,并且显著影响了物种多样性和物种分布[39]。考虑到本研究中的样地海拔高度均处于1000 m以下,这或许会影响研究结果。
最近的研究对谱系多样性还是物种丰富度能更好地预测生态系统功能大有争议。有研究表明:物种丰富度并不是一个很好的预测因子[38]。而在本研究中,物种丰富度和谱系多样性对阔叶林中的生物量有相似的显著性预测,其中,单独的物种丰富度及考虑其他环境因子均与生物量存在显著的正线性关系。许多研究都有相似的结论,如吴初平等[39]和YUAN等[44]研究表明:物种丰富度能较好地预测森林生态系统功能。本研究进一步发现:在阔叶林中,物种丰富度对生物量有较好的预测性。此外,谱系多样性对生物量的解释力还会因为其他环境因子的作用而不同,因此,需要更加深入的研究来验证进化历史对生态系统功能的影响。
-
对浙江省公益林不同森林类型进行生态系统功能的研究结果表明:物种丰富度、谱系多样性和环境因子对生物量的影响是显著不同的,这种复杂性或许解释了生物多样性和生物量之间存在多种关系。本研究揭示非生物因素和生物因素共同影响了公益林在发展过程中的生物量。此外,谱系多样性和物种丰富度均能解释阔叶林的生物量变化。为了提高浙江省公益林的生物量,建议改善及提升针阔混交林和针叶林的土壤肥力,而对阔叶林可以进行适当的物种管理,改善物种结构。
Impact of biodiversity and site factors on biomass of public welfare forests in Zhejiang Province
-
摘要:
目的 探究浙江省公益林生物多样性和立地因子对生物量的影响,研究公益林群落结构的稳定性。 方法 依托浙江省3个县的公益林调查数据,探索10个土壤和地形因子(土壤吸湿水、土壤pH、土壤有机质、土壤速效氮、土壤速效磷、土壤速效钾、海拔、坡度、坡向和土壤厚度)以及生物多样性(物种丰富度和谱系多样性)对3种森林类型(针叶林、针阔混交林和阔叶林)生物量的影响。 结果 谱系多样性较物种丰富度能更好地区分森林类型,其中阔叶林和混交林有较高的生物多样性,针叶林则拥有高生物量。仅考虑单独因子的作用,谱系多样性(P=0.041)和物种丰富度(P<0.001)在阔叶林中对生物量有显著的积极影响;而考虑环境因子的效应时,物种丰富度、谱系多样性、土壤速效氮、土壤厚度和土壤吸湿水对阔叶林中的生物量具有显著影响(P<0.05),土壤厚度和土壤酸碱度对针叶林的生物量具有显著影响(P<0.05)。谱系多样性在环境因子的共同作用下对生物量有消极影响。 结论 生物多样性和环境因素共同影响浙江公益林的生物量。在未来公益林的经营中,应对不同森林类型采取相应的措施,增加针阔混交林和针叶林的土壤肥力,改善阔叶林的物种结构,以期更好地维持和提升公益林的生态系统功能。图2表2参44 Abstract:Objective This research aims to explore the impact of biodiversity and site factors on biomass of public welfare forest in Zhejiang Province, and to study the stability of its community structure. Method Based on the survey data of public welfare forest from three counties in Zhejiang, the effects of ten soil and terrain factors (soil hygroscopic water, soil pH, soil organic matter, soil available nitrogen, soil available phosphorus, soil available potassium, altitude, slope, aspect and soil thickness) and biodiversity (species richness and phylogenetic diversity) on the biomass of three forest types (coniferous forest, coniferous and broad-leaved mixed forest, and broad-leaved forests) were explored. Result Compared with species richness, phylogenetic diversity was better to distinguish forest types, in which broad-leaved forest and mixed forest had higher biodiversity, and coniferous forest had higher biomass. When only considering the effects of individual factors, phylogenetic diversity (P=0.041) and species richness (P< 0.001) were significantly and positively correlated with biomass in broad-leaved forests. When considering the effects of environmental factors, species richness, phylogenetic diversity, soil available nitrogen, soil thickness and soil hygroscopic water had significant effects (P<0.05) on biomass of broad-leaved forests, while soil thickness and soil pH had significant effects (P< 0.05) on biomass of coniferous forests. Phylogenetic diversity was negatively correlated with biomass due to the joint effect of environmental factors. Conclusion Environmental factors and biodiversity jointly affect the biomass of public welfare forest in Zhejiang Province. In the future management of public welfare forest, measures should be taken to increase soil fertility of coniferous and broad-leaved mixed forest, and improve the species structure of broad-leaved forest, so as to better maintain and enhance the ecosystem function of public welfare forest. [Ch, 2 fig. 2 tab. 44 ref.] -
Key words:
- forest ecology /
- phylogenetic diversity /
- species richness /
- biomass /
- environment factors /
- forest type /
- public welfare forest
-
图 1 3种森林类型中谱系多样性、物种丰富度和生物量差异
Figure 1 Phylogenetic diversity, species richness and biomass per plot in three forest types
图 2 公益林3种森林类型谱系多样性、物种丰富度与生物量的关系
Figure 2 Relationship between phylogenetic diversity, species richness and above-ground biomass within each forest type
表 1 公益林3种森林类型样方概况
Table 1. Summary of three forest types of welfare forests
森林类型 样方数/个 优势树种 物种数/种 林龄范围/a 针叶林 49 杉木、马尾松Pinus massoniana 61 0~50 针阔混交林 69 杉木、马尾松、木荷和青冈 96 0~52 阔叶林 79 石栎Lithocarpus glaber、青冈、白栎Quercus fabri和苦槠Castanopsis sclerophylla 89 0~60  下载: 导出CSV
下载: 导出CSV
表 2 不同森林类型的生物量与生物多样性、环境因子之间的关系
Table 2. Relationship between total above-ground biomass (AGB) and biodiversity, environmental factors within each forest type
森林类型 解释因子 估计系数 标准误 Z 显著度 阔叶林 截距 1.849 8 0.426 5 4.337 <0.001*** 谱系多样性 −0.001 0 0.000 4 −2.413 0.016* 物种丰富度 0.164 8 0.040 3 4.084 <0.001*** 土壤速效氮 0.002 9 0.001 3 2.190 0.029* 土壤厚度 0.010 8 0.004 5 2.366 0.018* 土壤吸湿水 0.424 8 0.137 6 −3.160 0.002* 混交林 截距 1.701 2 0.253 9 6.700 <0.001*** 土壤厚度 0.008 5 0.005 1 1.679 0.093 针叶林 截距 3.965 4 0.940 7 4.216 <0.001*** 土壤厚度 0.014 9 0.004 5 3.325 <0.001*** 土壤酸碱度 −0.458 1 0.184 7 −2.481 0.013* 坡度 −0.011 2 0.006 3 −1.775 0.076 说明:*P<0.05, **P<0.01, ***P<0.001;Z值为用于检验相应估计值为0的假设的Wald统计量
下载: 导出CSV
-
[1] CHASE J M, BLOWES S A, KNIGHT T M,et al. Ecosystem decay exacerbates biodiversity loss with habitat loss [J]. Nature, 2020, 584: 238 − 243. [2] FAO. China-Global Forest Resources Assessment 2015-Country Report [EB/OL]. (2015-01-01) [2021-04-11]. http://www.fao.org/documents/card/en/c/a6afe3a7-0fb6-4920-bb18-ae3465c2f0a9/. [3] WU Chuping, VELLEND M, YUAN Weiguo, et al. Patterns and determinants of plant biodiversity in non-commercial forests of eastern China[J]. PLoS One, 2017, 12(11). doi: 10.1371/journal.pone.0188409. [4] 沈爱华, 袁位高, 张骏, 等. 浙江省生态公益林物种多样性时空格局研究[J]. 浙江林业科技, 2014, 34(1): 1 − 6. SHEN Aihua, YUAN Weigao, ZHANG Jun,et al. Spatial-temporal patterns of species diversity in ecological forests of Zhejiang [J]. J Zhejiang For Sci Tech, 2014, 34(1): 1 − 6. [5] 钱逸凡, 伊力塔, 钭培民, 等. 浙江缙云公益林生物量及固碳释氧效益[J]. 浙江农林大学学报, 2012, 29(2): 257 − 264. QIAN Yifan, Yilita, DOU Peimin,et al. Biomass and carbon fixation with oxygen release benefits in an ecological service forest of Jinyun County, China [J]. J Zhejiang A&F Univ, 2012, 29(2): 257 − 264. [6] BROSE U, HILLEBRAND H. Biodiversity and ecosystem functioning in dynamic landscapes[J]. Philos Trans Royal Soc B Biol Sci, 2016, 371(1694). doi: 10.1098/rstb.2015.0267. [7] EVANS E W. Biodiversity, ecosystem functioning, and classical biological control [J]. Appl Entomol Zool, 2016, 51(2): 173 − 184. [8] LOREAU M, NAEEM S, INCHAUSTI P,et al. Biodiversity and ecosystem functioning: current knowledge and future challenges [J]. Science, 2001, 294(5543): 804 − 808. [9] SUTHERLAND W J, FRECKLETON R P, CHARLES H,et al. Identification of 100 fundamental ecological questions [J]. J Ecol, 2013, 101(1): 58 − 67. [10] CAVENDER-BARES J, KOZAK K H, FINE P V A,et al. The merging of community ecology and phylogenetic biology [J]. Ecol Lett, 2009, 12(7): 693 − 715. [11] LASKY J R, URIARTE M, BOUKILI V,et al. The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession [J]. Ecol Lett, 2014, 17(9): 1158 − 1167. [12] CADOTTE M W. Experimental evidence that evolutionarily diverse assemblages result in higher productivity [J]. Proc Natl Acad Sci, 2013, 110(22): 8996 − 9000. [13] LARUE E A, CHAMBERS S M, EMERY N C. Eco-evolutionary dynamics in restored communities and ecosystems [J]. Restoration Ecol, 2017, 25(1): 19 − 26. [14] DAVIES T J, URBAN M, RAYFIELD B,et al. Deconstructing the relationships between phylogenetic diversity and ecology: a case study on ecosystem functioning [J]. Ecology, 2016, 97(9): 2212 − 2222. [15] SANTOS A M C, CIANCIARUSO M, BARBOSA A M,et al. Current climate, but also long-term climate changes and human impacts, determine the geographic distribution of European mammal diversity [J]. Global Ecol Biogeogr, 2020, 29(10): 1758 − 1769. [16] 王立竹, 于晓鹏, 管杰然, 等. 浙江缙云县公益林群落植物多样性及生物量动态分析[J]. 生态科学, 2018, 37(4): 147 − 153. WANG Lizhu, YU Xiaopeng, GUAN Jieran,et al. Plant diversity and biomass dynamics of the public-welfare forest in Jinyun County, Zhejiang Province [J]. Ecol Sci, 2018, 37(4): 147 − 153. [17] RUIZ-BENITO P, RATCLIFFE S, ZAVALA M A,et al. Climate- and successional-related changes in functional composition of European forests are strongly driven by tree mortality [J].Global Change Biol, 2017, 23(10): 4162 − 4176. [18] ALI A, YAN Enrong, CHANG S X,et al. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests [J]. Sci Total Environ, 2017, 574: 654 − 662. [19] BECKNELL J M, POWERS J S. Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest [J]. Can J For Res, 2014, 44(6): 604 − 613. [20] PEÑA-CLAROS M, POORTER L, ALARCON A,et al. Soil effects on forest structure and diversity in a moist and a dry tropical forest [J]. Biotropica, 2012, 44(3): 276 − 283. [21] van der SANDE M, PEÑA-CLAROS M, ASCARRUNZ N L,et al. Abiotic and biotic drivers of biomass change in a Neotropical forest [J]. J Ecol, 2017, 105(5): 1223 − 1234. [22] COOMES D A, KUNSTLER G, CANHAM C D,et al. A greater range of shade-tolerance niches in nutrient-rich forests: an explanation for positive richness-productivity relationships? [J]. J Ecol, 2009, 97(4): 705 − 717. [23] 闫东锋, 郭丹丹, 吴桂藏, 等. 栎类天然次生林不同组分及土壤碳氮分布对森林抚育的响应[J]. 浙江农林大学学报, 2017, 34(2): 215 − 224. YAN Dongfeng, GUO Dandan, WU Guizang,et al. Carbon and nitrogen distribution with forest tending in a natural secondary oak forest [J]. J Zhejiang A&F Univ, 2017, 34(2): 215 − 224. [24] 王涛, 万晓华, 程蕾, 等. 杉木采伐迹地营造阔叶树种对土壤微生物生态化学计量特征的影响[J]. 应用生态学报, 2020, 31(11): 3851 − 3858. WANG Tao, WANG Xiaohua, CHENG Lei,et al. Effects of broadleavedtree species on soil microbial stoichiometry in clear-cut patches of Cunninghamia lanceolata plantation [J]. Chin J Appl Ecol, 2020, 31(11): 3851 − 3858. [25] KHALIL M, GIBSON D J, BAER S G. Phylogenetic diversity reveals hidden patterns related to population source and species pools during restoration [J]. J Appl Ecol, 2017, 54(1): 91 − 101. [26] 袁位高. 浙江省生态公益林主要群落结构的比较研究[D]. 北京: 中国林业科学研究院, 2009. YUAN Weigao. Comparative Studies on Structure of Main Forest Type of Ecological Service Forest in Zhejiang Province[D]. Beijing: Chinese Academy of Forestry, 2009. [27] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. [28] The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG Ⅲ [J]. Bot J Linn Soc, 2009, 161(2): 105 − 121. [29] ZANNE A E, TANK D C, CORNWELL W K,et al. Three keys to the radiation of angiosperms into freezing environments [J]. Nature, 2014, 506(7486): 89 − 92. [30] OKSANEN J, BLANCHET F G, FRIENDLY M, et al. Package “vegan”: Community Ecology Package. R package version 2.5–6, 2019[CP/OL]. (2020-11-12) [2020-12-12]. http://CARN.R-project.org/package=vegan. [31] FAITH D P. Conservation evaluation and phylogenetic diversity [J]. Biol Conserv, 1992, 61(1): 1 − 10. [32] TUCKER C M, CADOTTE M W, CARVALHO S B,et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology [J]. Biol Rev, 2017, 92(2): 698 − 715. [33] HELMUS M R, BLAND T J, WILLIAMS C K,et al. Phylogenetic measures of biodiversity [J]. Am Nat, 2007, 169(3): 68 − 83. [34] KEMBEL S W, COWAN P D, HELMUS M R,et al. Picante: R tools for integrating phylogenies and ecology [J]. Bioinformatics, 2010, 26(11): 1463 − 1464. [35] 袁位高, 江波, 葛永金, 等. 浙江省重点公益林生物量模型研究[J]. 浙江林业科技, 2009, 29(2): 1 − 5. YUAN Weigao, JIANG Bo, GE Yongjin,et al. Study on biomass model of key ecological forest in Zhejiang Province [J]. J Zhejiang For Sci Tech, 2009, 29(2): 1 − 5. [36] HAIR J F, ANDERSON R E, TATHAM R L, et al. Multi-variate Data Analysis[M]. 5th ed. New Jersey: Prentice Hall, 1998. [37] BURNHAM K P, ANDERSON D R. Model Selection and Multi-model Inference: A Practical Information Theoretic Approach[M]. New York: Springer, 2010. [38] SATDICHANH M, MA H X, YAN K,et al. Phylogenetic diversity correlated with above-ground biomass production during forest succession: evidence from tropical forests in Southeast Asia [J]. J Ecol, 2019, 107(3): 1419 − 1432. [39] 吴初平, 韩文娟, 江波, 等. 浙江定海次生林内物种丰富度与生物量和生产力关系的环境依赖性[J]. 生物多样性, 2018, 26(6): 545 − 553. WU Chuping, HAN Wenjuan, JIANG Bo,et al. Relationships between species richness and biomass/productivity depend on environmental factors in secondary forests of Dinghai, Zhejiang Province [J]. Biodiversity Sci, 2018, 26(6): 545 − 553. [40] OUYANG Shuai, XIANG Wenhua, WANG Xiangping,et al. Significant effects of biodiversity on forest biomass during the succession of subtropical forest in south China [J]. For Ecol Manage, 2016, 372: 291 − 302. [41] VENAIL P, GROSS K, OAKLEY T H,et al. Species richness, but not phylogenetic diversity, influences community biomass production and temporal stability in a re-examination of 16 grassland biodiversity studies [J]. Funct Ecol, 2015, 29(5): 615 − 626. [42] 车盈, 金光泽. 物种多样性和系统发育多样性对阔叶红松林生产力的影响[J]. 应用生态学报, 2019, 30(7): 2241 − 2248. CHE Ying, JIN Guangze. Effects of species diversity and phylogenetic diversity on productivity of a mixed broad-leaved-Korean pine forest [J]. Chin J Appl Ecol, 2019, 30(7): 2241 − 2248. [43] 侯嫚嫚, 李晓宇, 王均伟, 等. 长白山针阔混交林不同演替阶段群落系统发育和功能性状结构简[J]. 生态学报, 2017, 37(22): 7502 − 7513. HOU Manman, LI Xiaoyu, WANG Junwei,et al. Phylogenetic development and functional structures during successional stages of conifer and broad-leaved mixed forest communities in Changbai Mountains, China [J]. Acta Ecol Sin, 2017, 37(22): 7502 − 7513. [44] YUAN Zuoqiang, WANG Shaopeng, GAZOL A,et al. Multiple metrics of diversity have different effects on temperate forest functioning over succession [J]. Oecologia, 2016, 182(4): 1175 − 1185. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200696
 点击查看大图
点击查看大图
计量
- 文章访问数: 3014
- HTML全文浏览量: 447
- PDF下载量: 290
- 被引次数: 0
