




-
中国西南地区的干热河谷气候干热、植被稀疏、土壤贫瘠、水土流失严重,是典型的生态脆弱区,也是植被恢复和生态治理极为困难的区域,主要分布在金沙江、红河、怒江和澜沧江等流域的中上游地区,其中金沙江干热河谷面积最大,也最典型[1]。金沙江干热河谷起于云南省丽江市永胜县与大理州鹤庆县交界地区,向金沙江下游延伸到四川省布拖县与云南省巧家县毗邻的干支流流域河段,海拔一般在1 600 m以下,全长约850 km,面积约2.9万 km2[2]。从20世纪50年代开始,许多学者在干热河谷开展了植被恢复研究与实践,但由于该地区地理环境异质性较高,植被类型和生态恢复方式也不相同,植被恢复效果差异明显[1]。植物多样性研究是植被恢复的前提和基础[3],其影响机制一直是生态学的基本问题和研究热点[4]。地形是影响植物多样性的重要因子[5],可以通过海拔、坡向和坡度等因子的变化以及由它们所决定的光照、水分和养分的空间再分配,引起局部环境变化,从而影响植被[6-8]。目前,国内外学者对于金沙江干热河谷的研究主要集中在植被恢复途径与方法[9]、植物逆境生理机制[10]、现有植被演变过程[11]以及植被恢复对土壤质量的影响[12-13]等,对金沙江干热河谷各区段植物多样性研究较少,尤其是在不同立地环境下[5]。本研究调查了金沙江干热河谷不同区段植被,探究海拔和坡向等立地环境对植物多样性的影响,以期为金沙江干热河谷各区段进行精准生态恢复的物种选择、森林经营和管理提供决策依据。
-
本研究以金沙江干热河谷为研究区域。区域最冷月平均气温>12 ℃,最暖月平均气温为24~28 ℃,年平均积温>7 000 ℃;全年几无霜日;年平均降水量为600~800 mm,年平均蒸发量达2 750~3 850 mm,年平均干燥度>2.0。土壤类型有燥红土、褐红壤、赤红壤、紫色土等。植被群落为稀树灌草丛,以锥连栎Quercus franchetii、滇缆仁Terminalia franchetii、山合欢Albizzia kalkora、车桑子Dodonaea viscose、余甘子Phyllanthus emblica、羊蹄甲Bauhinia purpurea和黄茅Heteropogon contortus等为主。
-
根据前期数据调查和全面实地考察,2021年1月在金沙江干热河谷上段的鹤庆县和永胜县设22个样方,中段的仁和区和元谋县设13个样方,下段的东川区和宁南县设12个样方,共47个样方,其中16个天然林样方,25个人工林样方(林龄20 a左右)和6个稀树灌草丛。各样方面积20 m×20 m,用于乔木调查;沿样方对角线的4个角及中心设置5个5 m×5 m 小样方,用于灌木和草本调查。在样方内取2个环刀样,用于测定土壤含水量。记录样方地理位置、地形地貌和群落特征等。
-
计算植物丰富度指数(R0)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)和Pielou均匀度指数(J)。公式如下[14-15]:R0=S;
$H=-\displaystyle \sum_{i=1}^{S} P_{i} \ln P_{i} $ ;$D=1 - \displaystyle \sum_{i=1}^{S} P_{i}^{2} $ ;J=H/lnS;Pi=Ni/N。其中:S为物种数目,Pi为第i个物种个体数占样方中所有物种个体总数的比例,Ni为种i的个体数目,N为群落中全部物种的个体数。 -
用方差分析(ANOVA,LSD检验)比较金沙江干热河谷各区段、海拔和利用方式间植物多样性指标的差异显著性,用独立样本t检验比较坡向间植物多样性指标的差异显著性,用线性回归分析探究海拔与植物多样性指标的拟合曲线。以上统计分析在SPSS 23.0上完成,图表用Excel 2010整理绘制。
-
金沙江干热河谷群落共调查到植物169种,隶属63科114属。其中金沙江干热河谷上段调查到植物86种,隶属43科73属;中段植物90种,隶属48科76属;下段植物95种,隶属53科80属(表1)。可见,金沙江干热河谷群落植物种类从上段到下段逐渐增加。
表 1 样地基本信息
Table 1. Basic characteristics of plots
区段 样方数 年平均气温/℃ 年均降水/mm 海拔/m 坡向 科 属 种 上段 22 19.4 775 1 200~2 000 阳坡(9),阴坡(13) 43 73 86 中段 13 21.2 800 800~2 000 阳坡(5),阴坡(8) 48 76 90 下段 12 23.5 812 800~2 000 阳坡(5),阴坡(7) 53 80 95 说明:坡向后括号里数字表示样方数。阴坡为0~90°和270°~360°;阳坡:90°~270° 金沙江干热河谷植物群落中调查到菊科Compositae有25种,占植物种类的14.8%;禾本科Gramineae 15种,占植物种类的8.9%;豆科Leguminosae 13种,占植物种类的7.7%;壳斗科Fagaceae 6种,占植物种类的3.6%;蔷薇科Rosaceae 5种,占植物种类的3.0%;桃金娘科Myrtaceae 5种,占植物种类的3.0%;无患子科Sapindaceae 5种,占植物种类的3.0%。就不同区段而言,上段菊科有15种,占植物种类的17.4%;禾本科8种,占植物种类的9.3%;豆科5种,占植物种类的5.8%;壳斗科5种,占植物种类的5.8%;蔷薇科3种,占植物种类的3.5%;柏科Cupressaceae 3种,占植物种类3.5%;莎草科Cyperaceae 3种,占植物种类3.5%。中段菊科17种,占植物种类的18.9%;禾本科10种,占植物种类的11.1%;豆科9种,占植物种类的10.0%;壳斗科4种,占植物种类的4.4%;蔷薇科3种,占植物种类的3.3%;杜鹃花科Ericaceae 3种,占植物种类3.3%。下段菊科18种,占植物种类的18.9%;禾本科13种,占植物种类的13.7%;豆科8种,占植物种类的8.4%;壳斗科4种,占植物种类的4.2%;蔷薇科3种,占植物种类的3.2%;桃金娘科3种,占植物种类3.2%;无患子科3种,占植物种类的3.2%。结果表明:金沙江干热河谷植物群落主要以菊科、禾本科、豆科、壳斗科、蔷薇科、桃金娘科、无患子科、柏科、莎草科、杜鹃花科为主。
-
表2表明:总体上金沙江干热河谷各区段的植物丰富度指数(R0)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)和Pielou均匀度指数(J)变化一致,即从上段到下段均表现为增加趋势。上段群落R0为11.55,分别是中段和下段的68.26%和66.96%。不同区段植物群落H变幅为1.52~2.31,最大值出现于下段(2.31),最小值出现在上段(1.52)。下段植物群落D (0.84)与中段(0.82)接近,但显著高于上段(0.64) (P<0.05)。下段植物J为0.84,与上段差异显著(P<0.05),分别是上段和中段的1.29倍和1.09倍。
对金沙江干热河谷不同利用方式植物多样性分析发现:4种植物多样性指数从大到小依次均为天然林、人工林、稀树灌草丛;天然林的植物丰富度显著高于人工林和稀树灌草丛(P<0.05),但人工林多样性和均匀度与天然林无明显差异;人工林植物丰富度从大到小依次表现为上段、中段、下段,但差异不显著,下段和中段的多样性均高于上段且差异显著(P<0.05);天然林的植物多样性从大到小均表现为上段、中段、下段,且下段和上段差异显著(P<0.05)。
表 2 金沙江干热河谷各区段植物的多样性指数
Table 2. Plant diversity indices in various sections of Jinsha River dry-hot valley
地区 利用方式 R0 H D J 上段 天然林 15.89±5.62 aB 1.80±0.32 aB 0.71±0.11 aB 0.68±0.13 aB 人工林 9.00±3.61 bA 1.47±0.42 abB 0.66±0.08 aB 0.68±0.07 aB 稀树灌草丛 7.50±4.65 bA 1.02±0.61 bA 0.44±0.28 bA 0.52±0.17 bA 合计 11.55±5.84 B 1.52±0.49 B 0.64±0.17 B 0.65±0.13 B 中段 天然林 24.00±5.70 aAB 2.44±0.27 aB 0.87±0.04 aA 0.78±0.06 aAB 人工林 13.67±6.53 bA 2.01±0.46 aA 0.83±0.08 aA 0.82±0.05 aA 稀树灌草丛 9.00±0.00 bA 1.29±0.30 bA 0.64±0.06 bA 0.59±0.14 bA 合计 16.92±8.08 A 2.07±0.53 AB 0.82±0.10 A 0.77±0.10 AB 下段 天然林 33.00±1.14 aA 3.10±0.27 aA 0.94±0.03 aA 0.89±0.07 aA 人工林 14.10±6.06 bA 2.15±0.46 bA 0.83±0.09 aA 0.83±0.08 aA 合计 17.25±9.19 A 2.31±0.56 A 0.84±0.10 A 0.84±0.08 A 金沙江干热河谷 天然林 20.56±7.96 a 2.17±0.55 a 0.79±0.13 a 0.74±0.12 a 人工林 12.16±5.73 b 1.87±0.53 a 0.77±0.12 a 0.77±0.10 a 稀树灌草丛 8.00±3.69 b 1.11±0.51 b 0.51±0.24 b 0.54±0.15 b 总计 14.49±7.78 1.87±0.61 0.74±0.16 0.73±0.14 说明:表中植被的多样性指数均为同类型样地的算术平均数。不同小写字母表示同一区段不同林分间差异显著(P<0.05),不同大写 字母表示同一利用方式不同区段间差异显著(P<0.05) -
在金沙江干热河谷,随海拔升高,R0、H和D均呈上升趋势,而J呈下降趋势(图1)。结合表3可知:R0最大值(23.22)出现在海拔1 800~2 000 m,显著高于其他海拔段(P<0.05),最小值出现在海拔1 200~1 400 m,是最大值的35.32%。H最大值出现在海拔1 000~1 200 m,其次是海拔1 800~2 000 m,其差异不显著,最小值出现在海拔1 200~1 400 m,是最大值的55.51%。除海拔1 200~1 400 m D显著低于其他海拔梯度(P<0.05),其他海拔梯度D差异不显著。在海拔1 200~1 400 m,J最小(0.62),在海拔1 000~1 200 m最大(0.83),其差异显著(P<0.05)。在同一海拔段,植物丰富度和多样性从大到小均表现为下段、中段、上段。
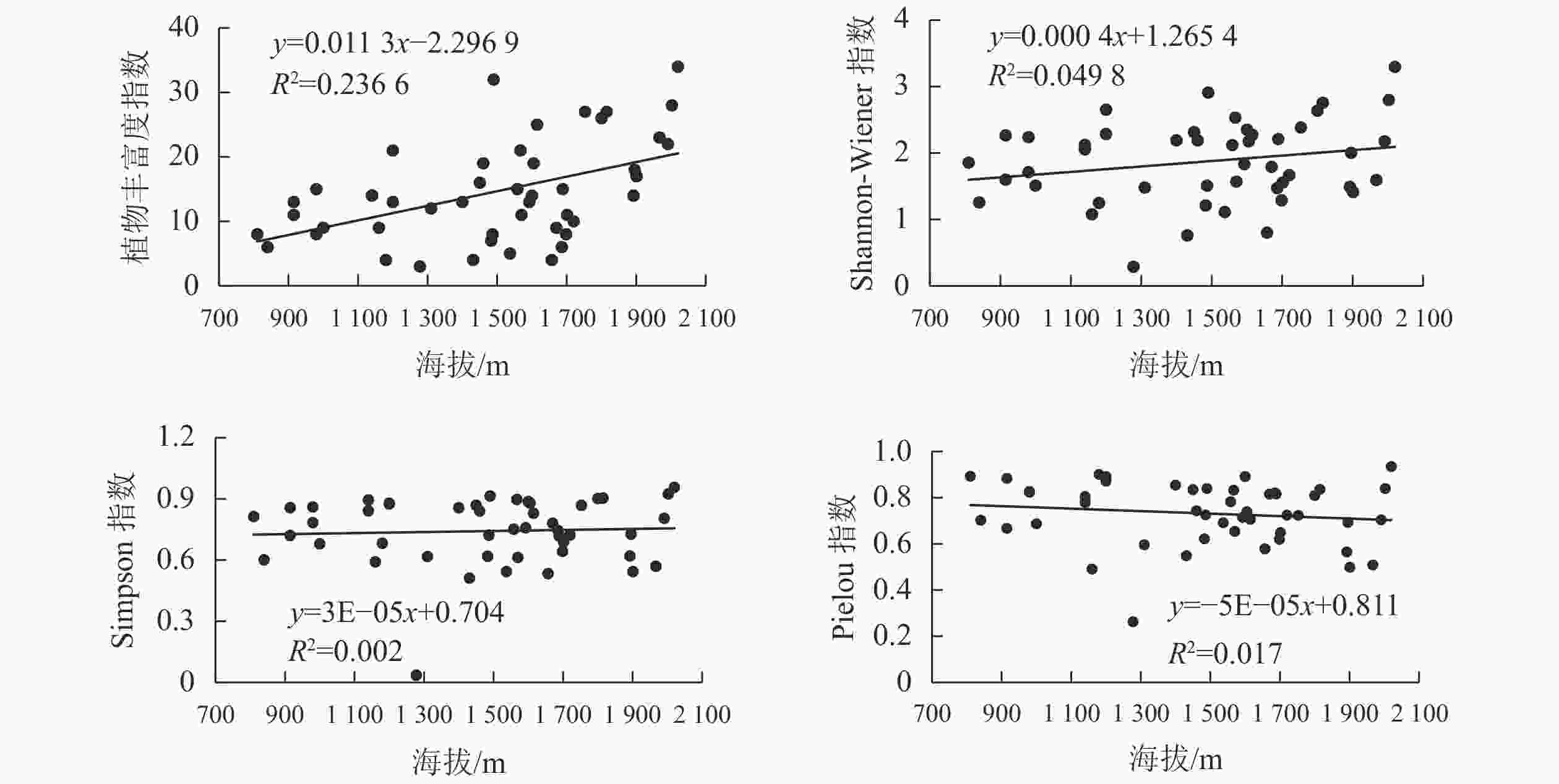
图 1 植物多样性与海拔的关系
Figure 1. Relationship between plant diversity and altitude
表 3 金沙江干热河谷各区段不同海拔段的植物多样性
Table 3. Plant diversity at different altitudes in various sections of the Jinsha River dry-hot valley
海拔/m 地区 R0 H D J 土壤含水量/% 1800~2000 上段 18.80±3.70 bA 1.74±0.34 bA 0.65±0.11 bA 0.59±0.10 bA 0.20±0.02 bA 中段 27.00±1.41 aA 2.72±0.11 aA 0.91±0.02 aA 0.82±0.02 aA 0.21±0.00 abA 下段 30.50±4.95 aA 3.23±0.38 aA 0.93±0.04 aA 0.89±0.07 aA 0.24±0.01 aA 合计 23.22±6.26 A 2.24±0.67 A 0.77±0.16 A 0.71±0.16 AB 0.21±0.02 A 1600~1800 上段 10.25±4.83 bB 1.62±0.46 aAB 0.71±0.10 aA 0.72±0.10 bA 0.15±0.02 bB 中段 26.00±1.41 aA 2.33±0.08 aB 0.85±0.03 aA 0.71±0.01 bA 0.19±0.00 aA 下段 14.00±0.00 bB 2.35±0.00 aBC 0.89±0.00 aAB 0.89±0.00 aA 0.17±0.00 abB 合计 13.45±7.50 B 1.82±0.51 AB 0.75±0.11 A 0.73±0.10 AB 0.16±0.02 B 1400~1600 上段 9.00±4.12 cB 1.44±0.46 bAB 0.65±0.10 bA 0.68±0.08 bA 0.12±0.03 bC 中段 18.67±2.52 bB 2.35±0.17 aB 0.87±0.03 aA 0.80±0.05 aA 0.14±0.01 abB 下段 32.00±0.00 aA 2.91±0.00 aAB 0.91±0.00 aA 0.84±0.00 aA 0.18±0.00 aB 合计 13.73±8.24 B 1.82±0.66 AB 0.73±0.14 A 0.73±0.09 AB 0.13±0.03 C 1200~1400 上段 7.50±6.36 aB 0.88±0.84 aB 0.32±0.41 aB 0.43±0.24 aB 0.09±0.00 cC 中段 6.50±3.54 aC 1.16±0.12 aD 0.64±0.06 aB 0.70±0.29 aA 0.13±0.01 bBC 下段 13.00±0.00 aB 2.19±0.00 aC 0.86±0.00 aAB 0.85±0.00 aA 0.17±0.00 aB 合计 8.20±4.55 B 1.26±0.69 B 0.56±0.31 B 0.62±0.26 B 0.12±0.03 CD 1000~1200 中段 14.00±0.00 aB 2.09±0.05 aB 0.87±0.04 aA 0.79±0.02 aA 0.11±0.02 aCD 下段 16.33±4.16 aB 2.39±0.23 aABC 0.87±0.01 aAB 0.86±0.03 aA 0.14±0.02 aC 合计 15.40±3.21 B 2.27±0.23 A 0.87±0.02 A 0.83±0.05 A 0.13±0.02 C 800~1000 中段 8.50±0.71 aC 1.61±0.14 aC 0.73±0.07 aB 0.76±0.10 aA 0.10±0.01 aD 下段 9.50±3.11 aB 1.75±0.42 aC 0.75±0.11 aB 0.79±0.12 aA 0.10±0.02 aD 合计 9.16±2.48 B 1.70±0.03 AB 0.74±0.09 A 0.78±0.10 AB 0.10±0.02 CD 说明:表中植被的多样性指数均为同类型样地的算术平均数。不同小写字母表示同一海拔不同区段间差异显著(P<0.05),不同大写 字母表示同一指数不同海拔间差异显著(P<0.05) 对比金沙江干热河谷不同坡向植物丰富度、多样性、均匀度发现(表4):阴坡R0、H、D和J均高于阳坡,除J差异不显著外,其他均存在显著差异(P<0.05)。对比各区段相同坡向的植物多样性发现:阳坡植物多样性指数从大到小依次为下段、中段、上段。单因素方差检验显示:下段的植物群落H、D与上段差异显著(P<0.05);阴坡变化趋势与阳坡相同。相同坡向条件下中段和下段的植物群落R0和H与上段存在显著差异(P<0.05),下段的植物群落H和J与上段存在显著差异(P<0.05)。
表 4 金沙江干热河谷各区段不同坡向的植物多样性
Table 4. Plant diversity of different slope downwards in different sections of Jinsha River dry-hot valley
坡向 地区 R0 H D J 土壤含水量/% 阳坡 上段 7.00±4.24 aB 1.15±0.47 bB 0.56±0.22 bB 0.62±0.17 aA 0.13±0.04 B 中段 8.80±3.56 aB 1.53±0.41 abB 0.72±0.88 abB 0.74±0.16 aA 0.11±0.02 B 下段 10.60±3.65 aB 1.85±0.43 aB 0.77±0.11 aB 0.80±0.11 aA 0.12±0.02 B 合计 8.42±4.02 aB 1.44±0.52 abB 0.66±0.19 abB 0.70±0.17 aA 0.12±0.03 B 阴坡 上段 14.69±4.63 bA 1.78±0.32 cA 0.70±0.09 bA 0.67±0.09 cA 0.16±0.03 A 中段 22.00±5.29 aA 2.40±0.24 abA 0.88±0.03 aA 0.78±0.05 abA 0.17±0.03 A 下段 22.00±9.10 aA 2.63±0.40 aA 0.89±0.04 aA 0.87±0.03 aA 0.17±0.05 A 合计 18.61±7.21 abA 2.17±0.49 bcA 0.80±0.11 bA 0.76±0.11 bcA 0.17±0.04 A 说明:表中植被的多样性指数均为同类型样地的算术平均数。不同小写字母表示同一坡向不同区段间差异显著(P<0.05),不同大写 字母表示同一指数不同坡向间差异显著(P<0.05) -
作为群落结构和功能的指示指标,植物多样性可以有效反映群落的组成、结构和演变以及环境现状,对珍稀濒危物种的保护具有至关重要的作用[16]。森林群落植物多样性最丰富,也是生态恢复保护的重要领域之一,研究群落植物多样性有助于维持生态系统的动态稳定[17]。研究表明[18]:提高人工林物种多样性可增加森林生态系统的稳定性。不过,人工纯林可能会导致生产力下降,不利于可持续经营[19]。本研究在金沙江干热河谷共调查记录了植物169种,隶属63科114属,以菊科、禾本科、豆科、壳斗科、蔷薇科、桃金娘科、无患子科、柏科、莎草科、杜鹃花科为主。植物多样性指数从上段到下段均有上升的趋势,这与前人的结论类似。金沙江干热河谷热量充沛,水分条件是主要的限制因子。各区段的年降水量从大到小表现为下段、中段、上段,植物多样性指数也表现为同样的趋势。张建利等[20]发现:金沙江干热河谷草地植物多样性和均匀性指数自上游至下游逐渐上升,认为与年均温和年降水量有关。本研究发现:不同利用方式的植物多样性指数从大到小依次为天然林、人工林、稀树灌草丛,天然林由于种群与环境之间的长期适应,群落内部种间关系协调,普遍表现出较高的植物多样性,然而除天然林和人工林植物丰富度指数有显著差异外,其他多样性指数和均匀度指数均无明显差异。经过大约20 a的发展,植被高度、凋落物量的增加,径级结构的改变,人工林的种间和种内竞争虽然激烈,很难容纳新的物种产生,但人工林植被结构较稀树灌草丛有显著改善,群落结构趋于稳定,有向天然林发展的趋势。人工林植物多样性提高,说明植树造林工程取得了一定的成效,干热河谷局部造林是可行和成功的。
-
海拔决定了区域的水、热条件,是生境影响植物多样性的主导因子[21]。植物多样性的海拔梯度格局,可反映植物生态学特性及对环境的适应性[22]。植物多样性沿海拔梯度的分布格局一般有5种形式,分别是沿海拔梯度先降后升、单峰曲线、单调升高、单调下降和无明显格局[23]。本研究发现:随着海拔上升,金沙江干热河谷植物丰富度和多样性指数均呈现逐渐上升趋势;这种格局通常出现在极端环境下,主要是由较小的海拔梯度引起的[24]。植物群落受坡位、坡向、坡度等小生境的影响,多样性指数会出现波动,但总体来说,由于海拔上升,干热河谷区域土壤含水量提高(表3),植物多样性也相应提高。原因是随着土壤含水量增加,植物群落种间竞争减少,降低了乔木和灌木对草本的影响,草本生存空间增加,植物多样性增加。此外1 600 m以上海拔段,其气候、土壤、植被类型与1 600 m以下不同。在1 600 m以上的林地主要为天然林,在1 800~2 000 m海拔范围内植物丰富度指数和Shannon-Wiener多样性指数分别达23.22和2.24,说明该区域植被结构稳定,物种适应了样地的局地环境条件,有利于木本幼苗、蚂蚁、爬行类和两栖类动物的迁入,增加林地植物多样性。
-
地形因子是影响植物丰富度及植物多样性的重要因素[25]。坡向通过改变太阳辐射和水分分布等生境条件影响群落物种的种类和数量[26],是重要地形因子之一。有研究表明[27]:阴坡土壤水分含量较高,林内阳光分布均匀,有利于灌木层物种良好发育,这是阴坡灌木层植物多样性指数显著高于阳坡的原因之一;阳坡的立地条件较为恶劣,严重风化使得土壤养分难以富集,同时较强的光照强度和较高的土壤水分蒸发速率,抑制了耐阴植物的正常生长,导致阳坡群落总体植物多样性降低。本研究发现:坡向对植物丰富度、Shannon-Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数都有显著影响,均表现为阴坡高于阳坡,与前人的研究结果一致。一方面是因为干热河谷月平均气温均高于5 ℃,植物均可生长,热量对植被恢复的限制较小,阳坡受光照直射时间长,获得太阳辐射多,土壤温度偏高,蒸发量大,在降水量相同的情况下,土壤含水量较阴坡低(表4),有利于物种的迁入和多样性的保持;另一方面,干热河谷植物群落多数为以禾草草丛为背景构成大片草地植被,在此基础上散生稀疏乔木和灌木的稀树灌草丛;群落结构多数分乔、灌、草3层,或灌、草2层,草本层为群落优势层;群落植物种类组成多数为热带起源耐干旱的种类,阳坡植被结构简单,多以灌草为主,除优势种外,其他各个种群的重要值都很小,群落的自我调节机制弱,群落容易在干热河谷恶劣的环境影响下发生改变,群落内部各种群的稳定性较差;阴坡植被结构复杂,以乔灌草结构为主,群落的自我调节机制强,群落内部各种群稳定,故植物多样性高。
Plant diversity in various sections of Jinsha River dry-hot valley under different site environments
-
摘要:
目的 为更精准地保护和修复金沙江干热河谷生态环境,明晰不同立地环境下金沙江干热河谷各区段植物多样性结构特征与差异。 方法 采用标准样地调查法调查金沙江干热河谷上段、中段和下段共47个样地的植物物种丰富度和多样性等。 结果 ① 金沙江干热河谷植物丰富度、多样性和均匀度从上段到下段有增加趋势,植物丰富度从大到小依次为天然林(20.56)、人工林(12.16)、稀树灌草丛(8.00),天然林和人工林之间多样性和均匀度差异均不显著。② 随海拔上升,植物多样性有增加趋势。阴坡植物多样性显著高于阳坡(P<0.05)。③ 在海拔800~1 400 m,上段、中段和下段多样性差异不显著,而在海拔1 400~2 000 m,多样性从大到小依次为下段、中段、上段;同一坡向下段的植物多样性均高于中段和上段,且下段的Shannon-Wiener多样性指数和Simpson多样性指数显著高于上段(P<0.05)。 结论 受海拔和坡向等立地环境的影响,金沙江干热河谷各区段植物多样性存在明显差异,局部造林可提高当地的植物多样性。图1表4参27 Abstract:Objective This study is aimed at an investigation of the structural features and differences of the plant diversity in various sections of the dry-hot valley under different site environments so as to efficiently protect and restore the biological environment of Jinsha River dry-hot valley. Method With the employment of standard sample plot survey method, a survey was conducted of the plant species richness and diversity of 47 plots in the upper, middle and lower sections of the Jinsha River dry-hot valley. Result The plant species richness, diversity and uniformity of the Jinsha River dry-hot valley increased from the upper section to the lower one with the order of species richness being natural forest (20.56)>plantation (12.16)>sparse trees and shrubs (8.00) and no significant difference in diversity or uniformity between natural forest and plantation. The plant diversity tends to increase as the altitude increases with the plant diversity of shade slopes being significantly higher than that of sunny slopes (P<0.05). No significant differences have been found in diversity among the upper, middle and lower sections in the range of 800−1 400 m, while in the range of 1 400−2 000 m, the diversity in the middle section was lower than that in the lower section, but higher than the upper section. The plant diversity in the lower section was always higher than that in the middle section and the upper section of the same slope, and the Shannon-Wiener diversity index and Simpson diversity index in the lower section were significantly different from those of the upper section (P< 0.05). Conclusion There are significant differences in plant diversity in various sections of the Jinsha River dry-hot valley, with altitude and slope as main environmental influencing factors, and local plantation may increase the plant diversity. [Ch, 1 fig. 4 tab. 27 ref.] -
Key words:
- dry-hot valley /
- watershed section /
- plant diversity /
- site environment
-
表 1 样地基本信息
Table 1. Basic characteristics of plots
区段 样方数 年平均气温/℃ 年均降水/mm 海拔/m 坡向 科 属 种 上段 22 19.4 775 1 200~2 000 阳坡(9),阴坡(13) 43 73 86 中段 13 21.2 800 800~2 000 阳坡(5),阴坡(8) 48 76 90 下段 12 23.5 812 800~2 000 阳坡(5),阴坡(7) 53 80 95 说明:坡向后括号里数字表示样方数。阴坡为0~90°和270°~360°;阳坡:90°~270°  下载: 导出CSV
下载: 导出CSV
表 2 金沙江干热河谷各区段植物的多样性指数
Table 2. Plant diversity indices in various sections of Jinsha River dry-hot valley
地区 利用方式 R0 H D J 上段 天然林 15.89±5.62 aB 1.80±0.32 aB 0.71±0.11 aB 0.68±0.13 aB 人工林 9.00±3.61 bA 1.47±0.42 abB 0.66±0.08 aB 0.68±0.07 aB 稀树灌草丛 7.50±4.65 bA 1.02±0.61 bA 0.44±0.28 bA 0.52±0.17 bA 合计 11.55±5.84 B 1.52±0.49 B 0.64±0.17 B 0.65±0.13 B 中段 天然林 24.00±5.70 aAB 2.44±0.27 aB 0.87±0.04 aA 0.78±0.06 aAB 人工林 13.67±6.53 bA 2.01±0.46 aA 0.83±0.08 aA 0.82±0.05 aA 稀树灌草丛 9.00±0.00 bA 1.29±0.30 bA 0.64±0.06 bA 0.59±0.14 bA 合计 16.92±8.08 A 2.07±0.53 AB 0.82±0.10 A 0.77±0.10 AB 下段 天然林 33.00±1.14 aA 3.10±0.27 aA 0.94±0.03 aA 0.89±0.07 aA 人工林 14.10±6.06 bA 2.15±0.46 bA 0.83±0.09 aA 0.83±0.08 aA 合计 17.25±9.19 A 2.31±0.56 A 0.84±0.10 A 0.84±0.08 A 金沙江干热河谷 天然林 20.56±7.96 a 2.17±0.55 a 0.79±0.13 a 0.74±0.12 a 人工林 12.16±5.73 b 1.87±0.53 a 0.77±0.12 a 0.77±0.10 a 稀树灌草丛 8.00±3.69 b 1.11±0.51 b 0.51±0.24 b 0.54±0.15 b 总计 14.49±7.78 1.87±0.61 0.74±0.16 0.73±0.14 说明:表中植被的多样性指数均为同类型样地的算术平均数。不同小写字母表示同一区段不同林分间差异显著(P<0.05),不同大写 字母表示同一利用方式不同区段间差异显著(P<0.05)
下载: 导出CSV
表 3 金沙江干热河谷各区段不同海拔段的植物多样性
Table 3. Plant diversity at different altitudes in various sections of the Jinsha River dry-hot valley
海拔/m 地区 R0 H D J 土壤含水量/% 1800~2000 上段 18.80±3.70 bA 1.74±0.34 bA 0.65±0.11 bA 0.59±0.10 bA 0.20±0.02 bA 中段 27.00±1.41 aA 2.72±0.11 aA 0.91±0.02 aA 0.82±0.02 aA 0.21±0.00 abA 下段 30.50±4.95 aA 3.23±0.38 aA 0.93±0.04 aA 0.89±0.07 aA 0.24±0.01 aA 合计 23.22±6.26 A 2.24±0.67 A 0.77±0.16 A 0.71±0.16 AB 0.21±0.02 A 1600~1800 上段 10.25±4.83 bB 1.62±0.46 aAB 0.71±0.10 aA 0.72±0.10 bA 0.15±0.02 bB 中段 26.00±1.41 aA 2.33±0.08 aB 0.85±0.03 aA 0.71±0.01 bA 0.19±0.00 aA 下段 14.00±0.00 bB 2.35±0.00 aBC 0.89±0.00 aAB 0.89±0.00 aA 0.17±0.00 abB 合计 13.45±7.50 B 1.82±0.51 AB 0.75±0.11 A 0.73±0.10 AB 0.16±0.02 B 1400~1600 上段 9.00±4.12 cB 1.44±0.46 bAB 0.65±0.10 bA 0.68±0.08 bA 0.12±0.03 bC 中段 18.67±2.52 bB 2.35±0.17 aB 0.87±0.03 aA 0.80±0.05 aA 0.14±0.01 abB 下段 32.00±0.00 aA 2.91±0.00 aAB 0.91±0.00 aA 0.84±0.00 aA 0.18±0.00 aB 合计 13.73±8.24 B 1.82±0.66 AB 0.73±0.14 A 0.73±0.09 AB 0.13±0.03 C 1200~1400 上段 7.50±6.36 aB 0.88±0.84 aB 0.32±0.41 aB 0.43±0.24 aB 0.09±0.00 cC 中段 6.50±3.54 aC 1.16±0.12 aD 0.64±0.06 aB 0.70±0.29 aA 0.13±0.01 bBC 下段 13.00±0.00 aB 2.19±0.00 aC 0.86±0.00 aAB 0.85±0.00 aA 0.17±0.00 aB 合计 8.20±4.55 B 1.26±0.69 B 0.56±0.31 B 0.62±0.26 B 0.12±0.03 CD 1000~1200 中段 14.00±0.00 aB 2.09±0.05 aB 0.87±0.04 aA 0.79±0.02 aA 0.11±0.02 aCD 下段 16.33±4.16 aB 2.39±0.23 aABC 0.87±0.01 aAB 0.86±0.03 aA 0.14±0.02 aC 合计 15.40±3.21 B 2.27±0.23 A 0.87±0.02 A 0.83±0.05 A 0.13±0.02 C 800~1000 中段 8.50±0.71 aC 1.61±0.14 aC 0.73±0.07 aB 0.76±0.10 aA 0.10±0.01 aD 下段 9.50±3.11 aB 1.75±0.42 aC 0.75±0.11 aB 0.79±0.12 aA 0.10±0.02 aD 合计 9.16±2.48 B 1.70±0.03 AB 0.74±0.09 A 0.78±0.10 AB 0.10±0.02 CD 说明:表中植被的多样性指数均为同类型样地的算术平均数。不同小写字母表示同一海拔不同区段间差异显著(P<0.05),不同大写 字母表示同一指数不同海拔间差异显著(P<0.05)
下载: 导出CSV
表 4 金沙江干热河谷各区段不同坡向的植物多样性
Table 4. Plant diversity of different slope downwards in different sections of Jinsha River dry-hot valley
坡向 地区 R0 H D J 土壤含水量/% 阳坡 上段 7.00±4.24 aB 1.15±0.47 bB 0.56±0.22 bB 0.62±0.17 aA 0.13±0.04 B 中段 8.80±3.56 aB 1.53±0.41 abB 0.72±0.88 abB 0.74±0.16 aA 0.11±0.02 B 下段 10.60±3.65 aB 1.85±0.43 aB 0.77±0.11 aB 0.80±0.11 aA 0.12±0.02 B 合计 8.42±4.02 aB 1.44±0.52 abB 0.66±0.19 abB 0.70±0.17 aA 0.12±0.03 B 阴坡 上段 14.69±4.63 bA 1.78±0.32 cA 0.70±0.09 bA 0.67±0.09 cA 0.16±0.03 A 中段 22.00±5.29 aA 2.40±0.24 abA 0.88±0.03 aA 0.78±0.05 abA 0.17±0.03 A 下段 22.00±9.10 aA 2.63±0.40 aA 0.89±0.04 aA 0.87±0.03 aA 0.17±0.05 A 合计 18.61±7.21 abA 2.17±0.49 bcA 0.80±0.11 bA 0.76±0.11 bcA 0.17±0.04 A 说明:表中植被的多样性指数均为同类型样地的算术平均数。不同小写字母表示同一坡向不同区段间差异显著(P<0.05),不同大写 字母表示同一指数不同坡向间差异显著(P<0.05)
下载: 导出CSV
-
[1] 李昆, 孙永玉. 干热河谷植被恢复技术[M]. 昆明: 云南科技出版社, 2011. LI Kun, SUN Yongyu. Vegetation Rehabilitation in the Arid-Hot Valleys [M]. Kunming: Yunnan Science and Technology Press, 2011. [2] 张荣祖. 横断山区干旱河谷[M]. 北京: 科学出版社, 1992. ZHANG Rongzu. The Dry Valleys of the Hengduan Mountains Region [M]. Beijing: Science Press, 1992. [3] HAMMOND R A, HUDSON M D. Environmental management of UK golf courses for biodiversity: attitudes and actions [J]. Landscape Urban Plann, 2007, 83(2/3): 127 − 136. [4] GRAHAM C H, FINE P V A. Phylogenetic beta diversity: linking ecological and evolutionary processes across space in time [J]. Ecol Lett, 2010, 11(12): 1265 − 1277. [5] 袁铁象, 张合平, 欧芷阳, 等. 地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响[J]. 应用生态学报, 2014, 25(10): 2803 − 2810. YUAN Tiexiang, ZHANG Heping, OU Zhiyang, et al. Effects of topography on the diversity and distribution pattern of ground plants in karst montane forests in Southwest Guangxi, China [J]. Chin J Appl Ecol, 2014, 25(10): 2803 − 2810. [6] 刘浩栋, 陈巧, 徐志扬, 等. 海南岛霸王岭陆均松天然群落物种多样性及地形因子的解释[J]. 生态学杂志, 2020, 39(2): 394 − 403. LIU Haodong, CHEN Qiao, XU Zhiyang, et al. Effects of topographical factors on speciesdiversity across Dacrydium pectinatum natural community in Hainan Island [J]. Chin J Ecol, 2020, 39(2): 394 − 403. [7] 温佩颖, 金光泽. 地形对阔叶红松林物种多样性的影响[J]. 生态学报, 2019, 39(3): 945 − 956. WEN Peiying, JIN Guangze. Effects of topography on species diversity in a typical mixed broadleaved-Korean pine forest [J]. Acta Ecol Sin, 2019, 39(3): 945 − 956. [8] 秦随涛, 龙翠玲, 吴邦利. 地形部位对贵州茂兰喀斯特森林群落结构及物种多样性的影响[J]. 北京林业大学学报, 2018, 40(7): 18 − 26. QIN Suitao, LONG Cuiling, WU Bangli. Effects of topographic sites on the community structure and species diversity of karst forest in Maolan, Guizhou Province of southwestern China [J]. J Beijing For Univ, 2018, 40(7): 18 − 26. [9] 纪中华, 方海东, 杨艳鲜, 等. 金沙江干热河谷退化生态系统植被恢复生态功能评价——以元谋小流域典型模式为例[J]. 生态环境学报, 2009, 18(4): 185 − 191. JI Zhonghua, FANG Donghai, YANG Yanxian, et al. Assessment of system functions after vegetation restoration of the degraded on of the degraded ecosystem ecosystem in arid-hot valleys of Jinsha River: a case study on small watershed of Yuanmou [J]. Ecol Environ Sci, 2009, 18(4): 185 − 191. [10] 朱俊杰, 曹坤芳. 元江干热河谷毛枝青冈和三叶漆抗氧化系统季节变化[J]. 植物生态学报, 2008, 32(5): 985 − 993. ZHU Junjie, CAO Kunfang. Seasonal changes in the foliar antioxidant systems in Cyclobalanopsis helferiana and Terminthia paniculata in the hot-dry valley of the Yuanjiang River, China [J]. Chin J Plant Ecol, 2008, 32(5): 985 − 993. [11] 刘耕武, 李代芸, 黄翡, 等. 云南元谋盆地上新世甘棠组植物和孢粉组合及其古气候意义[J]. 古生物学报, 2002, 41(1): 1 − 9. LIU Gengwu, LI Daiyun, HUANG Fei, et al. A pliocene flora from the gantang formation of Yuanmou Basin, Yunnan Province, SW China and ITS paleoclimate significance [J]. Acta Palaeontol Sin, 2002, 41(1): 1 − 9. [12] TANG Guoyong, LI Kun. Soil amelioration through afforestation and self-repair in a degraded valley-type savanna [J]. For Ecol Manage, 2014, 320: 13 − 20. [13] 唐国勇, 李昆, 孙永玉, 等. 干热河谷不同利用方式下土壤活性有机碳含量及其分配特征[J]. 环境科学, 2010, 31(5): 1365 − 1371. TANG Guoyong, LI Kun, SUN Yongyu, et al. Soil labile organic carbon contents and their allocation characteristics under different land uses at dry-hot valley [J]. Environ Sci, 2010, 31(5): 1365 − 1371. [14] 方精云, 沈泽昊, 唐志尧, 等. “中国山地植物物种多样性调查计划”及若干技术规范[J]. 生物多样性, 2004, 12(1): 5 − 9. FANG Jingyun, SHEN Zehao, TANG Zhiyao, et al. The protocol for the survey plan for plant species diversity of China’s mountains [J]. Biodiversity Sci, 2004, 12(1): 5 − 9. [15] RAN J C, SINGH A, SHARMA Y, et al. Dynamics of plant bioresources in Western Himalayan region of India-watershed based study [J]. Current Sci, 2010, 98(2): 192 − 203. [16] 贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征[J]. 生态学报, 1997, 17(1): 91 − 99. HE Jinsheng, CHEN Weilie. A review of gradient changes in species diversity of land plant communities [J]. Acta Ecol Sin, 1997, 17(1): 91 − 99. [17] 徐远杰, 陈亚宁, 李卫红, 等. 伊犁河谷山地植物群落物种多样性分布格局及环境解释[J]. 植物生态学报, 2010, 34(10): 22 − 34. XU Yuanjie, CHEN Yaning, LI Weihong, et al. Distribution pattern and environmental interpretation of plant species diversity in the mountainous region of Ili River Valley, Xinjiang, China [J]. Chin J Plant Ecol, 2010, 34(10): 22 − 34. [18] 盛炜彤. 人工林的生物学稳定性与可持续经营[J]. 世界林业研究, 2001, 14(6): 14 − 21. SHENG Weitong. The relationship between biological stability and sustainable management of plantation [J]. World For Res, 2001, 14(6): 14 − 21. [19] 孙长忠, 沈国舫. 对我国人工林生产力评价与提高问题的几点认识[J]. 世界林业研究, 2001, 14(1): 76 − 80. SUN Changzhong, SHEN Guofang. On the productivity of China’s plantation [J]. World For Res, 2001, 14(1): 76 − 80. [20] 张建利, 柳小康, 沈蕊, 等. 金沙江流域干热河谷草地群落物种数量及多样性特征[J]. 生态环境学报, 2010, 19(7): 1519 − 1524. ZHANG Jianli, LIU Xiaokang, SHEN Rui, et al. The species quantitative and diversity characteristic of grassland community in hot-dry valley of Jinsha River [J]. Ecol Environ Sci, 2010, 19(7): 1519 − 1524. [21] 沈泽昊, 刘增力, 伍杰. 贡嘎山东坡植物区系的垂直分布格局[J]. 生物多样性, 2004, 12(1): 89 − 98. SHEN Zehao, LIU Zengli, WU Jie. Altitudinal pattern of flora on the eastern slope of Mt. Gongga [J]. Biodiversity Sci, 2004, 12(1): 89 − 98. [22] 蒋艾平, 姜景民, 刘军. 檫木叶片性状沿海拔梯度的响应特征[J]. 生态学杂志, 2016, 35(6): 1467 − 1474. JIANG Aiping, JIANG Jingmin, LIU Jun. Responses of leaf traits of Sassafras tsumu (Hemsl.) Hemsl. along an altitudinal gradient [J]. Chin J Ecol, 2016, 35(6): 1467 − 1474. [23] 唐志尧, 方精云. 植物物种多样性的垂直分布格局[J]. 生物多样性, 2004, 12(1): 20 − 28. TANG Zhiyao, FANG Jingyun. A review on the elevational patterns of plant species diversity [J]. Biodiversity Sci, 2004, 12(1): 20 − 28. [24] BARUCH Z. Ordination and classification of vegetation along an altitudinal gradient in the Venezuelan páramos [J]. Vegetatio, 1984, 55: 115 − 126. [25] 周萍, 刘国彬, 侯喜禄. 黄土丘陵区不同坡向及坡位草本群落生物量及多样性研究[J]. 中国水土保持科学, 2009, 7(1): 67 − 73. ZHOU Ping, LIU Guobin, HOU Xilu. Biomass and species diversity of herbosa at different position and aspects of slope in the Hilly-gully Region of Loess Plateau [J]. Sci Soil Water Conserv, 2009, 7(1): 67 − 73. [26] AUSLANDER M, NEVO E, INBAR M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores [J]. J Arid Environ, 2003, 55(3): 405 − 416. [27] 刘旻霞, 南笑宁, 张国娟, 等. 高寒草甸不同坡向植物群落物种多样性与功能多样性的关系[J]. 生态学报, 2021, 41(13): 5398 − 5407. LIU Minxia, NAN Xiaoning, ZHANG Guojuan, et al. Relationship between species diversity and functional diversity of plant communities on different slopes in alpine meadow [J]. Acta Ecol Sin, 2021, 41(13): 5398 − 5407. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210572
 点击查看大图
点击查看大图

计量
- 文章访问数: 2643
- HTML全文浏览量: 459
- PDF下载量: 76
- 被引次数: 0


