





-
目前,由于化石燃料的逐渐枯竭及其利用过程中造成的环境污染问题,亟需寻求清洁可再生的能源利用方式。利用可再生的生物质资源,通过热化学转化法制备燃料和化学品,对中国实现碳达峰、碳中和目标具有重大意义[1]。玉米Zea mays秸秆、棕榈Trachycarpus fortunei壳与马尾松Pinus massoniana分别属于草本生物质、果壳类生物质和木本生物质,资源储量丰富。然而针对此类农林废弃物,传统的处理手段存在产物单一、附加值低、效率低等问题,因此开发新型的生物质资源利用方法,提高生物质资源利用率,增加其经济附加值,是目前生物质综合利用研究领域面临的重要问题[2]。
生物质气化多联产技术是以空气、氧气、水蒸气等为气化剂,通过热化学反应将生物质中的纤维素、半纤维素和木质素组分转化为可燃气体、生物质炭和焦油的过程[3]。可燃气体具有较高的热值,广泛应用于集中供气、发电、供暖等领域;通过物理和化学活化法,生物质炭可制成各种类型的活性炭,广泛应用于化工、环保及军工等领域;通过分馏和复配技术,生物质焦油可用于制成液体肥料、杀菌剂及防腐剂等[4−5]。成亮等[6]以糠醛渣和废菌棒为原料,王亮才等[7]以紫茎泽兰Eupatorium adenophorum秆为原料,分别采用生物质气化多联产技术制备了生物质炭与可燃气,生物质炭用于制成机制炭和炭基肥,而可燃气则用于锅炉燃烧供热。然而,将典型的3种不同类型的生物质(玉米秸秆、棕榈壳、马尾松)进行生物质气化多联产对比研究还未见相关报道。
本研究选取玉米秸秆、棕榈壳、马尾松等3种生物质为原料,以空气为气化剂,采用自主设计的微型气化反应器,开展3种生物质的气化多联产研究,系统研究了气化温度对气化性能的影响,并且分别对气化三相产物(可燃气、生物质炭与焦油)的特性进行了分析。
-
选取3种代表性的生物质原料进行气化实验,分别为木本生物质(马尾松)、草本生物质(玉米秸秆)和果壳类生物质(棕榈壳),其中玉米秸秆取自山西省忻州市西张村,棕榈壳取自南京菲尔莫材料科技有限公司,马尾松取自浙江省杭州市临安区木材加工厂。生物质原料经自然晾晒后,用粉碎机磨成粉末,筛选出粒径100~200目的生物质粉末,在105 ℃烘箱中烘至绝干,装袋保存。
-
参照国标GB/T 28731—2012《固体生物质燃料工业分析方法》,对3种生物质原料及其气化炭产物(挥发分、固定碳和灰分)进行测定;采用元素分析仪(Vario EL Ⅲ)对3种生物质原料及其气化炭产物的元素进行测定,其中氧(O)元素通过差减法计算得到;采用微机量热仪(ZDHW-8A)对原料及其气化炭产物的热值进行测量。采用X射线衍射仪(XRF-6000)对原料的结晶度进行测量,每次取样品粉末0.2 mg,经压片后送入样品室测试,扫描范围为5°~40°,扫描速度为2°·min−1,纤维素的结晶度按照式(1)进行计算。
$$ {I}_{\mathrm{C}\mathrm{r}}=\frac{{I}_{002}-{I}_{\mathrm{a}\mathrm{m}}}{{I}_{002}}\times 100\mathrm{\%} 。 $$ (1) 式(1)中:ICr为纤维素相对结晶度指数,I002是(002)面衍射角的极大强度,即结晶区的衍射强度,Iam是2θ为18°附近的非晶区衍射强度。采用热重分析仪(TG209 F1)对3种原料的热失重规律进行分析,每次实验取样品5 mg,置于石英坩埚中,氮气(N2)流量为40 mL·min−1,以20 ℃·min−1升温速率从室温升至800 ℃。
-
如图1所示,采用自主研发的微型气化反应装置开展玉米秸秆、棕榈壳与马尾松的气化实验。实验装置可分为供气系统、反应系统、焦油收集系统、气体收集系统4个部分。供气系统主要由高纯氮气瓶、高纯氧气瓶、气体流量控制器组成;反应系统主要由温度控制器和加热反应器组成,用于得到气化固体产物;焦油收集系统主要由低温浴槽和冷凝管组成,用于收集冷凝气体,得到气化液体产物;气体收集系统主要由干燥剂和集气袋组成,用于收集不可冷凝可燃气,得到气化气体产物。

图 1 微型气化反应器装置简图
Figure 1. Schematic diagram of miniature gasification reactor
气化实验开始前,将5 g样品装入填有石英棉的石英坩埚中并悬挂于石英管的冷却区,然后通入预混的气化剂[N2与氧气(O2)混合而成的气体],保持10 min,将反应器和链接管路内的其他气体排尽。当量比选取0.20,单位质量原料消耗的空气气化剂体积(Vair)的计算公式如下:
$$ {V}_{\mathrm{a}\mathrm{i}\mathrm{r}}=\lambda \frac{22.41}{0.21}\left(\frac{{W}_{\mathrm{C}}}{12}+\frac{{W}_{\mathrm{H}}}{4}-\frac{{W}_{\mathrm{O}}}{32}\right) 。 $$ (2) 式(2)中:λ为当量比;WC、WH、WO分别为玉米秸秆中碳(C)、氢(H)、O元素质量分数(%)。根据N2和O2的配比分别调节两者的进气量,玉米秸秆气化实验通入N2与O2的流量分别为42.2与158.9 mL·min−1,棕榈壳气化实验通入N2与O2的流量分别为45.1与169.6 mL·min−1,马尾松气化实验分别通入N2与O2的流量为46.0与173.0 mL·min−1。之后,采用20 ℃·min−1的升温速率,将反应器加热至不同的气化温度(700、800和900 ℃),待气化反应器加热至预设的气化温度后,迅速通过炉钩将坩埚垂直推至中部的反应区进行气化实验,同时连接好气袋,准备收集气化反应气体。气化产生的挥发分随气化剂进入冷凝管。冷凝管置于0 ℃以下的冰水混合物环境中,液体产物收集在冷凝管中,挥发分中的不可冷凝气体由集气袋收集。气化反应结束后,停止升温,立即用进样棒将石英坩埚拉至石英管的冷却区,关闭O2,继续通入N2,样品在冷却区逐渐降温。待坩埚内样品冷却后,取出得到气化炭,集气袋中收集到气化燃气,冷凝管中收集到气化液体产物。气化固体和液体产物的产率通过式(3)和(4)计算,气体产物的产率通过差减法获得。
$$ {Y}_{{\rm{solid}}}=\frac{{W}_{{\rm{solid}}}}{{W}_{{\rm{D}}}}\times 100\% \text{;} $$ (3) $$ {Y}_{{\rm{liquid}}}=\frac{{W}_{{\rm{liquid}}}}{{W}_{{\rm{D}}}}\times 100\% 。 $$ (4) 式(3)~(4)中:Ysolid、Yliquid分别为固体产物和液体产物的质量产率(%);Wsolid、Wliquid分别为气化炭和焦油的质量(mg);WD为原料的质量(mg)。
-
采用气相色谱仪(GC9890B)对可燃气的组分进行分析,包括一氧化碳(CO)、氢气(H2)、甲烷(CH4)、二氧化碳(CO2)、O2和N2,可燃气的低位热值通过式(5)进行计算。
$$ {V}_{\mathrm{L}\mathrm{H}}=\mathrm{ }12.6{V}_{\mathrm{C}\mathrm{O}}+10.8{V}_{{\mathrm{H}}_{2}}+35.9{V}_{{\mathrm{C}\mathrm{H}}_{4}} 。$$ (5) 式(5)中:VLH为可燃气的低位热值(MJ·m−3),$ {V}_{\mathrm{C}\mathrm{O}} $、$ {V}_{{\mathrm{H}}_{2}} $、$ {V}_{{\mathrm{C}\mathrm{H}}_{4}} $分别代表各气体成分在可燃气中的体积分数(%)。采用气质联用仪(7890B-5977B)对气化焦油组分进行分析,测试前先采用色谱级甲醇溶剂对液体产物进行稀释。气相色谱检测条件:载气为高纯氦气(99.999%),载气流速1 mL·min−1,色谱柱为HP-5MS毛细管柱,不分流。柱箱升温过程以4 ℃·min−1的升温速率升至260 ℃。质谱检测条件:离子源温度230 ℃,四级杆温度150 ℃,全扫描模式,溶剂延迟时间3 min。根据美国国家标准技术研究所谱库对液体产物进行组分鉴定和相对含量分析,用峰面积归一化法计算各组分的相对含量。
-
表1列出了3种不同类型生物质原料的元素分析、工业分析和热值结果。由元素分析结果可知:生物质原料的氮(N)和硫(S)元素都很低,其中棕榈壳的C元素质量分数最高(48.50%),马尾松的H元素质量分数最高(6.17%),玉米秸秆C元素最低,而O元素质量分数最高(47.75%),导致玉米秸秆的高位热值(16.16 MJ·kg−1)小于棕榈壳(17.06 MJ·kg−1)和马尾松(17.77 MJ·kg−1)。由工业分析结果可知:马尾松的挥发分质量分数最高,而灰分质量分数极低,仅为0.20%,远低于玉米秸秆(3.38%)和棕榈壳(4.51%),灰分中的碱和碱土金属在气化过程中将促进焦油的二次裂解,降低焦油质量分数。
表 1 玉米秸秆、棕榈壳和马尾松的元素分析、工业分析和热值
Table 1. Ultimate analysis, proximate analysis, and calorific values of corn stalk, palm kernel shell, and pine wood
样品 工业分析 元素分析 高位热值/(MJ·kg−1) m挥发分/% m灰分/% m固定碳/% mC/% mH/% mO/% mN/% mS/% 玉米秸秆 77.79 3.38 18.83 45.75 5.80 47.75 0.49 0.21 16.16 棕榈壳 73.04 4.51 22.45 48.50 5.53 44.80 1.08 0.09 17.06 马尾松 85.08 0.20 14.72 47.91 6.17 45.82 0.10 0.00 17.77 -
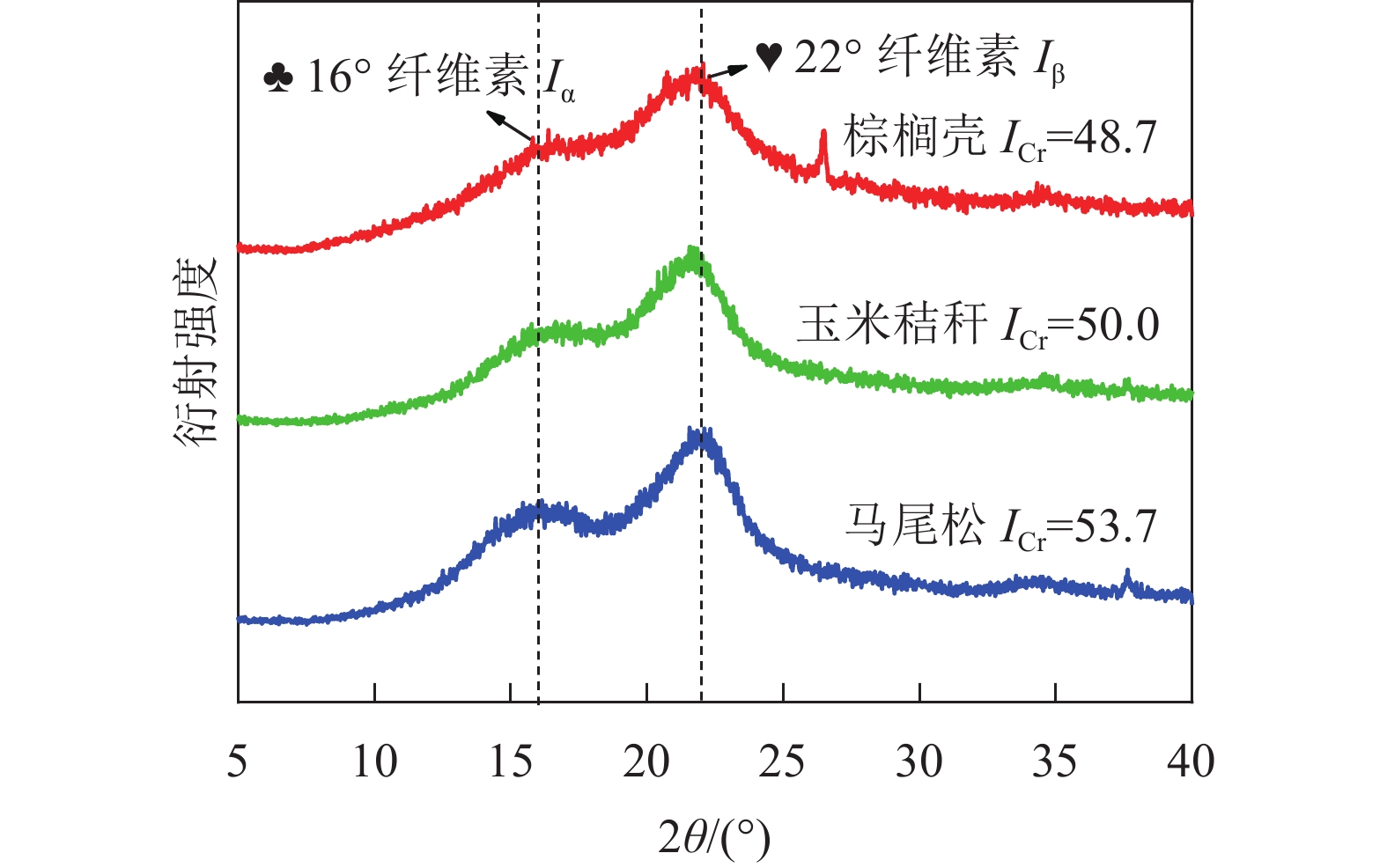
图2为3种不同类型生物质原料的XRD图谱。由图2可知:在衍射角为16°和22°处,3种原料均出现了2个衍射峰,分别代表纤维素结晶区的三斜晶系(Iα)和单斜晶系(Iβ)结构峰[8],但是3种原料的结晶度存在显著差别。在衍射角为22º时,马尾松的衍射峰较为尖锐,而玉米秸秆和棕榈壳的衍射峰则较为宽泛。根据式(1)计算得到3种原料的结晶度的大小依次为马尾松(53.7)、玉米秸秆(50.0)、棕榈壳(48.7)。生物质原料的结晶度与纤维素质量分数密切相关,马尾松作为木本生物质,纤维素质量分数最高(40%~60%),远高于草本生物质(30%~40%)与果壳类生物质(约25%)中纤维素质量分数,导致其结晶度最高[9]。
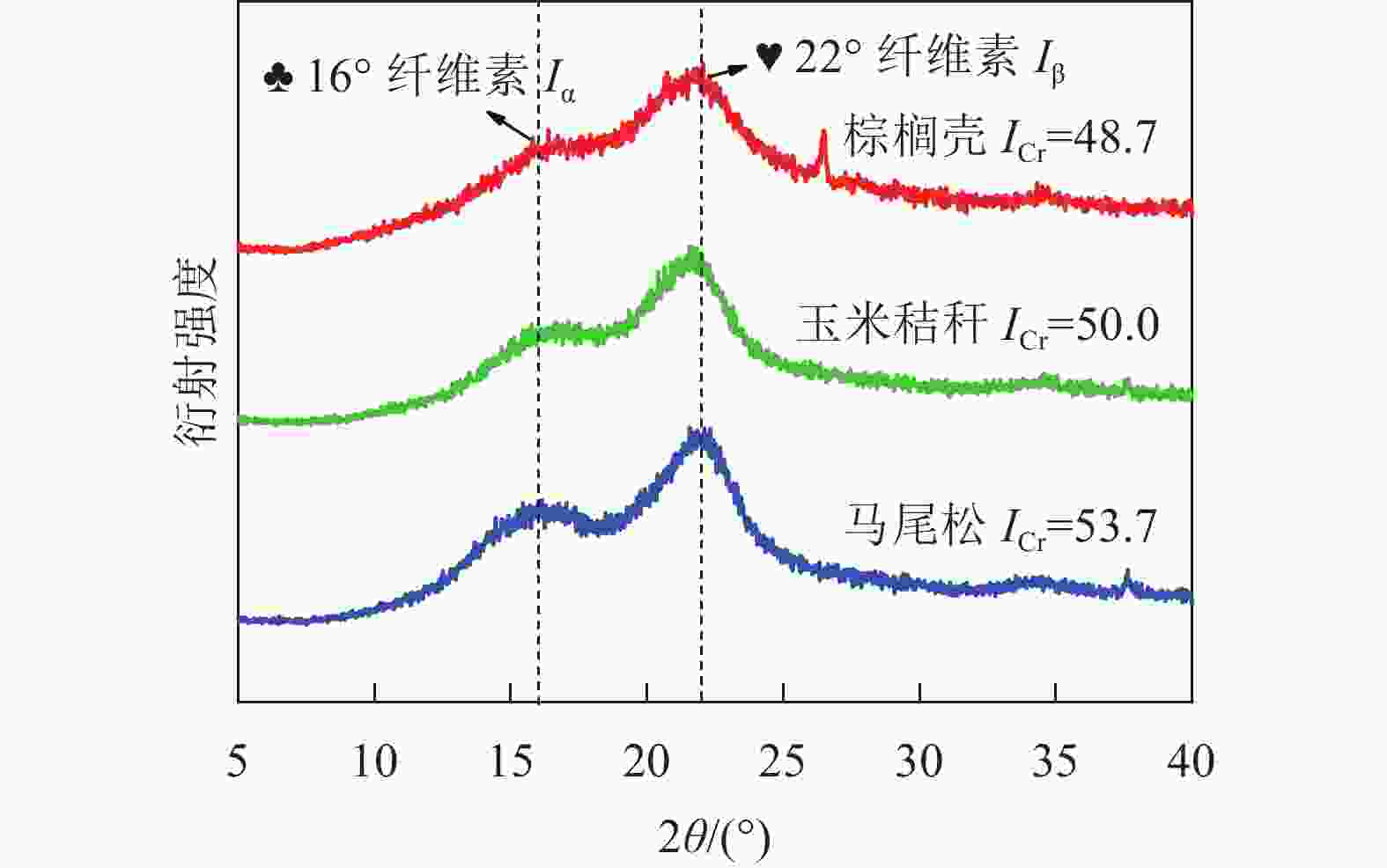
图 2 玉米秸秆、棕榈壳与马尾松原料的X射线衍射图
Figure 2. XRD patterns of corn stalk, palm kernel shell, and pine wood
-
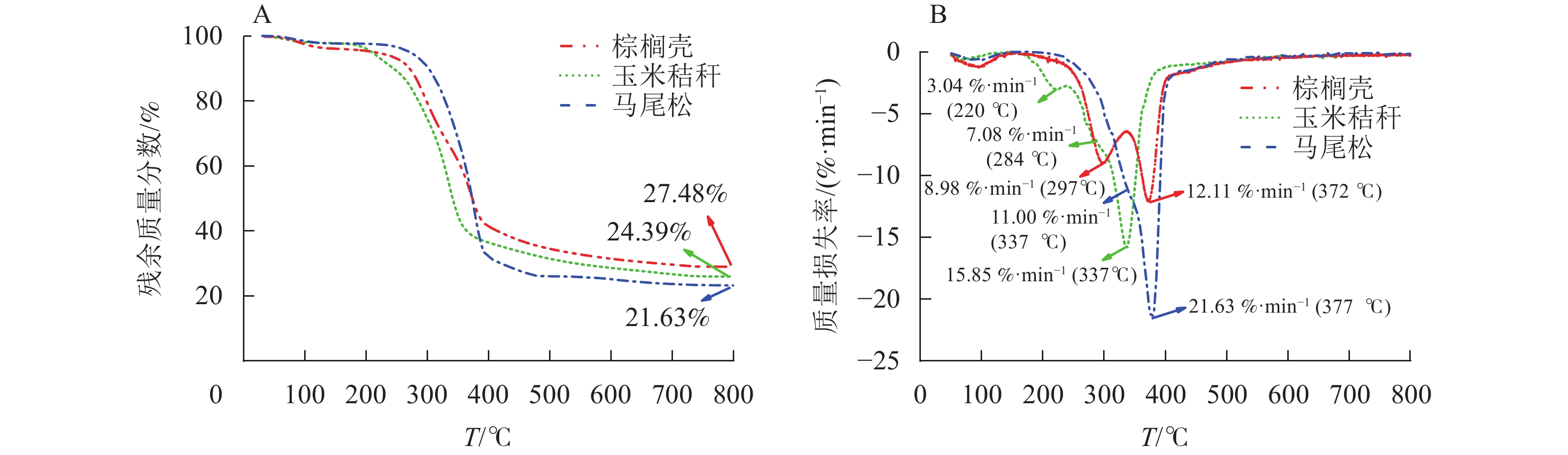
图3为3种不同类型生物质原料在升温速率为20 ℃·min−1时的热重(TG)和微商热重(DTG)曲线。由TG曲线可知:玉米秸秆、棕榈壳和马尾松的热解残余质量分别为24.39%、27.48%和21.63%,其中马尾松的残余质量最少,因为马尾松的挥发分质量分数最高,灰分质量分数最低。由DTG曲线可知:3种原料的热解过程主要分为3个阶段,分别为干燥阶段、快速热解阶段和慢速热解阶段。干燥阶段的温度范围为室温至150 ℃,其中105 ℃以下时,主要是自由水的挥发,而在105 ℃以上时,主要是结合水的挥发。快速热解阶段的温度范围为150~ 425 ℃,此阶段的失重主要来自于生物质中半纤维素、纤维素和木质素的逐步热降解[9−10]。结果表明:3种原料的快速热解阶段存在显著差异。玉米秸秆热解存在3个失重峰,分别位于220、284和337 ℃,位于左侧的2个失重峰主要来自于半纤维素的降解,失重率较小,而位于右侧的最大失重峰则来自于纤维素的降解,失重率为15.85%·min−1。棕榈壳的快速热解过程存在2个显著的失重峰,分别来自于半纤维素和纤维素。然而与其他2种原料相比,棕榈壳原料的2个失重峰间距较大,主要原因是棕榈壳中木质素质量分数较高。木质素是由苯丙烷基结构单元组成的具有三维结构的芳香族化合物,以共价键的形式与纤维素和半纤维素交联,形成稳定的生物质化学结构。因此,较高的木质素质量分数导致生物质热降解速率变慢,导致半纤维素和纤维素的热降解失重峰分离较为明显[11]。马尾松的快速热解阶段主要由1个轻微的肩峰和1个清晰的尖峰构成,分别来自于半纤维素和纤维素降解,尖峰处的失重率为21.63%·min−1,远高于其他2种原料的失重率,主要原因是马尾松的纤维素质量分数最高。慢速热解阶段的温度范围较广,主要是木质素的降解,木质素中连接键发生断裂,之后逐渐缩聚形成无定形结构炭[12]。

图 3 玉米秸秆、棕榈壳与马尾松热解过程的 TG (A)和 DTG (B)曲线
Figure 3. TG (A) and DTG (B) curves during pyrolysis of corn stalk, palm kernel shell, and pine wood
-
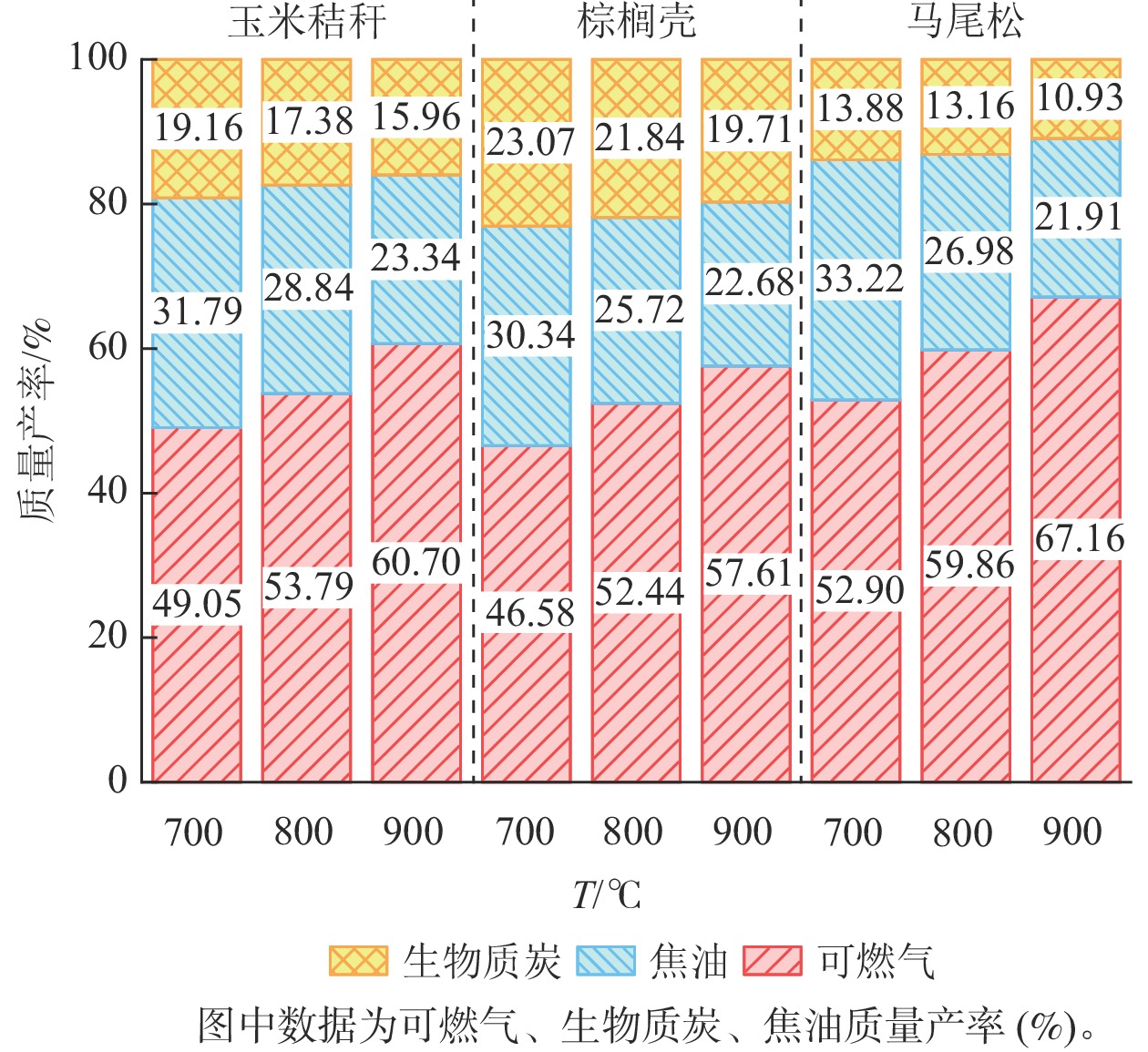
由图4可知:随着气化温度从700 ℃增加至900 ℃,所有原料的可燃气产率逐渐增加,而生物质炭和焦油的质量产率逐渐下降。生物质气化过程是由干燥、热解、氧化和还原等4个阶段组成。随着气化温度的增加,生物质原料在气化过程中的热解和氧化反应增强,消耗更多的固态原料,使其转化为挥发分气体,进而使得燃气的质量产率上升,生物质炭的质量产率下降。此外,较高的气化温度有利于焦油发生二次降解反应,转化为小分子的可燃气体,使得液体产物的质量产率下降[13−14]。MOHAMMED等[15]和GÁLVEZ-PÉREZ等[16]分别研究了气化温度对棕榈果废弃物和橄榄Canarium album饼气化产物产率的影响。结果表明:在更高的气化温度条件下,可燃气的产率呈上升的趋势,而生物质炭与焦油产率逐渐降低。主要原因是随着反应器内温度升高,气固反应更加充分,加剧了氧化和还原反应的发生,有利于焦油的裂解和重整反应,产气量逐渐增加,焦油产量减小。
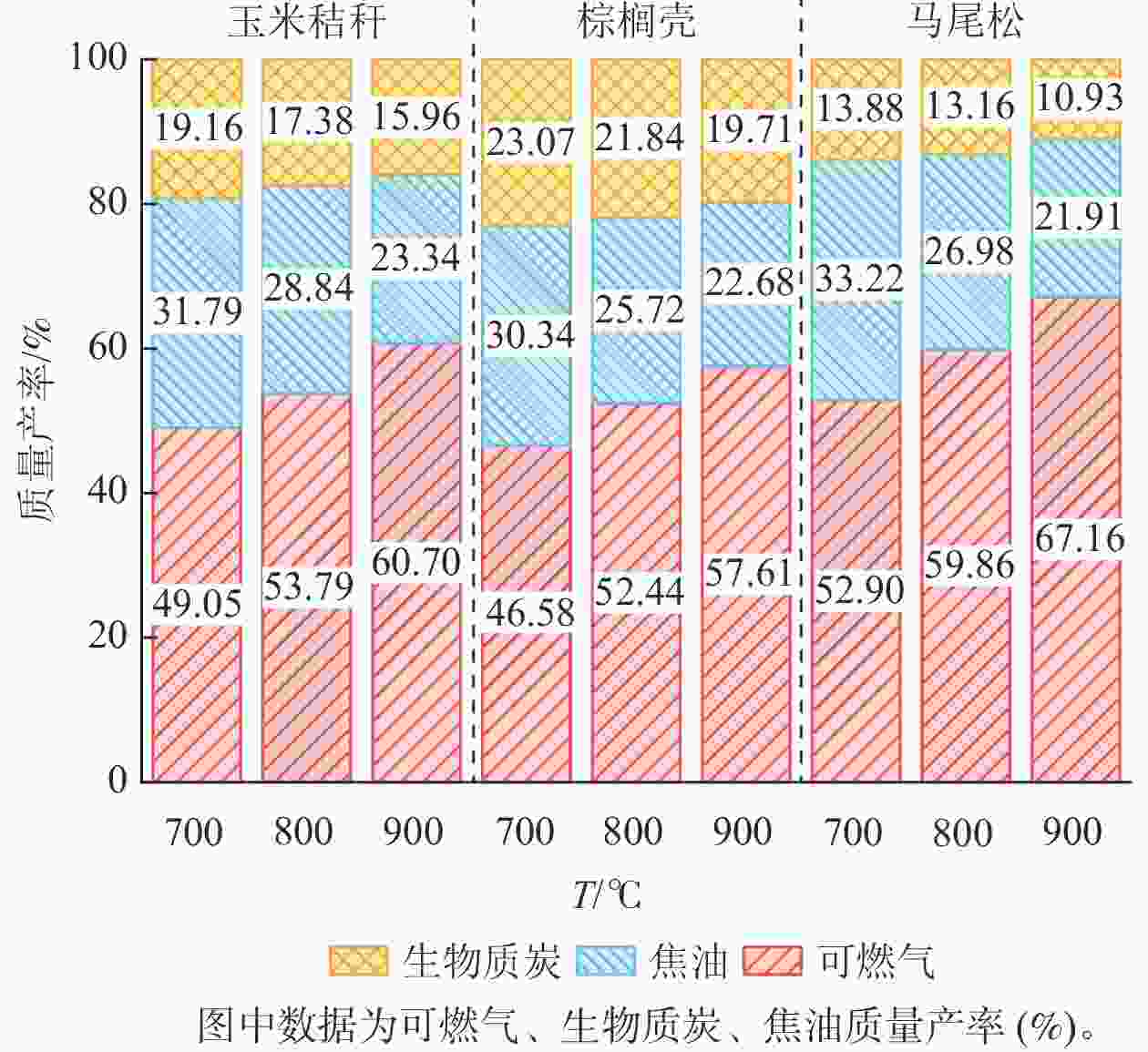
图 4 气化温度对玉米秸秆、棕榈壳与马尾松气化产物质量产率的影响
Figure 4. Effect of gasification temperatures on the mass yields of gasified products from corn stalk, palm kernel shell, and pine wood
然而,3种原料的气化产物的产率存在显著差异。当气化温度为900 ℃时,3种原料的可燃气质量产率从高到低依次为马尾松(67.16%)、玉米秸秆(60.70%)、棕榈壳(57.61%),生物质炭的质量产率从高到低依次为棕榈壳(19.72%)、玉米秸秆(15.96%)、马尾松(10.93%),焦油的质量产率从高到低依次为玉米秸秆(23.34%)、棕榈壳(22.68%)、马尾松(21.91%)。结果表明:在3种原料中,马尾松气化产生的可燃气质量产率最高,而生物质炭和焦油的质量产率最低。主要原因是马尾松的挥发分质量分数最高,导致其在热解和气化反应区产生更多的可燃气,并且马尾松的灰分质量分数最低,而灰分大部分都残留在生物质炭中,导致其生物质炭的产率最低[17]。
-
由图5可知:生物质原料气化的主要组分为H2、CO、CH4和CO2,其中CO为最主要的可燃气组分,体积分数远高于H2和CH4。随着气化温度从700 ℃增加至900 ℃,3种生物质可燃气中的CO、H2和CH4体积分数均逐渐增加,CO2体积分数逐渐减少,低位热值逐渐增加。以马尾松为例,CO、H2和CH4体积分数分别从37.95%、8.69%和15.67%增加至40.03%、18.27%和18.29%,而CO2体积分数则从14.88%减少至10.71%,低位热值从11.34 MJ·m−3增加至13.58 MJ·m−3。CERÓN等[18]研究了气化温度对挪威云杉Picea abies气化可燃气组分和热值的影响。结果表明:随着气化温度从750 ℃增加至950 ℃,CO和H2体积分数分别从10.4%和1.3%增加至21.4%和4.3%,而CO2则从3.8%下降至2.3%。主要原因如下:首先,气化温度的增加,促进了生物质在热解区的热降解反应,产生更多的挥发分气体;其次,较高的气化温度促进了燃烧区生物质炭中的C、H元素和气化剂中的O2之间的燃烧反应,进而生成更多的CO2和水(H2O);最后,燃烧区产生的CO2和H2O进入还原区,与生物质炭中的C元素发生水煤气转化反应和Boudouard反应,产生更多的CO和H2等可燃气体,而CH4主要来自于焦油的二次裂解反应和加氢气化反应,高温促进了加氢气化反应的发生,使得CH4体积分数增加[19−20]。
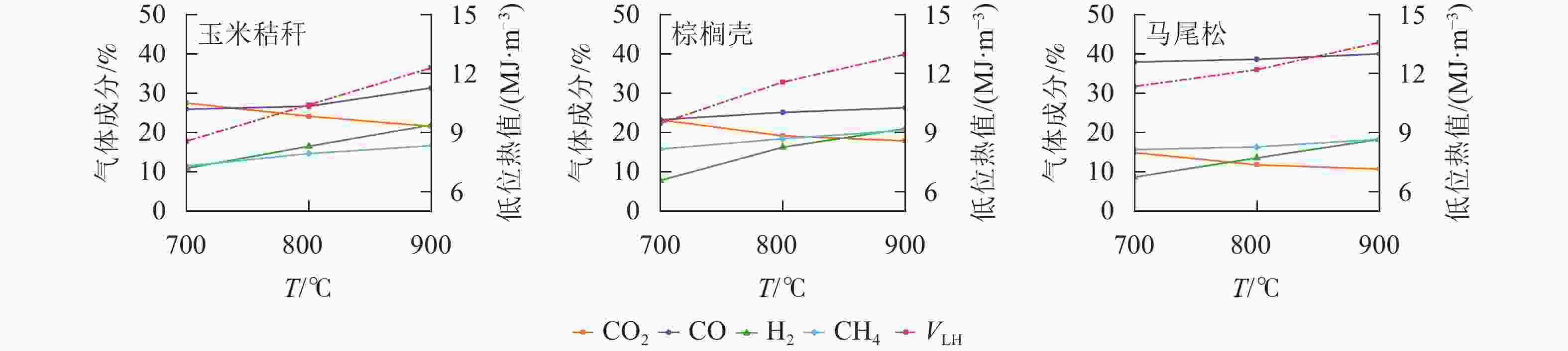
图 5 气化温度对玉米秸秆、棕榈壳与马尾松气化可燃气组分和热值的影响
Figure 5. Effect of gasification temperatures on the compound distribution and calorific values of gaseous product from corn stalk, palm kernel shell, and pine wood
3种生物质原料气化产生的可燃气组分和热值存在显著差异。当气化温度为900 ℃时,马尾松气化产生的CO体积分数最高,为40.03%,其次为玉米秸秆(31.37%)和棕榈壳(26.36%);玉米秸秆气化产生的H2体积分数最高,为21.93%,其次为棕榈壳(21.05%)和马尾松(18.27%);棕榈壳气化产生的CH4体积分数最高,为20.63%,其次为马尾松(18.29%)和玉米秸秆(16.65%)。马尾松气化产生可燃气的低位热值最高,达到13.59 MJ·m−3,表明在这3种原料中,马尾松最适宜作为气化原料。
-
通过对比表1和表2数据可知:与生物质原料相比,气化后生物质炭的C元素质量分数显著增加,H和O元素大幅度减少,挥发分质量分数显著下降,灰分质量分数大幅增加。首先,气化温度对生物质炭的基础特性具有显著影响。根据元素分析结果,随着气化温度的增加,玉米秸秆炭C元素质量分数逐渐减少,棕榈壳炭与马尾松炭的C元素质量分数逐渐增加,3种生物质炭的H与O元素质量分数均逐渐减少;其次,根据工业分析结果可知,随着气化温度的增加,3种生物质炭的挥发分质量分数均逐渐减少,灰分质量分数逐渐增加,玉米秸秆炭固定碳质量分数逐渐减少,而棕榈壳炭与马尾松炭的固定碳质量分数逐渐增加,因此,玉米秸秆炭的高位热值逐渐下降,而棕榈壳炭与马尾松炭的高位热值略有上升。其次,不同原料之间的基础特性差异也导致3种气化生物质炭之间存在显著差异。当气化温度为900 ℃时,马尾松炭的C元素和固定碳质量分数最高,为85.75%和79.37%;玉米秸秆炭的O元素、H元素、挥发分和灰分质量分数最高,分别为19.73%、1.77%、26.83%和24.11%。因此,3种原料中,马尾松气化炭的高位热值最高,而玉米秸秆炭高位热值最低,3种生物质炭的高位热值从高到低依次为马尾松炭(29.70 MJ·kg−1)、棕榈壳炭(23.97 MJ·kg−1)、玉米秸秆炭(17.69 MJ·kg−1)。
表 2 气化温度对玉米秸秆、棕榈壳与马尾松气化生物质炭基本特性的影响
Table 2. Effect of gasification temperature on the basic properties of bio-charfrom corn stalk, palm kernel shell, and pine wood
样品 T/℃ 元素分析 工业分析 高位热值/(MJ·kg−1) mC/% mH/% mO/% mN/% mS/% m挥发分/% m灰分/% m固定碳/% 玉米秸秆炭 700 65.03 2.02 20.45 0.02 0.03 31.49 12.45 56.06 21.51 玉米秸秆炭 800 61.93 1.82 20.26 0.01 0.00 29.52 15.98 54.50 20.21 玉米秸秆炭 900 54.37 1.77 19.72 0.01 0.02 26.83 24.11 49.06 17.69 棕榈壳炭 700 64.69 1.91 13.42 0.01 0.00 22.47 19.97 57.56 22.31 棕榈壳炭 800 68.49 1.71 9.73 0.01 0.00 19.61 20.06 60.33 23.88 棕榈壳炭 900 68.80 1.55 8.33 0.01 0.04 18.12 21.27 60.61 23.97 马尾松炭 700 83.58 2.16 10.93 0.00 0.05 21.35 2.71 75.94 29.39 马尾松炭 800 85.23 1.85 9.92 0.00 0.00 19.03 3.00 77.97 29.66 马尾松炭 900 85.75 1.74 9.78 0.00 0.02 17.92 3.28 79.37 29.70 说明:mO=100%−mC−mH−mN−mS−m灰分。 -
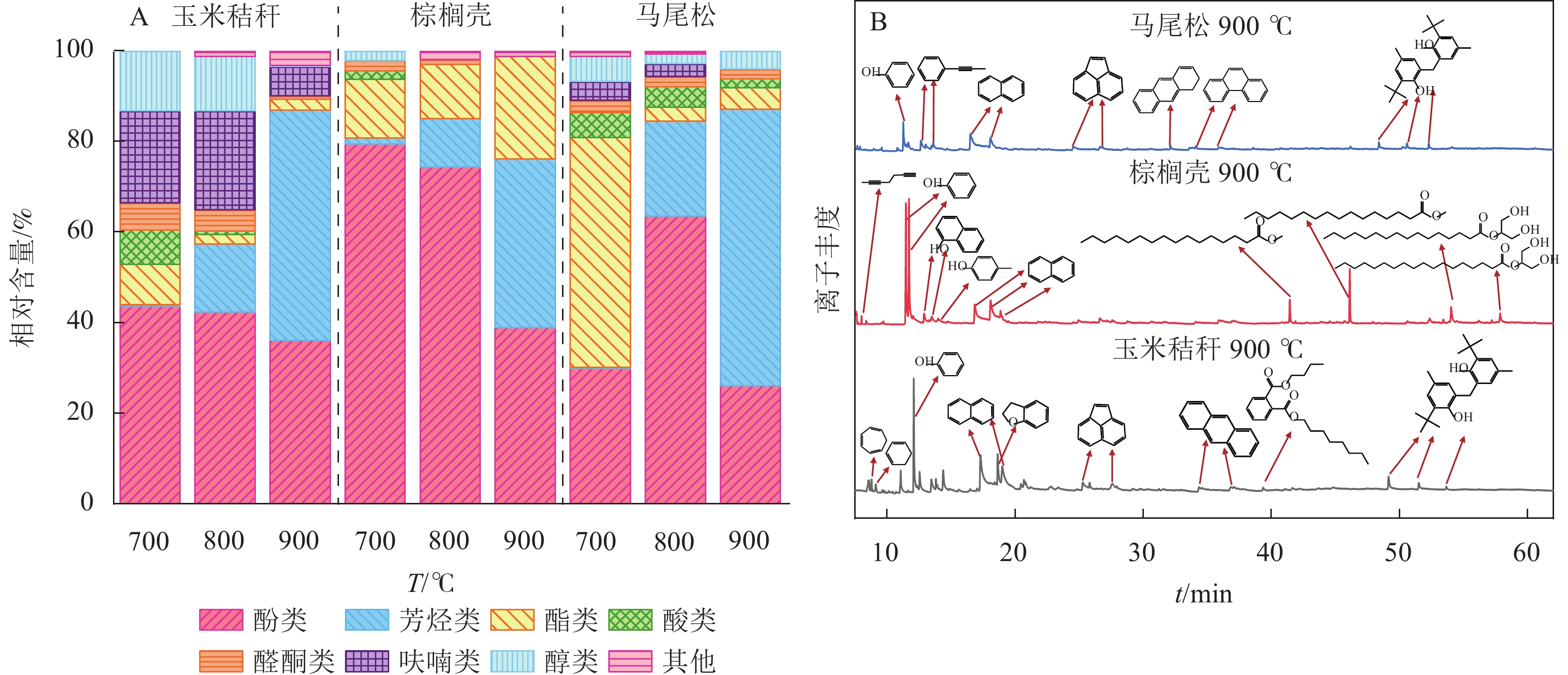
生物质气化的液体产物主要由酸类、醛类、酮类、醇类、酯类、酚类、芳烃类、呋喃类等有机化合物组成。由图6A可知:随着气化温度从700 ℃增加至900 ℃,所有原料气化后液体中酸类、醇类、醛酮类、呋喃类相对含量均呈递减趋势;玉米秸秆与马尾松气化液体中酯类相对含量逐渐下降,而棕榈壳呈相反趋势;玉米秸秆与棕榈壳气化液体产物的酚类物质相对含量的变化规律一致,而马尾松出现先增加后减小现象。此外,随气化温度的增加,玉米秸秆、棕榈壳、马尾松气化液体中芳烃类物质相对含量显著增加,分别从0.29%、1.30%、0.19%增加到50.86%、37.24%、61.12%。这是因为在高温下焦油中的酚类等含氧化合物逐步发生脱氧反应,使得芳构化程度逐渐加深,导致芳烃相对含量明显增加[21]。刘慧利等[22]研究了气化温度对液体产物组分含量的影响,发现在高温气化(700~900 ℃)条件下,液体产物中酚类化合物等含氧化合物极易发生脱氧反应,并最终形成了大量芳烃类化合物。
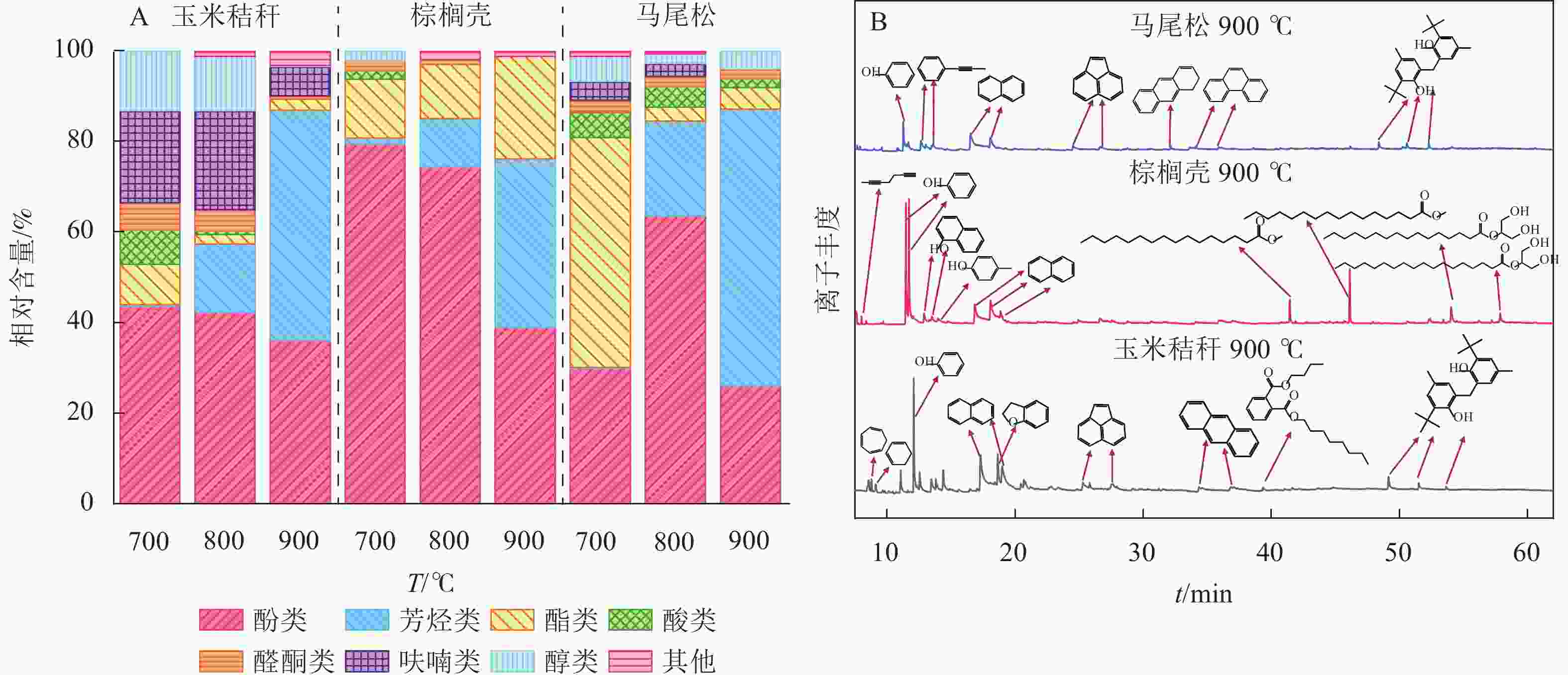
图 6 气化温度对玉米秸秆、棕榈壳与马尾松气化液体产物组分的影响
Figure 6. Effect of gasification temperatures on the liquid products of gaseous product from corn stalk, palm kernel shell, and pine wood
图6B为气化温度为900 ℃时,3种生物质原料气化液体产物的总离子色谱图。马尾松气化产生的芳烃类相对含量最高,为61.12%,其次为玉米秸秆(50.86%)和棕榈壳(37.24%),这是因为马尾松中纤维素相对含量最高(占40%~60%),在高温下裂解产生大量的含氧化合物,此类含氧化合物与烯烃会发生Diels-Alder反应,进而形成芳烃类物质[23]。玉米秸秆气化产生的呋喃类相对含量最高,为6.54%,棕榈壳和马尾松缺未检测出此类组分。这是因为半纤维素是草本植物的主要成分(占25%~39%),木聚糖经吡喃环开环反应生成木糖苷,C2或者C3位置上的4-O-甲基-D-葡萄糖醛酸取代基脱落,最终发生连续脱水反应形成多种呋喃类物质[24]。棕榈壳气化产生的酚类相对含量最高,为38.97%,其次为玉米秸秆(36.12%)和马尾松(26.13%),这与棕榈壳中木质素相对含量(约50%)较高有关,木质素主要是由愈创木基、紫丁香基和对羟基苯基等3种结构单元组成,通过木质素中的β—O—4醚键、C—C键等连接键的断裂,生成大量的酚类化合物[25]。
-
采用自主设计的微型固定床气化装置,研究了气化温度对马尾松、玉米秸秆和棕榈壳等3种生物质气化产物产率和特性的影响。结果表明:随着气化温度的增加,可燃气质量产率逐渐增加,而生物质炭和焦油质量产率逐渐减小;较高的气化温度有利于可燃气中可燃组分和热值的提高;较高的气化温度导致生物质炭中的C元素和灰分质量分数增加,而H和O元素及挥发分质量分数降低,热值增加;较高的气化温度导致焦油中酸类、醇类、醛酮类、呋喃类化合物相对含量逐渐减小,而芳烃类化合物相对含量大幅增加。
Effect of gasification temperature on the properties of the bio-gas, bio-char, and tar derived in biomass gasification
-
摘要:
目的 生物质气化技术是一种清洁可再生的能源利用技术。研究气化温度对气化产物特性的影响机制,构建气化产物特性的演变规律,对中国实现碳达峰与碳中和目标具有重大意义。 方法 以玉米Zea mays秸秆、棕榈Trachycarpus fortunei壳和马尾松Pinus massoniana等3种农林生物质废弃物为原料,以空气为气化剂,采用自制的微型固定床气化装置,开展3种生物质的气化多联产研究,系统研究了不同气化温度(700、800、900 ℃)对气化性能的影响,以及对气化三相产物(可燃气、生物质炭与焦油)特性的影响机制。 结果 随着气化温度从700 ℃增加至900 ℃,可燃气的质量产率逐渐增加,而生物质炭和焦油的质量产率逐渐减小。较高的气化温度有利于提升可燃气的热值。在3种原料中,马尾松可燃气的热值最高,一氧化碳、氢气和甲烷体积分数分别为40.03%、18.27%和18.29%,低位热值达13.58 MJ·m−3。随着气化温度的增加,生物质炭中的碳元素质量分数增加,氢元素质量分数下降,灰分质量分数大幅增加,其中马尾松炭的热值最高,达29.70 MJ·kg−1。随着气化温度的增加,3种原料的气化液体产物中酸类、醇类、醛酮类、呋喃类化合物相对含量逐渐减小,而芳烃类化合物相对含量大幅增加,其中马尾松的芳烃类化合物相对含量最高,达到61.12%。 结论 气化温度对生物质气化三相产物的产量及特性有显著的影响,选择合适的气化温度对提高生物质可燃气热值及气化效率至关重要。图6表2参25 Abstract:Objective Biomass gasification technology is a clean and renewable energy utilization approach. The purpose of this study is to investigate the influence mechanism of gasification temperature on the properties of gasification products, and to build the evolution law of the characteristics of gasification products, which is of great significance for China to achieve the goal of carbon peaking and carbon neutrality. Method Three types of lignocellulosic biomass, namely Zea mays, Trachycarpus fortunei, and Pinus massoniana, were gasified in a home-made fixed bed gasification reactor. The effect of different gasification temperatures (GT) (700, 800, 900 ℃) on gasification performance were systematically studied, as well as the mechanism of their effects on the characteristics of gasification products (bio-gas, bio-char, and tar). Result With the increase of GT from 700 ℃ to 900 ℃, the yield of bio-gas gradually increased, while the yields of bio-char and tar gradually decreased. Higher GT resulted in better quality of bio-gas. Among the three types of biomass, bio-gas from P. massoniana had the highest calorific value. The contents of CO, H2, and CH4 were 40.03%, 18.27%, and 18.29%, respectively, and the lower calorific value was 13.58 MJ·m−3. With the increase of GT, the contents of C and ash increased, while the contents of H decreased. The calorific value of bio-char from P. massoniana was the highest, reaching 29.70 MJ·kg−1. With the increase of GT, the content of acids, alcohols, aldehydes, ketones and furans in the liquid products of biomass gasification gradually decreased, while the content of aromatics increased significantly, with the highest aromatic content in P. massoniana, reaching 61.12%. Conclusion GT has a significant influence on the yield and properties of gasified products. Optimizing the gasification temperature is crucial for improving the calorific value and gasification efficiency. [Ch, 6 fig. 2 tab. 25 ref.] -
Key words:
- biomass /
- gasification poly-generation /
- gasification temperature /
- bio-gas /
- bio-char /
- tar
-
图 2 玉米秸秆、棕榈壳与马尾松原料的X射线衍射图
Figure 2 XRD patterns of corn stalk, palm kernel shell, and pine wood
图 3 玉米秸秆、棕榈壳与马尾松热解过程的 TG (A)和 DTG (B)曲线
Figure 3 TG (A) and DTG (B) curves during pyrolysis of corn stalk, palm kernel shell, and pine wood
图 4 气化温度对玉米秸秆、棕榈壳与马尾松气化产物质量产率的影响
Figure 4 Effect of gasification temperatures on the mass yields of gasified products from corn stalk, palm kernel shell, and pine wood
图 5 气化温度对玉米秸秆、棕榈壳与马尾松气化可燃气组分和热值的影响
Figure 5 Effect of gasification temperatures on the compound distribution and calorific values of gaseous product from corn stalk, palm kernel shell, and pine wood
图 6 气化温度对玉米秸秆、棕榈壳与马尾松气化液体产物组分的影响
Figure 6 Effect of gasification temperatures on the liquid products of gaseous product from corn stalk, palm kernel shell, and pine wood
表 1 玉米秸秆、棕榈壳和马尾松的元素分析、工业分析和热值
Table 1. Ultimate analysis, proximate analysis, and calorific values of corn stalk, palm kernel shell, and pine wood
样品 工业分析 元素分析 高位热值/(MJ·kg−1) m挥发分/% m灰分/% m固定碳/% mC/% mH/% mO/% mN/% mS/% 玉米秸秆 77.79 3.38 18.83 45.75 5.80 47.75 0.49 0.21 16.16 棕榈壳 73.04 4.51 22.45 48.50 5.53 44.80 1.08 0.09 17.06 马尾松 85.08 0.20 14.72 47.91 6.17 45.82 0.10 0.00 17.77  下载: 导出CSV
下载: 导出CSV
表 2 气化温度对玉米秸秆、棕榈壳与马尾松气化生物质炭基本特性的影响
Table 2. Effect of gasification temperature on the basic properties of bio-charfrom corn stalk, palm kernel shell, and pine wood
样品 T/℃ 元素分析 工业分析 高位热值/(MJ·kg−1) mC/% mH/% mO/% mN/% mS/% m挥发分/% m灰分/% m固定碳/% 玉米秸秆炭 700 65.03 2.02 20.45 0.02 0.03 31.49 12.45 56.06 21.51 玉米秸秆炭 800 61.93 1.82 20.26 0.01 0.00 29.52 15.98 54.50 20.21 玉米秸秆炭 900 54.37 1.77 19.72 0.01 0.02 26.83 24.11 49.06 17.69 棕榈壳炭 700 64.69 1.91 13.42 0.01 0.00 22.47 19.97 57.56 22.31 棕榈壳炭 800 68.49 1.71 9.73 0.01 0.00 19.61 20.06 60.33 23.88 棕榈壳炭 900 68.80 1.55 8.33 0.01 0.04 18.12 21.27 60.61 23.97 马尾松炭 700 83.58 2.16 10.93 0.00 0.05 21.35 2.71 75.94 29.39 马尾松炭 800 85.23 1.85 9.92 0.00 0.00 19.03 3.00 77.97 29.66 马尾松炭 900 85.75 1.74 9.78 0.00 0.02 17.92 3.28 79.37 29.70 说明:mO=100%−mC−mH−mN−mS−m灰分。
下载: 导出CSV
-
[1] 张会岩, 杨海平, 陆强, 等. 生物质定向热解制取高品质液体燃料、化学品和碳材料研究进展[J]. 工程热物理学报, 2021, 42(12): 3031 − 3044. ZHANG Huiyan, YANG Haiping, LU Qiang, et al. Progress of directional pyrolysis of biomass to produce high-quality liquid fuels, chemicals and carbon materials [J]. Journal of Engineering Thermophysics, 2021, 42(12): 3031 − 3044. [2] 马中青, 张齐生. 温度对马尾松热解产物产率和特性的影响[J]. 浙江农林大学学报, 2016, 33(1): 109 − 115. MA Zhongqing, ZHANG Qisheng. Pinus Massoniana pyrolysis: influence of temperature on yields and product properties [J]. Journal of Zhejiang A&F University, 2016, 33(1): 109 − 115. [3] 马中青, 张齐生, 周建斌, 等. 下吸式生物质固定床气化炉研究进展[J]. 南京林业大学学报(自然科学版), 2013, 37(5): 139 − 145. MA Zhongqing, ZHANG Qisheng, ZHOU Jianbin, et al. Research progress of downdraft biomass fixed-bed gasifier [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2013, 37(5): 139 − 145. [4] 周建斌, 周秉亮, 马欢欢, 等. 生物质气化多联产技术的集成创新与应用[J]. 林业工程学报, 2016, 1(2): 1 − 8. ZHOU Jianbin, ZHOU Bingliang, MA Huanhuan, et al. Integrated innovation and application of biomass gasification poly-generation technology [J]. Journal of Forestry Engineering, 2016, 1(2): 1 − 8. [5] 张齐生, 马中青, 周建斌. 生物质气化技术的再认识[J]. 南京林业大学学报(自然科学版), 2013, 37(1): 1 − 10. ZHANG Qisheng, MA Zhongqing, ZHOU Jianbin. History, challenge and solution of biomass gasification: a review [J]. Journal of Nanjing Forestry University (Natural Science Edition), 2013, 37(1): 1 − 10. [6] 成亮, 周建斌, 章一蒙, 等. 糠醛渣和废菌棒的热解气化多联产再利用[J]. 农业工程学报, 2017, 33(21): 231 − 236. CHENG Liang, ZHOU Jianbin, ZHANG Yimeng, et al. Re-utilization of furfural residues and wasted mushroom inoculation bags by multi-production gasification technology [J]. Transactions of the Chinese Society of Agricultural Engineering, 2017, 33(21): 231 − 236. [7] 王亮才, 朱正祥, 马欢欢, 等. 紫茎泽兰杆热解气化及其产物研究[J]. 林业工程学报, 2018, 3(6): 98 − 103. WANG Liangcai, ZHU Zhengxiang, MA Huanhuan, et al. Pyrolysis, gasification and its products of Eupatorium adenophorum [J]. Journal of Forestry Engineering, 2018, 3(6): 98 − 103. [8] 张雨, 王浚浩, 马中青, 等. 温度对竹材烘焙过程中气固液三相产物组成及特性的影响[J]. 农业工程学报, 2018, 34(18): 242 − 251. ZHANG Yu, WANG Junhao, MA Zhongqing, et al. Effects of torrefaction temperature on composition and characteristics of gas-solid-liquid three-phase products in bamboo torrefaction process [J]. Transactions of the Chinese Society of Agricultural Engineering, 2018, 34(18): 242 − 251. [9] 朱锡锋. 生物质热解原理与技术[M]. 合肥: 中国科学技术大学出版社, 2006: 43 − 49. ZHU Xifeng. Biomass Pyrolysis Principle and Technology [M]. Hefei: Press of University of Science and Technology of China, 2006: 43 − 49. [10] YANG Haiping, YAN Rong, CHEN Hanping, et al. Characteristics of hemicellulose, cellulose and lignin pyrolysis [J]. Fuel, 2007, 86(12/13): 1781 − 1788. [11] 马中青, 徐嘉炎, 叶结旺, 等. 基于热重红外联用和分布活化能模型的樟子松热解机理研究[J]. 西南林业大学学报, 2015, 35(3): 90 − 96. MA Zhongqing, XU Jiayan, YE Jiewang, et al. Determination of pyrolysis characteristics and kinetics of Pinus sylvestris var. mongolica using TGA-FTIR and distributed activation energy model [J]. Journal of Southwest Forestry University, 2015, 35(3): 90 − 96. [12] MA Zongqing, YANG Youyou, MA Qianqian, et al. Evolution of the chemical composition, functional group, pore structure and crystallographic structure of bio-char from palm kernel shell pyrolysis under different temperatures [J]. Journal of Analytical and Applied Pyrolysis, 2017, 127(38): 350 − 359. [13] GONZÁLEZ J F, SUEROS R, BRAGADO D, et al. Investigation on the reactions influencing biomass air and air/steam gasification for hydrogen production [J]. Fuel Processing Technology, 2008, 89(8): 764 − 772. [14] REN Jie, CAO Jingpei, ZHAO Xiaoyan, et al. Recent advances in syngas production from biomass catalytic gasification: acritical review on reactors, catalysts, catalytic mechanisms and mathematical models [J/OL]. Renewable and Sustainable Energy Reviews, 2019, 116: 109426[2022-10-01]. doi: 10.1016/j.rser.2019.109426. [15] MOHAMMED M A A, SALMIATON A, WAN A W A K G, et al. Air gasification of empty fruit bunch for hydrogen-rich gas production in a fluidized-bed reactor [J]. Energy Conversion and Management, 2011, 52(2): 1555 − 1561. [16] GÁLVEZ-PÉREZ A, MARTÍN-LARA M A, CALERO M, et al. Experimental investigation on the air gasification of olive cake at low temperatures [J/OL]. Fuel Processing Technology, 2021, 213: 106703[2022-11-01]. doi: 10.1016/j.fuproc.2020.106703. [17] 刘晓, 杨晓辉, 李现奇, 等. 生物质成型燃料化学实验分析[J]. 节能, 2017, 36(4): 16 − 19. LIU Xiao, YANG Xiaohui, LI Xianqi, et al. The chemical experiment analysis of the biomass briquette fuel [J]. Energy Conservation, 2017, 36(4): 16 − 19. [18] CERÓN L A, KONIST A, LEES H, et al. Effect of woody biomass gasification process conditions on the composition of the producer gas [J]. Sustainability, 2021, 13(21): 1 − 17. [19] JARUNGTHAMMACHOTE S, DUTTA A. Experimental investigation of a multi-stage air-steam gasification process for hydrogen enriched gas production [J]. International Journal of Energy Research, 2012, 36(3): 335 − 345. [20] 高宁博, 李爱民, 全翠. 生物质高温气化重整制氢实验研究[J]. 太阳能学报, 2014, 35(5): 911 − 917. GAO Ningbo, LI Aimin, QUAN Cui, et al. Hydrogen production of biomass high steam temperaturegasification and reforming [J]. Acta Energiae Solaris Sinica, 2014, 35(5): 911 − 917. [21] CORTAZAR M, ALVAREZ J, LOPEZ G, et al. Role of temperature on gasification performance and tar composition in a fountain enhanced conical spouted bed reactor [J]. Energy Conversion and Management, 2018, 171: 1589 − 1597. [22] 刘慧利, 胡建杭, 王华, 等. 温度对松木屑气化液体产物组成的影响[J]. 燃料化学学报, 2013, 41(2): 189 − 197. LIU Huili, HU Jianhang, WANG Hua, et al. Effect of temperature on composition of liquid products from pine sawdust gasification [J]. Journal of Fuel Chemistry and Technology, 2013, 41(2): 189 − 197. [23] 俞海淼, 刘阳, 武子璐. 生物质三组分催化气化后焦油析出特性研究[J]. 现代化工, 2018, 38(8): 90 − 93. YU Haimiao, LIU Yang, WU Zilu. Characteristics of tar formation during catalytic gasification of three-component biomass [J]. Modern Chemical Industry, 2018, 38(8): 90 − 93. [24] SHEN Dekui, GU Sai, BRIDGWATER A V. The thermal performance of the polysaccharides extracted from hardwood: cellulose and hemicellulose [J]. Carbohydrate Polymers, 2010, 82(1): 39 − 45. [25] 左宋林, 于佳, 车颂伟. 热解温度对酸沉淀工业木质素快速热解液体产物的影响[J]. 燃料化学学报, 2008, 36(2): 144 − 148. ZUO Songlin, YU Jia, CHE Songwei. Effect of pyrolysis temperature on pyrolysate during fast pyrolysis of industrial lignin made by acid precipitation [J]. Journal of Fuel Chemistry and Technology, 2008, 36(2): 144 − 148. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220693
 点击查看大图
点击查看大图

计量
- 文章访问数: 2803
- HTML全文浏览量: 996
- PDF下载量: 63
- 被引次数: 0
