




-
人为活动已导致全球气温升高大约1.0 ℃,若温室气体排放速率持续不变,则可能在2030—2052年气温升高1.5 ℃[1, 2]。作为一个大型农业国家,中国的农业碳排放量仅次于工业。如何确保达到2030年的碳达峰以及2060年的碳中和,控制农业中的碳排放量尤为重要[3]。生物质炭是一种热稳定、廉价的炭产品,人类在数千年前就开始了它的制作和应用[4],将被遗弃的生物资源转化成为多功能材料,为资源可持续利用提供有效的生产方式,甚至可以作为升级传统农业系统的有效工具[5]。在农业应用方面,生物质炭已被证明可以提高土壤肥力、促进植物生长、增加作物产量[6],尤其是与有机肥一起施用对营养不良以及退化土壤的改良效果更加明显[7]。此外,有研究表明:广泛施用生物质炭可以减轻当前人为产生的二氧化碳12%的排放量[8−9]。
随着人口增长和饮食习惯的改变,对水果的需求不断增长,园艺作物的生产规模逐年增加[10],然而水果不能食用的部分以及作物修剪产生的枝条和树叶等已成为数量巨大的废弃物[11]。据统计,全球每年大约13亿t的食物被丢弃或浪费[12],其中水果和蔬菜占总量的42%以上[13]。同时,许多水果产地土壤营养缺乏,影响水果的可持续产出,造成农户经济损失。水果生产过程中产生的果园废弃物是一种潜在的生物资源,可以将其转化为有价值的材料[5]。如何循环利用这些废弃物资源,如何用于土壤养分补充和土壤改良是一个亟待解决的重大课题。
本研究对目前果园废弃物制备生物质炭的原理和性质以及在果园生产中的应用进行综述,为果园废弃物生物质炭的制备提供借鉴,以促进果园废弃物生物质炭的应用推广,有效推动果园生产的可持续发展。
-
生物质炭的性状不仅因原料不同有所差异,而且还受到制备过程中各项参数的影响[14]。目前,可通过热解、气化、烘烤等热化学技术转化后获得生物质炭[15],其中热解因操作简单、方便、可靠,成为采用最广泛的方法[16−17]。用来制备生物质炭的果园废弃物有果皮、果壳、花梗、茎叶、树叶、树皮和树枝等(表1)。
表 1 果园废弃物生物质炭制作条件和性质
Table 1. Methods and properties of orchard waste biochar production
原材料 热解温度/℃ 热解时间/h 有机碳/(g·kg−1) 全碳/% 全氮/% 全磷/% 全钾/% 炭产量/% 橙子皮 350 3 65.58 2.26 0.66 0.05 32.50 100~600 1 10.85 1.56 300 2 80.72 2.55 350 2 8.58 0.17 12.20 18.62 柑橘枝 500 2 80.60 0.87 0.30 1.08 香蕉花梗 300 1 37.79 66.00 500 1 41.86 40.00 香蕉皮 350 3 5.54 1.90 1.26 0.42 40.00 香蕉茎叶 500 2 462.50 1.36 0.54 4.27 苹果树枝 450 1 82.15 1.28 苹果树皮 450 1 68.96 2.93 苹果枝 700 2 74.21 1.29 32.60 300 2 62.20 1.69 47.94 600 2 80.10 1.28 28.48 椰子壳 400 47.92 1.34 43.33 600 58.43 1.02 36.00 800 67.31 0.87 34.00 1000 74.08 0.50 31.00 海枣树废弃物 300 57.99 0.54 49.97 400 66.87 0.45 36.54 500 72.30 0.42 32.38 600 72.89 0.39 30.88 700 73.42 0.35 28.84 800 74.63 0.31 27.40 原材料 灰分/% 挥发物/% 水分/% pH 比表面积/(m2·g−1) 碳氮比 文献 橙子皮 7.80 21.40 6.20 9.97 SIAL等[22] 5.50 70.00 13.00 9.43 STELLA等[23] 352.50 31.65 ADENIYI等[24] 9.10 10.02 OGUNKUNLE等[25] 柑橘枝 9.23 4.85 92.64 逄玉万等[26] 香蕉花梗 24.84 33.69 26.28 8.10 KARIM等[27] 35.48 24.37 21.26 10.20 香蕉皮 9.80 26.50 5.20 10.88 SIAL等[28] 香蕉茎叶 19.05 10.33 80.51 340.07 徐广平等[29] 苹果树枝 2.01 9.47 40.38 64.18 石钧元等[30] 苹果树皮 12.00 12.36 55.14 23.54 苹果枝 8.90 10.00 57.53 曹辉等[31] 6.72 60.77 2.39 36.80 ZHAO等[32] 9.40 14.86 108.59 62.58 椰子壳 26.27 30.56 8.50 39.57 35.76 SUMAN等[33] 21.99 20.82 9.64 120.73 57.28 17.32 5.33 10.87 256.60 77.37 12.03 4.72 11.02 590.41 148.46 海枣树废弃物 14.42 40.08 3.29 107.39 USMAN等[34] 16.34 20.25 3.13 148.60 19.68 9.31 2.96 172.14 20.71 6.85 2.25 186.90 21.05 5.47 2.12 209.77 21.39 3.91 2.09 240.74 说明:橙子Citrus sinensis;柑橘C. reticulata;香蕉Musa nana;苹果Malus pumila;椰子Cocos nucifera;海枣树 Phoenix dactylifera。表中百分数均为质量分数 -
果园废弃物主要由半纤维素(质量分数为20%~40%)、纤维素(质量分数为35%~55%)和木质素(质量分数为10%~25%)组成[18],三者的组成差异将极大地影响热分解过程。半纤维素是具有支链结构的多糖,由于不同来源生物质材料的半纤维素组成不同,其结构比纤维素更复杂。在低温下(<250 ℃),半纤维素的分解主要包括脱羟、脱羧和去除不稳定的碳水化合物支链,随着温度的升高,主链上的糖苷键被切断释放出木糖;温度达400 ℃以上时,由于木糖和糖基的分解,脂肪烃支链的芳香环化反应得到加强,生物质炭中形成芳香环结构;在高温下(>600 ℃),单核芳香环之间的缩合反应得到加强,并逐渐形成融合环结构[19−20]。纤维素是一种由葡萄糖环的β-1,4-糖苷键连接的线性聚合物,与半纤维素结构不同,其很少有支链结构,热稳定性较高,且热解过程中吸热,而半纤维素和木质素为放热。在低温下(<300 ℃),纤维素主要通过分子间和分子内氢键的断裂以及羟基的脱水而降解;当温度超过500 ℃时,强烈的脱氧反应(如脱羧)会导致含氧官能团的去除,同时低聚物继续脱水,迅速形成苯环结构,由此产生的生物质炭呈现出高度无序的交联结构;在600~800 ℃时,形成的生物质炭发生脱氢、脱甲基等反应,使芳香环结构变为大分子碳结构,生物质炭变得更加石墨化[19−21]。木质素具有多种不同化学键的复杂三维大分子结构,其基本结构单元为苯丙烷,在三者中最为稳定。木质素大约在200 ℃时就开始分解,在200~300 ℃时木质素发生侧链键裂解反应,形成少量芳香环化合物;随着温度的升高,其中的官能团与苯基丙烷反应形成酚类化合物和芳香烃;在500 ℃以上时,会发生芳香族开环和缩合反应,且由联苯基反应和苯类化合物之间的缩合反应形成多环芳烃;在600~800 ℃时会形成更多的融合环结构。在木质素的分子结构中,连接苯基丙烷单元和侧链的氧桥键在加热时容易被切断,形成含有苯环的活性自由基。这些自由基很容易与其他分子或自由基反应生成具有更稳定结构的大分子,最终形成生物质炭[19, 35]。
总的来说,果园废弃物的热解过程大致包括生物质脱水、挥发性物质质量分数减少和固体分解[36−42]。随着温度升高,脱氧反应导致含氧官能团的数量减少,以及缩聚反应形成更多多环芳烃的热解,导致生物质炭中羧基碳和邻烷基碳的碳结构降低,而芳烃基碳的碳结构增强[19]。同时,随着温度升高,三者的演化相似,呈现出碳质量分数增加,氢、氧质量分数降低,颜色变暗的趋势;半纤维素生物质炭和木质素生物质炭的体积减小,纤维素生物质炭成为一种泡沫状材料,且体积变大。WAN等[43]研究发现:木质素和木聚糖生物质炭呈球形,而纤维素生物质炭为长条纤维状,进而推断纤维素可能在生物质炭的热解过程中首先形成,起到骨架的作用,对生物质炭的形状具有重要意义。此外,三者在相同的温度下碳质量分数存在显著差异。
-
果树不同部位的废弃物制得的生物质炭性质不同,因此对果皮类废弃物进一步分类有助于制备差异化的生物质炭。研究表明:把橙子皮分为橙子皮和橙子皮络可制得不同性状的生物质炭,与橙子皮络生物质炭相比,橙子皮生物质炭碳质量分数更高,表面更加光滑,其多孔性更明显,并具有更多的活性位点,因此具有更好的催化活性、保水能力和吸附能力,在温室气体排放和固碳效应方面更有效;而橙子皮络生物质炭具有无机元素质量分数高,表面高度曲卷,表面积略高等优点[24]。与废弃物原料相比,废弃物生物质炭固定碳质量分数高,挥发性物质质量分数低,比表面积和孔隙空间大;与传统的直接填埋相比,废弃物生物质炭可有效降低二氧化碳、甲烷和一氧化二氮排放,减少经济损失和环境破坏[22, 28]。
果园果壳、花梗类废弃物制备的生物质炭具有多孔、孔径多样、表面积大、含大量含氧官能团的特点,其中果壳生物质炭pH较低,茎叶和树叶类钾质量分数较高。有研究表明:由杏Prunus armeniaca壳制备的生物质炭在解决农业点源污染特别是农药残留污染防治方面具有良好的应用前景[44−45]。SUMAN等[33]研究表明:随着温度的升高,椰子壳废料产生的生物质炭产量、含水量以及挥发性物质、氮、氢元素质量分数均有所降低,但热值较高,灰分、碳元素、芳香性、pH和表面积有所增加,且在400~1 000 ℃下得到的生物质炭与土壤改良、固碳效果有更强的相关性。KARIM等[46]对香蕉花梗进行不同的等离子体处理研究表明:获得的富钾生物质炭在土壤固碳、保水能力方面效果显著,可应用于酸性、缺钾土壤和喜钾作物中。此外,等离子体处理有助于水溶性钾组分转化为可交换的碳酸氢钾(KHCO3)矿物形式,从而增强钾的有效利用。
果园树皮和树枝类废弃物制备的生物质炭通常具有较高的产量,其中含有较高的碳和较低的灰分。PARK等[47]研究了梨Pyrus、苹果、柿Diospyros kaki等3种树枝在300~600 ℃温度下制备的生物质炭,随着裂解温度的升高,3种原料生物质炭的产量均下降,其中苹果枝下降最慢,为14.0%;氢和氧元素均下降,氮元素却先上升后下降;比表面积均增加,并以梨枝增加最快,从1.5 m2·g−1增加到134.2 m2·g−1;产物中的碳质量分数也均增加,其中苹果枝增量最大,为11.92%;它们的傅里叶转换红外光谱(FTIR光谱)均相似(C—H拉伸带除外)。ZHAO等[32]研究发现:当热解温度从300 ℃升高到500 ℃时,苹果枝生物质炭的酸性官能团降低,以羧基官能团的含量降低最为明显,而基本官能团则呈相反的趋势。一般来说,较高裂解温度下(>400 ℃)生物质炭主要拥有芳香碳结构和高热稳定性,将其施于土壤中,可以储存碳的方式有助于缓解气候变化,同时低温(<400 ℃)制备的生物质炭可能更适合于干旱地区的碱性土壤。
-
生物质炭质地轻、孔隙多,施用于土壤中既能降低土壤容重,又能增加土壤总孔隙率,还会改变土体的孔径分布情况[48−49]。同时其生物质炭强大的吸附性能可以改变土壤中水分停留时间和流动路径,从而能直接吸附水分外,还间接与其他土壤成分结合,增加水和空气循环,进而有效缓解土壤压实问题,提高土壤持水、保水能力[50]。何秀峰等[51]研究表明:施用苹果枝生物质炭后葡萄Vitis vinifera园土壤容重呈下降趋势。颜永毫等[52]研究表明:苹果枝生物质炭施于黄土高原典型土壤后其田间持水量提高幅度是施用锯末生物质炭的2.3倍。此外,在土壤中添加高量的生物质炭可使土壤颜色变暗,有利于吸光和增温,从而影响土壤温度和热量状况[53]。
-
生物质炭表面的酸性芳香碳氧化后可形成丰富的官能团(—OH, —COOH),增强土壤阳离子的吸附能力[54−55],并在水的作用下与土壤中的H+和Al3+交换,调节土壤的pH[56−57]。生物质炭还可提高果园土壤细菌、放线菌的数量,促进土壤系统中易被利用有机碳源的转化,从而增加土壤有机质质量分数[26],其中细粒径生物质炭对土壤有机质的促进作用大于粗粒径生物质炭[58]。TAGHIZADEH-TOOSI等[59]通过同位素示踪技术发现:被生物质炭吸附的铵态氮可以被植物重复利用,同时生物质炭也可以吸附氨,从而减少氮的损失,提高氮的养分利用率。有研究表明:香蕉皮生物质炭可降低土壤铵态氮和硝态氮质量分数[28]。对苹果幼苗施用由桃Prunus persica树枝和葡萄藤制备的生物质炭可减少土壤中无机磷的损失,且在温带气候条件下短期内显著降低了亚碱性土壤表层的硝酸盐浸出[60]。可见生物质炭不仅可以调节土壤pH,增加土壤有机质质量分数,还可以减少土壤养分损失,增加植物养分利用率。
-
土壤微生物作为土壤中的主要生物体,是物质转化的动力,同时可以丰富土壤有机组分。生物质炭的结构和养分不仅能够促使大部分土壤微生物的生长,还可以改变土壤理化性质,从而直接或间接影响土壤微生物数量和活性。此外,生物质炭的高芳香烃结构和表面性质很容易成为土壤微生物,如藻类、细菌、真菌和土壤动物的栖息地[61−62]。LI等[63]研究表明:土壤生物对苹果枝生物质炭的响应受土壤氮水平的影响,对氮质量分数高的土壤中β-葡萄糖苷酶、β-氨基葡萄糖苷酶和脲酶的活性有积极影响,反之对氮质量分数低的土壤有负面影响。SIAL等[28]研究表明:香蕉皮生物质炭可显著提高土壤脲酶、转化酶和碱性磷酸酶活性。
-
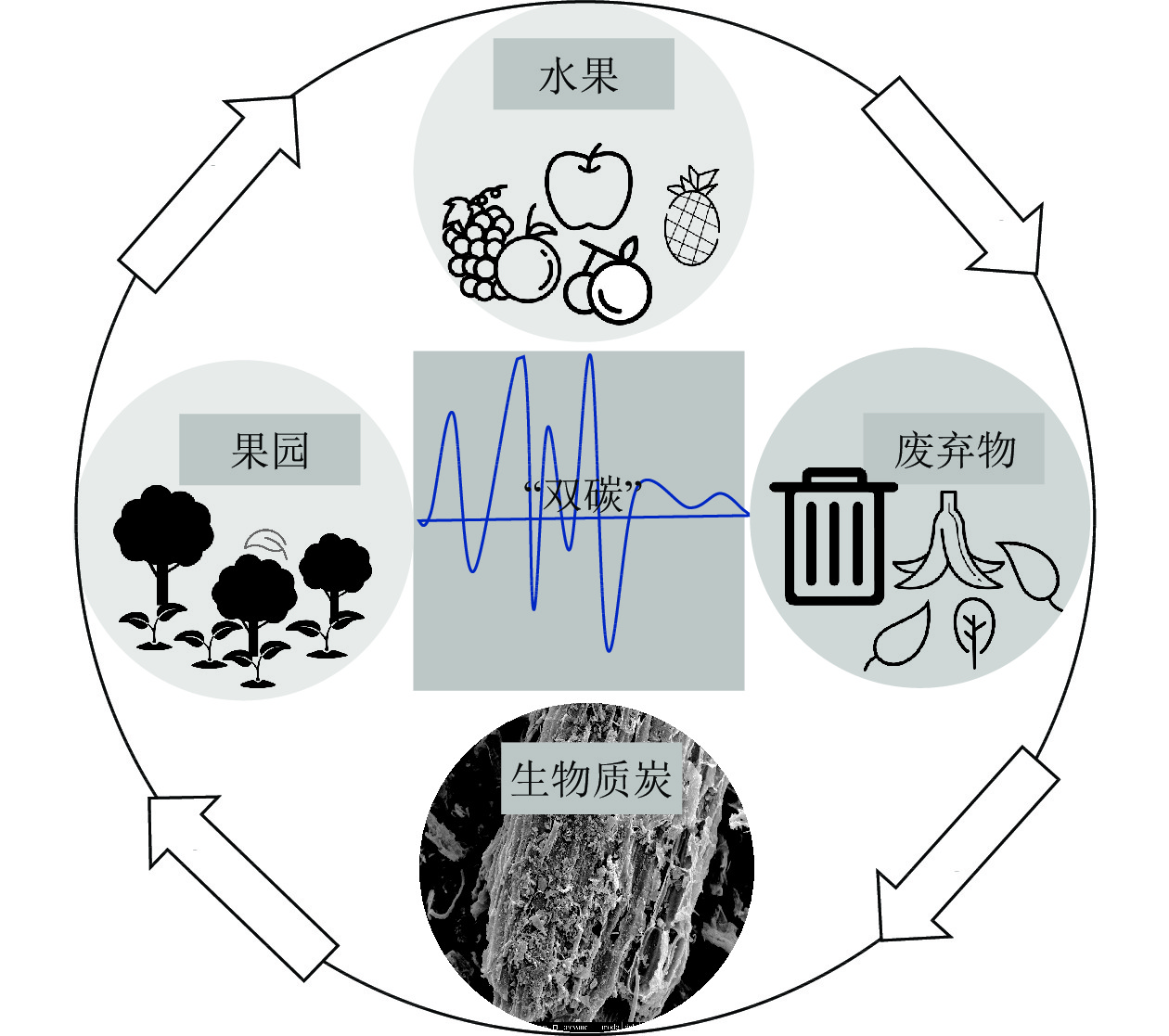
处理果园废弃物的传统方式主要有填埋、饲料、焚化、堆肥、厌氧消化、酶利用等,在整个过程中处理不当就会存在潜在风险(表2),对环境带来威胁[64]。果园废弃物炭化还田是一种有效处理果园废弃物,提高果园生产效率和调节果园生态环境的闭态循环生产方式(图1)。
表 2 废弃物处理方式和危害
Table 2. Waste disposal methods and hazards

图 1 果园废弃物生物质炭循环利用过程
Figure 1. Biochar recycling process for orchard waste
-
仁果类即梨果类。苹果和梨是中国第一、二大落叶果树,且每年树枝、树叶量较多。如果就地取材,就近利用果园废弃物制备生物质炭,并应用于当地果园中,是一种绿色环保的可持续资源利用途径。有研究表明:在干旱条件下秋子梨Pyrus ussuriensis枝条生物质炭的施用可以减轻秋子梨幼苗根系呼吸代谢功能的抑制作用,提高其对干旱胁迫的适应能力[70];梨树枝条生物质炭的施用可增加梨园土壤的pH、总碳、碳氮比[71];苹果枝生物质炭的施用可增加苹果幼苗盆栽土壤中可培养的细菌和放线菌的数量,提高细菌和真菌的比例,改变土壤微生物的群落结构[72],并为苹果园土壤提供碳源和可利用养分,促进根系与生物质炭的相互作用,调节苹果根系土壤氮转化和氮氧化物的浓度[73]。可见果园废弃物生物质炭施用于同类别果园中,改良土壤效果较为显著,但目前研究集中在盆栽试验。而秦亚旭[74]研究表明:苹果园施用以农林废弃物生物质炭为基质的生物质炭基系列专用肥后,其果实氮、磷、维生素C、可溶性固形物质量分数和糖酸比均显著提高。总的来说,仁果类果树大部分种植在中国北方,其土壤较为干旱,pH较高,对磷、钾元素的吸附性较大,具有针对性的施肥会使其生产效率显著增强。
-
核果类果树分布范围极为广泛,几乎覆盖中国全部地域。生物质炭对核果类果园土壤的影响不仅与其用量有关,而且施用年限的增加也对土壤性状有较大的影响。田间研究表明:施用生物质炭在前2 a内均增加了桃树土壤有机碳和土壤团聚体,而在3 a后才提高了土壤的酶活性,其中火炬松Pinus taeda 10%生物质炭的施用量更具有提高和改善桃树沙土的潜力[75]。不同用量及不同原料制备的生物质炭对植物生理生化性质以及果实品质和产量的影响也存在差异。如将3种生物质炭(苹果枝炭、烟草 Nicotiana tabacum秸秆炭和玉米 Zea mays秸秆炭)以2种施用量(30和60 g·kg−1)添加于樱桃 Prunus pseudocerasus幼苗的盆栽试验表明:施用生物质炭既有助于保护叶片光合色素,延缓衰老进程,又在一定程度上降低了细胞质膜透性,减轻了膜质过氧化反应的伤害,从而显著提高了樱桃幼苗叶片数、茎粗、新梢长度和株高,其中以30 g·kg−1施用量的苹果枝炭效果最为显著[76]。在果实品质和产量方面,ZHANG等[77]研究表明:施用质量分数为1.5%的生物质炭后,蓝莓 Vaccinium corymbosum果实维生素C、产量和数量分别增加了21.1%、4.82%和8.27%,而施用质量分数为3.0%生物质炭,蓝莓果实维生素C、可溶性糖分别增加了30.4%、7.24%。HARHASH等[78]研究表明:不同生物质炭用量对芒果 Mangifera indica的生长、产量和品质同样均有改善作用。
-
浆果类是成熟后果实呈浆液状而分类的,并不能真实反映果实的构造特征。葡萄是典型的浆果类水果,它对于水分要求较高,需要严格控制土壤中的水分,而果园废弃物生物质炭可以有效地提高葡萄园土壤水分含量,降低植物水分胁迫,增加光合活性[79]。有研究表明:苹果枝生物质炭施用于葡萄园后,不同程度提高了磷代谢相关菌属节细菌属Arthrobacter和钾代谢相关菌属假单胞菌属Pseudomonas的菌群数量,从而促进土壤固定态磷、钾转化为作物可直接吸收利用的有效态磷、钾[51]。也有研究表明:当养分依次转移到植物根系和叶片时,只有磷质量分数显著增加,而其他养分元素质量分数并没有显著增加[80]。可见果园废弃物生物质炭虽然对葡萄土壤中的有效养分影响显著,但施用后植物对养分元素的吸收并没有显著差异,其机制有待进一步研究。此外,在葡萄园中施用果园废弃物生物质炭4 a的研究表明:生物质炭在调节植物水分有效性中起着关键作用,而且与不施生物质炭相比,4个收获年内施用果园废弃物生物质炭的葡萄树生产力更高,但葡萄品质并没有显著差异[81]。
-
香蕉适宜在无霜冻或者少发生霜冻的温暖地区生长,主要分布于中国南方部分地区。有研究表明:施用香蕉假茎生物质炭能增加香蕉土壤中革兰氏阴性菌伯克氏菌Burkholderia、假单胞菌Pseudomonas、原小单孢菌Promicromonospora和罗思河小杆菌Rhodanobacter的丰度[82],提高香蕉苗根际微生物群落对糖类、羧酸类和氨基酸类碳源的利用能力[83]。2 a田间研究表明:生物质炭的添加促进了香蕉的生长,提高了其产量和果实品质[84]。此外,徐广平等[29]研究表明:香蕉茎叶生物质炭施用于酸化香蕉园土壤,可明显降低香蕉的黄叶率和枯萎病发病率。而在降低植物病虫害方面,果园废弃物生物质炭的合理利用有待进一步研究。
-
生物质炭是一种能促进植物生长和改善土壤特性的多功能材料[85],但不能作为植物长期和充足的营养来源,而有机肥可以通过缓慢释放可利用营养元素以及其他产物为植物提供养分[86]。有研究表明:果园废弃物生物质炭可能刺激了有机肥相对更不稳定的有机组分的微生物转化,产生了微生物副产物且吸附在土壤矿物表面,形成了有机物-矿物配合物的形式,最终增强了施用有机肥土壤中碳的稳定作用[87]。因此,将生物质炭和有机肥结合可以固定养分并长期缓慢释放,同时促进植物生长,改善土壤质量[88],两者可以起到互补的作用。如苹果枝生物质炭和牛粪、秸秆等有机物料混合施用后,土壤矿化氮、土壤酶活性及微生物数量均高于单独施用有机物料,且增强了土壤对氮的固持能力[89]。2 a年田间研究表明:生物质炭和有机肥结合,存在某种类型的协同效应[90],与单独施用生物质炭对比,有机肥为生物质炭提供了大量的养分元素,不仅改良了土壤,还显著提高了果实产量和品质。有研究表明:刺梨Hibiscus sabdariffa皮制备的生物质炭配施有机肥可通过改变土壤性质,提高土壤微生物活性和养分利用率,显著提高了作物产量和品质[91]。
EYLES等[92]进行了4 a的田间研究表明:与生物质炭结合有机肥相比,单独施用生物质炭对果树生长的积极影响仅在试验的最后1 a才有明显的体现。这表明生物质炭对土壤的改良作用可能需要进行更长时间尺度来研究。且把苹果枝生物质炭结合无机肥施用于黄土高原的苹果园,虽然苹果枝生物质炭有助于在土壤中储存碳,但在该研究中并没有显示出减轻温室气体排放的潜力,反而增加了二氧化碳和一氧化二氮的累积排放量[93]。值得关注的是生物质炭减缓气候变化的影响不仅取决于其材料特性,还取决于其生产和应用方式[94]。与施用其他生物质炭一样,需要长期探究果园废弃物生物质炭配施有机肥时,土壤中果园废弃物生物质炭的合理应用方式和演变机制,以稳定果园生产效率和生态环境。
-
果园废弃物炭化还田,不仅解决了果园废弃物的处理问题,还可以改良果园土壤,缓解养分释放,为果树提供养分,促进果树生长,并进一步提高了果实产量和品质。但还有很多相关问题亟待研究,如果园废弃物生物质炭原料选择性相对较少;果园施用的废弃物生物质炭专一性相对较弱;施用废弃物生物质炭也有可能对果园产量、果实品质以及果园生态环境产生负面影响。针对以上问题,今后应重点关注以下研究内容:①进一步分类果园废弃物原料,制备不同种类生物质炭以满足差异化需求,如不同果树种类、不同种植区域及果树不同生长阶段。②探索不同果园废弃物生物质炭施用于同类别果园的效果研究。生物质炭会保留原材料本身的一些特性,植物所需元素的种类和份量应该与同原料生物质炭的性质更加接近。应开展不同类别果园废弃物生物质炭施用于同类别果园的研究,探索适宜的果园废弃物生物质炭施用方案。③结合有机肥制备果园专用生物质炭基有机肥。生物质炭基有机肥是指以生物质炭为基质,添加某种或几种有机物料,通过一定的加工工艺制成的肥料,不仅可以长期缓慢释放养分,而且对于果园生产具有较高的针对性、科学性、全面性,可实现果园养分多样化,提高肥料利用效率。④关注果园废弃物生物质炭对果园生态环境的影响。生物质炭在土壤中缓慢降解,且理化性质和生态环境效益随时间的变化而不同。应研究长期添加生物质炭对果园土壤的影响及机制,特别要评估其对果园生态环境影响的负面机制,制定相应的改进方案。
Research progress on application effect of orchard waste biochar in orchard
-
摘要: 处理果园废弃物是果园生产中的重要环节,传统方式主要有填埋、饲料、焚化、堆肥、厌氧消化、酶利用等。果园废弃物生物质炭是以果园生产中的废弃物为原料,通过热解技术转化后获得生物质炭。将生物质炭返还于果园,这样不仅改良了果园土壤,还保障和提高了果实产量和品质。果园废弃物在果园中的合理应用对果园生产效率和生态环境具有重要的稳定作用,开展果园废弃物生物质炭的研究对其在果园中的合理应用和演变机质具有重要的科学意义和应用价值。目前,果园废弃物生物质炭的研究集中于不同类别果园废弃物生物质炭的制备原理和性质,改良土壤的效果以及有机肥配施对果园生产的影响。未来应重点关注:①进一步分类果园废弃物,制备更多满足差异化需求的生物质炭;②比较不同类别果园废弃物生物质炭施用的生产效益;③制备果园专用生物质炭基有机肥;④探讨果园废弃物生物质炭对果园生态环境的影响。图1表2参94Abstract: Treatment of orchard waste is an important link in orchard production. The carbonization of orchard waste is a green, environmentally friendly and low-carbon mode of production, which can improve orchard soil, as well as yield and quality of fruits. Therefore, it’s of great scientific significance and application value to carry out research on orchard waste biochar for its rational application and evolution in orchard. The current research on biochar from orchard wastes was reviewed, which mainly focused on the preparation principle and properties of biochar from different types of orchard wastes and the effects on soil improvement, and the impact of its application combined with organic fertilizer on orchard production. The future research directions are proposed: (1) To further classify the orchard wastes to prepare more biochar to meet the differentiated needs. (2) To compare the production benefits of different types of orchard waste biochar application. (3) To prepare biochar-based organic fertilizer for orchard. (4) To explore the effect of orchard waste biochar on orchard environment and ecology. [Ch, 1 fig. 2 tab. 94 ref.]
-
Key words:
- orchard /
- orchard waste /
- biochar
-
表 1 果园废弃物生物质炭制作条件和性质
Table 1. Methods and properties of orchard waste biochar production
原材料 热解温度/℃ 热解时间/h 有机碳/(g·kg−1) 全碳/% 全氮/% 全磷/% 全钾/% 炭产量/% 橙子皮 350 3 65.58 2.26 0.66 0.05 32.50 100~600 1 10.85 1.56 300 2 80.72 2.55 350 2 8.58 0.17 12.20 18.62 柑橘枝 500 2 80.60 0.87 0.30 1.08 香蕉花梗 300 1 37.79 66.00 500 1 41.86 40.00 香蕉皮 350 3 5.54 1.90 1.26 0.42 40.00 香蕉茎叶 500 2 462.50 1.36 0.54 4.27 苹果树枝 450 1 82.15 1.28 苹果树皮 450 1 68.96 2.93 苹果枝 700 2 74.21 1.29 32.60 300 2 62.20 1.69 47.94 600 2 80.10 1.28 28.48 椰子壳 400 47.92 1.34 43.33 600 58.43 1.02 36.00 800 67.31 0.87 34.00 1000 74.08 0.50 31.00 海枣树废弃物 300 57.99 0.54 49.97 400 66.87 0.45 36.54 500 72.30 0.42 32.38 600 72.89 0.39 30.88 700 73.42 0.35 28.84 800 74.63 0.31 27.40 原材料 灰分/% 挥发物/% 水分/% pH 比表面积/(m2·g−1) 碳氮比 文献 橙子皮 7.80 21.40 6.20 9.97 SIAL等[22] 5.50 70.00 13.00 9.43 STELLA等[23] 352.50 31.65 ADENIYI等[24] 9.10 10.02 OGUNKUNLE等[25] 柑橘枝 9.23 4.85 92.64 逄玉万等[26] 香蕉花梗 24.84 33.69 26.28 8.10 KARIM等[27] 35.48 24.37 21.26 10.20 香蕉皮 9.80 26.50 5.20 10.88 SIAL等[28] 香蕉茎叶 19.05 10.33 80.51 340.07 徐广平等[29] 苹果树枝 2.01 9.47 40.38 64.18 石钧元等[30] 苹果树皮 12.00 12.36 55.14 23.54 苹果枝 8.90 10.00 57.53 曹辉等[31] 6.72 60.77 2.39 36.80 ZHAO等[32] 9.40 14.86 108.59 62.58 椰子壳 26.27 30.56 8.50 39.57 35.76 SUMAN等[33] 21.99 20.82 9.64 120.73 57.28 17.32 5.33 10.87 256.60 77.37 12.03 4.72 11.02 590.41 148.46 海枣树废弃物 14.42 40.08 3.29 107.39 USMAN等[34] 16.34 20.25 3.13 148.60 19.68 9.31 2.96 172.14 20.71 6.85 2.25 186.90 21.05 5.47 2.12 209.77 21.39 3.91 2.09 240.74 说明:橙子Citrus sinensis;柑橘C. reticulata;香蕉Musa nana;苹果Malus pumila;椰子Cocos nucifera;海枣树 Phoenix dactylifera。表中百分数均为质量分数  下载: 导出CSV
下载: 导出CSV
-
[1] FAWZY S, OSMAN A I, YANG Haiping, et al. Industrial biochar systems for atmospheric carbon removal: a review [J]. Environmental Chemistry Letters, 2021, 19(4): 3023 − 3055. [2] FAWZY S, OSMAN A I, DORAN J, et al. Strategies for mitigation of climate change: a review [J]. Environmental Chemistry Letters, 2020, 18(6): 2069 − 2094. [3] HUANG Xiaobing, GAO Shiqi. Temporal characteristics and influencing factors of agricultural carbon emission in Jiangxi Province of China [J/OL]. Environmental Research Communications, 2022, 4(4): 045006[2022-07-15]. doi: 10.1088/2515-7620/ac6380. [4] GABHANE J W, BHANGE V P, PATIL P D, et al. Recent trends in biochar production methods and its application as a soil health conditioner: a review [J/OL]. Sn Applied Sciences, 2020, 2(7): 1307[2022-07-15]. doi: 10.1007/s42452-020-3121-5. [5] SHARMA S, RANA V S, RANA N, et al. Biochar from fruit crops waste and its potential impact on fruit crops [J/OL]. Scientia Horticulturae, 2022, 299: 111052[2022-07-15]. doi: org/10.1016/j. scienta. 2022.111052. [6] DING Yang, LIU Yunguo, LIU Shaobo, et al. Biochar to improve soil fertility: a review [J/OL]. Agronomy for Sustainable Development, 2016, 36(2): 36[2022-07-15]. doi: 10.1007/s13593-016-0372-z. [7] EL-NAGGAR A, LEE S S, RINKLEBE J, et al. Biochar application to low fertility soils: a review of current status, and future prospects [J]. Geoderma, 2019, 337: 536 − 554. [8] LEHMANN J, GAUNT J, RONDON M. Bio-char sequestration in terrestrial ecosystems: a review [J]. Mitigation and Adaptation Strategies for Global Change, 2006, 11(2): 403 − 427. [9] WOOLF D, AMONETTE J E, STREET-PERROTT F A, et al. Sustainable biochar to mitigate global climate change [J/OL]. Nature Communications, 2010, 1(1): 56[2022-07-15]. doi: 10.1038/ncomms1053. [10] SAGAR N A, PAREEK S, SHARMA S, et al. Fruit and vegetable waste: bioactive compounds, their extraction, and possible utilization [J]. Comprehensive Reviews in Food Science and Food Safety, 2018, 17(3): 512 − 531. [11] MAGAMA P, CHIYANZU I, MULOPO J. A systematic review of sustainable fruit and vegetable waste recycling alternatives and possibilities for anaerobic biorefinery [J/OL]. Bioresource Technology Reports, 2022, 18: 101031[2022-07-15]. doi: 10.1016/j.biteb.2022.101031. [12] DU Chenyu, ABDULLAH J J, GREETHAM D, et al. Valorization of food waste into biofertiliser and its field application [J]. Journal of Cleaner Production, 2018, 187: 273 − 284. [13] GANESH K S, SRIDHAR A, VISHALI S. Utilization of fruit and vegetable waste to produce value-added products: conventional utilization and emerging opportunities-a review [J/OL]. Chemosphere, 2022, 287: 132221[2022-07-15]. doi: 10.1016/j.chemosphere.2021.132221. [14] LENG Lijian, HUANG Huajun. An overview of the effect of pyrolysis process parameters on biochar stability [J]. Bioresource Technology, 2018, 270: 627 − 642. [15] PATRA B R, MUKHERJEE A, NANDA S, et al. Biochar production, activation and adsorptive applications: a review [J]. Environmental Chemistry Letters, 2021, 19(3): 2237 − 2259. [16] HUANG He, REDDY N G, HUANG Xilong, et al. Effects of pyrolysis temperature, feedstock type and compaction on water retention of biochar amended soil [J]. Scientific Reports, 2021, 11(1): 1 − 19. [17] FOONG S Y, LIEW R K, YANG Yafeng, et al. Valorization of biomass waste to engineered activated biochar by microwave pyrolysis: progress, challenges, and future directions [J/OL]. Chemical Engineering Journal, 2020, 389: 124401[2022-07-15]. doi: 10.1016/j.cej.2020.124401. [18] NANDA S, MOHANTY P, PANT K K, et al. Characterization of north American lignocellulosic biomass and biochars in terms of their candidacy for alternate renewable fuels [J]. BioEnergy Research, 2013, 6(2): 663 − 677. [19] CHEN Dengyu, CEN Kehui, ZHUANG Xiaozhuang, et al. Insight into biomass pyrolysis mechanism based on cellulose, hemicellulose, and lignin: evolution of volatiles and kinetics, elucidation of reaction pathways, and characterization of gas, biochar and bio-oil [J/OL]. Combustion and Flame, 2022, 242: 112142[2022-07-15]. doi: 10.1016/j.combustflame.2022.112142. [20] WEBER K, QUICKER P. Properties of biochar [J]. Fuel, 2018, 217: 240 − 261. [21] YANG Haiping, YAN Rong, CHEN Hanping, et al. Characteristics of hemicellulose, cellulose and lignin pyrolysis [J]. Fuel, 2007, 86(12/13): 1781 − 1788. [22] SIAL T A, LAN Zhilong, KHAN M N, et al. Evaluation of orange peel waste and its biochar on greenhouse gas emissions and soil biochemical properties within a loess soil [J]. Waste Management, 2019, 87: 125 − 134. [23] STELLA MARY G, SUGUMARAN P, NIVEDITHA S, et al. Production, characterization and evaluation of biochar from pod (Pisum sativum), leaf (Brassica oleracea) and peel (Citrus sinensis) wastes [J]. International Journal of Recycling of Organic Waste in Agriculture, 2016, 5(1): 43 − 53. [24] ADENIYI A G, IGHALO J O, ONIFADE D V. Biochar from the thermochemical conversion of orange (Citrus sinensis) peel and albedo: product quality and potential applications [J]. Chemistry Africa, 2020, 3(2): 439 − 448. [25] OGUNKUNLE C O, VARUN M, OGUNDELE I G, et al. Citrus epicarp-derived biochar reduced Cd uptake and ameliorates oxidative stress in young Abelmoschus esculentus (L. ) Moench (okra) under low Cd stress [J]. Bulletin of Environmental Contamination and Toxicology, 2018, 100(6): 827 − 833. [26] 逄玉万, 唐拴虎, 林焕嘉, 等. 酸性硫酸盐土壤对柑桔枝生物炭改良的响应[J]. 中国农学通报, 2018, 34(23): 86 − 91. PANG Yuwan, TANG Shuanhu, LIN Huanjia, et al. Response of acid sulfate soil to amelioration by citrus branch biochar [J]. Chinese Agricultural Science Bulletin, 2018, 34(23): 86 − 91. [27] KARIM A A, KUMAR M K, MOHAPATRA S, et al. Banana peduncle biochar: characteristics and adsorption of hexavalent chromium from aqueous solution [J]. International Research Journal of Pure and Applied Chemistry, 2015, 7(1): 1 − 10. [28] SIAL T A, KHAN M N, LAN Zhilong, et al. Contrasting effects of banana peels waste and its biochar on greenhouse gas emissions and soil biochemical properties [J]. Process Safety and Environmental Protection, 2019, 122: 366 − 377. [29] 徐广平, 滕秋梅, 沈育伊, 等. 香蕉茎叶生物炭对香蕉枯萎病防控效果及土壤性状的影响[J]. 生态环境学报, 2020, 29(12): 2373 − 2384. XU Guangping, TENG Qiumei, SHEN Yuyi, et al. Effects of banana stems-leaves biochar on soil properties and control of banana fusarium wilt [J]. Ecology and Environmental Sciences, 2020, 29(12): 2373 − 2384. [30] 石钧元, 荀咪, 崔迎宾, 等. 炭化苹果枝皮和木材对水中硝态氮和铵态氮的差异吸附[J]. 水土保持学报, 2020, 34(2): 253 − 260. SHI Junyuan, XUN Mi, CUI Yingbin, et al. Differential sorption of nitrate and ammonium nitrogen in water by charred apple branch bark and wood [J]. Journal of Soil and Water Conservation, 2020, 34(2): 253 − 260. [31] 曹辉, 李燕歌, 周春然, 等. 炭化苹果枝对苹果根区土壤细菌和真菌多样性的影响[J]. 中国农业科学, 2016, 49(17): 3413 − 3424. CAO Hui, LI Yan’ge, ZHOU Chunran, et al. Effect of carbonized apple branches on bacterial and fungal diversities in apple root-zone soil [J]. Scientia Agricultura Sinica, 2016, 49(17): 3413 − 3424. [32] ZHAO Shixiang, TA Na, WANG Xudong. Effect of temperature on the structural and physicochemical properties of biochar with apple tree branches as feedstock material [J/OL]. Energies, 2017, 10(9): 1293[2022-07-15]. doi: 10.3390/en10091293. [33] SUMAN S, GAUTAM S. Pyrolysis of coconut husk biomass: analysis of its biochar properties [J]. Energy Sources,Part A:Recovery,Utilization,and Environmental Effects, 2017, 39(8): 761 − 767. [34] USMAN A R A, ABDULJABBAR A, VITHANAGE M, et al. Biochar production from date palm waste: charring temperature induced changes in composition and surface chemistry [J]. Journal of Analytical and Applied Pyrolysis, 2015, 115: 392 − 400. [35] YOGALAKSHMI K N, POORNIMA D T, SIVASHANMUGAM P, et al. Lignocellulosic biomass-based pyrolysis: a comprehensive review [J/OL]. Chemosphere, 2022, 286: 131824[2022-07-15]. doi: 10.1016/j.chemosphere.2021.131824. [36] SAIT H H, HUSSAIN A, SALEMA A A, et al. Pyrolysis and combustion kinetics of date palm biomass using thermogravimetric analysis [J]. Bioresource Technology, 2012, 118: 382 − 389. [37] CHAIWONG K, KIATSIRIROAT T, VORAYOS N, et al. Study of bio-oil and bio-char production from algae by slow pyrolysis [J]. Biomass and Bioenergy, 2013, 56: 600 − 606. [38] FERNANDES E R K, MARANGONI C, SOUZA O, et al. Thermochemical characterization of banana leaves as a potential energy source [J]. Energy Conversion and Management, 2013, 75: 603 − 608. [39] MAIA B G D O, SOUZA O, MARANGONI C, et al. Production and characterization of fuel briquettes from banana leaves waste [J]. Chemical Engineering Transactions, 2014, 37: 439 − 444. [40] ABDULLAH N, SULAIMAN F, MISKAM M A, et al. Characterization of banana (Musa spp. ) pseudo-stem and fruit-bunch-stem as a potential renewable energy resource [J]. International Journal of Biological,Veterinary,Agricultural and Food Engineering, 2014, 8(8): 712 − 716. [41] BISWAS B, RAWEL S, KUMAR J, et al. Slow pyrolysis of prot, alkali and dealkaline lignins for production of chemicals [J]. Bioresource Technology, 2016, 213: 319 − 326. [42] KABENGE I, OMULO G, BANADDA N, et al. Characterization of banana peels wastes as potential slow pyrolysis feedstock [J/OL]. Journal of Sustainable Development, 2018, 11(2): 14[2022-07-15]. doi: 10.5539/jsd.v1n2p14. [43] WAN Jiang, LIU Lin, AYUB K S, et al. Characterization and adsorption performance of biochars derived from three key biomass constituents [J/OL]. Fuel, 2020, 269: 117142[2022-07-15]. doi: 10.1016/j.fuel.2020.117142. [44] JANKOVIĆ B, MANIĆ N, DODEVSKI V, et al. Physico-chemical characterization of carbonized apricot kernel shell as precursor for activated carbon preparation in clean technology utilization [J/OL]. Journal of Cleaner Production, 2019, 236: 117614[2022-07-15]. doi: 10.1016/j.jclepro.2019.117614. [45] ZHANG Zhongqing, ZHOU Chenhui, YANG Jingmin, et al. Preparation and characterization of apricot kernel shell biochar and its adsorption mechanism for atrazine [J/OL]. Sustainability, 2022, 14(7): 4082[2022-07-15]. doi: 10.3390/su14074082. [46] KARIM A A, KUMAR M, SINGH S K, et al. Potassium enriched biochar production by thermal plasma processing of banana peduncle for soil application [J]. Journal of Analytical and Applied Pyrolysis, 2017, 123: 165 − 172. [47] PARK J H, OK Y S, KIM S H, et al. Characteristics of biochars derived from fruit tree pruning wastes and their effects on lead adsorption [J]. Journal of the Korean Society for Applied Biological Chemistry, 2015, 58(5): 751 − 760. [48] 李金文, 顾凯, 唐朝生, 等. 生物炭对土体物理化学性质影响的研究进展[J]. 浙江大学学报(工学版), 2018, 52(1): 192 − 206. LI Jinwen, GU Kai, TANG Chaosheng, et al. Advances in effects of biochar on physical and chemical properties of soil [J]. Journal of Zhejiang University (Engineering Science), 2018, 52(1): 192 − 206. [49] 郭茜, 陆扣萍, 胡国涛, 等. 死猪炭和竹炭对菜地土壤理化性质和蔬菜产量的影响[J]. 浙江农林大学学报, 2017, 34(2): 244 − 252. GUO Xi, LU Kouping, HU Guotao, et al. Greenhouse soil properties and vegetable yield with dead pig and bamboo biochars [J]. Journal of Zhejiang A&F University, 2017, 34(2): 244 − 252. [50] 王瑞峰, 赵立欣, 沈玉君, 等. 生物炭制备及其对土壤理化性质影响的研究进展[J]. 中国农业科技导报, 2015, 17(2): 126 − 133. WANG Ruifeng, ZHAO Lixin, SHEN Yujun, et al. Research progress on preparing biochar and its effect on soil physio-chemical properties [J]. Journal of Agricultural Science and Technology, 2015, 17(2): 126 − 133. [51] 何秀峰, 赵丰云, 于坤, 等. 生物炭对葡萄幼苗根际土壤养分、酶活性及微生物多样性的影响[J]. 中国土壤与肥料, 2020(6): 19 − 26. HE Xiufeng, ZHAO Fengyun, YU Kun, et al. Effect of biochar on nutrient, enzyme activities and microbial diversity of rhizosphere soil of grape seedlings [J]. Soil and Fertilizer Sciences in China, 2020(6): 19 − 26. [52] 颜永毫, 郑纪勇, 张兴昌, 等. 生物炭添加对黄土高原典型土壤田间持水量的影响[J]. 水土保持学报, 2013, 27(4): 120 − 124, 190. YAN Yonghao, ZHENG Jiyong, ZHANG Xingchang, et al. Impact of biochar addition into typical soil on field capacity in loess plateau [J]. Journal of Soil and Water Conservation, 2013, 27(4): 120 − 124, 190. [53] ZHANG Yafu, WANG Jinman, FENG Yu. The effects of biochar addition on soil physicochemical properties: a review [J/OL]. Catena, 2021, 202: 105284[2022-07-15]. doi: 10.1016/j.catena.2021.105284. [54] ATKINSON C J, FITZGERALD J D, HIPPS N A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: a review [J]. Plant and Soil, 2010, 337(1/2): 1 − 18. [55] GASKIN J W, STEINER C, HARRIS K, et al. Effect of low-temperature pyrolysis conditions on biochar for agricultural use [J]. Transactions of the ASABE, 2008, 51(6): 2061 − 2069. [56] van ZWIETEN L, KIMBER S, MORRIS S, et al. Effects of biochar from slow pyrolysis of papermill waste on agronomic performance and soil fertility [J]. Plant and Soil, 2010, 327(1/2): 235 − 246. [57] 包骏瑶, 赵颖志, 严淑娴, 等. 不同农林废弃物生物质炭对雷竹林酸化土壤的改良效果[J]. 浙江农林大学学报, 2018, 35(1): 43 − 50. BAO Junyao, ZHAO Yingzhi, YAN Shuxian, et al. Soil amelioration with biochars pyrolyzed from different feedstocks of an acidic bamboo (Phyllostachys violascens) plantation [J]. Journal of Zhejiang A&F University, 2018, 35(1): 43 − 50. [58] 宿贤超, 胡杨勇, 赵薇, 等. 添加竹炭对土壤化学性质和重金属有效性及水稻生长的影响[J]. 浙江农业学报, 2014, 26(2): 439 − 443. SU Xianchao, HU Yangyong, ZHAO Wei, et al. Effects of addition of bamboo charcoal on soil chemical properties, heavy metal availability, and rice growth [J]. Acta Agriculturae Zhejiangensis, 2014, 26(2): 439 − 443. [59] TAGHIZADEH-TOOSI A, CLOUGH T J, SHERLOCK R R, et al. Biochar adsorbed ammonia is bioavailable [J]. Plant and Soil, 2012, 350(1/2): 57 − 69. [60] VENTURA M, SORRENTI G, PANZACCHI P, et al. Biochar reduces short-term nitrate leaching from a horizon in an apple orchard [J]. Journal of Environmental Quality, 2013, 42(1): 76 − 82. [61] KNICKER H. How does fire affect the nature and stability of soil organic nitrogen and carbon? a review [J]. Biogeochemistry, 2007, 85(1): 91 − 118. [62] SUN Wei, GU Jie, WANG Xiaojuan, et al. Impacts of biochar on the environmental risk of antibiotic resistance genes and mobile genetic elements during anaerobic digestion of cattle farm wastewater [J]. Bioresource Technology, 2018, 256: 342 − 349. [63] LI Shuailin, LIANG Chutao, SHANGGUAN Zhouping. Effects of apple branch biochar on soil C mineralization and nutrient cycling under two levels of N [J]. Science of the Total Environment, 2017, 607/608: 109 − 119. [64] SRIDHAR A, KAPOOR A, KUMAR P S, et al. Conversion of food waste to energy: a focus on sustainability and life cycle assessment[J/OL]. Fuel, 2021, 302: 121069[2022-07-15]. doi: 10.1016/j.fuel.2021.121069. [65] GARCÍA A J, ESTEBAN M B, MÁRQUEZ M C, et al. Biodegradable municipal solid waste: characterization pand potential use as animal feedstuffs [J]. Waste Management, 2005, 25(8): 780 − 787. [66] DENG Guifang, SHEN Chen, XU Xiangrong, et al. Potential of fruit wastes as natural resources of bioactive compounds [J]. International Journal of Molecular Sciences, 2012, 13(7): 8308 − 8323. [67] SAYARA T, BASHEER-SALIMIA R, HAWAMDE F, et al. Recycling of organic wastes through composting: process performance and compost application in agriculture [J/OL]. Agronomy, 2020, 10(11): 1838[2022-07-15]. doi: 10.3390/agronomy10111838. [68] CHATTERJEE B, MAZUMDER D. New approach of characterizing fruit and vegetable waste (FVW) to ascertain its biological stabilization via two-stage anaerobic digestion (AD) [J/OL]. Biomass and Bioenergy, 2020, 139: 105594[2022-07-15]. doi: 10.1016/j.biombioe.2020.105594. [69] KAUR S, KAUR H P, PRASAD B, et al. Production and optimization of pectinase by Bacillus sp. isolated from vegetable waste soil [J]. Indo American Journal of Pharmaceutical Research, 2016, 6(1): 4185 − 4190. [70] 杜国栋, 刘志琨, 赵玲, 等. 生物炭减缓干旱胁迫对秋子梨根系呼吸生理功能的影响[J]. 果树学报, 2016, 33(增刊1): 90 − 97. DU Guodong, LIU Zhikun, ZHAO Ling, et al. Biochar alleviates effects of drought stress on root respiration in Pyrus ussuriensis [J]. Journal of Fruit Science, 2016, 33(suppl 1): 90 − 97. [71] OO A Z, SUDO S, WIN K T, et al. Influence of pruning waste biochar and oyster shell on N2O and CO2 emissions from Japanese pear orchard soil [J/OL]. Heliyon, 2018, 4(3): e00568[2022-07-15]. doi: 10.1016/j.heliyon.2018.e00568. [72] LIU Yinghao, MA Zhiting, CHEN Ran, et al. Biochar promotes the growth of apple seedlings by adsorbing phloridzin [J/OL]. Scientia Horticulturae, 2022, 303: 111187[2022-07-15]. doi: 10.1016/j.scienta.2022.111187. [73] CAO Hui, JIA Mingfang, XUN Mi, et al. Nitrogen transformation and microbial community structure varied in apple rhizosphere and rhizoplane soils under biochar amendment [J]. Journal of Soils and Sediments, 2021, 21(2): 853 − 868. [74] 秦亚旭. 生物质炭基专用肥对苹果产量品质及土壤肥力的影响[D]. 杨凌: 西北农林科技大学, 2020. QIN Yaxu. Effects of Biochar-based Special Compound Fertilizer on Yield and Quality of Apple and Soil Fertility [D]. Yangling: Northwest A&F University, 2020. [75] FRENE J P, FRAZIER M, RUTTO E, et al. Early response of organic matter dynamics to pine-biochar in sandy soil under peach trees [J/OL]. Agrosystems, Geosciences & Environment, 2020, 3(1): e20094[2022-07-15]. doi: 10.1002/agg2.20094. [76] 王健宁, 文晓鹏, 洪怡, 等. 生物炭对玛瑙红樱桃苗期生理生化特征的影响[J]. 华中农业大学学报, 2019, 38(3): 19 − 24. WANG Jianning, WEN Xiaopeng, HONG Yi, et al. Effects of biochar on physiological and biochemical characteristics of Prunus pseudocerasus ‘Manaohong’ seedling [J]. Journal of Huazhong Agricultural University, 2019, 38(3): 19 − 24. [77] ZHANG Yuchan, WANG Xiao, LIU Bingjie, et al. Comparative study of individual and co-application of biochar and wood vinegar on blueberry fruit yield and nutritional quality [J/OL]. Chemosphere, 2020, 246: 125699[2022-07-15]. doi: 10.1016/j.chemosphere.2019.125699. [78] HARHASH M M, AHAMED M M M, MOSA W F A. Mango performance as affected by the soil application of zeolite and biochar under water salinity stresses [J/OL]. Environmental Science and Pollution Research, 2022[2022-07-15]. doi: 10.1007/s11356-022-21503-4. [79] BARONTI S, VACCARI F P, MIGLIETTA F, et al. Impact of biochar application on plant water relations in Vitis vinifera (L. ) [J]. European Journal of Agronomy, 2014, 53: 38 − 44. [80] CHANG Yuru, ROSSI L, ZOTARELLI L, et al. Greenhouse evaluation of pinewood biochar effects on nutrient status and physiological performance in muscadine grape (Vitis rotundifolia L. ) [J]. HortScience, 2021, 56(2): 277 − 285. [81] GENESIO L, MIGLIETTA F, BARONTI S, et al. Biochar increases vineyard productivity without affecting grape quality: Results from a four years field experiment in Tuscany [J]. Agriculture,Ecosystems &Environment, 2015, 201: 20 − 25. [82] 王明元, 侯式贞, 董涛, 等. 香蕉假茎生物炭对根际土壤细菌丰度和群落结构的影响[J]. 微生物学报, 2019, 59(7): 1363 − 1372. WANG Mingyuan, HOU Shizhen, DONG Tao, et al. Effects of banana pseudostem biochar on bacterial abundance and community structure in rhizosphere soil [J]. Acta Microbiologica Sinica, 2019, 59(7): 1363 − 1372. [83] 李航, 董涛, 王明元. 生物炭对香蕉苗根际土壤微生物群落与代谢活性的影响[J]. 微生物学杂志, 2016, 36(1): 42 − 48. LI Hang, DONG Tao, WANG Mingyuan. Effects of biochar on microbial communities and metabolic activity in rhizospheric soil of banana seedlings [J]. Journal of Microbiology, 2016, 36(1): 42 − 48. [84] ABO-OGIALA A. Impact of biochar on vegetative parameters, leaf mineral content, yield and fruit quality of grande naine banana in saline-sodic soil [J]. Egyptian Journal of Horticulture, 2018, 45(2): 315 − 330. [85] SATTAR A, SHER A, IJAZ M, et al. Interactive effect of biochar and silicon on improving morpho-physiological and biochemical attributes of maize by reducing drought hazards [J]. Journal of Soil Science and Plant Nutrition, 2020, 20(4): 1819 − 1826. [86] MÖLLER K, SCHULTHEIΒ U. Chemical characterization of commercial organic fertilizers [J]. Archives of Agronomy and Soil Science, 2015, 61(7/9): 989 − 1012. [87] PLAZA C, GIANNETTA B, FERNÁNDEZ J M, et al. Response of different soil organic matter pools to biochar and organic fertilizers [J]. Agriculture,Ecosystems &Environment, 2016, 225: 150 − 159. [88] BOLAN N, HOANG S A, BEIYUAN J, et al. Multifunctional applications of biochar beyond carbon storage [J]. International Materials Reviews, 2022, 67(2): 150 − 200. [89] 王璐, 朱占玲, 刘照霞, 等. 多种有机物料混施对苹果幼苗生长、氮素利用及土壤特性的影响[J]. 水土保持学报, 2021, 35(5): 362 − 368. WANG Lu, ZHU Zhanling, LIU Zhaoxia, et al. Effects of mixtures of diffierent organic materials on apples seeding growth, nitrogen utilization and soil properties [J]. Journal of Soil and Water Conservation, 2021, 35(5): 362 − 368. [90] SÁNCHEZ-GARCÍA M, SÁNCHEZ-MONEDERO M A, ROIG A, et al. Compost vs biochar amendment: a two-year field study evaluating soil C build-up and N dynamics in an organically managed olive crop [J]. Plant and Soil, 2016, 408(1/2): 1 − 14. [91] AL-SAYED H M, ALI A M, MOHAMED M A, et al. Combined effect of prickly pear waste biochar and azolla on soil fertility, growth, and yield of roselle (Hibiscus sabdariffa L. ) plants [J]. Journal of Soil Science and Plant Nutrition, 2022, 22: 3541 − 3552. [92] EYLES A, BOUND S A, OLIVER G, et al. Impact of biochar amendment on the growth, physiology and fruit of a young commercial apple orchard [J]. Trees, 2015, 29(6): 1817 − 1826. [93] HAN Jiale, ZHANG Afeng, KANG Yanhong, et al. Biochar promotes soil organic carbon sequestration and reduces net global warming potential in apple orchard: a two-year study in the Loess Plateau of China [J/OL]. Science of the Total Environment, 2022, 803: 150035[2022-07-15]. doi: 10.1016/j.scitotenv.2021.150035. [94] LEHMANN J, COWIE A, MASIELLO C A, et al. Biochar in climate change mitigation [J]. Nature Geoscience, 2021, 14(12): 883 − 895. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220535
 点击查看大图
点击查看大图
图(1) / 表(2)
计量
- 文章访问数: 2269
- HTML全文浏览量: 393
- PDF下载量: 105
- 被引次数: 0
