






-
MYB蛋白质家族成员数量庞大,功能多样,存在于绝大多数真核生物中。大多数MYB蛋白作为转录因子发挥作用,MYB结合域有结合DNA的能力[1−2],在植物生长发育、次生代谢及抗逆胁迫等方面具有重要作用[3]。R2R3-MYB作为家族中成员最多的一类,对植物生长发育意义重大[4−7]。
MYB基因家族根据MYB结构域的数量和类型分为4类:1R-MYB (或MYB相关)、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3-MYB)和4R-MYB[8]。2R-MYB类是植物特有的,并且是最大的亚家族,已在许多植物中进行了全基因组注释。R2R3-MYB家族成员数量各不相同,拟南芥Arabidopsis thaliana[9]有126个,水稻Oryza sativa[10]有102个,葡萄Vitis vinifera[11]有108个,甜橙Citrus sinensis[12]有100个,苹果Malus domestica[13]有222个,毛果杨Populus trichocarpa[14]有192个,玉米Zea mays[15]有200多个。LI等[16]在黄瓜Cucumis sativus中仅鉴定出55个R2R3-MYB基因,而黄瓜基因组V2版本中鉴定出69个R2R3-MYB基因[17]。通过最新版黄瓜基因组数据库可知:黄瓜基因组V3版本基因组数量为24 317个,较V2版本(23 248个)增多。鉴于此,本研究对黄瓜基因组数据库V3版本中的R2R3-MYB亚家族进行鉴定,分析黄瓜CsMYB基因家族成员,为研究黄瓜CsMYB家族功能提供参考。
-
全基因组序列来源于黄瓜基因组数据库(
http://www.cucurbitgenomics.org/organism/20 )。序列中的MYB结合域(PF00249)使用PFam数据库(http://pfam.xfam.org/ )的隐马尔可夫模型(HMM)配置文件进行识别和确认,该配置文件用作对先前注释的黄瓜MYB序列的查询,通过HMMER SEARCH进一步识别并使用默认参数手动筛选(目标序列的期望值E≤1.2×10−8)[17]。应用HMMER SEARCH建立黄瓜MYB (CsMYB)基因家族特异性模型[18],使用黄瓜MYB特异性HMM选择E低于0.001的所有黄瓜MYB蛋白。CsMYB家族的候选基因根据黄瓜特异性MYB HMM确定。以上最初获得的所有候选CsMYB基因均通过保守域数据库(conserved domain database,CDD)[19]和简单模块化架构研究工具(simplemodular architecture research tool,SMART)使用默认参数E≤0.010[20]。根据MYB序列内的特征性保守结构域,分离黄瓜R2R3-MYB基因。 -
通过ClustalW对模式植物拟南芥、木本植物杨梅Morella rubra[21]和草本植物黄瓜[1]的R2R3-MYB序列进行比对,采用MEGA-11软件的邻接法(neighbor-joining,NJ),设置自展法系数(bootstrap)为1 000次,其余参数按系统默认构建进化树[22]。
-
通过筛选结果得到的黄瓜R2R3-MYB基因的序列信息,利用TBtools将其定位于染色体上。采用ExPASy-ProtParam (
https://www.expasy.org/resources/protparam )在线预测黄瓜R2R3-MYB蛋白的氨基酸数量、分子量、理论等电点、不稳定指数及脂肪系数;使用Plant-mPLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )预测亚细胞定位。 -
利用TBtools中的Gene Location Visualize功能绘制黄瓜R2R3-MYB家族成员的染色体定位图,并通过MCscanX将黄瓜R2R3-MYB亚家族成员基因与拟南芥同源基因进行共线性分析。
-
利用TBtools对黄瓜R2R3-MYB亚家族成员的基因结构进行可视化分析,并对其蛋白的保守基序进行分析,限值为15,绘制结果图。
-
利用TBtools提取黄瓜R2R3-MYB亚家族成员上游1 500 bp的序列,通过PlantCARE 数据库(
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ )获得启动子顺式作用元件,利用Excel筛选结果,再利用TBtools对其最终结果进行可视化。 -
通过网站STRING (
https://www.string-db.org/ )对黄瓜R2R3-MYB蛋白成员之间进行互作预测分析,研究黄瓜R2R3-MYB蛋白成员之间的作用关系。 -
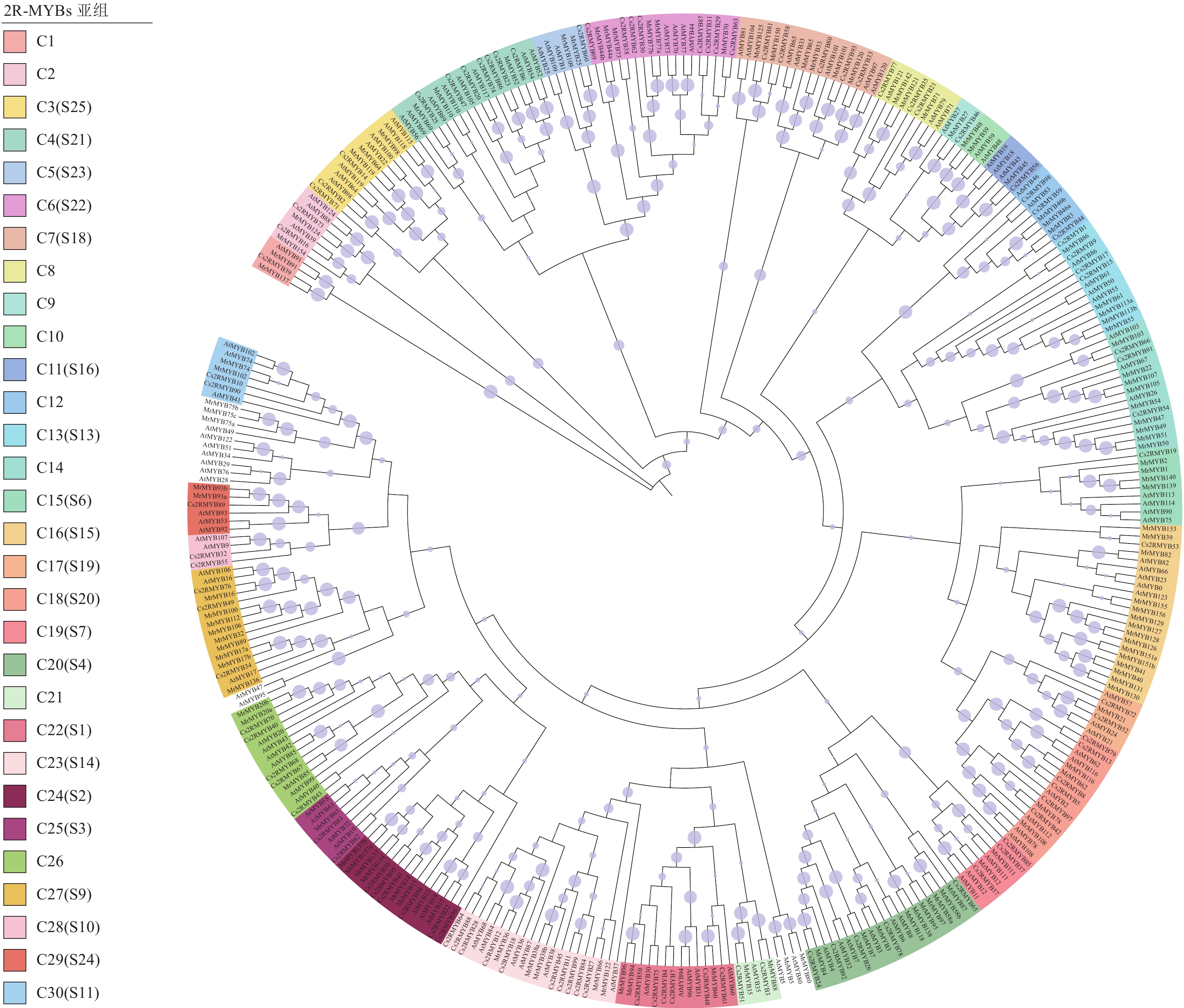
将黄瓜基因组数据与拟南芥进行比对,根据HMM模型进行搜索,取其交集,筛选鉴定其具有特殊结构域的序列,最终获得黄瓜99个R2R3-MYB基因家族成员。参照DUBOS等[23]的分类方法,将鉴定出的成员进一步分为30个亚组。为了探索系统发育关系,通过ClustalW进行序列比对,使用黄瓜99个CsMYB蛋白、拟南芥125个AtMYB蛋白和杨梅122个MrMYB蛋白构建了系统发育树(图1)。按照染色体定位结果,分别命名为Cs2RMYB1~Cs2RMYB99 (表1)。根据蛋白质序列的相似性和聚类结果的高自举性,将CsMYB进一步分为30个进化枝(指定为C1~C30),黄瓜R2R3-MYB基因家族成员被分成30个亚组。其中,C16、C20和C23亚组的家族成员数量最多,有21个成员;C18亚组成员数为16个;C14、C24和C27亚组家族成员数均为15个;C10和C28亚组家族成员数量等同,为4个;C9亚组成员数最少,仅3个。黄瓜R2R3-MYB亚家族的进化树聚类方式与拟南芥有一定的相似性。
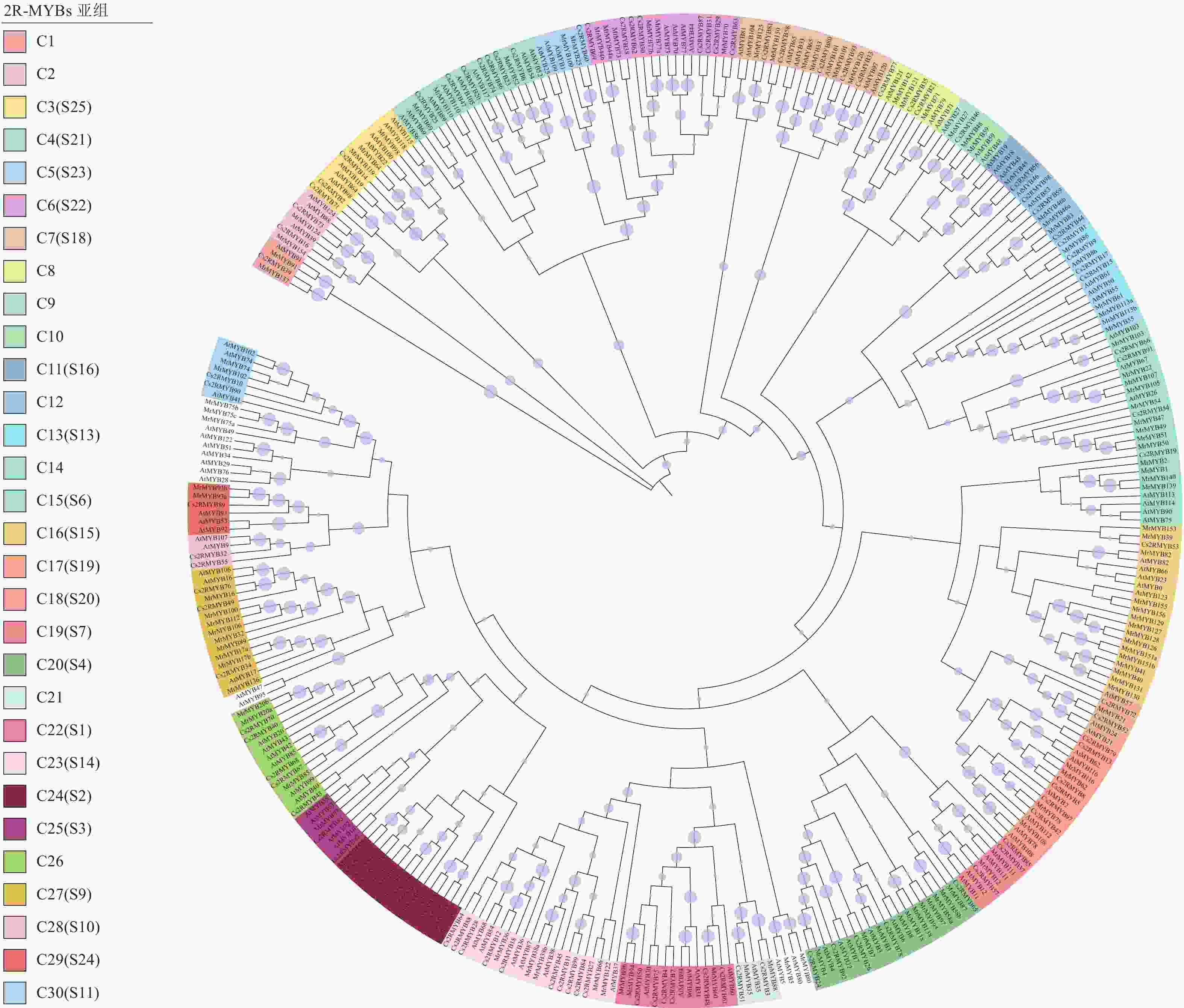
图 1 黄瓜与拟南芥、杨梅R2R3-MYB亚家族成员的系统进化树构建
Figure 1. Phylogenetic tree construction of C. sativus, A. thaliana and M. rubra R2R3-MYB subfamily members
表 1 黄瓜R2R3-MYB亚家族成员信息
Table 1. Cucumber R2R3-MYB subfamily member information
基因命名 氨基酸
数量/个分子量/kDa 等电点 不稳定
指数脂肪指数 染色体
位置基因命名 氨基酸
数量/个分子量/
kDa等电点 不稳定
指数脂肪指数 染色体
位置Cs2RMYB1 360 40.91 6.44 52.41 57.17 Chr 1 Cs2RMYB51 268 30.01 8.59 54.65 64.40 Chr 3 Cs2RMYB2 471 54.88 5.91 42.84 66.43 Chr 1 Cs2RMYB52 217 25.13 7.21 55.75 65.21 Chr 3 Cs2RMYB3 367 41.94 5.83 59.22 75.50 Chr 1 Cs2RMYB53 195 22.67 8.37 72.59 73.54 Chr 3 Cs2RMYB4 331 37.60 9.02 65.15 61.90 Chr 1 Cs2RMYB54 355 39.21 6.25 54.46 82.96 Chr 3 Cs2RMYB5 277 31.19 5.90 78.88 66.21 Chr 1 Cs2RMYB55 305 34.19 6.19 45.79 74.52 Chr 3 Cs2RMYB6 232 26.59 8.83 57.26 66.51 Chr 1 Cs2RMYB56 253 28.52 9.10 49.09 65.53 Chr 3 Cs2RMYB7 326 36.76 6.42 53.91 67.94 Chr 1 Cs2RMYB57 313 35.05 4.97 68.24 75.97 Chr 4 Cs2RMYB8 288 32.50 6.31 71.30 64.03 Chr 1 Cs2RMYB58 314 36.06 6.10 62.98 58.06 Chr 4 Cs2RMYB9 377 42.09 6.46 50.23 66.71 Chr 1 Cs2RMYB59 322 35.68 6.14 54.46 67.55 Chr 4 Cs2RMYB10 337 37.92 5.74 45.24 71.19 Chr 1 Cs2RMYB60 377 40.50 5.41 39.90 73.24 Chr 4 Cs2RMYB11 299 33.99 9.17 58.17 61.64 Chr 1 Cs2RMYB61 353 39.81 6.22 60.28 59.92 Chr 4 Cs2RMYB12 372 42.33 6.12 54.32 67.63 Chr 1 Cs2RMYB62 229 25.19 9.29 58.90 61.70 Chr 4 Cs2RMYB13 277 32.11 5.83 63.46 79.96 Chr 1 Cs2RMYB63 263 29.14 8.34 61.14 67.15 Chr 4 Cs2RMYB14 337 38.24 8.66 52.38 59.08 Chr 1 Cs2RMYB64 280 31.51 8.92 59.53 72.11 Chr 4 Cs2RMYB15 370 41.15 6.13 51.97 61.95 Chr 1 Cs2RMYB65 260 28.84 5.00 38.86 76.54 Chr 5 Cs2RMYB16 364 41.75 9.54 55.58 69.81 Chr 1 Cs2RMYB66 323 36.41 6.00 53.24 72.45 Chr 5 Cs2RMYB17 327 37.06 7.68 62.31 69.48 Chr 2 Cs2RMYB67 258 29.46 5.84 45.28 67.67 Chr 5 Cs2RMYB18 295 32.99 9.07 45.41 70.10 Chr 2 Cs2RMYB68 236 26.98 6.71 53.48 74.83 Chr 5 Cs2RMYB19 222 25.74 6.00 53.86 72.43 Chr 2 Cs2RMYB69 284 31.23 6.41 56.18 72.46 Chr 5 Cs2RMYB20 264 29.98 8.97 58.24 63.14 Chr 2 Cs2RMYB70 300 33.77 5.29 55.23 71.50 Chr 5 Cs2RMYB21 265 31.42 9.64 54.52 67.02 Chr 2 Cs2RMYB71 337 38.25 7.69 55.10 66.59 Chr 5 Cs2RMYB22 286 32.59 5.60 58.72 59.69 Chr 2 Cs2RMYB72 210 24.43 5.19 51.25 60.38 Chr 5 Cs2RMYB23 228 26.87 10.27 58.08 65.44 Chr 2 Cs2RMYB73 464 52.29 6.82 66.09 66.85 Chr 5 Cs2RMYB24 256 29.15 8.24 48.25 79.22 Chr 2 Cs2RMYB74 369 41.40 6.61 50.68 56.34 Chr 5 Cs2RMYB25 210 23.54 10.31 65.32 59.48 Chr 2 Cs2RMYB75 307 34.25 7.51 48.64 64.59 Chr 5 Cs2RMYB26 230 26.32 8.83 57.64 63.65 Chr 2 Cs2RMYB76 398 44.52 6.49 53.56 62.29 Chr 5 Cs2RMYB27 204 23.04 8.44 43.78 67.94 Chr 2 Cs2RMYB77 202 23.93 8.25 43.61 58.37 Chr 5 Cs2RMYB28 338 38.46 8.56 58.00 68.99 Chr 2 Cs2RMYB78 294 33.13 8.72 52.07 70.41 Chr 5 Cs2RMYB29 290 31.80 9.23 59.33 65.93 Chr 2 Cs2RMYB79 286 32.77 6.40 64.03 65.17 Chr 6 Cs2RMYB30 297 33.70 9.97 55.65 69.97 Chr 2 Cs2RMYB80 552 61.13 5.50 52.51 61.54 Chr 6 Cs2RMYB31 301 32.30 8.19 55.90 64.15 Chr 2 Cs2RMYB81 301 34.40 5.56 70.97 65.08 Chr 6 Cs2RMYB32 308 34.86 5.93 48.47 84.90 Chr 2 Cs2RMYB82 269 31.33 6.45 45.95 63.42 Chr 6 Cs2RMYB33 519 57.03 7.46 59.70 57.53 Chr 3 Cs2RMYB83 334 38.24 4.96 46.95 59.85 Chr 6 Cs2RMYB34 305 34.64 5.70 57.58 70.72 Chr 3 Cs2RMYB84 286 32.92 5.80 56.50 54.58 Chr 6 Cs2RMYB35 248 29.14 9.10 56.61 66.49 Chr 3 Cs2RMYB85 291 33.53 6.63 61.70 46.94 Chr 6 Cs2RMYB36 259 29.98 6.50 48.74 59.54 Chr 3 Cs2RMYB86 277 32.26 9.35 51.45 59.49 Chr 6 Cs2RMYB37 296 33.70 5.35 56.27 67.20 Chr 3 Cs2RMYB87 301 32.45 9.32 50.91 64.82 Chr 6 Cs2RMYB38 219 25.25 6.32 59.79 70.32 Chr 3 Cs2RMYB88 319 35.85 6.53 48.51 65.49 Chr 6 Cs2RMYB39 354 40.50 9.20 58.22 77.40 Chr 3 Cs2RMYB89 344 38.44 6.12 57.72 80.00 Chr 6 Cs2RMYB40 267 30.10 5.59 55.70 69.33 Chr 3 Cs2RMYB90 387 44.36 5.81 54.93 61.01 Chr 6 Cs2RMYB41 245 28.13 5.75 48.70 62.57 Chr 3 Cs2RMYB91 289 33.19 6.45 53.15 70.17 Chr 6 Cs2RMYB42 224 25.60 7.64 65.43 74.87 Chr 3 Cs2RMYB92 248 27.74 9.11 46.24 76.73 Chr 7 Cs2RMYB43 275 31.61 8.71 39.67 79.75 Chr 3 Cs2RMYB93 500 54.62 5.89 55.46 57.82 Chr 7 Cs2RMYB44 316 36.22 5.63 58.87 56.52 Chr 3 Cs2RMYB94 254 29.45 5.36 74.13 75.63 Chr 7 Cs2RMYB45 312 34.71 6.99 55.70 62.79 Chr 3 Cs2RMYB95 271 31.90 5.56 53.17 62.66 Chr 7 Cs2RMYB46 248 29.70 8.46 52.73 69.60 Chr 3 Cs2RMYB96 254 29.40 5.02 43.86 79.49 Chr 7 Cs2RMYB47 348 39.13 8.86 46.82 64.77 Chr 3 Cs2RMYB97 280 32.36 5.20 59.21 68.86 Chr 7 Cs2RMYB48 294 32.66 6.17 57.40 64.42 Chr 3 Cs2RMYB98 285 32.82 5.32 58.47 64.70 Chr 7 Cs2RMYB49 233 26.13 9.33 51.33 65.41 Chr 3 Cs2RMYB99 248 27.78 6.24 44.80 65.28 Scaffold 72 Cs2RMYB50 313 35.13 5.95 54.63 65.50 Chr 3 说明:Cs2RMYB48基因定位于叶绿体,其他基因定位于细胞核。 -
表1表明:黄瓜R2R3-MYB亚家族成员编码195~552个氨基酸,分子量为22.67~61.13 kDa,理论等电点为4.96~10.31,59个R2R3-MYB蛋白的等电点小于7,40个R2R3-MYB蛋白的等电点大于7,超过半数的蛋白为酸性。除Cs2RMYB43、Cs2RMYB60、Cs2RMYB65外,其余CsR2R3-MYB亚家族成员蛋白质不稳定指数为42.84~78.88,大于40.00,为不稳定蛋白。所有成员的脂肪系数相对较高,说明黄瓜R2R3-MYB亚家族成员适应环境的多样性。Plant-mPLoc 2.0网站预测结果显示:除Cs2RMYB48定位于叶绿体,其他成员均定位于细胞核。
-
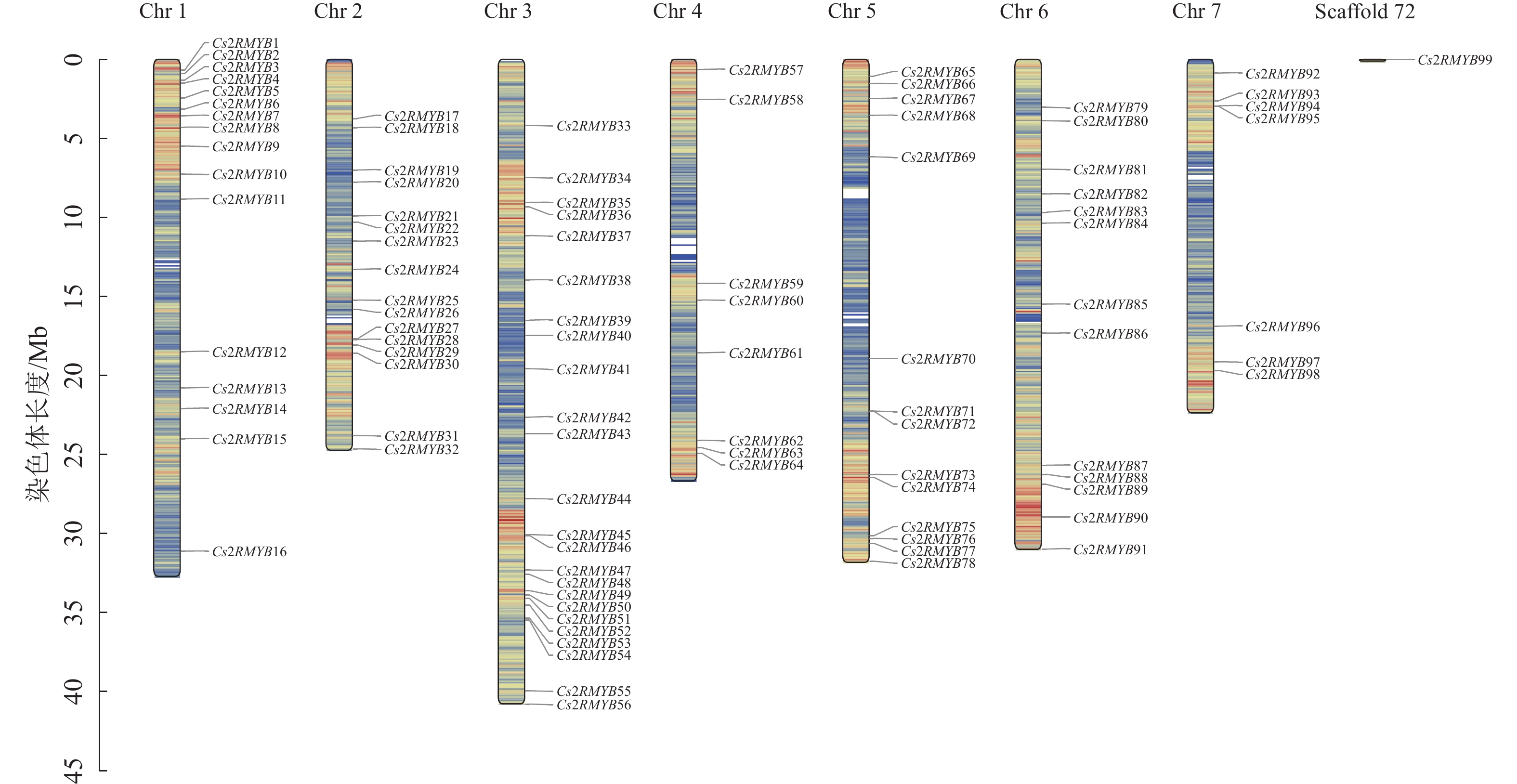
黄瓜R2R3-MYB亚家族成员映射于1~7号染色体上,Cs2RMYB99的染色体定位结果未知(图2)。每条染色体上的基因分布相对不均匀,1、4、5、7号染色体上基因大多分布在上下两端,而剩下的染色体上基因分布较为均匀。3号染色体上数量最多,可以定位到24个R2R3-MYB亚家族成员;1、2号染色体各有16个R2R3-MYB亚家族成员;5、6号染色体分别为14和13个R2R3-MYB亚家族成员;7号染色体上最少,为7个R2R3-MYB亚家族成员。此外,2号染色体上的Cs2RMYB27和Cs2RMYB28、3号染色体上的Cs2RMYB45和Cs2RMYB46、5号染色体上的Cs2RMYB71和Cs2RMYB72、7号染色体上的Cs2RMYB94和Cs2RMYB95均在各自染色体上形成基因簇。根据进化树分析可知:形成基因簇的成员之间同源性较高,推测成员间功能相对保守。
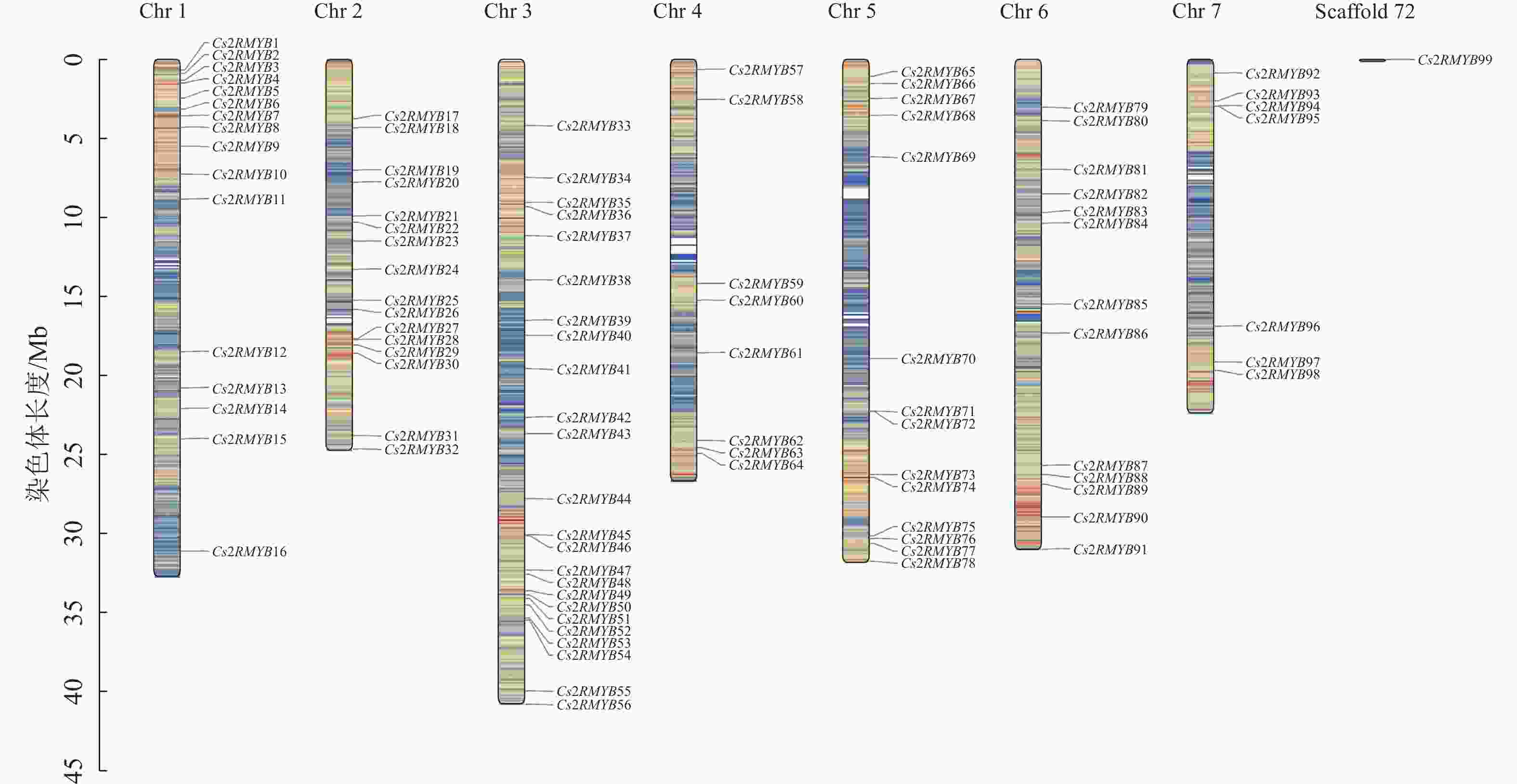
图 2 黄瓜R2R3-MYB亚家族染色体定位图
Figure 2. Chromosome location map of C. sativus R2R3-MYB subfamily
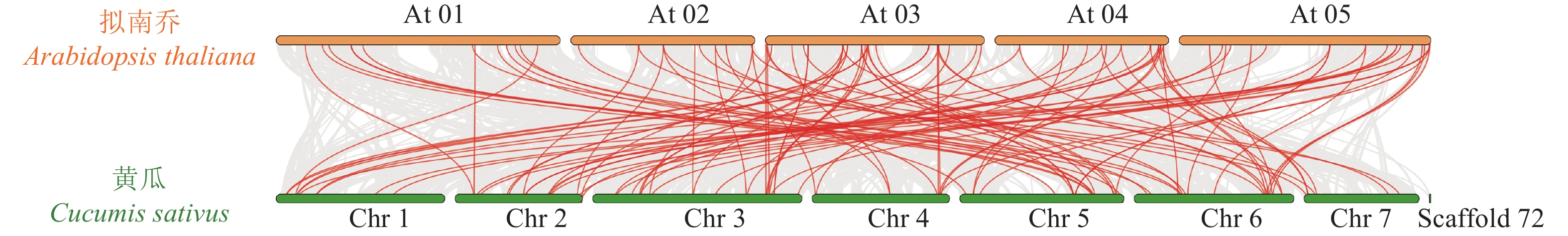
为揭示黄瓜R2R3-MYB亚家族成员在整个黄瓜基因组中的连锁关系,对黄瓜与拟南芥MYB基因进行共线性分析(图3),结果表明:25对基因表现出共线性和保守的连锁关系,即表明2个物种之间MYB基因家族存在高同源性。
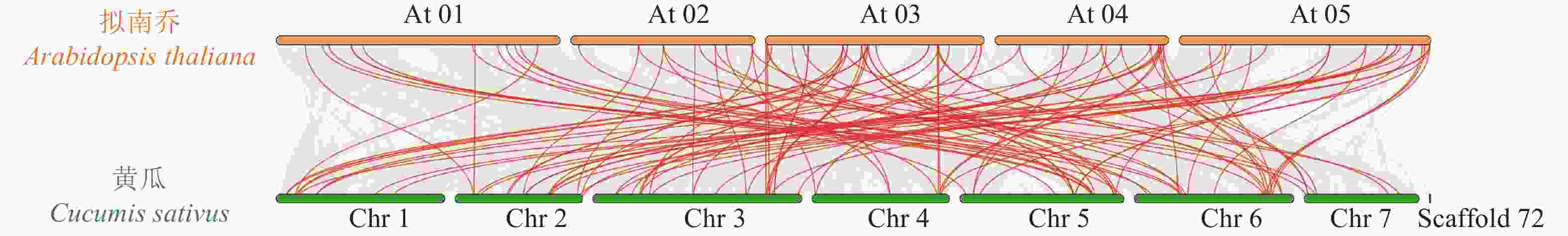
图 3 黄瓜R2R3-MYB亚家族与拟南芥共线性分析图
Figure 3. Analysis of the collinearity between C. sativus R2R3-MYB subfamily and A. thaliana
-
利用TBtools对黄瓜R2R3-MYB亚家族进行保守基序分析(图4)。结果显示:在99个R2R3-MYB蛋白中鉴定出10个不同的基序,其中Motif 3、Motif 7、Motif 2和Motif 4是高度保守的基序,所有成员均含有Motif 3,导致成员间功能相似。表明这些黄瓜R2R3-MYB亚家族成员间拥有特有或相同的基序,导致了它们之间具有特定的或者相似的功能。
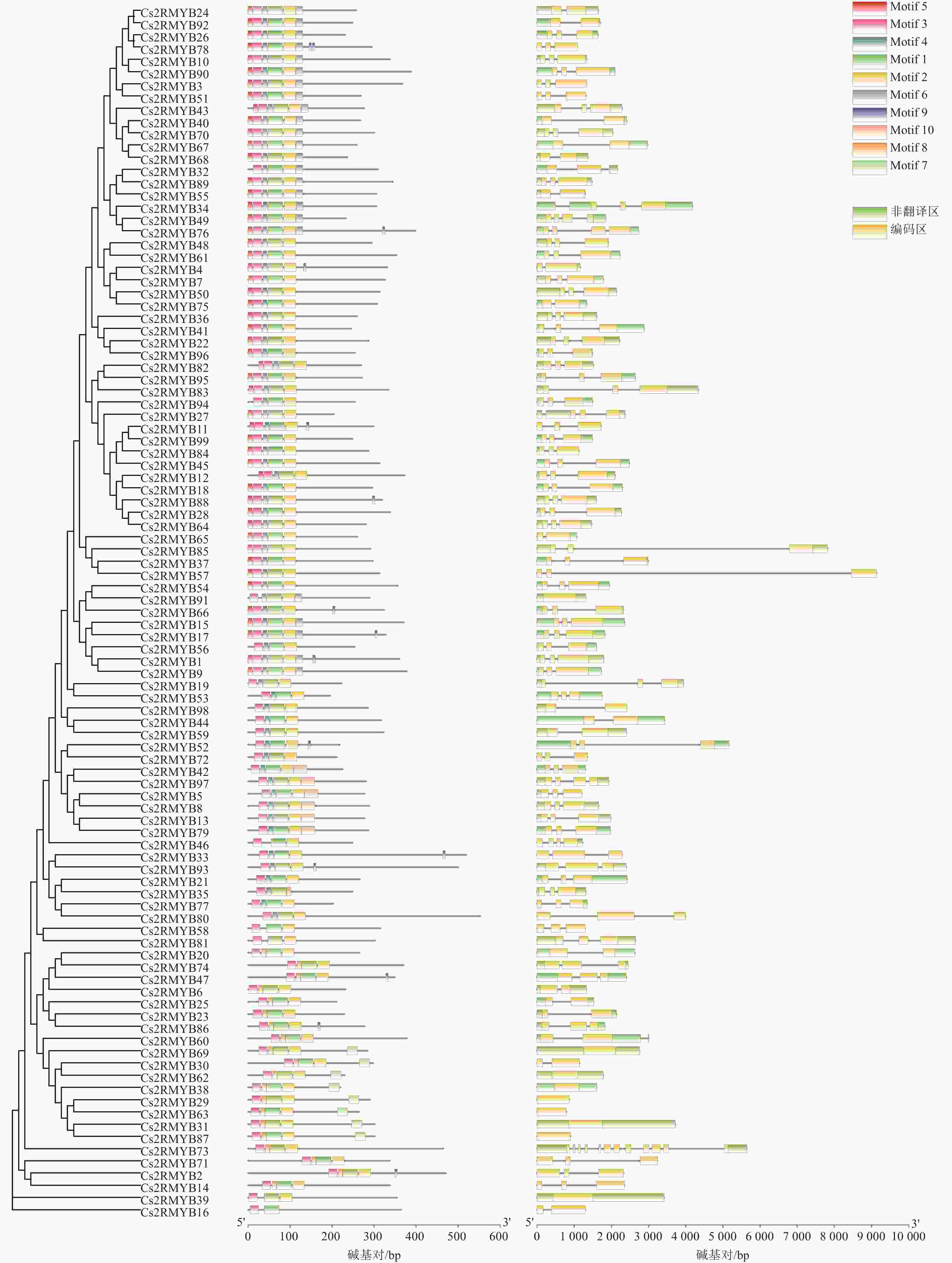
图 4 黄瓜R2R3-MYB亚家族蛋白保守基序和基因结构
Figure 4. Protein conservative motifs and gene structure of R2R3-MYB subfamily in C. sativus
通过Jalview对黄瓜R2R3-MYB亚家族进行序列比对(图5)。结果显示:2R重复序列中有3个色氨酸残基(W)及3R重复序列中有3个色氨酸残基,形成疏水结构,此为鉴定MYB家族典型的结构域特征。除保守的W外,2R重复序列的末端还存在12个高度保守的氨基酸残基,如精氨酸(R)、甘氨酸(G)、赖氨酸(K)、丝氨酸(S)、半胱氨酸(C)、亮氨酸(L)等,3R重复序列中的谷氨酸(E)、G、丙氨酸(A)、R、苏氨酸(T)等。

图 5 黄瓜R2R3-MYB亚家族蛋白序列比对图
Figure 5. Sequence comparison of C. sativus R2R3-MYB subfamily proteins
-
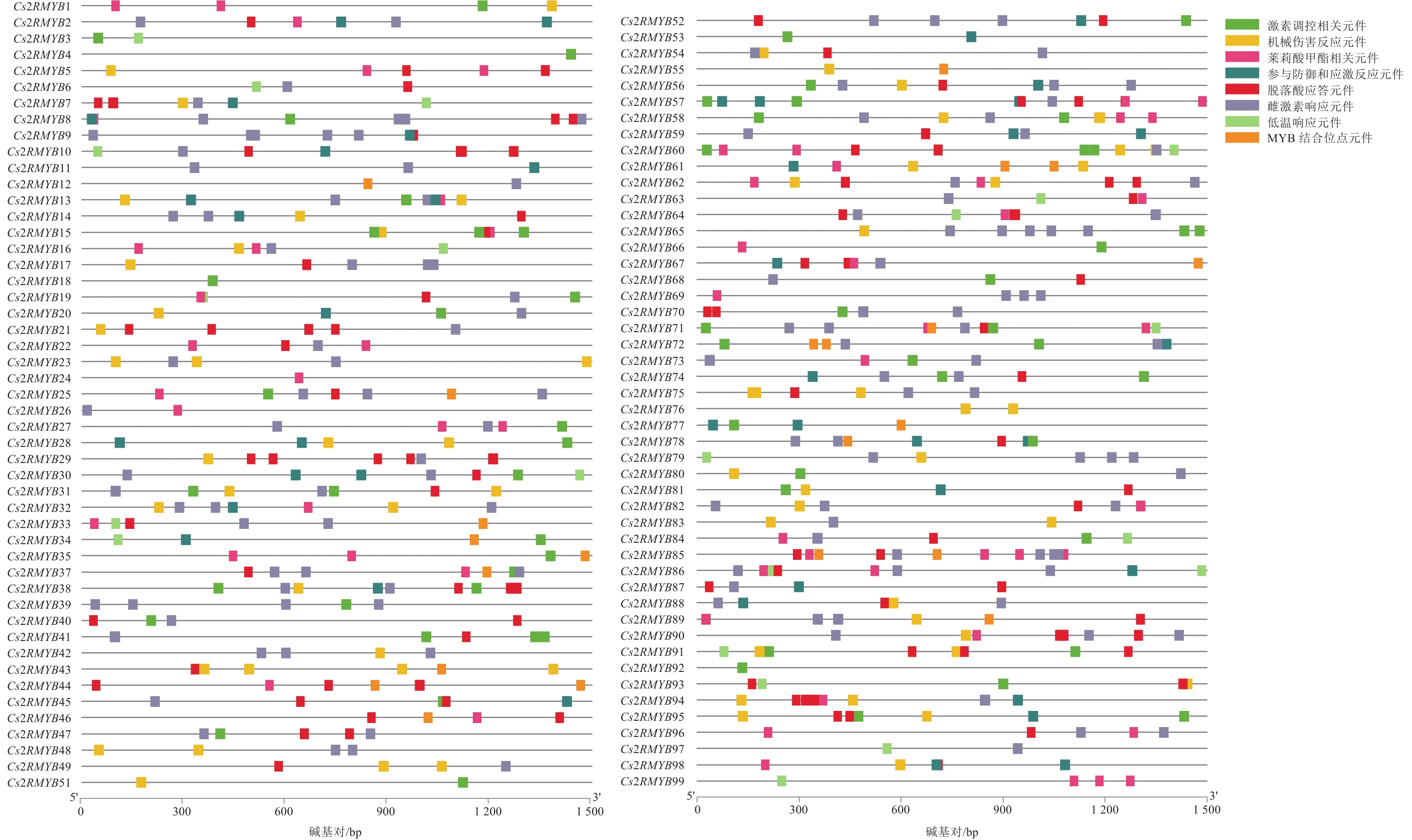
为了进一步研究黄瓜R2R3-MYB亚家族成员启动子区的顺式作用元件,通过PlantCARE在线网站分析了该亚家族上游1 500 bp区域(图6)。其启动子元件多与激素、抗逆及厌氧诱导相关,大部分黄瓜R2R3-MYB亚家族成员含雌激素响应(ERE)作用元件。从元件类型来看,ERE元件最多,分布于绝大多数成员的启动子区域上。从单个元件的具体分布来看,约68%家族成员启动子区含ERE元件,约60%成员启动子区含脱落酸应答元件(ABRE)。从单个基因家族成员来看,Cs2RMYB92启动子只有TCA-element激素调控相关元件;Cs2RMYB98有MYB结合位点,并富含激素响应与厌氧诱导元件,例如ABRE、ERE、茉莉酸甲酯相关元件(CGTCA)和MYB结合位点元件(MBS)。Cs2RMYB5、Cs2RMYB7和Cs2RMYB8等基因上有ABRE元件,说明这些基因可能与脱落酸响应有关;Cs2RMYB2、Cs2RMYB6和Cs2RMYB9等基因上含ERE元件,说明这些基因可能响应乙烯的调控;Cs2RMYB1、Cs2RMYB16和Cs2RMYB25等基因上有CGTCA元件,说明这些基因可能与茉莉酸甲酯响应有关;Cs2RMYB12、Cs2RMYB25、Cs2RMYB33和Cs2RMYB34等基因启动子元件上都含MBS元件,说明这些基因可能响应干旱胁迫的调控。
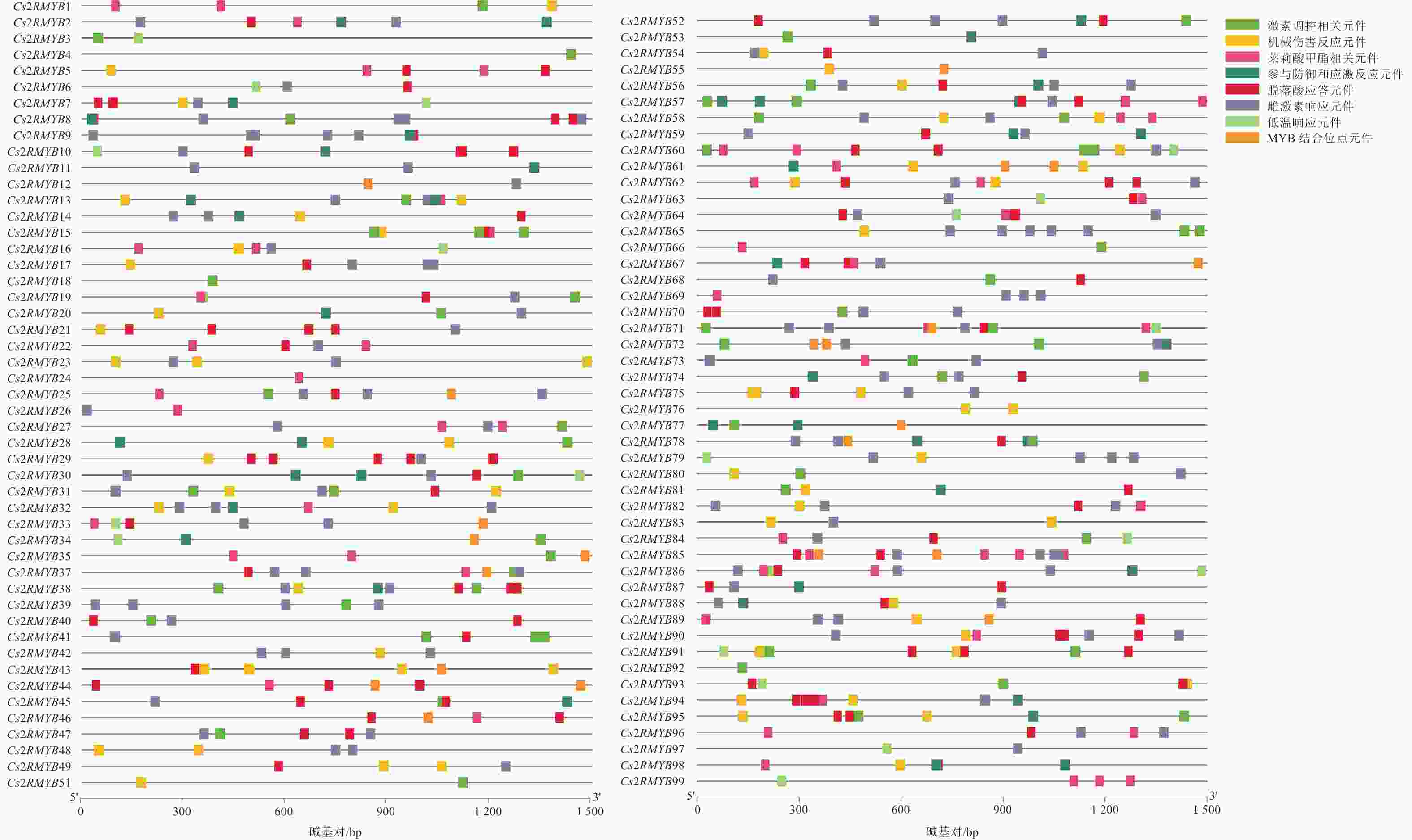
图 6 黄瓜R2R3-MYB亚家族上游顺式元件预测
Figure 6. Prediction of upstream homeopathic elements of C. sativus R2R3-MYB subfamily
-
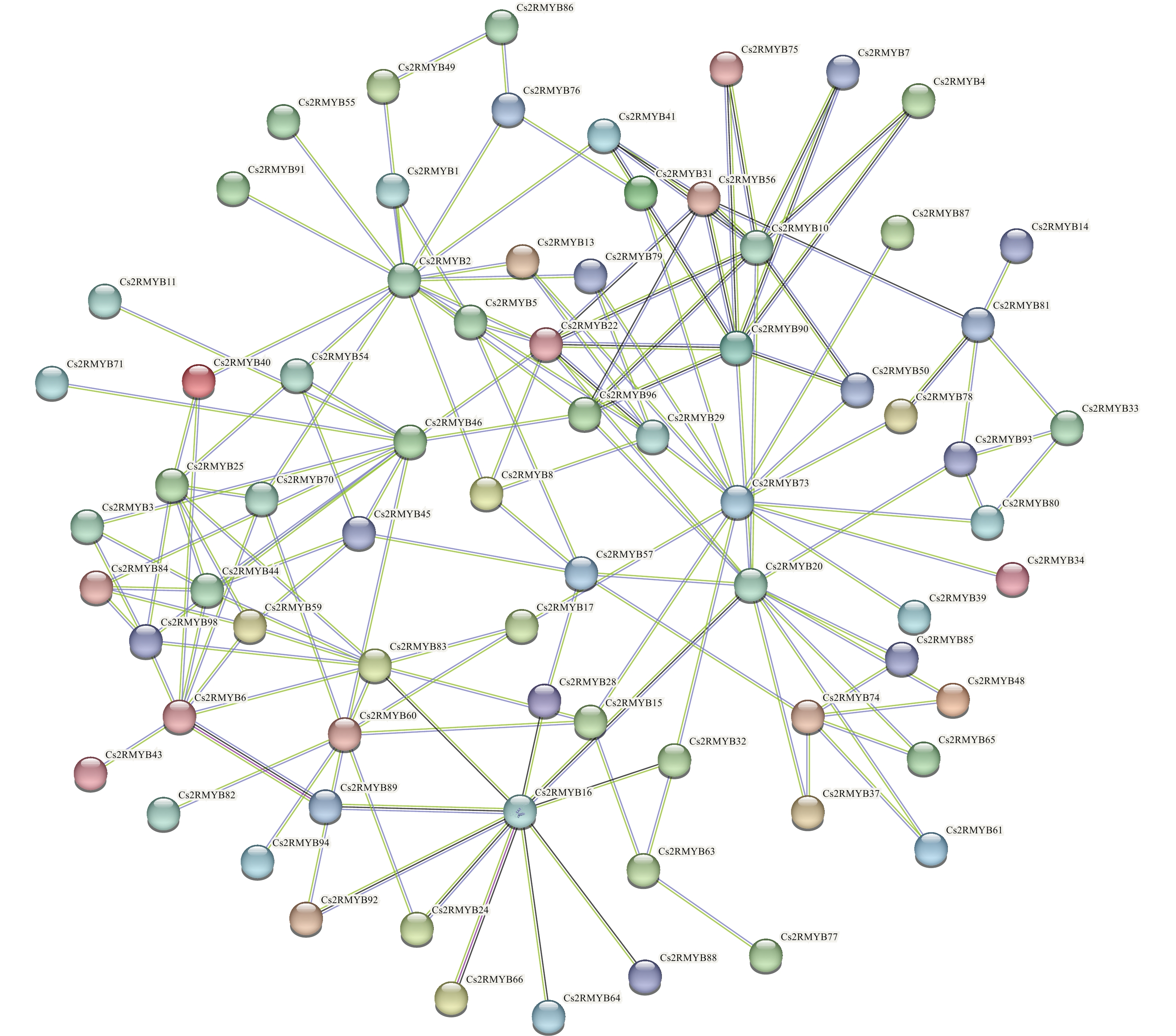
为深入了解黄瓜R2R3-MYB亚家族成员蛋白的互作情况,对99个成员进行蛋白质互作预测(图7),结果表明:Cs2RMYB25、Cs2RMYB3、Cs2RMYB84、Cs2RMYB98、Cs2RMYB6、Cs2RMYB6均与Cs2RMYB44存在互作关系;Cs2RMYB96、Cs2RMYB22、Cs2RMYB31、Cs2RMYB41、Cs2RMYB56、Cs2RMYB75、Cs2RMYB7、Cs2RMYB4、Cs2RMYB50均与Cs2RMYB90存在互作关系,且有较强的关联性。此外,部分成员间虽存在互作相关性,但各成员之间的具体机制并不明晰,仍需进一步深入研究。
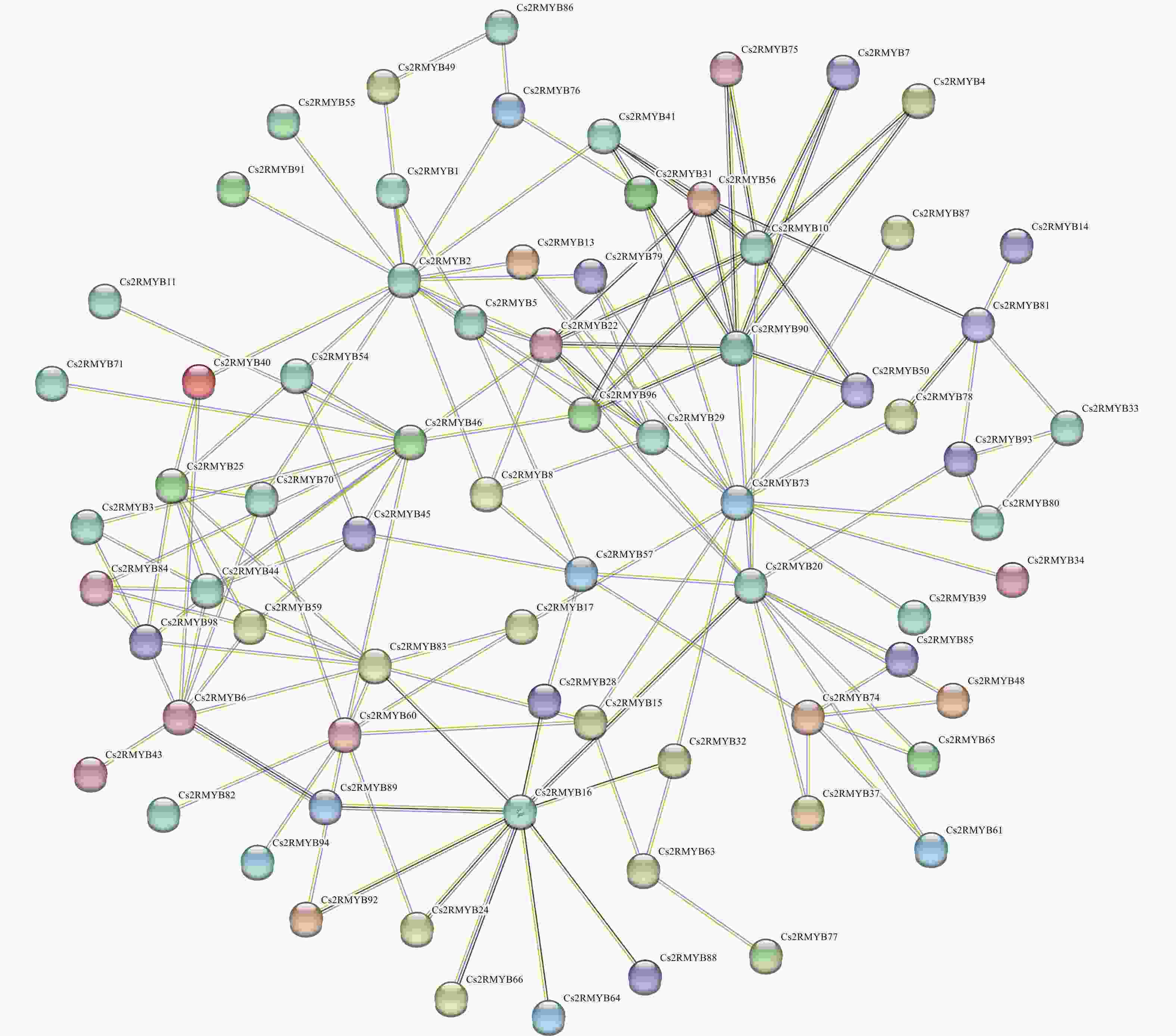
图 7 黄瓜R2R3-MYB亚家族成员蛋白的互作预测
Figure 7. Prediction of protein interactions among R2R3-MYB subfamily members in C. sativus
-
本研究共鉴定出99个黄瓜R2R3-MYB亚家族成员,约占黄瓜总MYB数的60%,数量多于西瓜(89个)[4],少于拟南芥(126个)[9]、水稻(102个)[10]和玉米(200个)[15]。黄瓜的基因组大小为224.8~251.1 Mb[24],而拟南芥、水稻、玉米和西瓜的基因组大小分别为125.0、430.0、2 300.0和365.1 Mb[25−28],R2R3-MYB亚家族成员数量与植物的基因组大小并没有直接的相关性。
作为MYB家族最大的亚家族,R2R3-MYB家族成员具有典型的模式结构,在植物发育、新陈代谢、信号转导及响应生物和非生物胁迫中起着非常重要的作用[29]。为了研究黄瓜基因组进化过程中功能保守的R2R3-MYB基因功能,构建其系统进化树并进行分析,结果表明:黄瓜Cs2RMYB 30个亚组中大多数包含来自拟南芥和杨梅R2R3-MYB成员,然而,C28亚组仅拥有黄瓜和拟南芥R2R3-MYB成员,表明它们可能来自共同的祖先。同时,外显子和内含子的结构分析有利于研究基因家族内部的进化关系,通过分组可以初步推测其功能,聚类较近的基因存在功能相似性,如在C30亚组中,拟南芥AtMYB42和AtMYB85被证明参与木质素生物合成代谢调控[30],黄瓜Cs2RMYB72和Cs2RMYB73与其表现出较近的亲缘关系,可能与木质素生物合成代谢有关;在C15亚组中,AtMYB75、AtMYB90、AtMYB113和AtMYB114过量表达会促进植物花青素的积累[31],与黄瓜Cs2RMYB19同属于C15亚组;在C22亚组中,AtMYB93是其分支上唯一一个影响拟南芥侧根发育的负调控因子[32],而黄瓜Cs2RMYB89与AtMYB93进化关系最近;宫思宇等[33]研究表明:AtMYB94通过调节拟南芥表皮的蜡质含量提高植株耐旱性,而C22亚组中的Cs2RMYB48是其同源基因,可能具有类似功能,同一分支亚家族转录因子可能存在功能相似性[23, 34]。
基序分析结果发现:所有的CsR2R3-MYB基因都有共同的保守基序,但也有独特的基序,这为CsR2R3-MYB亚家族成员的分类提供参考,且可能与CsR2R3-MYB基因的功能分化相关。此外,启动子序列上的顺式作用元件是转录因子识别并发挥转录调控作用的重要识别位点[35]。本研究发现:黄瓜CsR2R3-MYB亚家族成员启动子上的顺式作用元件种类繁多,绝大多数CsR2R3-MYB基因家族成员基因启动子区都含有ERE元件,暗示其可能参与黄瓜性别分化等生长发育过程;富含ABRE和CGTCA等激素响应顺式作用元件,表明CsR2R3-MYB基因家族在黄瓜应答逆境胁迫中发挥重要作用。由于CsR2R3-MYB基因调控元件的不同功能,启动子区域中这些高度多样化的顺式调控元件也可能反映了转录水平上的功能分歧[36]。本研究初步预测了MYB家族CsR2R3-MYB基因与植物生长发育及逆境胁迫响应相关,可为后续该亚家族成员基因功能深入研究提供参考。
Identification and bioinformatics analysis of R2R3-MYB subfamily in cucumber
-
摘要:
目的 深入研究黄瓜Cucumis sativus R2R3-MYB亚家族成员的相关功能。 方法 利用生物信息学手段分析黄瓜全基因组,鉴定R2R3-MYB亚家族成员,对其系统进化关系、蛋白理化性质、染色体定位、基因结构、保守基序、顺式作用元件、蛋白质互作进行分析。 结果 黄瓜全基因组中含99个具有典型结构域的R2R3-MYB转录因子,蛋白序列含195~552个氨基酸,有保守基序及氨基酸位点;基因在染色体上分布不均匀;大部分亚家族成员蛋白质的不稳定指数大于40,属于不稳定蛋白。顺式作用调控元件分析发现:大部分基因启动子区所含元件与激素调节、MYB结合位点、胁迫密切相关。 结论 通过黄瓜全基因组鉴定,获得黄瓜基因组99个R2R3-MYB家族成员,分为30个亚组,映射于7条染色体上,该家族成员的上游启动子区含逆境相关作用元件。图7表1参36 Abstract:Objective This study is aimed to conduct an in-depth study of the functions associated with members of the R2R3-MYB subfamily in cucumber (Cucumis sativus). Method First, an analysis was conducted of the whole cucumber genome so as to identify R2R3-MYB subfamily members. Then, bioinformatics was employed to analyze their phylogenetic relationships, physicochemical properties of proteins, chromosomal localization, gene structure, conserved motifs, and cis-acting elements, as well as the protein interactions. Result The whole cucumber genome contained 99 R2R3-MYB transcription factors with typical structural domains, and the protein sequences contained amino acids ranging from 195 to 552, with conserved motifs and amino acid sites. They were unevenly distributed on chromosomes. They were unstable proteins with the instability index of most subfamily members being greater than 40. As was shown by the analysis of cis-acting regulatory elements, most of the elements contained in the promoter regions of genes were closely related to hormone regulation, MYB binding sites, and stress. Conclusion The R2R3-MYB family members were obtained through whole cucumber genome identification, which were divided into 30 subgroups and mapped on 7 chromosomes and the upstream promoter region of this family members contained stress-related action elements. [Ch, 7 fig. 1 tab. 36 ref.] -
Key words:
- Cucumis sativus /
- R2R3-MYB /
- transcription factor /
- whole genome identification
-
图 1 黄瓜与拟南芥、杨梅R2R3-MYB亚家族成员的系统进化树构建
Figure 1 Phylogenetic tree construction of C. sativus, A. thaliana and M. rubra R2R3-MYB subfamily members
图 2 黄瓜R2R3-MYB亚家族染色体定位图
Figure 2 Chromosome location map of C. sativus R2R3-MYB subfamily
图 3 黄瓜R2R3-MYB亚家族与拟南芥共线性分析图
Figure 3 Analysis of the collinearity between C. sativus R2R3-MYB subfamily and A. thaliana
图 4 黄瓜R2R3-MYB亚家族蛋白保守基序和基因结构
Figure 4 Protein conservative motifs and gene structure of R2R3-MYB subfamily in C. sativus
图 5 黄瓜R2R3-MYB亚家族蛋白序列比对图
Figure 5 Sequence comparison of C. sativus R2R3-MYB subfamily proteins
图 6 黄瓜R2R3-MYB亚家族上游顺式元件预测
Figure 6 Prediction of upstream homeopathic elements of C. sativus R2R3-MYB subfamily
图 7 黄瓜R2R3-MYB亚家族成员蛋白的互作预测
Figure 7 Prediction of protein interactions among R2R3-MYB subfamily members in C. sativus
表 1 黄瓜R2R3-MYB亚家族成员信息
Table 1. Cucumber R2R3-MYB subfamily member information
基因命名 氨基酸
数量/个分子量/kDa 等电点 不稳定
指数脂肪指数 染色体
位置基因命名 氨基酸
数量/个分子量/
kDa等电点 不稳定
指数脂肪指数 染色体
位置Cs2RMYB1 360 40.91 6.44 52.41 57.17 Chr 1 Cs2RMYB51 268 30.01 8.59 54.65 64.40 Chr 3 Cs2RMYB2 471 54.88 5.91 42.84 66.43 Chr 1 Cs2RMYB52 217 25.13 7.21 55.75 65.21 Chr 3 Cs2RMYB3 367 41.94 5.83 59.22 75.50 Chr 1 Cs2RMYB53 195 22.67 8.37 72.59 73.54 Chr 3 Cs2RMYB4 331 37.60 9.02 65.15 61.90 Chr 1 Cs2RMYB54 355 39.21 6.25 54.46 82.96 Chr 3 Cs2RMYB5 277 31.19 5.90 78.88 66.21 Chr 1 Cs2RMYB55 305 34.19 6.19 45.79 74.52 Chr 3 Cs2RMYB6 232 26.59 8.83 57.26 66.51 Chr 1 Cs2RMYB56 253 28.52 9.10 49.09 65.53 Chr 3 Cs2RMYB7 326 36.76 6.42 53.91 67.94 Chr 1 Cs2RMYB57 313 35.05 4.97 68.24 75.97 Chr 4 Cs2RMYB8 288 32.50 6.31 71.30 64.03 Chr 1 Cs2RMYB58 314 36.06 6.10 62.98 58.06 Chr 4 Cs2RMYB9 377 42.09 6.46 50.23 66.71 Chr 1 Cs2RMYB59 322 35.68 6.14 54.46 67.55 Chr 4 Cs2RMYB10 337 37.92 5.74 45.24 71.19 Chr 1 Cs2RMYB60 377 40.50 5.41 39.90 73.24 Chr 4 Cs2RMYB11 299 33.99 9.17 58.17 61.64 Chr 1 Cs2RMYB61 353 39.81 6.22 60.28 59.92 Chr 4 Cs2RMYB12 372 42.33 6.12 54.32 67.63 Chr 1 Cs2RMYB62 229 25.19 9.29 58.90 61.70 Chr 4 Cs2RMYB13 277 32.11 5.83 63.46 79.96 Chr 1 Cs2RMYB63 263 29.14 8.34 61.14 67.15 Chr 4 Cs2RMYB14 337 38.24 8.66 52.38 59.08 Chr 1 Cs2RMYB64 280 31.51 8.92 59.53 72.11 Chr 4 Cs2RMYB15 370 41.15 6.13 51.97 61.95 Chr 1 Cs2RMYB65 260 28.84 5.00 38.86 76.54 Chr 5 Cs2RMYB16 364 41.75 9.54 55.58 69.81 Chr 1 Cs2RMYB66 323 36.41 6.00 53.24 72.45 Chr 5 Cs2RMYB17 327 37.06 7.68 62.31 69.48 Chr 2 Cs2RMYB67 258 29.46 5.84 45.28 67.67 Chr 5 Cs2RMYB18 295 32.99 9.07 45.41 70.10 Chr 2 Cs2RMYB68 236 26.98 6.71 53.48 74.83 Chr 5 Cs2RMYB19 222 25.74 6.00 53.86 72.43 Chr 2 Cs2RMYB69 284 31.23 6.41 56.18 72.46 Chr 5 Cs2RMYB20 264 29.98 8.97 58.24 63.14 Chr 2 Cs2RMYB70 300 33.77 5.29 55.23 71.50 Chr 5 Cs2RMYB21 265 31.42 9.64 54.52 67.02 Chr 2 Cs2RMYB71 337 38.25 7.69 55.10 66.59 Chr 5 Cs2RMYB22 286 32.59 5.60 58.72 59.69 Chr 2 Cs2RMYB72 210 24.43 5.19 51.25 60.38 Chr 5 Cs2RMYB23 228 26.87 10.27 58.08 65.44 Chr 2 Cs2RMYB73 464 52.29 6.82 66.09 66.85 Chr 5 Cs2RMYB24 256 29.15 8.24 48.25 79.22 Chr 2 Cs2RMYB74 369 41.40 6.61 50.68 56.34 Chr 5 Cs2RMYB25 210 23.54 10.31 65.32 59.48 Chr 2 Cs2RMYB75 307 34.25 7.51 48.64 64.59 Chr 5 Cs2RMYB26 230 26.32 8.83 57.64 63.65 Chr 2 Cs2RMYB76 398 44.52 6.49 53.56 62.29 Chr 5 Cs2RMYB27 204 23.04 8.44 43.78 67.94 Chr 2 Cs2RMYB77 202 23.93 8.25 43.61 58.37 Chr 5 Cs2RMYB28 338 38.46 8.56 58.00 68.99 Chr 2 Cs2RMYB78 294 33.13 8.72 52.07 70.41 Chr 5 Cs2RMYB29 290 31.80 9.23 59.33 65.93 Chr 2 Cs2RMYB79 286 32.77 6.40 64.03 65.17 Chr 6 Cs2RMYB30 297 33.70 9.97 55.65 69.97 Chr 2 Cs2RMYB80 552 61.13 5.50 52.51 61.54 Chr 6 Cs2RMYB31 301 32.30 8.19 55.90 64.15 Chr 2 Cs2RMYB81 301 34.40 5.56 70.97 65.08 Chr 6 Cs2RMYB32 308 34.86 5.93 48.47 84.90 Chr 2 Cs2RMYB82 269 31.33 6.45 45.95 63.42 Chr 6 Cs2RMYB33 519 57.03 7.46 59.70 57.53 Chr 3 Cs2RMYB83 334 38.24 4.96 46.95 59.85 Chr 6 Cs2RMYB34 305 34.64 5.70 57.58 70.72 Chr 3 Cs2RMYB84 286 32.92 5.80 56.50 54.58 Chr 6 Cs2RMYB35 248 29.14 9.10 56.61 66.49 Chr 3 Cs2RMYB85 291 33.53 6.63 61.70 46.94 Chr 6 Cs2RMYB36 259 29.98 6.50 48.74 59.54 Chr 3 Cs2RMYB86 277 32.26 9.35 51.45 59.49 Chr 6 Cs2RMYB37 296 33.70 5.35 56.27 67.20 Chr 3 Cs2RMYB87 301 32.45 9.32 50.91 64.82 Chr 6 Cs2RMYB38 219 25.25 6.32 59.79 70.32 Chr 3 Cs2RMYB88 319 35.85 6.53 48.51 65.49 Chr 6 Cs2RMYB39 354 40.50 9.20 58.22 77.40 Chr 3 Cs2RMYB89 344 38.44 6.12 57.72 80.00 Chr 6 Cs2RMYB40 267 30.10 5.59 55.70 69.33 Chr 3 Cs2RMYB90 387 44.36 5.81 54.93 61.01 Chr 6 Cs2RMYB41 245 28.13 5.75 48.70 62.57 Chr 3 Cs2RMYB91 289 33.19 6.45 53.15 70.17 Chr 6 Cs2RMYB42 224 25.60 7.64 65.43 74.87 Chr 3 Cs2RMYB92 248 27.74 9.11 46.24 76.73 Chr 7 Cs2RMYB43 275 31.61 8.71 39.67 79.75 Chr 3 Cs2RMYB93 500 54.62 5.89 55.46 57.82 Chr 7 Cs2RMYB44 316 36.22 5.63 58.87 56.52 Chr 3 Cs2RMYB94 254 29.45 5.36 74.13 75.63 Chr 7 Cs2RMYB45 312 34.71 6.99 55.70 62.79 Chr 3 Cs2RMYB95 271 31.90 5.56 53.17 62.66 Chr 7 Cs2RMYB46 248 29.70 8.46 52.73 69.60 Chr 3 Cs2RMYB96 254 29.40 5.02 43.86 79.49 Chr 7 Cs2RMYB47 348 39.13 8.86 46.82 64.77 Chr 3 Cs2RMYB97 280 32.36 5.20 59.21 68.86 Chr 7 Cs2RMYB48 294 32.66 6.17 57.40 64.42 Chr 3 Cs2RMYB98 285 32.82 5.32 58.47 64.70 Chr 7 Cs2RMYB49 233 26.13 9.33 51.33 65.41 Chr 3 Cs2RMYB99 248 27.78 6.24 44.80 65.28 Scaffold 72 Cs2RMYB50 313 35.13 5.95 54.63 65.50 Chr 3 说明:Cs2RMYB48基因定位于叶绿体,其他基因定位于细胞核。  下载: 导出CSV
下载: 导出CSV
-
[1] LIU Yi, ZHANG Zhishuo, FANG Ke, et al. Genome-wide analysis of the MYB-related transcription factor family in pepper and functional studies of CaMYB37 involvement in capsaicin biosynthesis [J/OL]. International Journal of Molecular Sciences, 2022, 23(19): 11667[2023-05-01]. doi: 10.3390/ijms231911667. [2] LI Jinlu, HAN Guoliang, SUN Cuifeng, et al. Research advances of MYB transcription factors in plant stress resistance and breeding [J/OL]. Plant Signaling & Behavior, 2019, 14(8): e1613131[2023-05-01]. doi: 10.1080/15592324.2019.1613131. [3] STRACKE R, HOLTGRÄWE D, SCHNEIDER J, et al. Genome-wide identification and characterization of R2R3-MYB genes in sugar beet (Beta vulgaris) [J/OL]. BMC Plant Biology, 2014, 14: 249[2023-05-01]. doi:10.1186/s12870-014-0249-8. [4] WANG Jin, LIU Yue, CHEN Xueliang, et al. Characterization and divergence analysis of duplicated R2R3-MYB genes in watermelon [J]. Journal of the American Society for Horticultural Science, 2020, 145(5): 281 − 288. [5] LI Sijia, HUANG Hu, MA Xianjin, et al. Characterizations of MYB transcription factors in Camellia oleifera reveal the key regulators involved in oil biosynthesis [J]. Horticulturae, 2022, 8(8): 742 − 761. [6] ZHAO Kai, CHENG Zihan, GUO Qing, et al. Characterization of the poplar R2R3-MYB gene family and over-expression of PsnMYB108 confers salt tolerance in transgenic tobacco [J/OL]. Frontiers in Plant Science, 2020, 11: 571881[2023-05-01]. doi: 10.3389/fpls.2020.571881. [7] SABIR I A, MANZOOR M A, SHAH I H, et al. MYB transcription factor family in sweet cherry (Prunus avium L.): genome-wide investigation, evolution, structure, characterization and expression patterns [J/OL]. BMC Plant Biology, 2022, 22: 2[2023-05-01]. doi: 10.1186/s12870-021-03374-y. [8] 邱文怡, 王诗雨, 李晓芳, 等. MYB转录因子参与植物非生物胁迫响应与植物激素应答的研究进展[J]. 浙江农业学报, 2020, 32(7): 1317 − 1328. QIU Wenyi, WANG Shiyu, LI Xiaofang, et al. Progress of MYB transcription factors involved in plant abiotic stress response and plant hormone response [J]. Zhejiang Journal of Agriculture, 2020, 32(7): 1317 − 1328. [9] STRACKE R, WERBER M, WEISSHAAR B. The R2R3-MYB gene family in Arabidopsis thaliana [J]. Current Opinion in Plant Biology, 2001, 4(5): 447 − 456. [10] CHEN Yanhui, YANG Xiaoyuan, HE Kun, et al. The MYB transcription factor superfamily of Arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family [J]. Plant Molecular Biology, 2006, 60(1): 107 − 124. [11] MATUS J T, AQUEA F, ARCE-JOHNSON P. Analysis of the grape MYB-R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across vitis and Arabidopsis genomes [J/OL]. BMC Plant Biology, 2008, 8: 83[2023-05-01]. doi: 10.1186/1471-2229-8-83. [12] LIU Chaoyang, WANG Xia, XU Yuantao, et al. Genome-wide analysis of the R2R3-MYB transcription factor gene family in sweet orange (Citrus sinensis) [J]. Molecular Biology Reports, 2014, 41(10): 6769 − 6785. [13] CAO Zhonghui, ZHANG Shizhong, WANG Rongkai, et al. Genome wide analysis of the apple MYB transcription factor family allows the identification of MdoMYB121 gene confering abiotic stress tolerance in plants [J/OL]. PLoS One, 2013, 8(7): e69955[2023-05-01]. doi:10.1371/journal.pone.0069955. [14] WILKINS O, NAHAL H, FOONG J, et al. Expansion and diversification of the Populus R2R3-MYB family of transcription factors [J]. Plant Physiology, 2009, 149(2): 981 − 993. [15] DIAS A P, BRAUN E L, MCMULLEN M D, et al. Recently duplicated maize R2R3-MYB genes provide evidence for distinct mechanisms of evolutionary divergence after duplication [J]. Plant Physiology, 2003, 131(2): 610 − 620. [16] LI Qiang, ZHANG Cunjia, LI Jing, et al. Genome-wide identification and characterization of R2R3-MYB family in Cucumis sativus [J]. PLoS One, 2012, 7(10): 1 − 18. [17] CHENG Chunyan, LI Qingrong, WANG Xing, et al. Identification and expression analysis of the CsMYB gene family in root knot nematode-resistant and susceptible cucumbers [J/OL]. Frontiers in Genetics, 2020, 11: 550677[2023-05-01]. doi: 10.3389/FGENE.2020.550677. [18] LOZANO R, HAMBLIN M T, PROCHNIK S, et al. Identification and distribution of the NBS-LRR gene family in the Cassava genome [J]. BMC Genomics, 2015, 16(1): 360 − 374. [19] MARCHLER-BAUER A, BO Y, HAN L Y, et al. CDD/SPARCLE: functional classification of proteins via subfamily domain architectures [J]. Nucleic Acids Research, 2017, 45(D1): D200 − D203. [20] MARCHLER-BAUER A, DERBYSHIRE M K, GONZALES N R, et al. CDD: NCBI’ s conserved domain database [J]. Nucleic Acids Research, 2015, 43(D1): D222 − D226. [21] CAO Yunlin, JIA Huimin, XING Mengyun, et al. Genome-wide analysis of MYB gene family in Chinese bayberry (Morella rubra) and identification of members regulating flavonoid biosynthesis [J/OL]. Frontiers in Plant Science, 2021, 12: 691384[2023-05-01]. doi: 10.3389/fpls.2021.691384. [22] KUMAR S, STECHER G, TAMURA K. MEGA 7: molecular evolutionary genetics analysis version 7.0 for bigger datasets [J]. Molecular Biology and Evolution, 2016, 33(7): 1870 − 1874. [23] DUBOS C, STRACKE R, GROTEWOLD E, et al. MYB transcription factors in Arabidopsis [J]. Trends in Plant Science, 2010, 15(10): 573 − 581. [24] LI Hongbo, WANG Shenhao, CHAI Sen, et al. Graph-based pan-genome reveals structural and sequence variations related to agronomic traits and domestication in cucumber [J]. Nature Communications, 2022, 13(1): 682 − 696. [25] KAUL S, KOO H L, JENKINS J, et al. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana [J]. Nature, 2000, 408(6814): 796 − 815. [26] BURR B. Mapping and sequencing the rice genome [J]. The Plant Cell, 2002, 14(3): 521 − 523. [27] SCHNABLE P S, WARE D, FULTON R S, et al. The B73 maize genome: complexity, diversity, and dynamics [J]. Science, 2009, 326(5956): 1112 − 1115. [28] GUO Shaogui, ZHAO Shengjie, SUN Honghe, et al. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits [J]. Nature Genetics, 2019, 51(11): 1616 − 1623. [29] BALDONI E, GENGA A, COMINELLI E. Plant MYB transcription factors: their role in drought response mechanisms [J]. International Journal of Molecular Sciences, 2015, 16(7): 15811 − 15851. [30] ZHONG Ruiqin, LEE Chanhui, ZHOU Jianli, et al. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis [J]. The Plant Cell, 2008, 20(10): 2763 − 2782. [31] FENG Shouqian, XU Yongchao, YANG Long, et al. Genome-wide identification and characterization of R2R3-MYB transcription factors in pear [J]. Scientia Horticulturae, 2015, 197: 176 − 182. [32] GIBBS D J, VOß U, HARDING S A, et al. AtMYB93 is a novel negative regulator of lateral root development in Arabidopsis [J]. New Phytologist, 2014, 203(4): 1194 − 1207. [33] 宫思宇, 陈海燕, 郭冬雪, 等. 黄瓜抗旱相关基因CsMYB94的克隆与表达分析[J]. 华北农学报, 2020, 35(增刊1): 18 − 23. GONG Siyu, CHEN Haiyan, GUO Dongxue, et al. Cloning and expression analysis of drought-related gene CsMYB94 in cucumber [J]. Journal of North China Agriculture, 2020, 35(suppl 1): 18 − 23. [34] HUANG Wenjun, SUN Wei, LÜ Haiyan, et al. Isolation and molecular characterization of thirteen R2R3-MYB transcription factors from Epimedium sagittatum [J]. International Journal of Molecular Sciences, 2013, 14(1): 594 − 610. [35] LI Junren, CHEN Xiuzhen, ZHOU Xuanxuan, et al. Identification of trihelix transcription factors in Pogostemon cablin reveals PatGT-1 negatively regulates patchoulol biosynthesis [J/OL]. Industrial Crops and Products, 2020, 161: 113182[2023-05-01]. doi:10.1016/j.indcrop.2020.113182. [36] YANG Jiahan, ZHANG Binghui, GU Gang, et al. Genome-wide identification and expression analysis of the R2R3-MYB gene family in tobacco (Nicotiana tabacum L.) [J]. BMC Genomics, 2022, 23(1): 432 − 453. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230278
 点击查看大图
点击查看大图
计量
- 文章访问数: 2355
- HTML全文浏览量: 925
- PDF下载量: 58
- 被引次数: 0
