





-
优化植物株型已被证明是提高作物种植密度、抗逆性和整体生产力的最佳方法之一[1]。 随着基因组学的发展,科学家们发现了被命名为IGT的保守基因家族,与植物分枝角度调控密切相关[2]。这个基因家族拥有1个保守的结构域[GϕL(A/T)IGT],包含着LAZY和TILLER ANGLE CONTROL (TAC)基因。这些基因在植物侧枝的遗传控制中扮演着重要角色[3]。
近几十年来的研究已经确认,LAZY基因在植物根部的向地生长和茎部的背地生长中起到了重要的调节作用。“LAZY”这个术语来源于被认为是“懒惰”的经典水稻Oryza sativa和玉米Zea mays植株,它们的特点在水稻中表现为分蘖扩散,而在玉米中表现为平卧[4−5]。LI等[6]通过研究水稻LAZY变异体,成功克隆了1个名为LAZY1的基因,该基因调控着分蘖角度。拟南芥Arabidopsis thaliana中也发现存在LAZY1突变体,被命名为AtLAZY1。拟南芥分枝朝外生长,并且比野生型植株的分枝角度更宽[3]。LAZY基因还被命名为其他名称,如DEEPER ROOTING(DRO)。水稻基因组携带有4个LAZY基因,分别是LAZY1、DRO1、DRO1-like 1(qSOR1, DRL1)和DRO1-like 2(DRL2)[7]。DRO1和DRL1的自然突变以及DRL2的CRISPR/Cas9技术产生的敲除突变都导致根部重力性反应受损,根系结构也发生改变,甚至出现裸露在地面的根[8]。GE等[9]在拟南芥中发现了3个类似DRO1的基因:AtDRO1(AtNGR2和At1g72490)、AtDRO2(AtNGR3和At1g19115)及AtDRO3(AtNGR1和At1g17400)。这3个基因都具有最保守的LAZY1结构域Ⅴ,即EAR基序,因而它们又分别被命名为AtLAZY4、AtLAZY3、AtLAZY2;相应地,At3g24750和At3g27025分别被命名为AtLAZY5和AtLAZY6。它们分别在根尖、根和叶柄中表达[10]。YU等[11]以水稻‘IR24’和‘Asominori’杂交,获得‘IL55’紧凑型杂交系,并成功定位克隆到水稻分蘖角度控制基因TAC1(TILLER ANGLE CONTROLLING)。TAC1基因增加了分枝角度,与LAZY1基因突变植物表型相反。研究表明:AtTAC1的表达具有光依赖性[12],并且在拟南芥中,几个LAZY基因的缺失导致其无法调整枝条角度以响应光照[13]。这些研究表明:IGT基因家族在响应光和重力刺激的植物株型调节中发挥作用。
大豆Glycine max是重要的粮食和油料作物,营养成分丰富。当前,大豆为全球供应了71%的植物蛋白和29%的植物油脂[14−16]。然而2022年中国进口大豆数量达9 108万t,对外依存度高达80%以上,大豆产需缺口高达9 000万t。因此,优化大豆株型对提高中国大豆种植密度以及提升中国大豆产量有着重要意义。IGT基因家族作为株型调控的潜在基因,在大豆中的系统鉴定和功能尚不明确。本研究利用生物信息学方法对大豆IGT基因家族(GmIGTs)进行全基因组水平鉴定,并对IGT基因家族成员进化关系、基因结构、保守基序、顺式作用元件、共线性关系及组织表达量等进行分析,以期为深入探究IGT基因家族在大豆生长发育过程中的作用机制奠定理论基础。
-
在Phytozome (
https://phytozome-next.jgi.doe.gov/ )和美国国家生物技术信息中心(NCBI)数据库(https://www.ncbi.nlm.nih.gov )对大豆‘天隆1号’‘Tianlong 1’ IGT基因家族氨基酸序列与拟南芥进行BLAST比对,确认大豆IGT基因家族成员,收集大豆基因组数据(www.soybase.org)中的大豆IGT基因家族成员遗传信息,并利用蛋白质数据库分析大豆IGT基因家族成员基本信息。在Phytozome (
https://phytozome-next.jgi.doe.gov/ )下载拟南芥以及大豆IGT基因家族氨基酸序列,对氨基酸序列进行多重比对。采用最大似然法构建进化树(MEGA 5.0软件)。 -
下载大豆基因组Phytozome (
https://phytozome-next.jgi.doe.gov/ ),使用TBtools 对IGT基因家族外显子-内含子结构分析,并绘制可视化图。同时,对大豆IGT基因家族进行蛋白质保守基序研究(MEME),利用TBtools进行可视化绘图。 -
提取大豆IGT基因家族上游2 000 bp序列,并利用PlantCARE数据库对启动子区域的顺式作用元件进行分析。随后,使用在线工具GSDS (
http://gsds.gao-lab.org/ )对结果进行可视化处理。 -
通过Phytozome (
https://phytozome-next.jgi.doe.gov/ )下载大豆的全基因组文件,利用 TBtools软件进行共线性分析,并进一步可视化。 -
通过从Phytozome (
https://phytozome-next.jgi.doe.gov/ )数据库下载的大豆在不同组织的基因表达数据,选择分生组织、根、茎、叶、花、荚、种子和根瘤等8个组织的基因表达数据进行热图分析,分析IGT基因家族在大豆中的表达情况。采用TBtools 热图软件进行数据可视化处理。采用TransZol Up Plus RNA Kit (ER501,Transgene)提取大豆的根、茎、叶、叶柄、花、果荚、种子、根瘤各部位RNA,采用Hifair® Ⅲ 1 st Strand cDNA Synthesis SuperMix for qPCR (11141ES60, 翌圣)进行反转录合成 cDNA,所得的cDNA用于检测GmIGT基因的表达水平。在Phytozome (
https://phytozome-next.jgi.doe.gov/ )上检索获得IGT基因的序列,以大豆cons4为内参基因,使用Snap Gene软件设计跨内含子的上下游引物(表1),由擎科生物技术有限公司(杭州)合成。将大豆不同组织(根、茎、叶、叶柄、花、果荚、种子、根瘤)的cDNA质量浓度定量至50 mg·L−1,按照Hieff UNICON® qPCR SYBR Green Master Mix (抗体法,No Rox,11198ES08, 翌圣)操作说明进行实时荧光定量PCR(RT-qPCR)。每个样品含3个生物学重复。利用$2^{-\Delta \Delta C{{t}}} $法计算表达水平[17]。表 1 用于RT-qPCR的引物序列
Table 1. Primer sequences for RT-qPCR
引物名称 引物序列(5′→3′) 引物名称 引物序列(5′→3′) IGT1-qRT-F TTGGTCATTGGGAATTCTTGCAA IGT9-qRT-R CAATCGCTGAATTCTTCCCT IGT1-qRT-R CTTCTACACCAGCAAAAGAGT IGT10-qRT-F GAAAGATGGGTTTGCTACAAAC IGT2-qRT-F ATGCAATTCCTCAGCTGGA IGT10-qRT-R CCGAGAGTGCCAATGGTT IGT2-qRT-R TGCTAGTAAACCATCAGGC IGT11-qRT-F TTCACACTTGGAAACCCTTGTA IGT3-qRT-F AAGTTCCTCAGCTGGATG IGT11-qRT-R CAACCCAGAGTAAGATGTTTGA IGT3-qRT-R GGCCAATCGCTGAATTC IGT12-qRT-F ATGCAGATTCTTCAATGGATCTCC IGT4-qRT-F CTGAAAGATGGGTTTGCTTCA IGT12-qRT-R CCTCCCTTCGATTGTCTTT IGT4-qRT-R GATCATAGCCAAGAGTGCCAA IGT13-qRT-F CAGTGAACAAAATGAAGGGAAAG IGT5-qRT-F TCAGTGGGAAAGAGGTAAATC IGT13-qRT-R GTTGAGGGTGCTGTAAAAGAA IGT5-qRT-R CTGAACATTAAAACCTTTCTTCCA IGT14-qRT-F CTTGGTCATTGGGAATTCTTGC IGT6-qRT-F GAATGGAAAAGATTTTGAGGGCAATAC IGT14-qRT-R CCAGCAAAAGAGTTTCTAAGGT IGT6-qRT-R AAGCTCCTCTTCATCATCATC IGT15-qRT-F GATTTCACACTTGGAAATCCCTG IGT7-qRT-F TTGATCATCGGAGGGAATTCTT IGT15-qRT-R CCCAGAGTAAGATGTTTGATTTTCC IGT7-qRT-R CTGAATCTTCCTCATACTCATCATG IGT16-qRT-F AAATGATCATGAAATAGGCCGG IGT8-qRT-F CAACTACCCATTACATGAGGC IGT16-qRT-R TCCTCCCTTCGATTGTCTTT IGT8-qRT-R GCGTGGTTACTCCTTCCA IGT17-qRT-F TTGATCATCGGAGGAGGGA IGT9-qRT-F CAATGAAGTTCCTCAGCTGGAT IGT17-qRT-R CTTCCTCATACTCATCATGATCC -
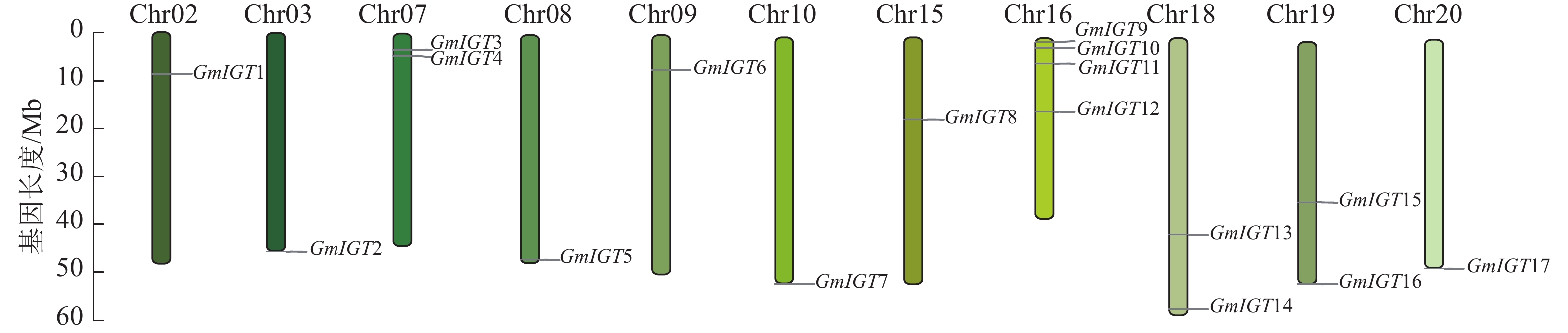
将大豆基因组数据库与拟南芥序列进行比对,识别大豆中的同源基因。获得比对结果后,删除重复序列,以确保结果的准确性和可靠性。随后,对筛选后的序列进行Pfam分析,进一步验证其作为大豆IGT基因家族的潜在候选。最终成功鉴定出17个大豆IGT基因,并按照其在染色体上的顺序命名为GmIGT1~GmIGT17(图1,表2)。
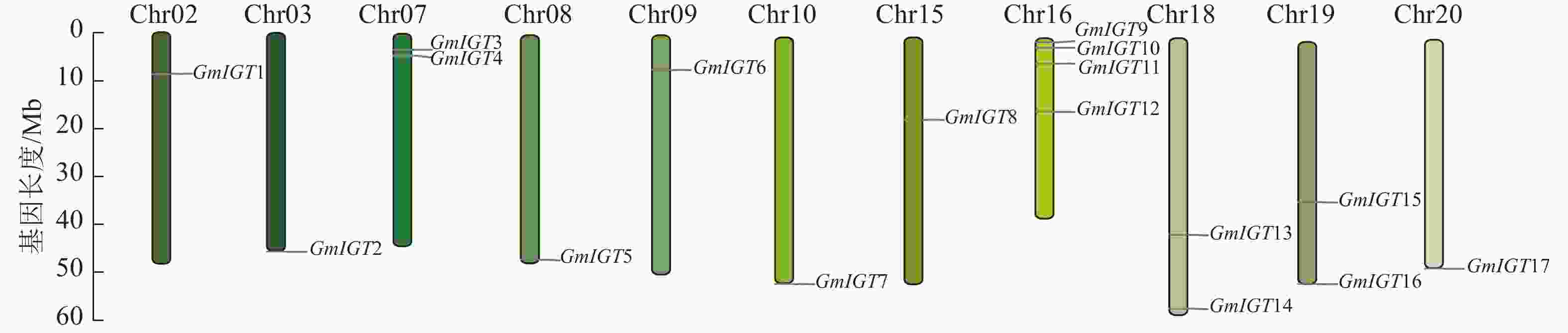
图 1 大豆IGT基因家族的染色体定位
Figure 1. Chromosome mapping of soybean IGT gene family
表 2 大豆IGT基因家族成员特征
Table 2. Properties of members of the IGT gene family in soybean
基因名称 序列号 染色体 位置 碱基对/bp 预测蛋白质长度/个 理论等电点 分子量/Da 亚细胞定位 GmIGT1 Glyma.02G097100 Chr02 8677685~8680757 1 101 367 6.91 40 326.05 细胞核 GmIGT2 Glyma.03G264200 Chr03 46860193~46864806 822 274 5.52 31 485.80 细胞核 GmIGT3 Glyma.07G040800 Chr07 3391340~3394088 846 282 7.58 32 278.71 叶绿体 GmIGT4 Glyma.07G052500 Chr07 4555925~4560419 858 286 5.45 32 189.21 细胞核 GmIGT5 Glyma.08G356200 Chr08 46252966~46256085 852 284 10.03 32 316.57 细胞核 GmIGT6 Glyma.09G071100 Chr09 7218596~7221206 789 263 5.11 29 050.36 细胞核 GmIGT7 Glyma.10G298100 Chr10 51546352~51548699 1068 356 5.96 39 907.67 细胞质 GmIGT8 Glyma.15G179200 Chr15 17222927~17225610 789 263 5.12 29 253.52 细胞核 GmIGT9 Glyma.16G009400 Chr16 842813~845650 855 285 8.03 32 507.05 细胞核 GmIGT10 Glyma.16G021400 Chr16 2005820~2009925 870 290 5.45 32 743.91 细胞质 GmIGT11 Glyma.16G054300 Chr16 5286678~5290902 1 149 383 6.65 43 362.59 叶绿体 GmIGT12 Glyma.16G094000 Chr16 15479890~15482382 786 262 9.67 30 379.48 细胞核 GmIGT13 Glyma.18G173300 Chr18 41543428~41546170 861 287 9.92 32 629.87 细胞核 GmIGT14 Glyma.18G284400 Chr18 56775415~56778618 1 098 366 6.91 40 390.19 细胞核 GmIGT15 Glyma.19G094600 Chr19 33952844~33956542 1 149 383 7.18 43 382.51 细胞核 GmIGT16 Glyma.19G263200 Chr19 51104292~51108788 807 269 5.68 30 957.17 细胞核 GmIGT17 Glyma.20G249200 Chr20 47749661~47752260 1 080 360 5.87 40 051.88 细胞核 GmIGT的cDNA长度为786~1 149 bp,有明显差异。它们所编码的蛋白质长度为263~384个氨基酸,其理论等电点为5.11~10.03。在这些蛋白质中,有6个的等电点超过7,11个的等电点低于7。这表明该基因家族具有多样性。亚细胞定位进一步揭示了这些蛋白的潜在功能。预测发现:GmIGT3与GmIGT11位于叶绿体,GmIGT10定位于细胞质,其余14个GmIGTs均定位于细胞核(表2)。
GmIGT的17个成员分布在11条染色体上,分别为Chr02、Chr03、Chr07、Chr08、Chr09、Chr10、Chr15、Chr16、Chr18、Chr19、Chr20,其中有4个成员位于Chr16上(图1)。
由IGT基因家族鉴定与理化性质分析及染色体位置分析可知:IGT基因家族在序列长度、氨基酸个数、等电点、亚细胞定位与染色体分布各方面均有差异。
-
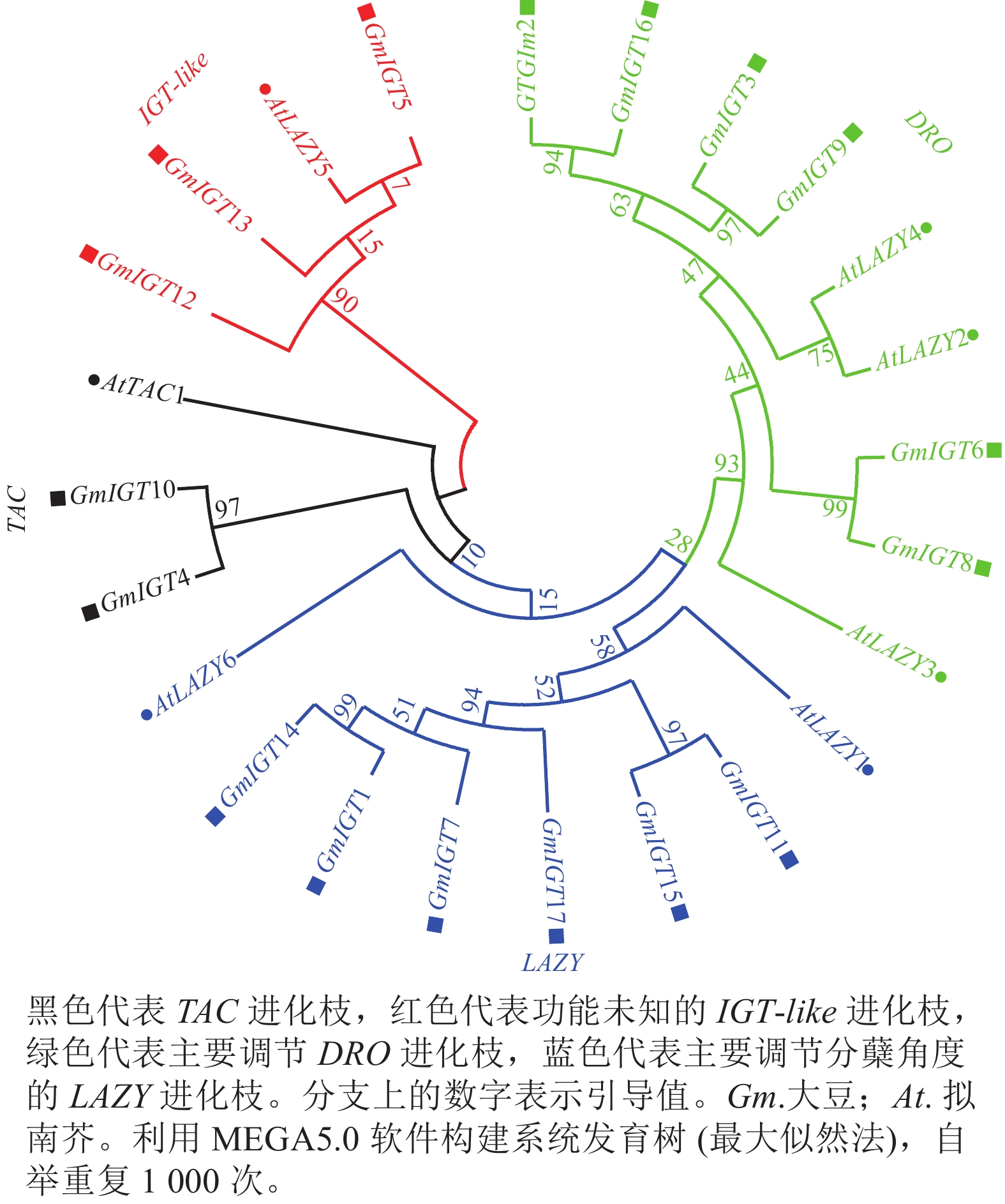
对大豆IGT基因家族进行系统进化树分析,结果(图2)显示:IGT基因家族可以分为4个进化枝,即TAC进化枝、IGT-like进化枝、DRO进化枝、LAZY进化枝。每个进化枝包含了2~6个大豆IGT基因家族成员。GmIGT4/10与AtTAC1同源,属于TAC进化枝;GmIGT5/12/13与AtLAZY5同源,属于功能未知的进化枝;GmIGT2/3/6/8/9/16与AtLAZY2/3/4同源,属于DRO进化枝;GmIGT1/4/7/11/15/17与AtLAZY1同源,属于主要调节向地性的进化枝。该结果表明:IGT基因家族成员可能在进化上存在功能分化。
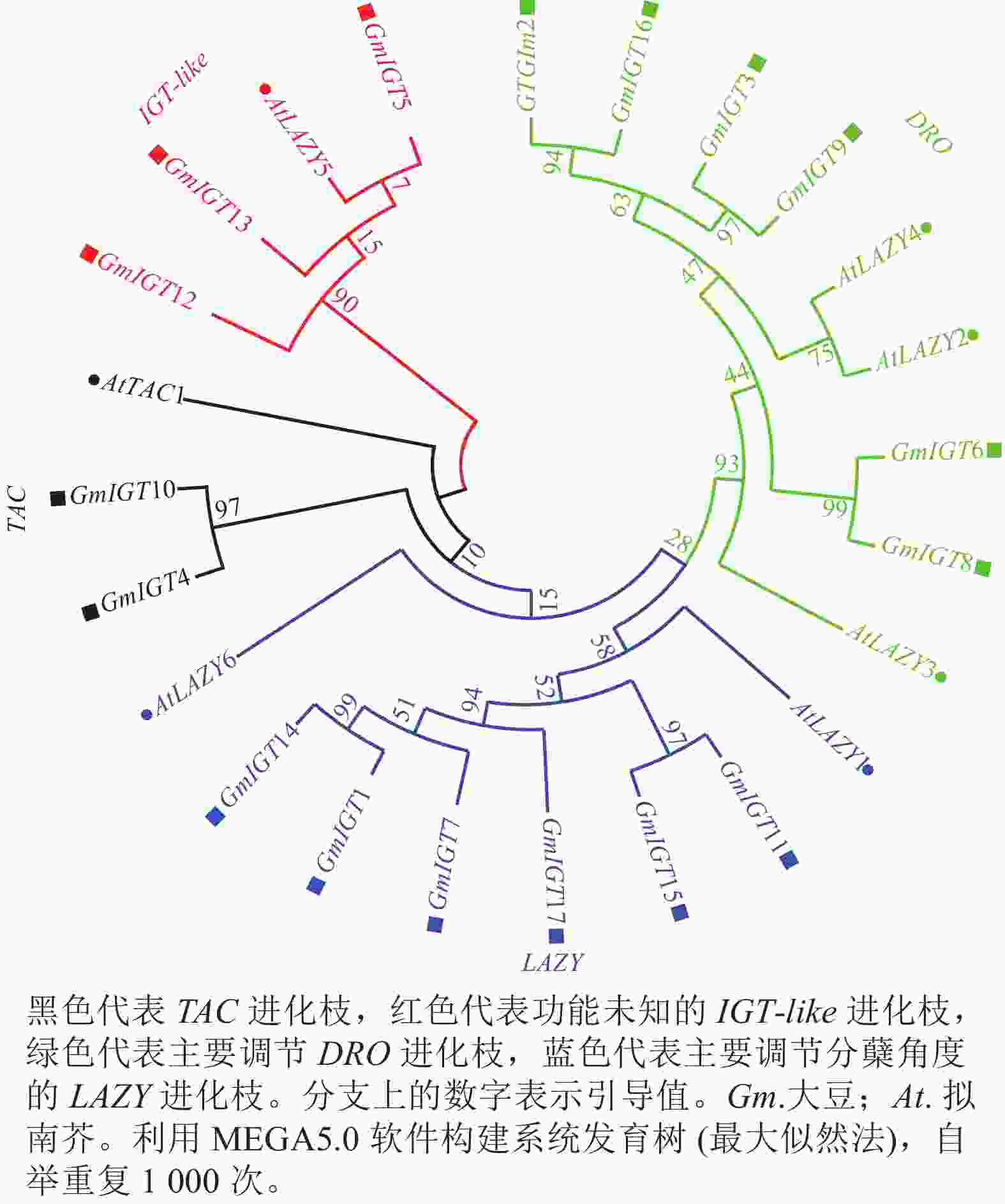
图 2 IGT基因家族系统进化树分析
Figure 2. Phylogenetic tree analysis of the IGT gene family
-
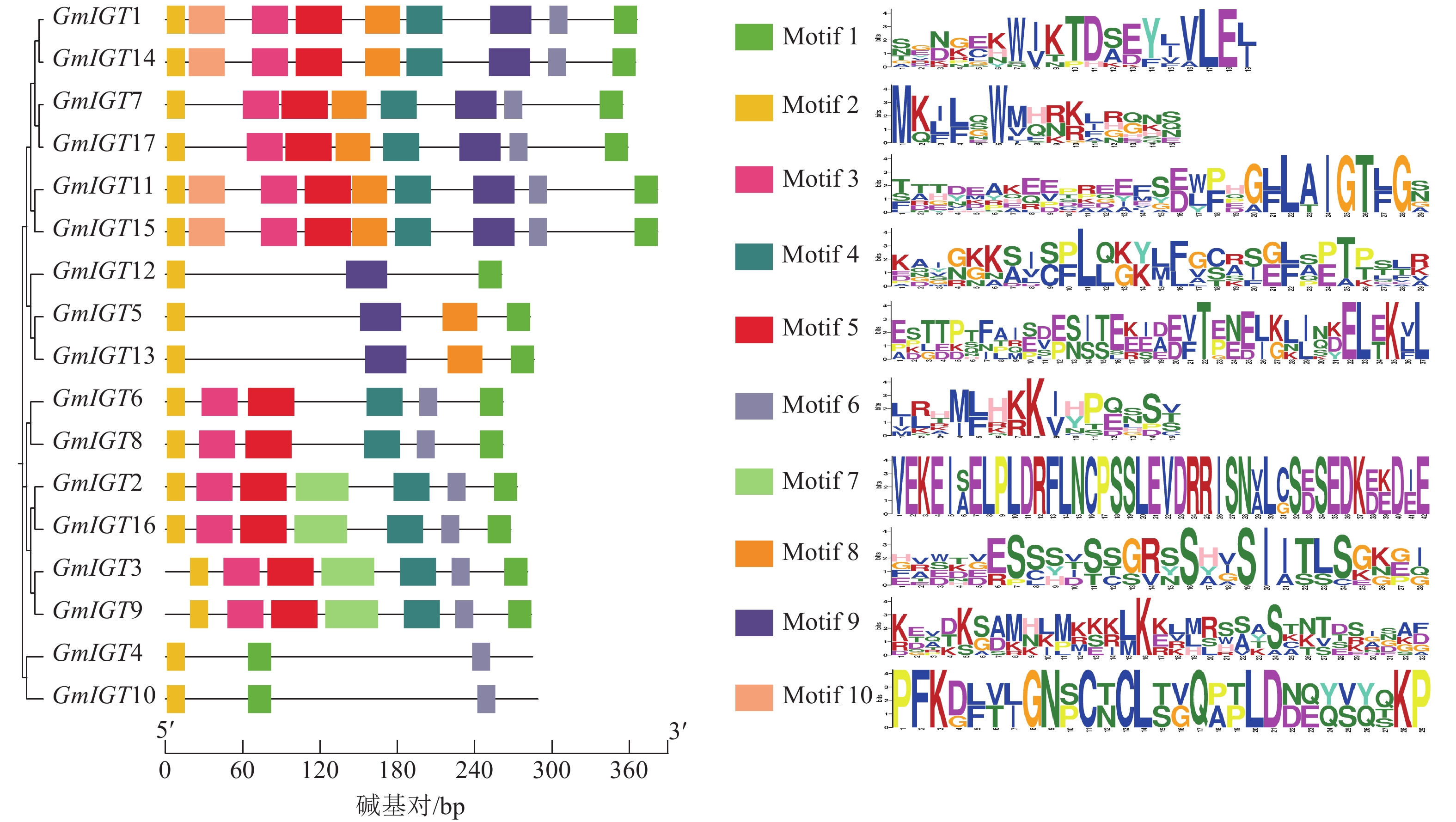
为了进一步探究大豆IGT基因家族功能,绘制了IGT基因家族的结构模型(图3)。由图3可知:GmIGT基因家族成员基因结构简单,其外显子数目仅3~5个,这表明该基因家族在基因结构上具有一定的保守性。尽管在结构上存在保守性,但也存在长度上的差异,这些差异可能源于基因组中的多态性和突变,也可能反映了这些基因在进化过程中的功能细分。此外,家族中有3个基因不存在UTR区域。这些结构上的差异可能暗示GmIGT基因成员在功能或表达调控上存在差异,为进一步的功能试验和机制研究提供了方向。大豆IGT基因家族展现出高度保守的基因结构,这表明了IGT基因家族在功能上的相似性较高,仅功能细分上存在差异。
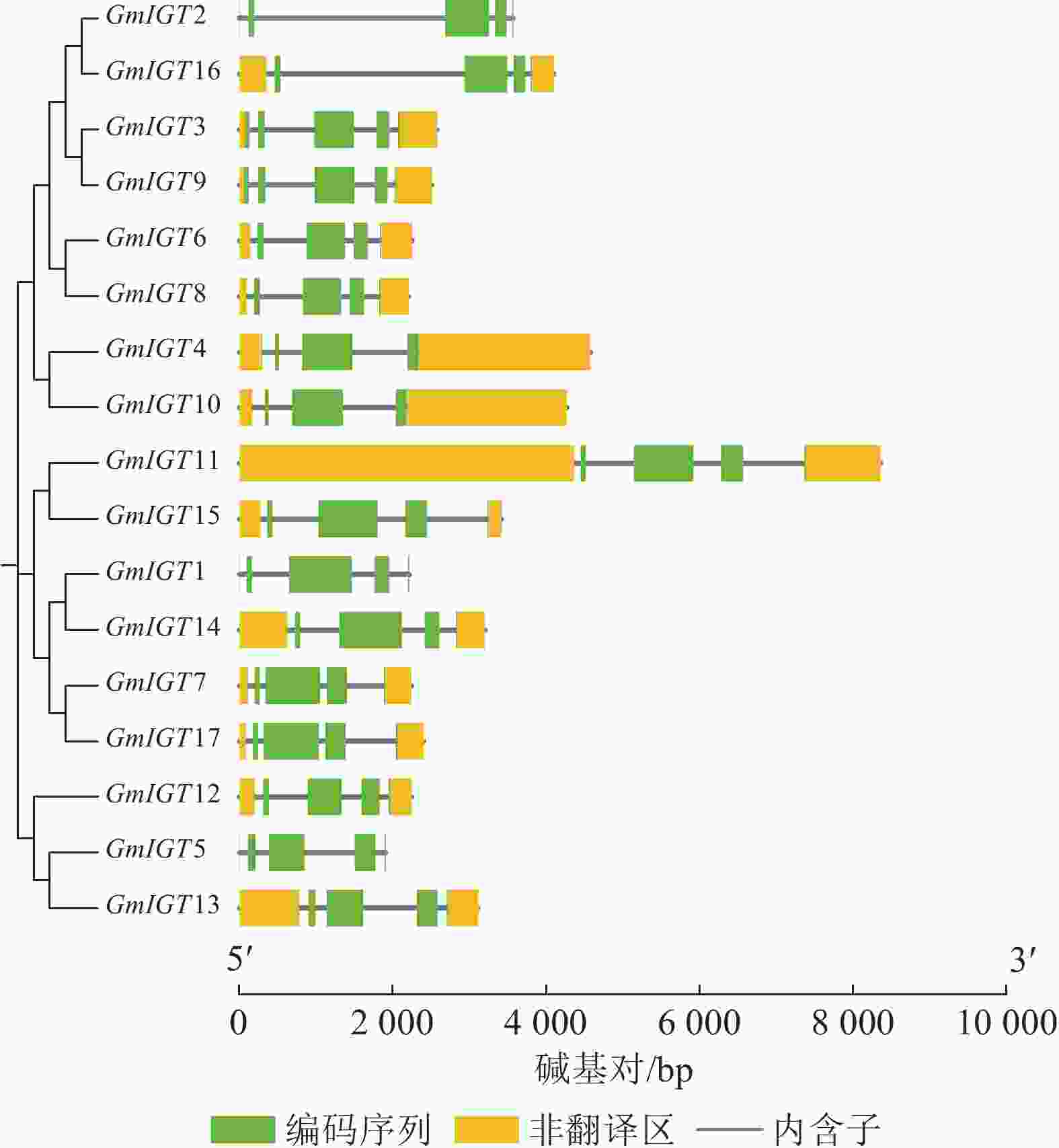
图 3 GmIGT基因的结构分析
Figure 3. Structure analysis of the GmIGT genes
-
为了深入了解GmIGT成员蛋白保守基序组成,通过MEME软件对其保守基序进行预测(图4)。大豆IGT基因家族包含10个保守基序,Motif 3含有家族特有的GφL(A/T)IGT序列,该序列在除GmIGT4/5/10/12/13以外的所有GmIGT蛋白中都存在。所有GmIGT的N末端都含有Motif 2。属于LAZY进化枝的GmIGT1/7/11/14/15/17,都具有Motif 1、Motif 2、Motif 3、Motif 4、Motif 5、Motif 6、Motif 8、Motif 9,其中GmIGT1/11/14/15具有特异性的Motif 10;属于IGT-like进化枝的GmIGT5/12/13,都具有Motif 1、Motif 2、Motif 9,其中GmIGT5/13具有特异性的Motif 8;属于DRO进化枝的GmIGT2/3/6/8/9/16,都具有Motif 1、Motif 2、Motif 3、Motif 4、Motif 5、Motif 6,其中GmIGT2/3/9/16具有特异性的Motif 7;属于TAC进化枝的GmIGT4/10,都具有Motif 1、Motif 2、Motif 6。

图 4 不同GmIGT蛋白质之间的保守基序分析
Figure 4. Analysis of conserved motifs occurring in different GmIGT proteins
结果表明:大豆IGT基因家族保守基序分布、基因结构和系统发育有相关性。相比基因结构,GmIGT的保守基序的相似度比较小,但同属同一进化枝的保守基序的相似度高,除了TAC进化枝,LAZY、IGT-like、DRO进化枝的成员并不都具有完全相同的Motif,这可能意味着其进化枝上的基因在功能上存在差异。这种基序的差异可能反映了这些进化枝在生物学功能或调控机制上的不同。
-
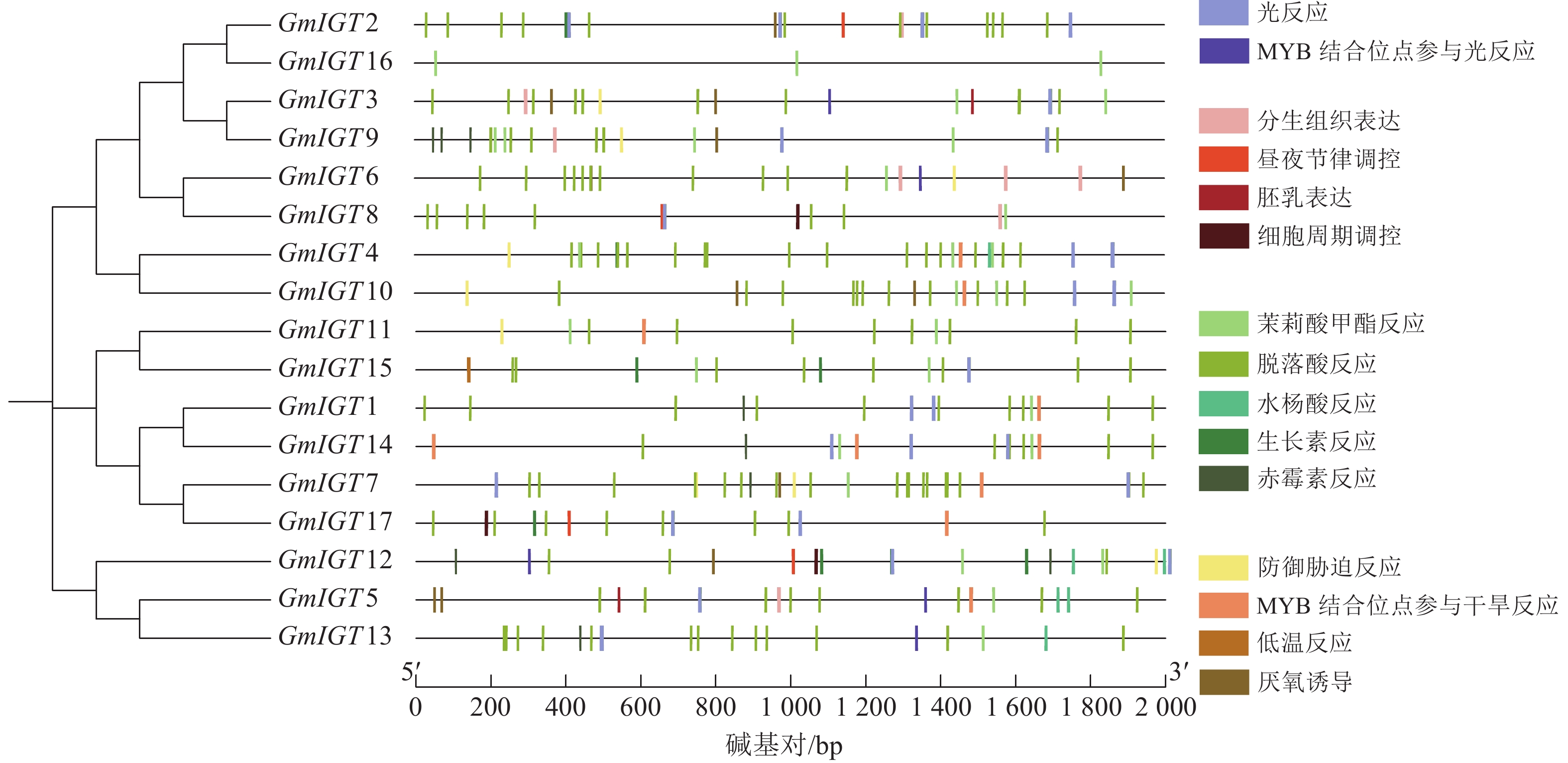
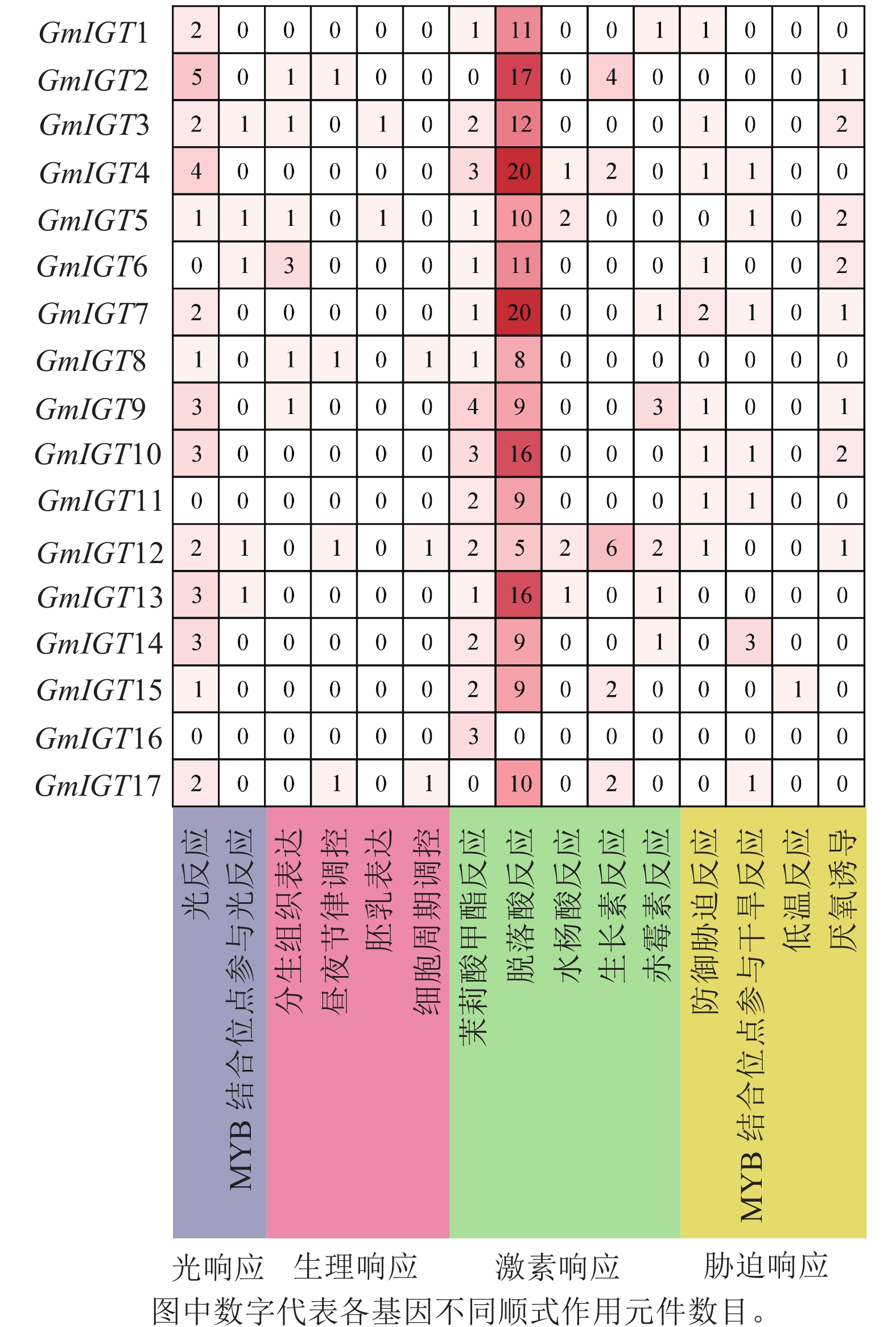
为了进一步了解大豆IGT基因家族的潜在功能和调控机制,使用PlantCARE对翻译起始位点上游近2 000 bp的序列区域进行顺式作用元件分析。结果显示:几乎所有的IGT基因家族成员在启动子区域都有至少3个以上顺式作用元件,GmIGT4含有的元件数最多,为32个,其次是GmIGT2,含有 29个顺式作用元件,GmIGT16仅包含3个顺式作用元件(图5)。共检测到15种顺式调控元件,将它们分为4个大类:光响应、生理响应、植物激素响应和胁迫响应。第 1 类是光响应元件,包括光响应与MYB结合位点参与光响应。第2类是生理响应元件,包括分生组织表达、昼夜节律调控、胚乳表达等。第3类是植物激素响应元件,由茉莉酸甲酯反应、脱落酸反应等组成,其中脱落酸反应元件是在GmIGT基因成员启动子中最丰富的顺式作用元件,而GmIGT4与茉莉酸甲酯、脱落酸、生长素、赤霉素响应有关。推测GmIGT4可能参与多种激素响应。第4类是非生物与生物胁迫相关的顺式作用元件,包括防御胁迫反应、低温反应等,值得注意的是,GmIGT8/13/16基因中不包含任何与非生物或生物胁迫相关的元件。在4类顺式作用元件中,激素响应元件最多,占比74.12%,光响应元件排第2位(11.47%),胁迫响应元件为9.41%,参与植物生理响应最少,占5.00%(图6)。这些结果表明:GmIGT基因可能与光响应、生长发育、激素和胁迫响应有关。
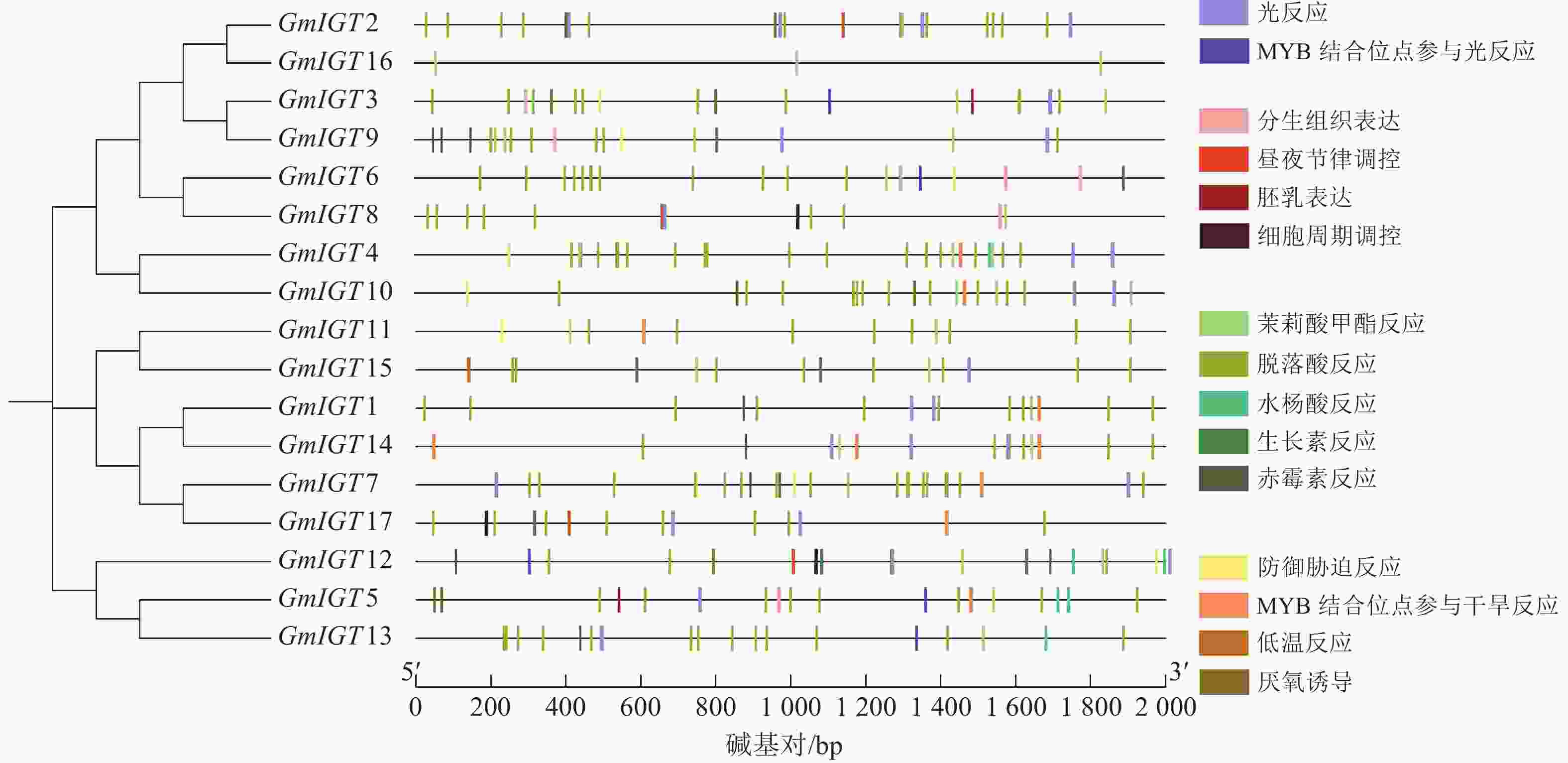
图 5 GmIGT基因启动子区域顺式作用元件分析
Figure 5. Analysis of cis-elements in the promoters of the GmIGT genes
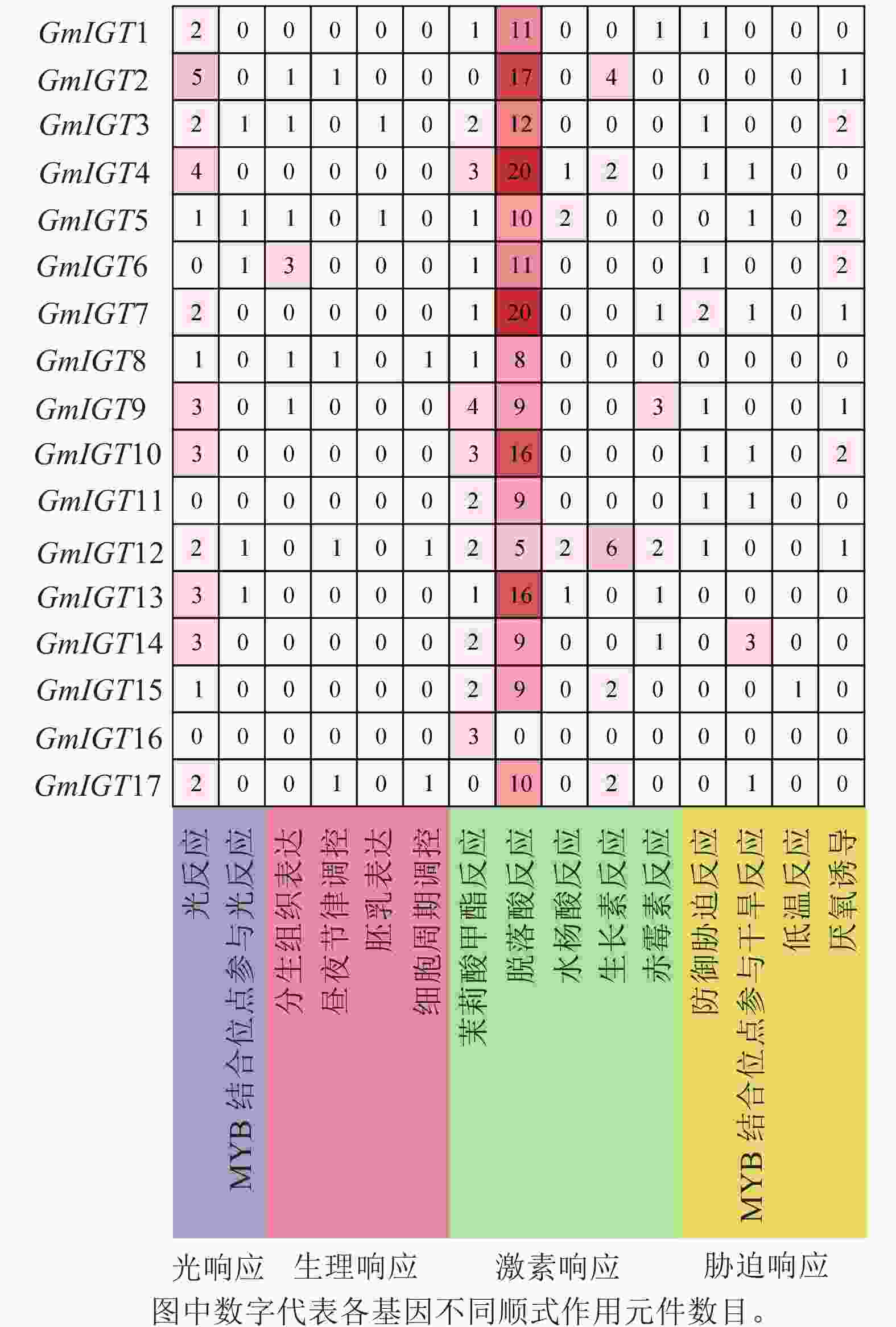
图 6 GmIGT基因启动子区域顺式作用元件数量统计分析
Figure 6. Statistical analysis of number of cis-acting elements in the promoters of the GmIGT genes
-
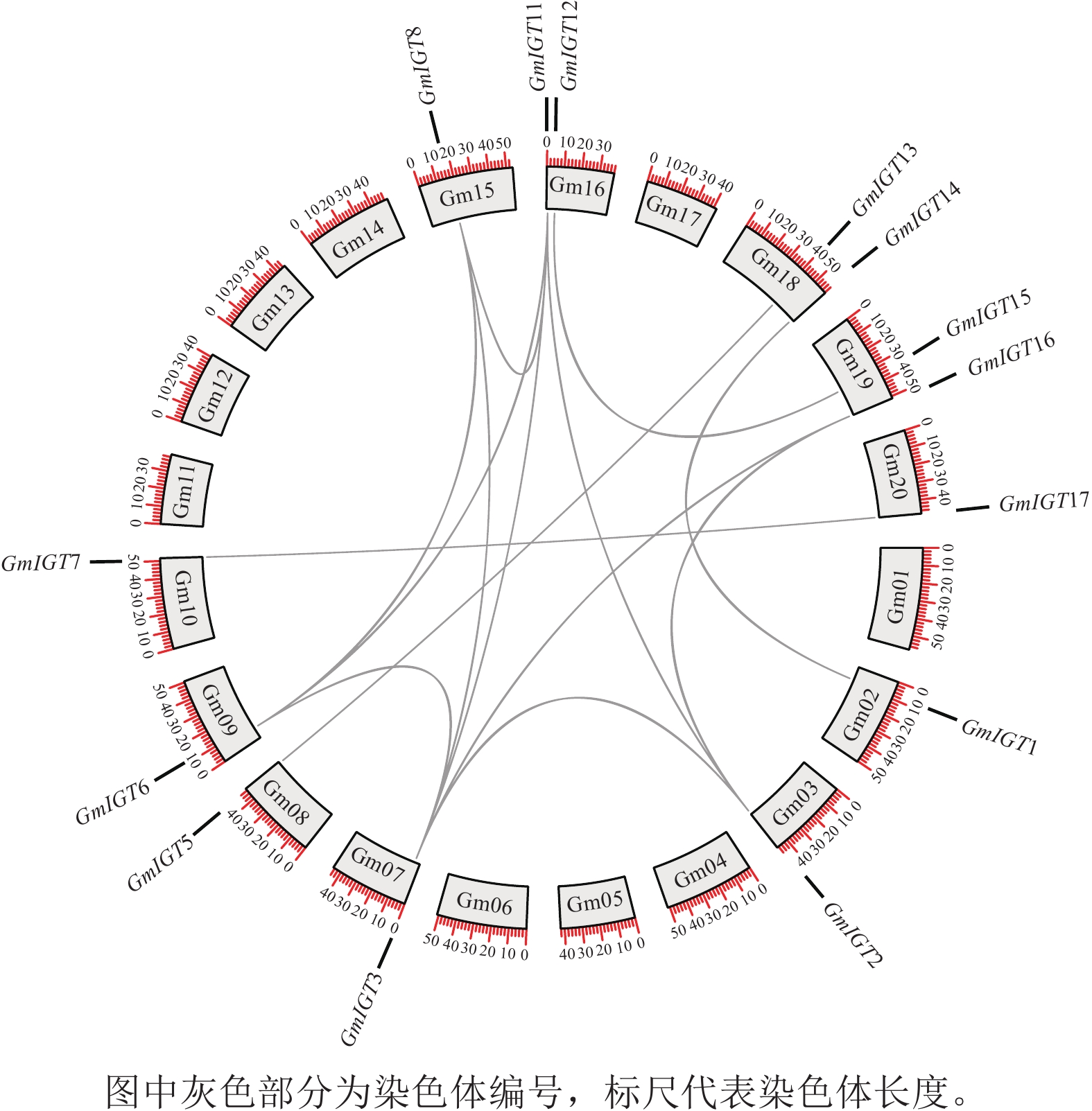
GmIGT基因遍布于11条染色体上(图1)。为了解其扩增机制,基因共线性分析揭示:除了GmIGT4/9/10以外,其余GmIGT基因均参与了片段复制事件(图7)。其中有14条片段复制事件(GmIGT1-GmIGT14、GmIGT2-GmIGT3/12/16、GmIGT3-GmIGT6/8/12/16、GmIGT5-GmIGT13、GmIGT6-GmIGT8/12、GmIGT7-GmIGT17、GmIGT8-GmIGT12、GmIGT11-GmIGT15),GmIGT2、GmIGT3、GmIGT6、GmIGT8、GmIGT12、GmIGT16之间存在1个或多个共同的复制源 。这表明染色体或基因组经历了多次或层叠的片段复制事件,导致产生特定基因或基因区域的多重复制品。
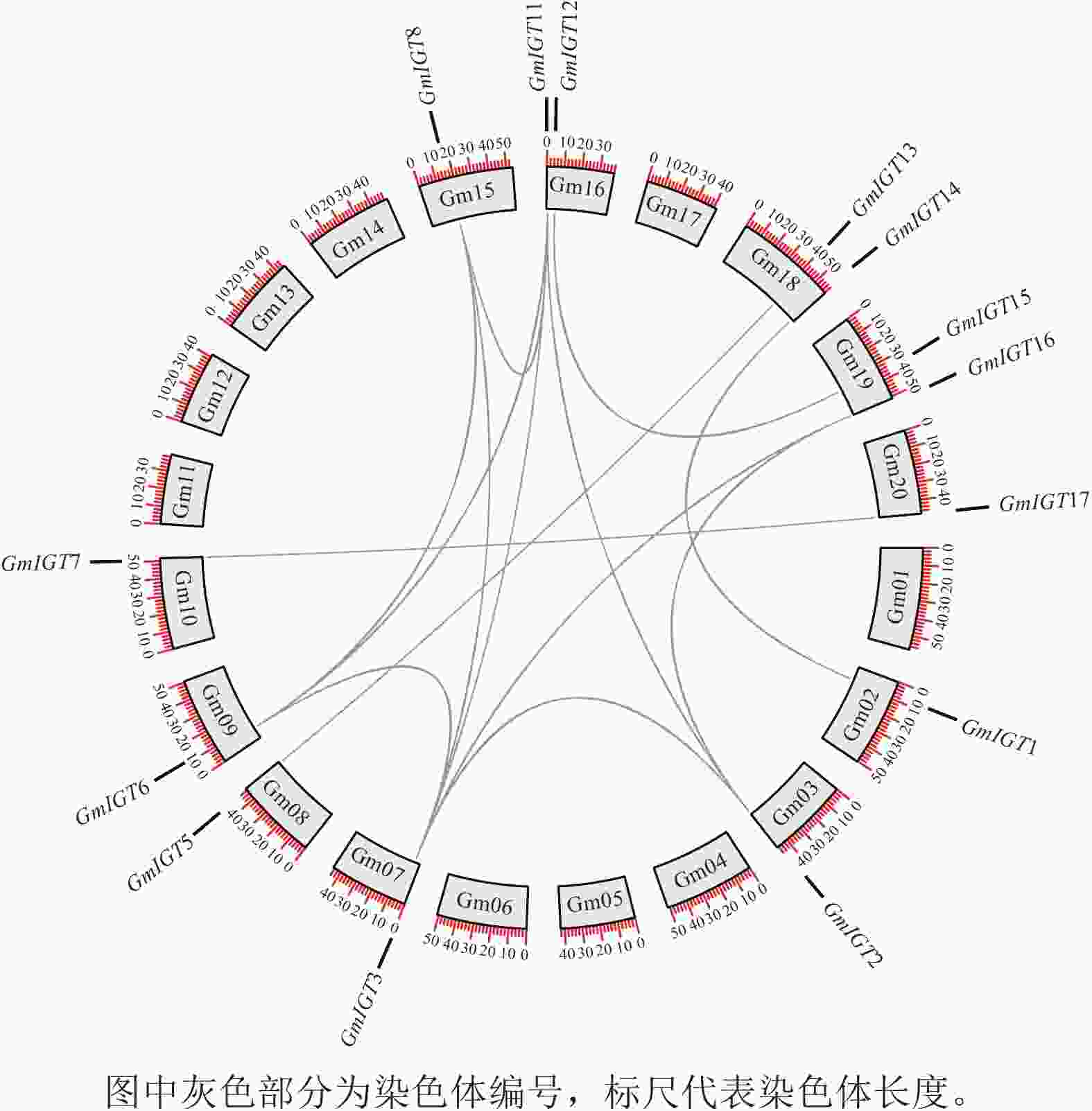
图 7 GmIGT基因共线性分析
Figure 7. Collinearity analysis of the GmIGT genes
-
为探索GmIGT基因在不同时期以及组织部位的表达模式,借助大豆Phytozome(
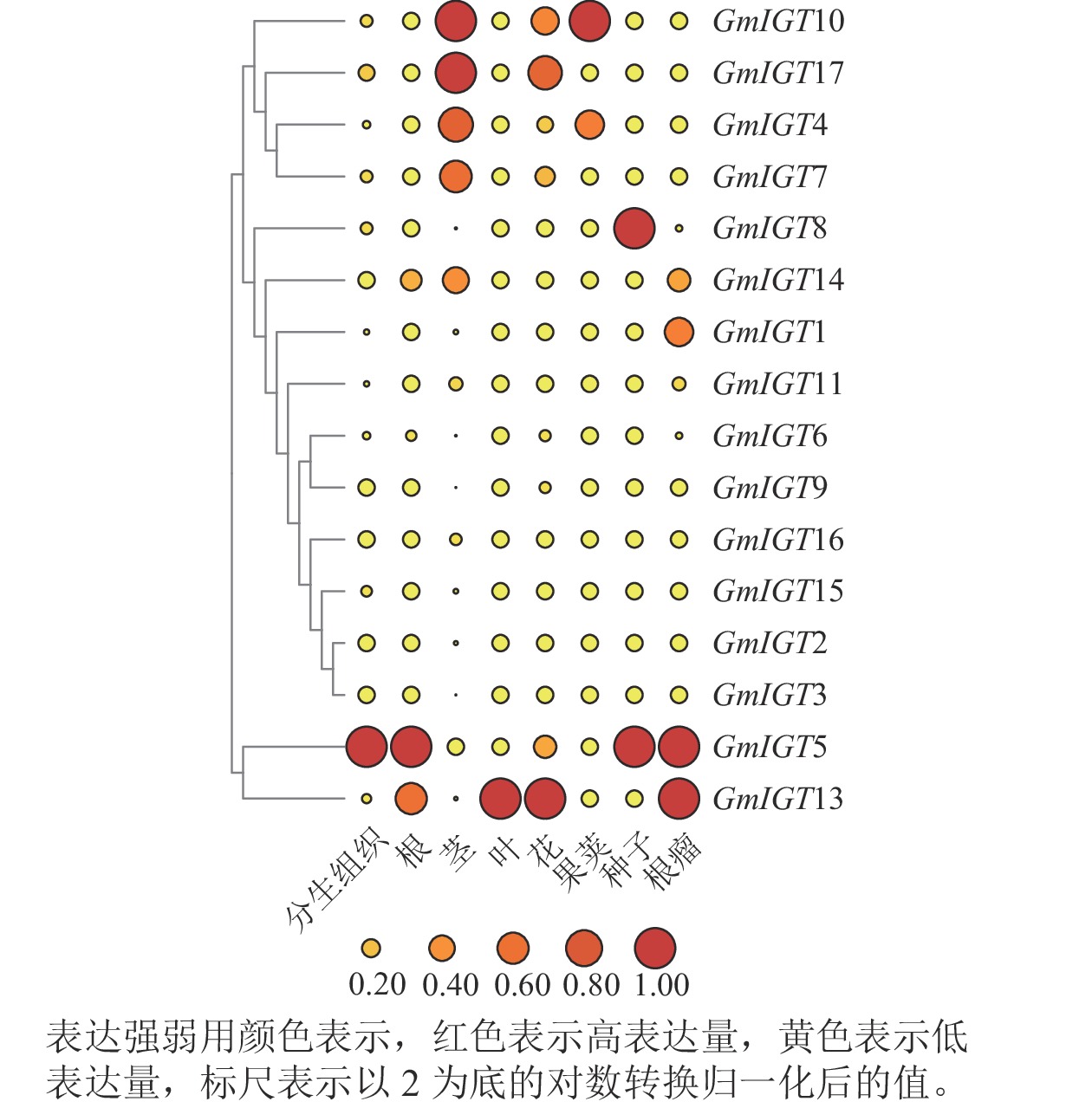
https://phytozome-next.jgi.doe.gov/ )数据,对17个GmIGTs在大豆的分生组织、根、茎、叶、花、荚、种子与根瘤的表达量进行计算机分析。结果表明:GmIGTs基因在这8个组织中显示出特异性的表达模式。具体而言,GmIGT5和GmIGT13在不同组织中都表现出较高的表达水平。在分生组织中,GmIGT5的表达高;在根中,GmIGT5和GmIGT13表现出高表达水平;在茎中,GmIGT4、GmIGT7、GmIGT10、GmIGT14与GmIGT17的表达高;在叶中,GmIGT13的表达量高;在花中,GmIGT17的表达水平高;而在荚中,GmIGT10表达水平高;在种子中,GmIGT8和GmIGT5的表达高;在根瘤中,GmIGT1、GmIGT14、GmIGT5和GmIGT13具有高表达(图8)。IGT基因家族在大豆不同组织中的表达差异较大,说明IGT基因家族可能多方面参与大豆生长发育调控。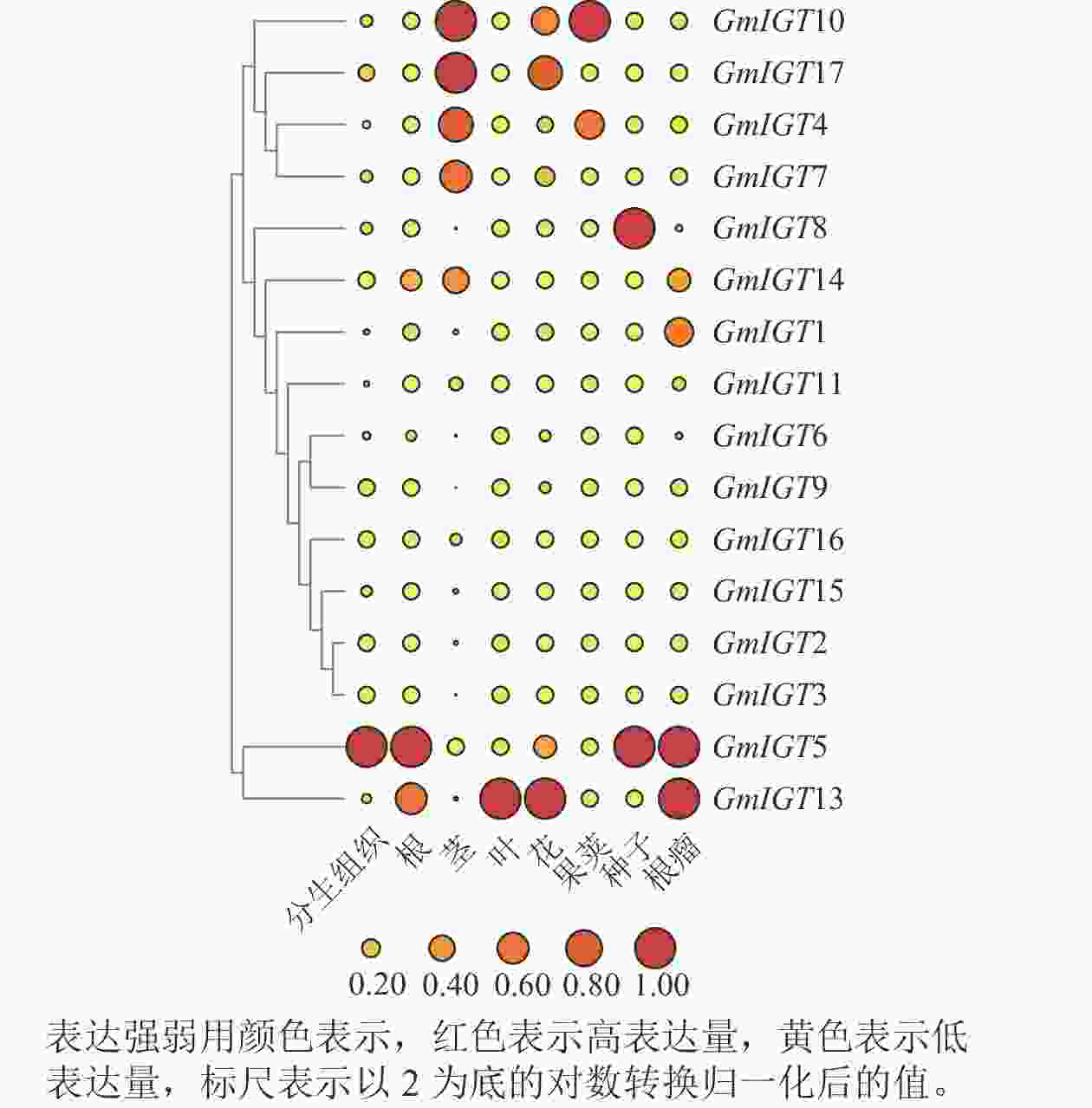
图 8 GmIGT在不同组织中表达模式的计算机分析
Figure 8. In silico analysis of expression patterns of different GmIGT genes in different tissues
-
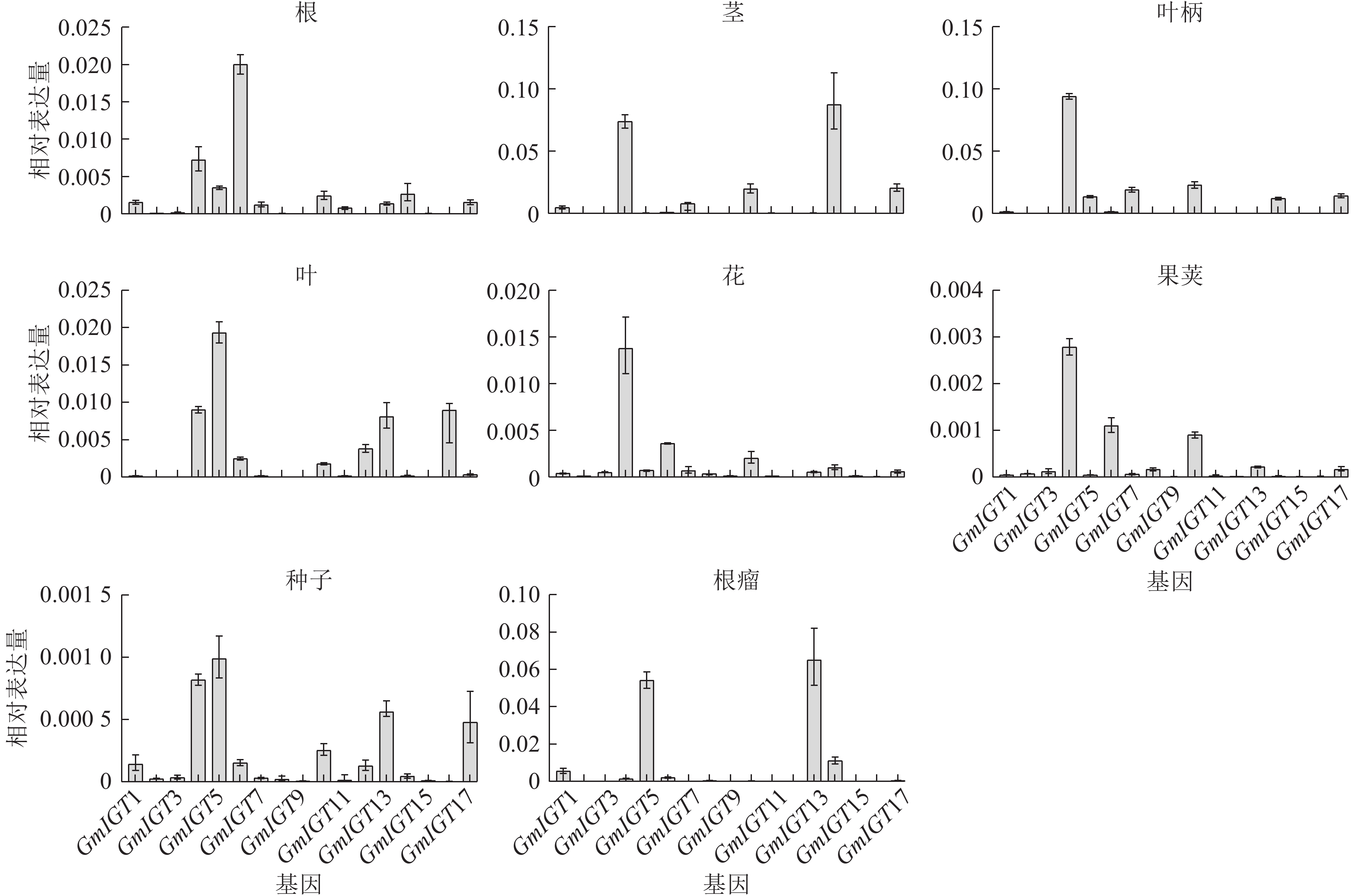
为了进一步验证GmIGT在大豆中的时空表达模式,对17个GmIGTs在大豆不同组织中的表达量进行RT-qPCR分析 (图9)。结果显示:GmIGTs基因在8个组织中表现出特异性的表达模式。具体而言,GmIGT4在除根瘤外的其他组织中都表现出较高的表达水平。在根中,GmIGT6的相对表达量最高;在茎中,GmIGT4、GmIGT10、GmIGT14与GmIGT17的相对表达量较高;在叶柄中,GmIGT4、GmIGT7、GmIGT10与GmIGT17的相对表达量较高;在叶中,GmIGT5的表达量高;在花中,GmIGT4与GmIGT6的表达水平高;在荚中,GmIGT4与GmIGT6表达水平高;在种子中,GmIGT4、GmIGT5、GmIGT13和GmIGT17的表达高;在根瘤中,GmIGT5和GmIGT13具有高表达。GmIGT4、GmIGT10与GmIGT17均在茎与叶柄中高度表达,说明其可能在塑造大豆株型中发挥作用。综上所述,IGT基因家族可能在多方面参与大豆生长发育调控,并且与生物信息学预测基本一致。
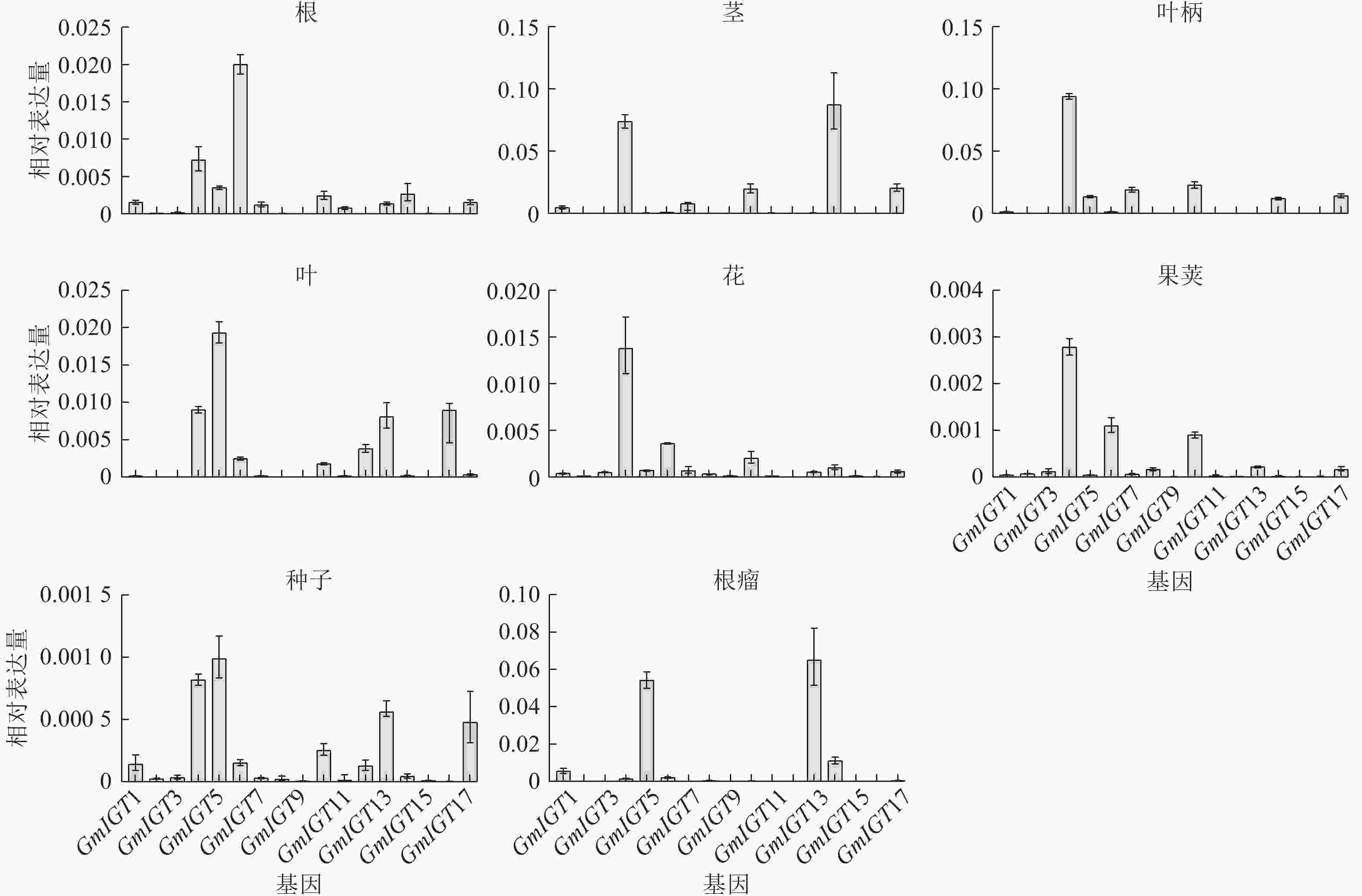
图 9 GmIGT基因家族成员在不同组织中的相对表达水平
Figure 9. Relative expression levels of members of the GmIGT gene family in different tissues of soybean
-
IGT基因家族在重力感应中起着关键作用,因此推测该基因家族参与植物株型的调控。IGT基因家族广泛存在于植物基因组中,且在不同物种中的数量差距较大。目前,已在水稻、玉米、油菜Brassica campestris和苹果Malus domestica中分别鉴定出4、2、27和4个IGT基因家族成员[1,18−20]。本研究在深入分析大豆基因组基础上,鉴定得到17个大豆IGT基因家族成员,它们不均匀分布在11条染色体上。其基因总数与其他物种中IGT基因家族成员数目有所差异,这可能与不同物种间基因组大小、基因扩张程度不同有关。基因组内共线性分结果表明:GmIGT复制类型主要为片段性复制,有14条片段复制事件,这说明片段性复制在GmIGT基因扩张中起了至关重要的作用。这些家族成员的扩增可能会导致不同成员产生功能冗余性或专化性,从而为生物过程的精细调控提供基础。
在拟南芥中,IGT基因家族存在5个较短的保守结构域和1个独特的内含子排列[1],GmIGT基因中也出现了类似的内含子排列。外显子和内含子排列可在一定程度上解释该基因家族的进化史。在很多情况下,不同物种中具有相似基因结构和保守基序的同源蛋白可能具有相同或相似的功能。由此推测,犹如拟南芥AtIGT基因,大豆GmIGT可能参与大豆的重力传感和植物株型的形成。
前人研究表明:LAZY基因往往在拟南芥、水稻、蒺藜Tribulus terrestris和日本百里香Thymus vulgaris等高等植物中形成1个小基因家族[3, 21]。在拟南芥中,该基因家族有6个成员,不同成员在不同器官中的功能可能存在差异,已知AtLAZY1调节芽和花序茎的向地性[3, 10],但AtLAZY5、AtLAZY6在向地性方面的功能尚未见报道。在许多植物中,TAC在调节分枝角度方面起着重要作用,OsTAC1参与水稻分蘖角度的调节,并影响内源生长素的含量[22]。在玉米中,ZmTAC1的表达水平与叶片角度呈正相关,该基因在叶片发育中起着重要的调控作用[23]。系统发育分析显示:IGT基因家族在大豆中可以分为4个进化枝,TAC、IGT-like、DRO以及LAZY进化枝。同时,LAZY、IGT-like、DRO进化枝的成员并不都具有完全相同的Motif,这可能意味着其进化枝上的基因在功能上存在差异,这种基序的差异可能反映了这些进化枝在生物学功能或调控机制上的不同。因此,根据聚类结果,大豆的IGT基因家族可能在不同的器官中起着不同的作用,但其作用有待进一步研究。
根据启动子预测结果,IGT基因家族存在光响应、生长发育、激素响应及胁迫响应元件,表明其可能受到光照、植物激素的调控诱导或参与其信号反应途径,同时还可能参与大豆生长发育的某些过程以及参与胁迫过程中的抗逆调控。
通过数据库预测,GmIGT基因成员在不同的组织和器官中显示出特异性的表达模式, RT-qPCR分析结果与这些基因表达模式的预测结果基本一致。组织特异性表达分析发现:整体相对表达量最高的为GmIGT4,推测该基因可能在很多组织、器官中发挥作用;GmIGT4、GmIGT10和GmIGT17在茎和叶柄中的相对表达量显著高于其他组织,推测其可能在大豆株型调控方面发挥重要作用。不同IGT基因家族成员在大豆不同组织中表达模式的差异暗示了其可能在根的向地性调节、茎的生长发育等方面存在功能上的分化,这为进一步解析GmIGT基因在大豆生长发育和株型形成中的作用奠定基础。
-
本研究对大豆IGT基因家族从全基因组层面进行了全面的生物信息学分析,共鉴定出17个GmIGT,分为TAC、IGT-like、DRO以及LAZY等4个进化枝。表达模式分析显示:GmIGT基因成员在大豆不同组织中表达模式具有差异性,GmIGT4、GmIGT10和GmIGT17可能在塑造大豆株型中发挥主要作用。该结果为进一步了解GmIGT基因在塑造植物株型中的生物学功能提供了理论基础。
Genome-wide identification and tissue expression analysis of IGT gene family in soybean
-
摘要:
目的 利用生物信息学方法对大豆‘天隆1号’Glycine max‘Tianlong 1’ IGT基因家族进行全基因组鉴定,探究IGT基因家族在大豆中的潜在功能。 方法 通过对大豆‘天隆1号’IGT蛋白结构域进行BLASTP搜索,确定GmIGTs;以生物信息学方法分析其进化关系、基因结构、保守基序、顺式作用元件、共线性关系,以及在不同组织部位表达模式。 结果 共鉴定出17个GmIGT基因,并根据其系统发育关系将其分为4个分支:TAC、IGT-like、DRO和LAZY。蛋白质保守基序分析发现: ‘天隆1号’IGT蛋白均含有Motif2。染色体定位和共线性分析显示:GmIGT不均匀地定位在11条染色体上,并且片段复制可能在GmIGT基因的扩张中发挥了重要作用。顺式作用元件分析表明:GmIGT的表达可能与光响应、生理响应、植物激素响应和胁迫等相关。此外,实时荧光定量聚合酶链式反应分析表明:GmIGT基因具有明显的组织特异表达特点,GmIGT4在所有组织中表达量都相对较高,GmIGT4、GmIGT10与GmIGT17在茎与叶柄中高度表达。 结论 GmIGT基因可能在塑造大豆‘天隆1号’株型中发挥某些潜在作用,GmIGT4、GmIGT10与GmIGT17可能为该过程中的核心基因。图9表2参23 Abstract:Objective This study aims to employ bioinformatics methods to perform a comprehensive genome-wide identification of the soybean IGT gene family and explore the potential functions of IGT gene family in soybean. Method Soybean cultivar Glycine max ‘Tianlong 1’ was selected and GmIGTs were identified by BLASTP search of IGT protein domain. The evolutionary relationships, gene structure, conserved motifs, cis-acting elements, and collinearity relationships were analyzed by bioinformatics methods. The expression patterns of GmIGTs in different tissue parts were analyzed. Result A total of 17 GmIGT genes were identified and classified into 4 branches based on their phylogenetic relationships: TAC, IGT-like, DRO, and LAZY. Protein conserved motif analysis revealed that all IGT proteins contained Motif2. Chromosomal localization and collinearity analysis showed that GmIGT genes were unevenly distributed on 11 chromosomes, and segmental duplication might have played a significant role in the expansion of GmIGT gene. Cis-acting element analysis indicated that GmIGT expression might be associated with light response, physiological response, plant hormone response and stress. RT-qPCR analysis demonstrated that GmIGT gene had obvious tissue-specific expression characteristics. Among them, GmIGT5 exhibited relatively high expression levels in all tissues, while GmIGT4 and GmIGT10 were highly expressed in stems and petioles. Conclusion GmIGT gene may play a potential role in shaping soybean plant architecture, and GmIGT4 and GmIGT10 may be the core genes involved in this process. [Ch, 9 fig. 2 tab. 23 ref.] -
图 4 不同GmIGT蛋白质之间的保守基序分析
Figure 4 Analysis of conserved motifs occurring in different GmIGT proteins
图 5 GmIGT基因启动子区域顺式作用元件分析
Figure 5 Analysis of cis-elements in the promoters of the GmIGT genes
图 6 GmIGT基因启动子区域顺式作用元件数量统计分析
Figure 6 Statistical analysis of number of cis-acting elements in the promoters of the GmIGT genes
图 8 GmIGT在不同组织中表达模式的计算机分析
Figure 8 In silico analysis of expression patterns of different GmIGT genes in different tissues
图 9 GmIGT基因家族成员在不同组织中的相对表达水平
Figure 9 Relative expression levels of members of the GmIGT gene family in different tissues of soybean
表 1 用于RT-qPCR的引物序列
Table 1. Primer sequences for RT-qPCR
引物名称 引物序列(5′→3′) 引物名称 引物序列(5′→3′) IGT1-qRT-F TTGGTCATTGGGAATTCTTGCAA IGT9-qRT-R CAATCGCTGAATTCTTCCCT IGT1-qRT-R CTTCTACACCAGCAAAAGAGT IGT10-qRT-F GAAAGATGGGTTTGCTACAAAC IGT2-qRT-F ATGCAATTCCTCAGCTGGA IGT10-qRT-R CCGAGAGTGCCAATGGTT IGT2-qRT-R TGCTAGTAAACCATCAGGC IGT11-qRT-F TTCACACTTGGAAACCCTTGTA IGT3-qRT-F AAGTTCCTCAGCTGGATG IGT11-qRT-R CAACCCAGAGTAAGATGTTTGA IGT3-qRT-R GGCCAATCGCTGAATTC IGT12-qRT-F ATGCAGATTCTTCAATGGATCTCC IGT4-qRT-F CTGAAAGATGGGTTTGCTTCA IGT12-qRT-R CCTCCCTTCGATTGTCTTT IGT4-qRT-R GATCATAGCCAAGAGTGCCAA IGT13-qRT-F CAGTGAACAAAATGAAGGGAAAG IGT5-qRT-F TCAGTGGGAAAGAGGTAAATC IGT13-qRT-R GTTGAGGGTGCTGTAAAAGAA IGT5-qRT-R CTGAACATTAAAACCTTTCTTCCA IGT14-qRT-F CTTGGTCATTGGGAATTCTTGC IGT6-qRT-F GAATGGAAAAGATTTTGAGGGCAATAC IGT14-qRT-R CCAGCAAAAGAGTTTCTAAGGT IGT6-qRT-R AAGCTCCTCTTCATCATCATC IGT15-qRT-F GATTTCACACTTGGAAATCCCTG IGT7-qRT-F TTGATCATCGGAGGGAATTCTT IGT15-qRT-R CCCAGAGTAAGATGTTTGATTTTCC IGT7-qRT-R CTGAATCTTCCTCATACTCATCATG IGT16-qRT-F AAATGATCATGAAATAGGCCGG IGT8-qRT-F CAACTACCCATTACATGAGGC IGT16-qRT-R TCCTCCCTTCGATTGTCTTT IGT8-qRT-R GCGTGGTTACTCCTTCCA IGT17-qRT-F TTGATCATCGGAGGAGGGA IGT9-qRT-F CAATGAAGTTCCTCAGCTGGAT IGT17-qRT-R CTTCCTCATACTCATCATGATCC  下载: 导出CSV
下载: 导出CSV
表 2 大豆IGT基因家族成员特征
Table 2. Properties of members of the IGT gene family in soybean
基因名称 序列号 染色体 位置 碱基对/bp 预测蛋白质长度/个 理论等电点 分子量/Da 亚细胞定位 GmIGT1 Glyma.02G097100 Chr02 8677685~8680757 1 101 367 6.91 40 326.05 细胞核 GmIGT2 Glyma.03G264200 Chr03 46860193~46864806 822 274 5.52 31 485.80 细胞核 GmIGT3 Glyma.07G040800 Chr07 3391340~3394088 846 282 7.58 32 278.71 叶绿体 GmIGT4 Glyma.07G052500 Chr07 4555925~4560419 858 286 5.45 32 189.21 细胞核 GmIGT5 Glyma.08G356200 Chr08 46252966~46256085 852 284 10.03 32 316.57 细胞核 GmIGT6 Glyma.09G071100 Chr09 7218596~7221206 789 263 5.11 29 050.36 细胞核 GmIGT7 Glyma.10G298100 Chr10 51546352~51548699 1068 356 5.96 39 907.67 细胞质 GmIGT8 Glyma.15G179200 Chr15 17222927~17225610 789 263 5.12 29 253.52 细胞核 GmIGT9 Glyma.16G009400 Chr16 842813~845650 855 285 8.03 32 507.05 细胞核 GmIGT10 Glyma.16G021400 Chr16 2005820~2009925 870 290 5.45 32 743.91 细胞质 GmIGT11 Glyma.16G054300 Chr16 5286678~5290902 1 149 383 6.65 43 362.59 叶绿体 GmIGT12 Glyma.16G094000 Chr16 15479890~15482382 786 262 9.67 30 379.48 细胞核 GmIGT13 Glyma.18G173300 Chr18 41543428~41546170 861 287 9.92 32 629.87 细胞核 GmIGT14 Glyma.18G284400 Chr18 56775415~56778618 1 098 366 6.91 40 390.19 细胞核 GmIGT15 Glyma.19G094600 Chr19 33952844~33956542 1 149 383 7.18 43 382.51 细胞核 GmIGT16 Glyma.19G263200 Chr19 51104292~51108788 807 269 5.68 30 957.17 细胞核 GmIGT17 Glyma.20G249200 Chr20 47749661~47752260 1 080 360 5.87 40 051.88 细胞核
下载: 导出CSV
-
[1] WAITE J M, DARDICK C. The roles of the IGT gene family in plant architecture: past, present, and future [J/OL]. Current Opinion in Plant Biology, 2021, 59 : 101983[2024-04-15]. DOI: 10.1016/j.pbi.2020.101983. [2] YOSHIHARA T, SPALDING E P. Switching the direction of stem gravitropism by altering two amino acids in AtLAZY1[J]. Plant Physiology, 2020, 182(2): 1039−1051. [3] YOSHIHARA T, SPALDING E P, IINO M. AtLAZY1 is a signaling component required for gravitropism of the Arabidopsis thaliana inflorescence[J]. The Plant Journal, 2013, 74(2): 267−279. [4] van OVERBEEK J. “LAZY ” , an a-geotropic form of maize: “gravitational indifference” rather than structural weakness accounts for prostrate growth-habit of this form [J]. Journal of Heredity, 1936, 27 : 93−96. [5] JONES J W, ADAIR C R. A “LAZY” mutation in rice[J]. Journal of Heredity, 1938, 29(8): 315−318. [6] LI Peijing, WANG Yonghong, QIAN Qiang, et al. LAZY1 controls rice shoot gravitropism through regulating polar auxin transport [J]. Cell Research, 2007, 17 : 402−410. [7] UGA Y, SUGIMOTO K, OGAWA S, et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions[J]. Nature Genetics, 2013, 45: 1097−1102. [8] KITOMI Y, HANZAWA E, KUYA N, et al. Root angle modifications by the DRO1 homolog improve rice yields in saline paddy fields[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(35): 21242−21250. [9] GE Liangfa, CHEN Rujin. Negative gravitropism in plant roots [J/OL]. Nature Plants, 2016, 2 (11): 16155[2024-04-15]. DOI: 10.1038/nplants.2016.155. [10] YOSHIHARA T, SPALDING E P. LAZY genes mediate the effects of gravity on auxin gradients and plant architecture[J]. Plant Physiology, 2017, 175(2): 959−969. [11] YU Baisheng, LIN Zhongwei, LI Haixia, et al. TAC1, a major quantitative trait locus controlling tiller angle in rice [J]. The Plant Journal, 2007, 52 (5): 891−898. [12] WAITE J M, DARDICK C. TILLER ANGLE CONTROL 1 modulates plant architecture in response to photosynthetic signals[J]. Journal of Experimental Botany, 2018, 69(20): 4935−4944. [13] WAITE J M, DARDICK C. IGT/LAZY family genes are differentially influenced by light signals and collectively required for light-induced changes to branch angle [J/OL]. BMC Biology, 2024 , 22 (1): 8[2024-04-15]. DOI: 10.1186/s12915-024-01813-4 [14] ZHANG Min, LIU Shulin, WANG Zhao, et al. Progress in soybean functional genomics over the past decade[J]. Plant Biotechnology Journal, 2022, 20(2): 256−282. [15] XU Cailong, HE Yanqin, SUN Shi, et al. Analysis of soybean yield formation differences across different production regions in China[J]. Agronomy Journal, 2020, 112(5): 4195−4206. [16] LIN Feng, CHHAPEKAR S S, VIEIRA C C, et al. Breeding for disease resistance in soybean: a global perspective[J]. Theoretical and Applied Genetics, 2022, 135(11): 3773−3872. [17] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆ Ct method[J]. Methods, 2001, 25(4): 402−408. [18] ZHAO Jianping, JIANG Lihui, BAI Hanrui, et al. Characteristics of members of IGT family genes in controlling rice root system architecture and tiller development [J/OL]. Frontiers in Plant Science, 2022, 13 : 961658[2024-04-15]. DOI: 10.3389/fpls.2022.961658. [19] SUN Chengming, ZHANG Chun, WANG Xiadong, et al. Genome-wide identification and characterization of the IGT gene family in allotetraploid rapeseed (Brassica napus L. )[J]. DNA and Cell Biology, 2021, 40(3): 441−456. [20] WANG Limin, CAI Wenbo, DU Chuanhui, et al. The isolation of the IGT family genes in Malus × domestica and their expressions in four idiotype apple cultivars [J/OL]. Tree Genetics & Genomes, 2018, 14 : 46[2024-04-15]. DOI: 10.1007/s11295-018-1258-9. [21] NAKAMURA M, NISHIMURA T, MORITA M T. Bridging the gap between amyloplasts and directional auxin transport in plant gravitropism[J]. Current Opinion in Plant Biology, 2019, 52: 54−60. [22] JIANG Jianhuan, TAN Lubin, ZHU Zuofeng, et al. Molecular evolution of the TAC1 gene from rice (Oryza sativa L.) [J]. Journal of Genetics and Genomics, 2012, 39 (10): 551−560. [23] KU Lixia, WEI Xiaomin, ZHANG Shaofang. Cloning and characterization of a putative TAC1 ortholog associated with leaf angle in maize (Zea mays L.) [J/OL]. PLoS One, 2011, 6 : e20621[2024-04-15]. DOI: 10.1371/journal.pone.0020621. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240354
 点击查看大图
点击查看大图
计量
- 文章访问数: 3056
- HTML全文浏览量: 893
- PDF下载量: 51
- 被引次数: 0
