








-
生境质量指生态系统为生物个体及种群提供适宜其持续发展条件的能力,在一定程度上可以反映区域生物多样性的状态[1]。近20 a的高速城镇化发展,使城市土地利用格局发生变化,从而对生境质量产生较大影响,也给区域生态安全带来巨大挑战[2]。城市建设及发展带来的土地利用/覆被变化(land use/cover change, LUCC)是影响区域生境质量的主要因素[3],因此,在区域土地利用状况及变化的基础上研究生境质量,有利于促进土地资源可持续利用及保障城市生态安全。
生境质量研究是当前国内外生态学者关注的热点,依据研究尺度和研究方法可分为2类:一类是单个物种小尺度的生境质量研究[4],主要通过对生物所处自然环境进行样方调查,构建评价体系,评估其生境质量;另一类是区域生境质量研究[5],研究尺度相对较大,主要在遥感数据的基础上,使用模型进行运算。常用模型如SolVES 模型[6],用于生态系统服务功能社会价值评估;MaxEnt模型[7],通过推算物种生态需求和模拟物种潜在分布,进行生境适宜性评价;InVEST模型[8],用于生境质量评估;IDRISI模型[9],用于动植物生境和生物多样性评价等。其中,InVEST模型是在LUCC下根据威胁因子和敏感度来评估区域生境质量[10],因参数获取方便、易于操作、分析结果准确、空间表达清晰等优点,被广泛运用于城市群[11]、省域[12]、自然保护区[13]、湿地[14]等多尺度多类型生境质量相关的研究中。
随着海南自由贸易港建设的加速推进,海南省会海口市正经历着新一轮土地开发浪潮,同时也面临着城市生境破碎程度上升、生物多样性保护压力增大的挑战。目前,有关海口市生境质量的研究主要集中在海岸带[15]、滨海旅游区[16]、建城区[17]或某类土地利用类型,如湿地[18]、耕地[19]等,但欠缺对海口市全域土地利用变化及其生境质量演变规律的研究,在一定程度上影响其总体生态保护规划及策略制定。因此,本研究采用InVEST模型,基于海口市2000、2010、2020年3期土地利用数据,在分析土地利用动态变化规律的基础上,探究其生境质量时空演变特征,总结演变机制和影响因素,以期为保护海口市生态环境,提高区域生态质量提供科学支撑。
-
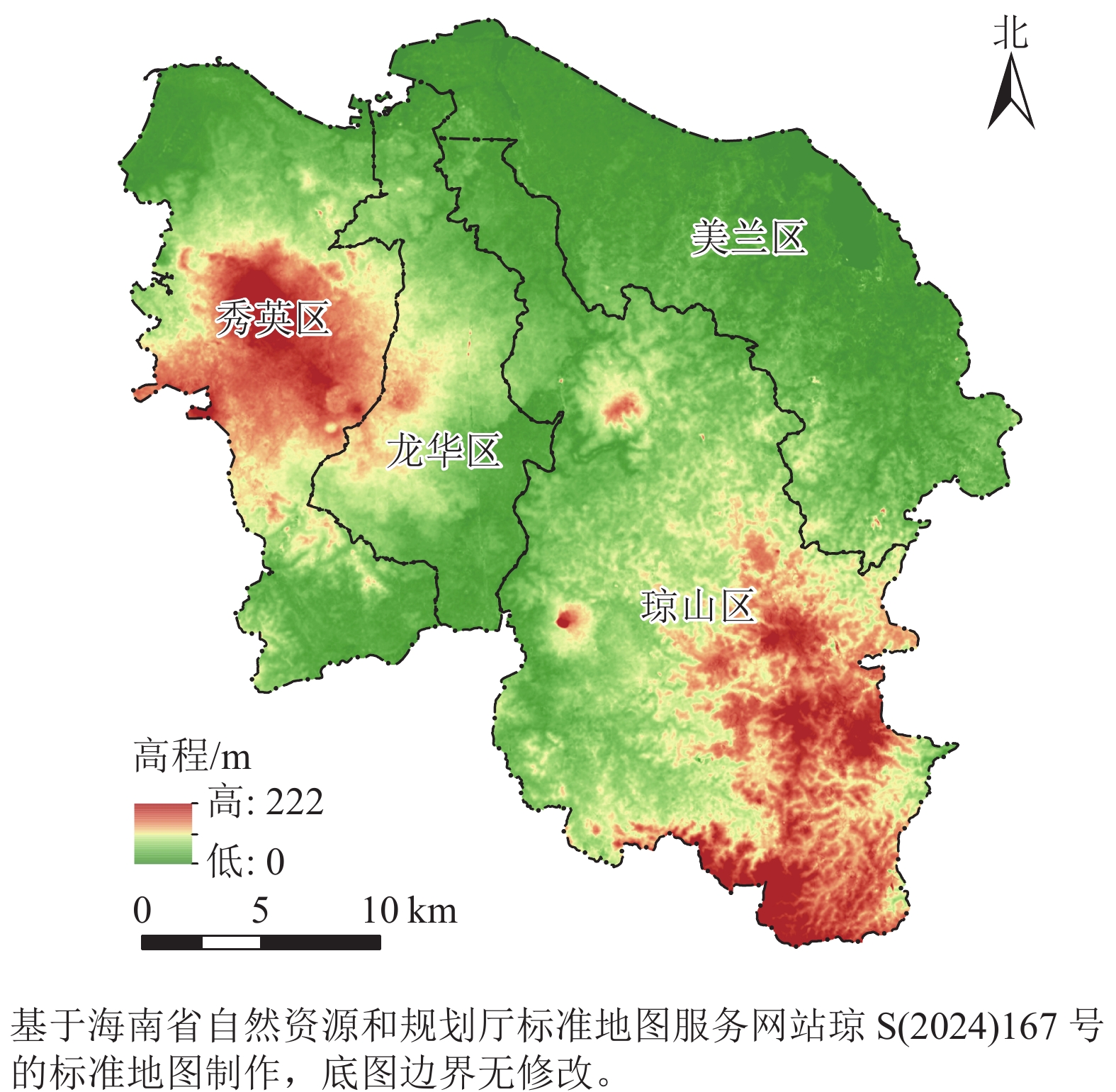
海口市(19°31′32″~20°04′52″N,110°07′22″~110°42′32″E)是海南省政治、经济、科技和文化中心,陆域面积为2 296.8 km2,下辖美兰、龙华、秀英、琼山等4个区。本研究选取海口市陆域(不含周边岛屿)为研究区(图1),常住人口为290.8万。全市地势平缓,水系发达,河流湖泊众多,在海口市东部沿海分布着红树林湿地,生境质量优良,生物多样性丰富。据海南省第三次森林资源调查统计,海口市森林覆盖率为39.60%,林木绿化率为40.08%。近20 a来,随着城镇化加速,以及海南国际旅游岛和自由贸易港政策的相继出台,海口市经历了人口大量涌入、城市建设持续蔓延、土地利用类型不断变化的过程,城市生态格局和生境质量也发生了相应的变化。
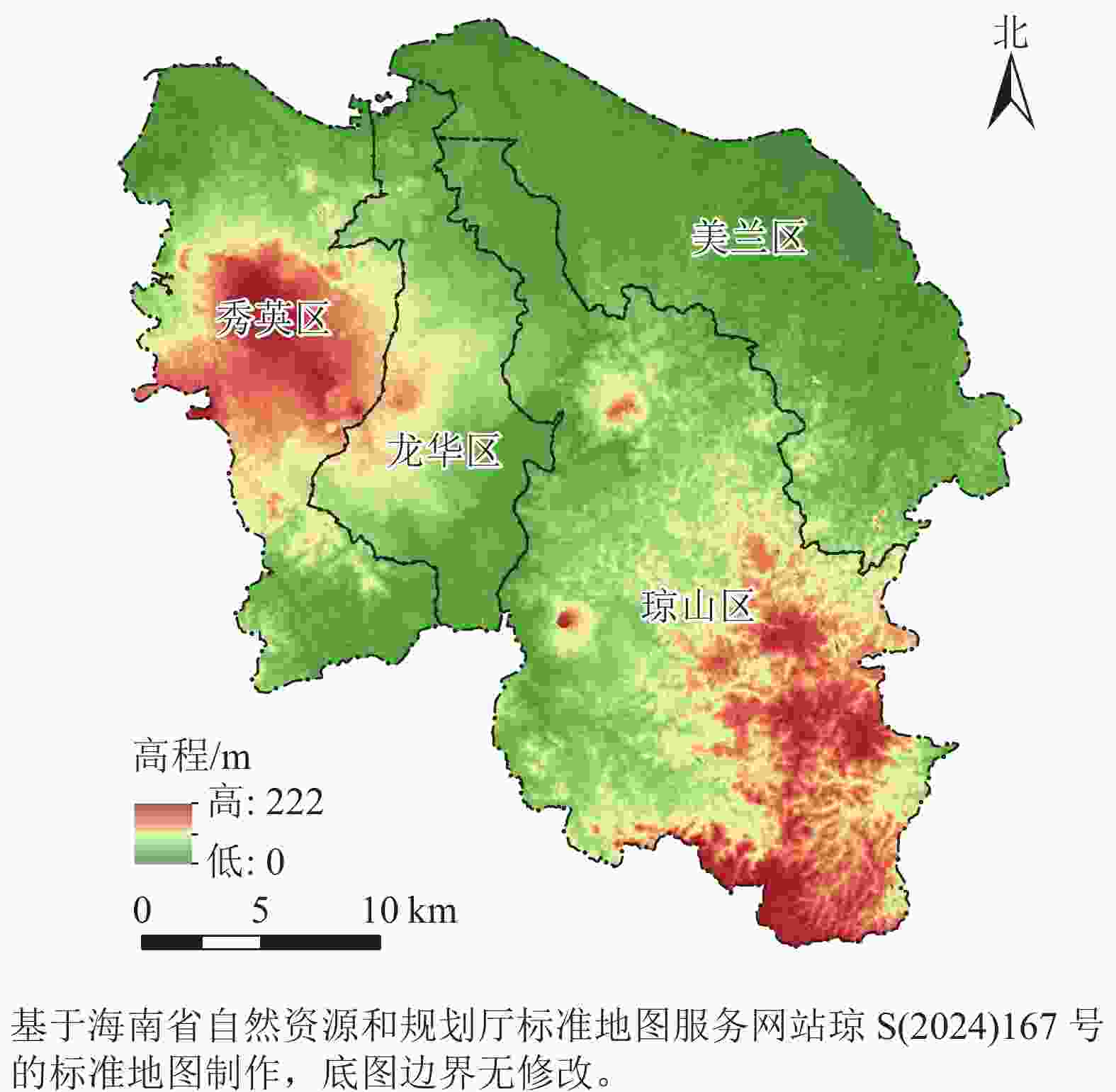
图 1 研究区位置及地形示意图
Figure 1. Location and topography of study area
-
所用数据包括海口市行政边界数据、土地利用数据(2000、2010、2020年)、数字高程数据(DEM)及道路矢量数据等。其中,海口市行政边界数据来源于海南测绘地理信息局网站;数字高程数据和道路矢量数据来源于地理空间数据云(
http://www.gscloud.cn/ );土地利用数据来源于国家基础地理信息中心全球地表覆盖数据产品服务网站,分辨率为30 m×30 m,总体精度达0.8以上。依据全国土地利用遥感监测分类体系,研究区土地利用类型分为耕地、林地、草地、湿地、水体、建设用地等6类一级类型。 -
土地利用转移矩阵是研究各种土地利用类型相互转换的方法之一,不仅可以定量研究不同土地利用类型之间的转化情况,还可以揭示不同土地利用类型之间的转移速率[20]。本研究通过ArcGIS软件对海口市2000和2020年2个时期的土地利用类型进行分析,建立2000—2020年海口市土地利用类型转移矩阵。计算公式如下:
$$ {{\boldsymbol{S}}_{ij}} = \left[ {\begin{array}{*{20}{c}} {{{\boldsymbol{S}}_{11}}}&{{{\boldsymbol{S}}_{12}}}& \cdots &{{\boldsymbol{S}}{}_{1n}} \\ {{{\boldsymbol{S}}_{21}}}&{{{\boldsymbol{S}}_{22}}}& \cdots &{{{\boldsymbol{S}}_{2n}}} \\ \vdots & \vdots & \vdots & \vdots \\ {{{\boldsymbol{S}}_{n1}}}&{{{\boldsymbol{S}}_{n2}}}& \cdots &{{{\boldsymbol{S}}_{nn}}} \end{array}} \right]。 $$ (1) 式(1)中:$ {\boldsymbol{S}} $为面积,$ n $为土地利用类型的数量;$ i $为研究初期土地利用类型;$ j $为研究末期土地利用类型。
-
InVEST模型可利用土地覆被和生物多样性威胁因素等信息生成生境质量地图,直观反映出研究区的生物多样性。本研究采用InVEST3.2.0模型中的生境质量模块(Habitat Quality)对海口市生境质量水平进行定量评估。计算公式如下:
$$ {Q_{xj}} = {H_j}\left[ {{{1 - }}\left( {\frac{{D_{xj}^z}}{{D_{xj}^z + {k^z}}}} \right)} \right]; $$ (2) $$ {D_{xj}} = \sum\limits_{r = 1}^R {} \sum\limits_{y = 1}^{Y_r} {} \left( {\frac{{{\omega _r}}}{{\displaystyle \sum\limits_{r = 1}^R {{\omega _r}} }}} \right){{{r}}_y}{i_{rxy}}{\beta _x}{S _{jr}} 。$$ (3) 式(2)~(3)中:$ {Q_{xj}} $为土地利用类型$ j $中栅格单元$ x $的生境质量指数大小;$ {H_j} $为土地利用类型$ j $的生境适宜度;$ {D_{xj}} $为土地利用类型$ j $中栅格单元$ x $的生境退化程度;$ z $为归一化常量,模型中设置为2.5;$ k $为半饱和系数,模型中设置为0.5;$ R $为威胁因子个数;$ {Y_r} $为威胁因子$ r $栅格单元的总数;$ {\omega _r} $为威胁因子$ r $的权重;$ {r_y} $为威胁因子$ r $在栅格$ y $的胁迫值;$ {i_{rxy}} $为栅格$ y $中的威胁因子$ r $对栅格$ x $的胁迫程度;$ {\beta _x} $为所有威胁因子在栅格单元$ x $的可达性;$ {S _{jr}} $为土地利用类型$ j $对威胁因子$ r $的敏感度。
$$ i_{rxy} = 1 - \left( {\frac{{d_{xy}}}{{d_{r\max} }}} \right);{\text{ }} $$ (4) $$ i_{rxy} = \exp \left[ { - \left( {\frac{{2.9{\text{9}}}}{{d_{r\max} }}} \right){{d}}_{xy}} \right]。 $$ (5) 式(4)为线性衰减,式(5)为指数衰减。$ {i_{rxy}} $为栅格$ y $中的威胁因子$ r $对栅格$ x $的胁迫程度;$ {d_{xy}} $为栅格$ x $和栅格$ y $之间的线性距离;$ {d_{r\max }} $为威胁因子$ r $的最大作用距离。
根据海口市地理环境和土地利用类型,参考文献[21]将耕地、建设用地、道路等人类活动密集区作为威胁因子,并进行比对校正,最终确定各相关数据及参数(表1和2)。
表 1 威胁因子最大影响距离及权重
Table 1. Maximum influence distance and weight of the threat source
威胁因子 最大影响距离/km 权重 衰减类型 耕地 4.0 0.7 线性 建设用地 7.0 1.0 指数 道路 2.0 0.6 指数 表 2 不同土地利用类型对威胁因子的敏感度
Table 2. Sensitivity of different land use types to threat factors
土地利用类型 生境适宜度 威胁因子 耕地 建设用地 道路 耕地 0.40 0 0.70 0.20 林地 1.00 0.70 0.80 0.50 草地 0.70 0.75 0.65 0.30 湿地 0.90 0.90 0.70 0.30 水体 0.80 0.75 0.85 0.70 建设用地 0 0 0 0 在模型中输入土地利用及威胁因子的栅格数据,并输入上述参数(格式为.csv),运行模型输出生境质量结果图。生境质量指数取值范围为0~1.0,利用自然间断点分级法将其分为4个等级:0~0.4(差)、>0.4~0.7(中)、>0.7~0.9(良)、>0.9~1.0(优)。
-
生境退化度(生境质量)变化率表示在某一时间段内,区域生境退化度(生境质量)时期末值相较于初始值的变化百分比[22],其计算公式为:
$$ v = \left( {{P_{{t1}}} - {P_{{t0}}}} \right){\raise0.7ex\hbox{${}$} \mathord{\left/ {\vphantom {{} {}}}\right.} \lower0.7ex\hbox{${}$}}{P_{{t0}}} \times 100\% 。$$ (6) 式(6)中:$ v $为生境退化度或生境质量的变化率(%),正值表示生境退化度或生境质量上升,负值表示下降;$ {P_{{t0}}} $、$ {P_{{t1}}} $分别为生境退化度或生境质量的初始值、末尾值。
-
由图2可知:研究区土地利用格局以林地和耕地为主。其中,林地主要分布在西南部和东南部区域;耕地广布于中南部区域;草地主要集中在秀英区北部和美兰区东部;水体主要由流经城市中部的南渡江及其支流,和分布各区的湖泊、水库组成;湿地主要为东北部的沿海红树林湿地;建设用地主要集中在北部平原区,且呈逐年扩张趋势。
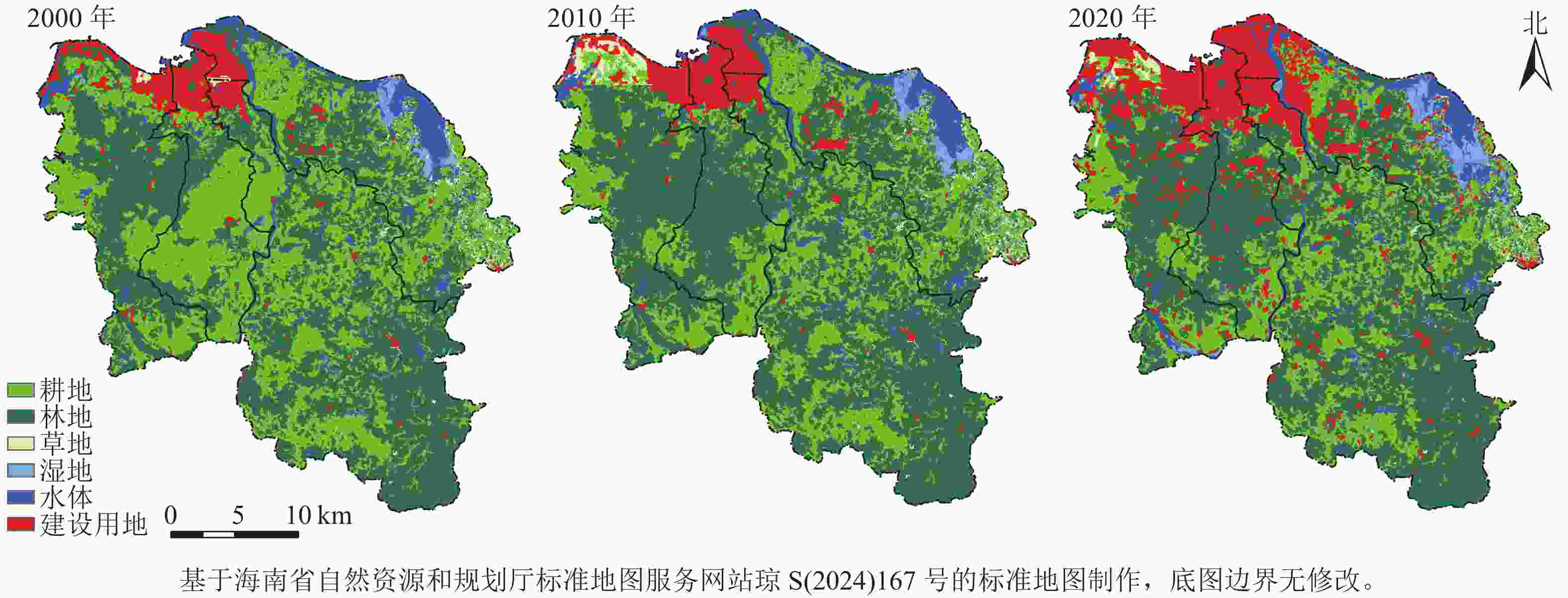
图 2 2000—2020年研究区土地利用类型空间分布示意图
Figure 2. Spatial distribution of land use types in the study area from 2000 to 2020
-
由表3可知:林地各期占比均最大,为海口市主要土地利用类型;其次是耕地,平均占比为30.54%;其他土地利用类型占比相对较少。2000—2020年,海口市耕地面积逐期下降,从2000年的871.32 km2下降到2020年的580.17 km2,占比从37.73%下降到25.12%;林地面积在2010年明显回升,但到2020年,林地面积回落到1 125.74 km2,占比为48.75%;湿地呈逐年上升趋势,面积从2000年的19.17 km2上升到2020年的39.38 km2,占比从0.83%上升到1.71%;建设用地面积增量最大,从2000年的133.58 km2增加到2020年的363.95 km2,占比从5.78%上升到15.76%,尤其是2010—2020年,建设用地面积增加了约1.5倍。
表 3 2000—2020年研究区土地利用类型面积及占比
Table 3. Area and proportion of land use types in the study area from 2000 to 2020
土地利用类型 2000年 2010年 2020年 面积/km2 占比/% 面积/km2 占比/% 面积/km2 占比/% 耕地 871.32 37.73 664.41 28.77 580.17 25.12 林地 1 090.34 47.22 1 270.70 55.03 1 125.74 48.75 草地 36.46 1.58 59.90 2.59 36.36 1.57 湿地 19.17 0.83 24.50 1.06 39.38 1.71 水体 158.34 6.86 141.79 6.14 163.61 7.09 建设用地 133.58 5.78 147.91 6.41 363.95 15.76 总计 2 309.21 100.00 2 309.21 100.00 2 309.21 100.00 -
由表4可知:耕地的转出面积最大,为311.10 km2,主要转为林地(200.36 km2)和建设用地(89.48 km2);其次是林地,转出面积为185.40 km2,主要转为建设用地和水体;其他土地利用类型的转出面积相对较小。建设用地的转入面积最大,为234.22 km2,主要来自于林地、耕地、水体;其次是林地,转入面积为220.87 km2,主要来自耕地。
表 4 2000—2020年研究区土地利用类型转移矩阵
Table 4. Conversion of different land use types in the study area from 2000 to 2020
年份 土地利用类型 2020年 耕地/km2 林地/km2 草地/km2 湿地/km2 水体/km2 建设用地/km2 转出/km2 2000 耕地 559.64 200.36 4.33 3.34 13.58 89.48 311.10 林地 13.24 904.18 17.85 7.49 23.02 123.81 185.40 草地 0.30 8.94 12.50 1.77 4.94 7.96 23.91 湿地 0.19 0.27 0.09 16.01 2.22 0.39 3.16 水体 5.33 9.24 1.40 10.75 96.84 12.58 39.29 建设用地 0.98 2.05 0.15 0.01 0.86 129.42 4.04 转入 20.03 220.87 23.82 23.35 44.62 234.22 -
由表5所示:2000—2020年海口市生境退化度平均值先下降后升高。2000—2010年,生境退化度变化率为−1.77%,主要与该时期实施的退耕还林政策有关;2010—2020年,生境退化度变化率为12.20%,主要与国际旅游岛建设和海南自由贸易港政策落地所带动的经济发展和城市建设有关,该时期人类活动程度加强,导致研究区建设用地大量增加,生态系统遭受到的人为干扰增大,生境退化程度增强。
表 5 2000—2020年研究区生境退化度变化率
Table 5. Change rate of habitat degradation in study area from 2000 to 2020
年份 生境退化度平均值 生境退化度变化率/% 2000 0.050 9 2010 0.050 0 −1.77 2020 0.056 1 12.20 -
利用自然间断点分级法对生境退化度进行分级,将其分为5个等级(图3):无退化(0)、轻度退化(>0~<0.02)、中度退化(0.02~<0.05)、高度退化(0.05~<0.09)和严重退化(≥0.09)。
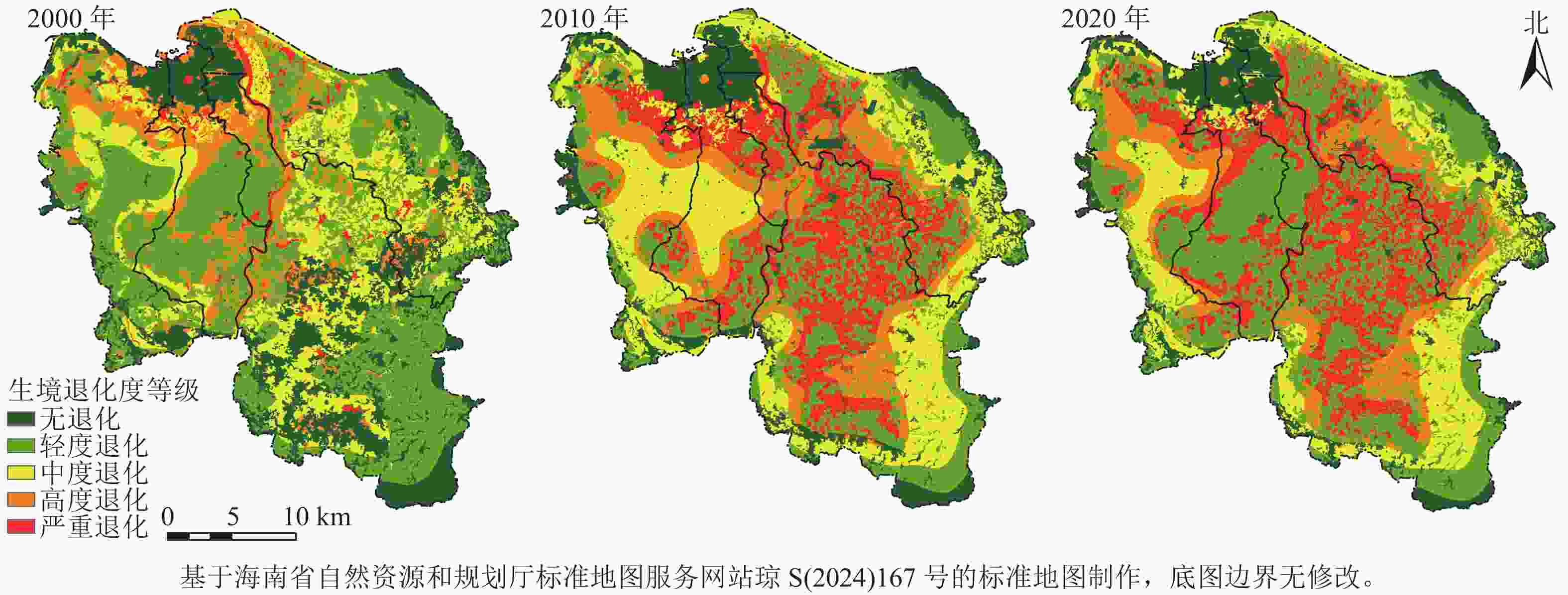
图 3 2000—2020年研究区生境退化度等级示意图
Figure 3. Habitat quality degradation grade in the study area from 2000 to 2020
总体上看,海口市生境退化度呈现出3个圈层递变的分布态势。第1个圈层的中心位于海口市主城区,土地利用类型为建设用地,人为干扰强度大,生境退化度等级为无退化。中心建城区边界周围,受城市扩张影响,生境退化等级为严重退化的范围逐期扩大,随后高度退化和中度退化穿插着往主城区外围扩散,呈递减现象;第2个圈层的中心在海口市秀英区休眠火山群周围的城镇建设区,该中心2000年为轻度退化,并与龙华区、琼山区、美兰区的轻度退化区域连成片,后退化等级逐期加强,到2020年演变为中度退化和高度退化;第3个圈层分布在琼山区东南角,该区域受台地和火山地貌影响,且离主城区较远,其生境质量难以受到干扰,故生境退化度等级由轻度退化到中度退化,再到高度退化。
-
由表6所示:海口市生境质量在2000—2010年处于上升趋势,生境质量变化率为6.92%,2010—2020年为下降趋势,生境质量变化率为−10.44%。从各区生境质量来看,2000年龙华区的生境质量指数最低,为0.525 1,2000—2010年该区大面积退耕还林,使得其生境质量指数大幅提高,为0.717 1,生境质量指数变化率为36.56% ;2010和2020年这2个时期,美兰区生境质量指数都最低,分别为0.697 8和0.598 7,这与美兰区建设用地大面积增加有关;琼山区各期生境质量指数都最高,分别为0.738 9、0.777 6和0.741 2,主要由于琼山区离主城区相对较远,且多火山丘陵地貌,林地面积占比高。
表 6 2000—2020年研究区及各区生境质量指数平均值及变化率
Table 6. Average value and change rate of habitat quality of the study area and its districts from 2000 to 2020
区域 生境质量指数平均值 2000年 2010年 变化率/% 2020年 变化率/% 海口市(不含周边岛屿) 0.688 0 0.735 6 6.92 0.658 8 −10.44 美兰区 0.683 4 0.697 8 2.11 0.598 7 −14.20 龙华区 0.525 1 0.717 1 36.56 0.629 3 −12.24 秀英区 0.697 0 0.711 1 2.02 0.600 7 −15.53 琼山区 0.738 9 0.777 6 5.24 0.741 2 −4.68 -
由图4可知:生境质量等级为差的区域主要分布在海口市北部建城区,随着城市扩张逐期扩大。生境质量等级为优的区域主要分布在秀英区、龙华区和琼山区,尤其是秀英区火山地质公园所处的马鞍岭—雷虎岭火山群一带,以及琼山区的旧州岭—日晒岭一带。从时空变化来看,2000—2010年,龙华区大面积耕地转化为林地,且建设用地扩张较为缓慢,使得研究区生境质量总体水平上升。2010—2020年,城市建设突飞猛进,导致海口市主城区周边生境质量等级为中的区域逐渐被等级为差的区域吞并。主城区外围,随着高铁、高速公路网等基础交通设施的修建,生境频繁被分割,破碎化程度加重。在南渡江、湖泊、水库等水域周边,人为建设量增加,导致生境质量逐期下降,生境破碎化程度同样加重。
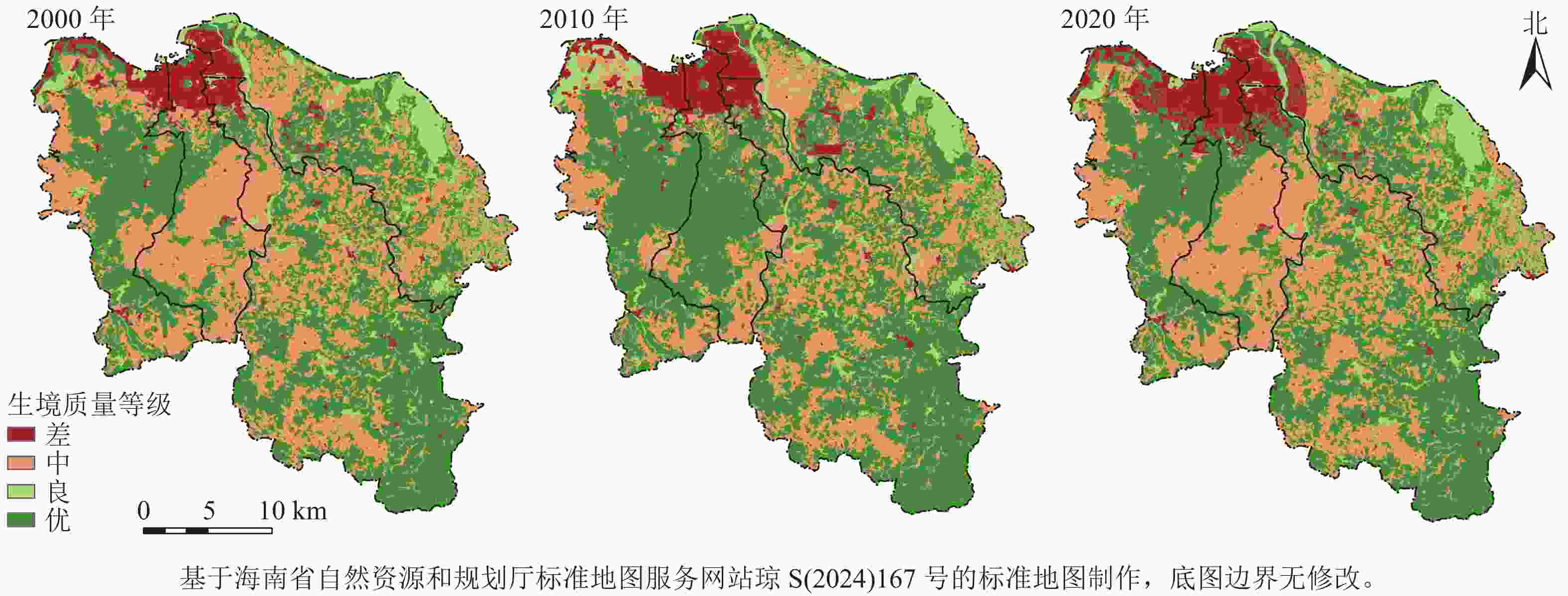
图 4 2000—2020年研究区生境质量等级空间分布示意图
Figure 4. Spatial distribution of habitat quality grades in the study area from 2000 to 2020
由表7所示:海口市2000—2020年生境质量等级为差的占比从5.33%上升到16.83%,但生境质量等级为优的区域占比各期均在50.00%左右,表明区域整体生境质量水平较高,生态环境较好。从4个区的变化上看,生境质量等级为差的面积均逐年增加,增幅最大的为美兰区,从2000年的23.13 km2增加到2020年的140.49 km2,增加了约5倍。琼山区生境质量等级为优的土地面积各期占比均最大,表明海口市4个区中琼山区的生境质量最好。
表 7 2000—2020年研究区生境质量等级面积及占比
Table 7. Habitat quality grade area and percentage in the study area from 2000 to 2020
区域 年份 差 中 良 优 面积/km2 占比/% 面积/km2 占比/% 面积/km2 占比/% 面积/km2 占比/% 美兰区 2000 23.13 3.96 173.38 29.72 131.69 22.57 255.27 43.75 2010 31.77 5.44 153.80 26.36 120.03 20.57 277.88 47.62 2020 140.49 24.08 109.81 18.82 78.95 13.53 254.19 43.57 龙华区 2000 33.40 11.02 174.63 57.64 10.95 3.61 83.99 27.72 2010 35.65 11.76 76.70 25.31 10.48 3.46 180.20 59.47 2020 66.34 21.90 66.86 22.07 10.57 3.49 159.21 52.55 秀英区 2000 42.91 8.67 165.62 33.47 21.52 4.35 264.78 53.51 2010 47.01 9.50 136.57 27.60 44.56 9.01 266.64 53.89 2020 112.42 22.72 120.63 24.38 32.25 6.52 229.51 46.38 琼山区 2000 23.64 2.55 340.81 36.73 33.94 3.66 529.55 57.07 2010 25.25 2.72 281.71 30.36 29.97 3.23 590.99 63.69 2020 69.50 7.49 259.17 27.93 36.89 3.98 562.42 60.61 海口市
(不含周边岛屿)2000 123.08 5.33 854.44 37.00 198.10 8.58 1 133.59 49.09 2010 139.68 6.05 648.78 28.10 205.04 8.88 1 315.71 56.98 2020 388.75 16.83 556.47 24.10 158.66 6.87 1 205.33 52.20 -
由图5所示:海口市生境质量等级转换频繁的区域主要在主城区外围,南渡江流域周边,美兰区沿海湿地,以及龙华区大面积耕地范围内;生境质量由中转为差的区域主要集中在海口市主城区周边、美兰区及秀英区长流镇,这与海口市城市发展建设的重点区域相吻合。生境质量由中转为良和优的区域主要集中在龙华区和秀英区,这是由于该区域分布着火山口地质公园,生境质量较高;生境质量由良转为中和优的区域,主要集中在海口市东北部和北部的滨海湿地内。
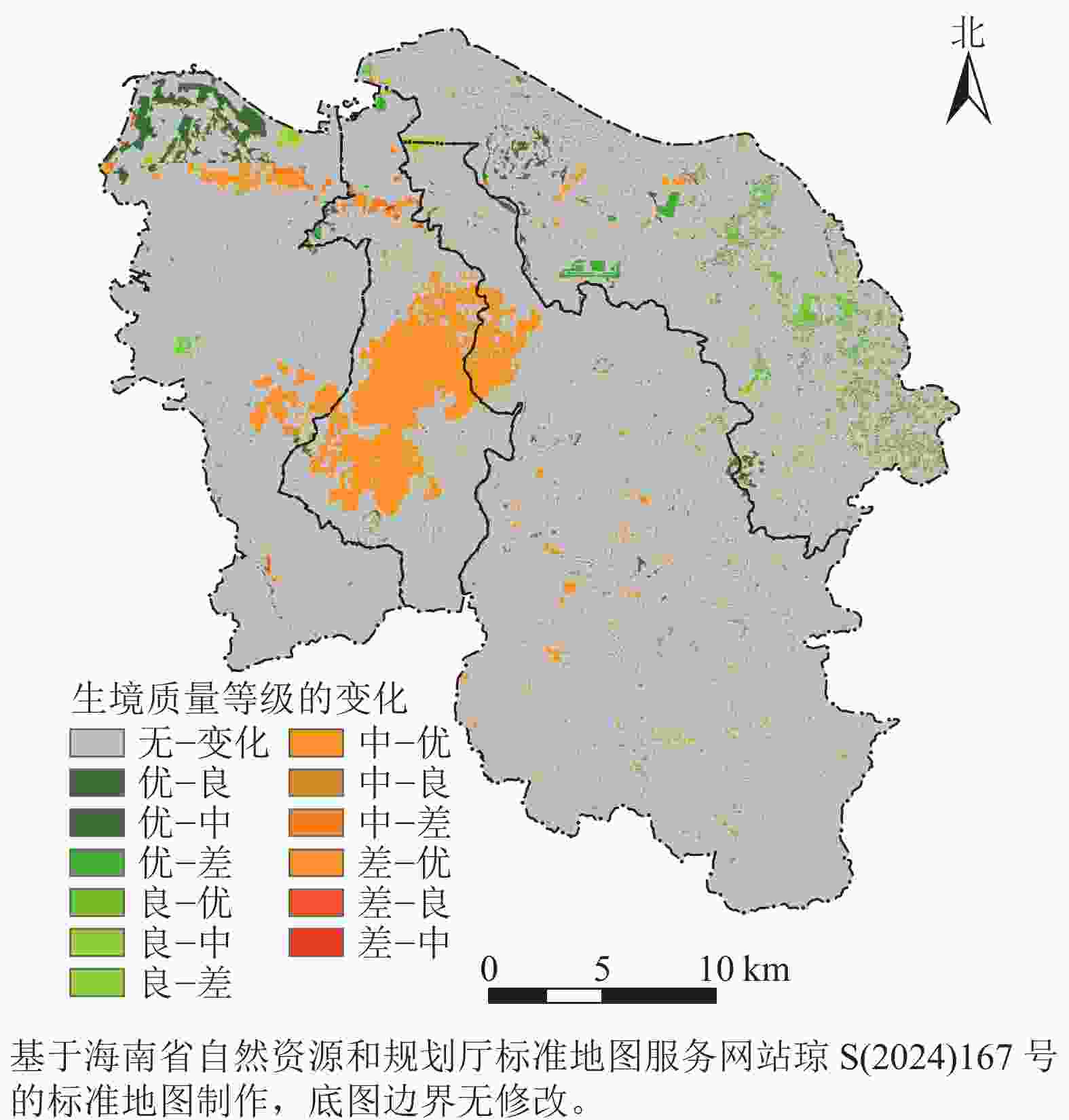
图 5 2000—2020年研究区生境质量变化空间分布示意图
Figure 5. Spatial distribution of habitat quality changes in study area from 2000 to 2020
-
自然地理环境和人类社会经济活动是区域生境质量空间分布格局的主要影响因素。本研究通过InVEST模型对海口市的生境质量进行评估,结果显示海口市生境质量整体呈现出北部主城区低,西部和东南部台地区域高的空间分布格局,与全市“核心城区—城乡交错带—乡村”的土地利用格局相契合,说明生境质量空间分布与地貌形态和土地利用类型有关。海口市作为海南“三区一中心”战略定位和“中国特色自由贸易港”建设的前沿阵地,城市发展向西拓展长流组团,向东建设江东新区,并加强建设城际铁路、区域快线等高效便捷的交通网络。与此同时,与澄迈、文昌协同打造“海澄文”经济圈。密集的社会经济活动导致土地利用强度提高,给城市生态环境带来巨大挑战,如城市热岛、洪水、环境污染、废弃物和野生动物栖息地缺失等问题,也对主城区生境质量产生巨大威胁,因此海口市主城区以及重点乡镇区域的生境质量有所降低。
-
2000—2020年,海口市生境质量总体呈先上升后下降的态势。自2002年起,海南省实施了大面积的退耕还林政策,土地利用向高质量生境类型转变,使区域生境质量指数有较为明显的提升;2010—2020年,海口市生境质量总体下降10.44%,随着该时段社会经济发展,由于人口增加、城市化建设等因素,尤其是2010年海南国际旅游岛政策提出后,全市建设用地面积从133.58 km2激增到363.95 km2,增加了172.46%,城市发展和土地开发“摊大饼”式扩张引起自然生境斑块丧失,生境破碎化加剧,导致区域生境质量退化。这与同处沿海地区的福建省[23]、京津冀区域[24]生境质量的演变规律一致,也说明了海口市生境质量变化与城市土地利用类型及地表覆盖物变化有关,且随着海南自由贸易港建设的持续推进,未来海口市建设用地仍将有巨大增量,生态环境将面临更为严峻的考验。
-
城市的生态系统极为脆弱,各种基础设施占据了城市中大量面积,并挤压着城市自然空间的范围,导致局部生境质量退化甚至丧失[25]。海口市规划建设应以其自然地理格局为基础,立足海南自由贸易港核心城市和国际化滨江滨海花园城市,统筹国土空间开发与生态系统保护制度,突出优质生态产品供给、生态价值实现、绿色发展成果共享的生态经济模式,进一步提升生态安全保障和生态系统服务功能。首先,要以生态优先的规划方法指导城市建设,建立蓝绿生态网络,强化城市建设集约化用地,避免建设空间无序蔓延;北部主要建设区采用组团发展、绿地楔入的空间模式;培育滨海生态绿带,保护滨水生态廊道,优化湿地自然公园等生态斑块。其次,加强城区生态修复和综合治理。注重绿化植物选材本土化,构建以提高生态服务功能为导向的生态修复体系,通过自然恢复、生态修复等措施改善生境质量,提高城市生态韧性及生态安全自我保障能力。
-
2000—2020年,研究区各土地利用类型面积占比从大到小为:林地、耕地、建设用地、水体、草地、湿地,建设用地增幅最大,主要由林地、耕地和草地转变而来;海口市生境退化度先下降后上升,在空间上呈圈层递进分布规律;海口市生境质量总体呈先升后降趋势,与城市建设强度密切相关。本研究从生境退化度和生境质量两方面阐释了海口市生境的变化情况,可为该区域生境保护政策的制定提供参考,为长时间序列的生境质量研究提供了思路。
Spatio-temporal evolution of habitat quality in Haikou from 2000 to 2020 based on InVEST model
-
摘要:
目的 分析海口市生境质量及其影响因素,为生态环境保护及城市建设提供数据支撑及科学参考。 方法 基于2000、2010、2020年3期土地利用数据,采用InVEST模型,综合应用土地利用转移、生境质量变化率和空间统计分析等方法,剖析海口市生境退化度和生境质量时空动态变化特征,并对其影响因素进行总结分析。 结果 ①2000—2020年,林地是海口市主要土地利用类型,各期面积占比为47.22%、55.03%、48.75%,其次是耕地;研究期内建设用地面积增幅最大,增量为230.37 km2,主要由林地、耕地和草地转变而来。②2000—2010年,海口市生境退化度变化率为−1.77%,2010年后生境退化明显,生境退化度变化率为12.20%;研究期内海口市生境退化等级在空间上呈现出以建城区为中心,圈层递减的分布格局。③海口市生境质量总体较好,生境质量等级为优的面积占比各期均在50.00%左右,等级为差的面积占比从2000年的5.33%上升到2020年的16.83%;生境质量等级在海口市中心城区城乡结合部等区域变化较大,转换频繁;研究期内海口市生境质量指数先上升后降低,整体由0.688 0下降至0.658 8。 结论 2000—2020年海口市生境质量总体呈先升后降趋势,与城市建设强度密切相关。海口市应保持生态优先,注重主城区生态修复和基础设施生态化建设,加强主城区外围自然环境生态系统的保护和综合治理。图5表7参25 Abstract:Objective This study is to analyze the habitat quality and its influencing factors in Haikou, so as to provide data support and scientific reference for ecological environment protection and urban construction. Method Based on the land use data of 2000, 2010 and 2020, InVEST model and the methods of land use transfer, habitat quality change rate and spatial statistical analysis were used to analyze the spatial-temporal dynamic changes in habitat degradation and habitat quality in Haikou, and summarize the influencing factors. Result ① From 2000 to 2020, forestland was the main type of land use in Haikou, accounting for 47.22%, 55.03%, and 48.75% of the total area in each period, followed by cropland. During the research period, the increase of built land area was the largest, with an increase of 230.37 km2, mainly transformed from forestland, cropland, and grassland. ②The change rate of habitat degradation degree was −1.77% from 2000 to 2010, and 12.20% from 2010 to 2020. The level of habitat degradation showed a spatial distribution pattern centered around the urban area and decreasing in circles. ③The overall habitat quality in Haikou was good. The proportion of areas with excellent habitat quality grades was around 50.00% in all stages, while the proportion of areas with poor grades increased from 5.33% in 2000 to 16.83% in 2020. The habitat quality level changed significantly and frequently in areas such as the urban-rural fringe in the central urban area of Haikou. During the research period, the habitat quality index of Haikou firstly increased and then decreased, and showed an overall decrease from 0.6880 to0.6588 .Conclusion From 2000 to 2020, the overall habitat quality in Haikou firstly increased and then decreased, which is closely related to the intensity of urban construction. Haikou should maintain the ecological priority and pay attention to ecological restoration of the main urban area, ecological infrastructure construction, and strengthen the protection and comprehensive management of the ecological system of the peripheral natural environment of the main urban area. [Ch, 5 fig. 7 tab. 25 ref.] -
Key words:
- Haikou /
- land use /
- habitat degradation degree /
- habitat quality /
- InVEST model
-
图 2 2000—2020年研究区土地利用类型空间分布示意图
Figure 2 Spatial distribution of land use types in the study area from 2000 to 2020
图 3 2000—2020年研究区生境退化度等级示意图
Figure 3 Habitat quality degradation grade in the study area from 2000 to 2020
图 4 2000—2020年研究区生境质量等级空间分布示意图
Figure 4 Spatial distribution of habitat quality grades in the study area from 2000 to 2020
图 5 2000—2020年研究区生境质量变化空间分布示意图
Figure 5 Spatial distribution of habitat quality changes in study area from 2000 to 2020
表 1 威胁因子最大影响距离及权重
Table 1. Maximum influence distance and weight of the threat source
威胁因子 最大影响距离/km 权重 衰减类型 耕地 4.0 0.7 线性 建设用地 7.0 1.0 指数 道路 2.0 0.6 指数  下载: 导出CSV
下载: 导出CSV
表 2 不同土地利用类型对威胁因子的敏感度
Table 2. Sensitivity of different land use types to threat factors
土地利用类型 生境适宜度 威胁因子 耕地 建设用地 道路 耕地 0.40 0 0.70 0.20 林地 1.00 0.70 0.80 0.50 草地 0.70 0.75 0.65 0.30 湿地 0.90 0.90 0.70 0.30 水体 0.80 0.75 0.85 0.70 建设用地 0 0 0 0
下载: 导出CSV
表 3 2000—2020年研究区土地利用类型面积及占比
Table 3. Area and proportion of land use types in the study area from 2000 to 2020
土地利用类型 2000年 2010年 2020年 面积/km2 占比/% 面积/km2 占比/% 面积/km2 占比/% 耕地 871.32 37.73 664.41 28.77 580.17 25.12 林地 1 090.34 47.22 1 270.70 55.03 1 125.74 48.75 草地 36.46 1.58 59.90 2.59 36.36 1.57 湿地 19.17 0.83 24.50 1.06 39.38 1.71 水体 158.34 6.86 141.79 6.14 163.61 7.09 建设用地 133.58 5.78 147.91 6.41 363.95 15.76 总计 2 309.21 100.00 2 309.21 100.00 2 309.21 100.00
下载: 导出CSV
表 4 2000—2020年研究区土地利用类型转移矩阵
Table 4. Conversion of different land use types in the study area from 2000 to 2020
年份 土地利用类型 2020年 耕地/km2 林地/km2 草地/km2 湿地/km2 水体/km2 建设用地/km2 转出/km2 2000 耕地 559.64 200.36 4.33 3.34 13.58 89.48 311.10 林地 13.24 904.18 17.85 7.49 23.02 123.81 185.40 草地 0.30 8.94 12.50 1.77 4.94 7.96 23.91 湿地 0.19 0.27 0.09 16.01 2.22 0.39 3.16 水体 5.33 9.24 1.40 10.75 96.84 12.58 39.29 建设用地 0.98 2.05 0.15 0.01 0.86 129.42 4.04 转入 20.03 220.87 23.82 23.35 44.62 234.22
下载: 导出CSV
表 5 2000—2020年研究区生境退化度变化率
Table 5. Change rate of habitat degradation in study area from 2000 to 2020
年份 生境退化度平均值 生境退化度变化率/% 2000 0.050 9 2010 0.050 0 −1.77 2020 0.056 1 12.20
下载: 导出CSV
表 6 2000—2020年研究区及各区生境质量指数平均值及变化率
Table 6. Average value and change rate of habitat quality of the study area and its districts from 2000 to 2020
区域 生境质量指数平均值 2000年 2010年 变化率/% 2020年 变化率/% 海口市(不含周边岛屿) 0.688 0 0.735 6 6.92 0.658 8 −10.44 美兰区 0.683 4 0.697 8 2.11 0.598 7 −14.20 龙华区 0.525 1 0.717 1 36.56 0.629 3 −12.24 秀英区 0.697 0 0.711 1 2.02 0.600 7 −15.53 琼山区 0.738 9 0.777 6 5.24 0.741 2 −4.68
下载: 导出CSV
表 7 2000—2020年研究区生境质量等级面积及占比
Table 7. Habitat quality grade area and percentage in the study area from 2000 to 2020
区域 年份 差 中 良 优 面积/km2 占比/% 面积/km2 占比/% 面积/km2 占比/% 面积/km2 占比/% 美兰区 2000 23.13 3.96 173.38 29.72 131.69 22.57 255.27 43.75 2010 31.77 5.44 153.80 26.36 120.03 20.57 277.88 47.62 2020 140.49 24.08 109.81 18.82 78.95 13.53 254.19 43.57 龙华区 2000 33.40 11.02 174.63 57.64 10.95 3.61 83.99 27.72 2010 35.65 11.76 76.70 25.31 10.48 3.46 180.20 59.47 2020 66.34 21.90 66.86 22.07 10.57 3.49 159.21 52.55 秀英区 2000 42.91 8.67 165.62 33.47 21.52 4.35 264.78 53.51 2010 47.01 9.50 136.57 27.60 44.56 9.01 266.64 53.89 2020 112.42 22.72 120.63 24.38 32.25 6.52 229.51 46.38 琼山区 2000 23.64 2.55 340.81 36.73 33.94 3.66 529.55 57.07 2010 25.25 2.72 281.71 30.36 29.97 3.23 590.99 63.69 2020 69.50 7.49 259.17 27.93 36.89 3.98 562.42 60.61 海口市
(不含周边岛屿)2000 123.08 5.33 854.44 37.00 198.10 8.58 1 133.59 49.09 2010 139.68 6.05 648.78 28.10 205.04 8.88 1 315.71 56.98 2020 388.75 16.83 556.47 24.10 158.66 6.87 1 205.33 52.20
下载: 导出CSV
-
[1] RIEDLER B, LANG S. A spatially explicit patch model of habitat quality, integrating spatio-structural indicators [J]. Ecological Indicators, 2018, 94: 128−141. [2] SUN Xiaoyin, JIANG Zhai, LIU Fei, et al. Monitoring spatio-temporal dynamics of habitat quality in Nansihu Lake Basin, Eastern China, from 1980 to 2015 [J]. Ecological Indicators, 2019, 102: 716−723. [3] 雷金睿, 陈宗铸, 陈小花, 等. 1980—2018年海南岛土地利用与生态系统服务价值时空变化[J]. 生态学报, 2020, 40(14): 4760−4773. LEI Jinrui, CHEN Zongzhu, CHEN Xiaohua, et al. Spatio-temporal changes of land use and ecosystem services value in Hainan Island from 1980 to 2018 [J]. Acta Ecologica Sinica, 2020, 40(14): 4760−4773. [4] 吴健生, 曹祺文, 石淑芹, 等. 基于土地利用变化的京津冀生境质量时空演变[J]. 应用生态学报, 2015, 26(11): 3457−3466. WU Jiansheng, CAO Qiwen, SHI Shuqin, et al. Spatio-temporal variability of habitat quality in Beijing-Tianjin-Hebei Area based on land use change [J]. Chinese Journal of Applied Ecology, 2015, 26(11): 3457−3466. [5] 王军, 严有龙, 王金满, 等. 闽江流域生境质量时空演变特征与预测研究[J]. 生态学报, 2021, 41(14): 5837−5848. WANG Jun, YAN Youlong, WANG Jinman, et al. Temporal-spatial variation characteristics and prediction of habitat quality in Min River Basin [J]. Acta Ecologica Sinica, 2021, 41(14): 5837−5848. [6] 赵琪琪, 李晶, 刘婧雅, 等. 基于SolVES模型的关中-天水经济区生态系统文化服务评估[J]. 生态学报, 2018, 38(10): 3673−3681. ZHAO Qiqi, LI Jing, LIU Jingya, et al. Assessment and analysis of social values of cultural ecosystem services based on the SolVES model in the Guanzhong-Tianshui Economic Region [J]. Acta Ecologica Sinica, 2018, 38(10): 3673−3681. [7] 李莎, 莫舜华, 胡兴华, 等. 基于MaxEnt和ArcGIS预测濒危植物资源冷杉潜在适生区分析[J]. 生态学杂志, 2024, 43(2): 533−541. LI Sha, MO Shunhua, HU Xinghua, et al. Prediction of potential suitable areas of endangered plant Abies ziyuanensis based on MaxEnt and ArcGIS [J]. Chinese Journal of Ecology, 2024, 43(2): 533−541. [8] 蔡梦卿, 黄璐, 严力蛟. 基于InVEST模型的杭州市典型年份年产水量时空变化特征及其影响因素[J]. 浙江农林大学学报, 2022, 39(1): 127−135. CAI Mengqing, HUANG Lu, YAN Lijiao. Temporal and spatial variation characteristics of annual water yield and its influencing factors in Hangzhou based on InVEST model [J]. Journal of Zhejiang A&F University, 2022, 39(1): 127−135. [9] DEVI B S, MURTHY M S R, BIJAN D, et al. Identification of potential habitat patches for connectivity using Weighted Linear Combination (WLC) and Integral Index of Connectivity (IIC) at East Godavari District, Andhra Pradesh, India [J]. Journal of the Indian Society of Remote Sensing, 2016, 44(3): 385−394. [10] 雷金睿, 陈毅青, 陈宗铸, 等. 基于InVEST模型的海南岛三大流域生境质量时空演变[J]. 应用生态学报, 2022, 33(9): 2511−2520. LEI Jinrui, CHEN Yiqing, CHEN Zongzhu, et al. Spatiotemporal evolution of habitat quality in three basins of Hainan Island based on InVEST model [J]. Chinese Journal of Applied Ecology, 2022, 33(9): 2511−2520. [11] 邱天琦, 王向荣. 基于InVEST模型的长株潭城市群生境质量时空演变分析研究[J]. 林业资源管理, 2022(5): 99−106. QIU Tianqi, WANG Xiangrong. Research on the temporal and spatial evolution of habitat quality in Changsha-Zhuzhou-Xiangtan urban agglomeration based on InVEST model [J]. Forest Resources Management, 2022(5): 99−106. [12] 梁晓瑶, 袁丽华, 宁立新, 等. 基于InVEST模型的黑龙江省生境质量空间格局及其影响因素[J]. 北京师范大学学报(自然科学版), 2020, 56(6): 864−872. LIANG Xiaoyao, YUAN Lihua, NING Lixin, et al. Spatial pattern of habitat quality and driving factors in Heilongjiang Province [J]. Journal of Beijing Normal University (Natural Science), 2020, 56(6): 864−872. [13] SALLUSTIO L, de TONI A, STROLLO A, et al. Assessing habitat quality in relation to the spatial distribution of protected areas in Italy [J]. Journal of Environmental Management, 2017, 201: 129−137. [14] 刘志伟. 基于InVEST的湿地景观格局变化生态响应分析——以杭州湾南岸地区为例[D]. 杭州: 浙江大学, 2014. LIU Zhiwei. Ecological Response Analysis of Wetland Landscape Pattern Change Based on InVEST−a Case Study of South Bank of Hangzhou Bay [D]. Hangzhou: Zhejiang University, 2014. [15] 罗文霞, 钟康正, 周晓娟, 等. 海口市江东新区30年来海岸带城市扩展与生态格局演变[J]. 生态学报, 2022, 42(6): 2164−2174. LUO Wenxia, ZHONG Kangzheng, ZHOU Xiaojuan, et al. Urban expansion and ecological pattern evolution of coastal zone in Jiangdong New District of Haikou City in the past 30 years [J]. Acta Ecologica Sinica, 2022, 42(6): 2164−2174. [16] 张明湖, 李鹏山, 苏珊, 等. 海口市滨海旅游区LUCC变化趋势预测[J]. 海南师范大学学报(自然科学版), 2010, 23(2): 198−203. ZHANG Minghu, LI Pengshan, SU Shan, et al. Study on the LUCC trend of the coastal resorts of Haikou City [J]. Journal of Hainan Normal University (Natural Science), 2010, 23(2): 198−203. [17] 雷金睿, 陈宗铸, 杨琦, 等. 基于GIS的海口市建成区景观格局分析[J]. 福建林业科技, 2018, 45(2): 78−82. LEI Jinrui, CHEN Zongzhu, YANG Qi, et al. GIS-based analysis of the urban landscape pattern of Haikou built-up areas [J]. Journal of Fujian Forestry Science and Technology, 2018, 45(2): 78−82. [18] 陈圣天, 付晖, 付广, 等. 海口湿地生态系统服务价值时空演变及其对景观格局变化的响应[J]. 西北林学院学报, 2023, 38(6): 236−242. CHEN Shengtian, FU Hui, FU Guang, et al. The spatial and temporal evolution of wetland ecosystem service value and its response to landscape pattern changes in Haikou [J]. Journal of Northwest Forestry University, 2023, 38(6): 236−242. [19] 王婷, 韦仕川. 2009—2018年海口市土地利用转型特征及其影响因素[J]. 水土保持通报, 2022, 42(1): 344−352. WANG Ting, WEI Shichuan. Characteristics and influencing factors of land use transition in Haikou City during 2009−2018 [J]. Bulletin of Soil and Water Conservation, 2022, 42(1): 344−352. [20] 杨清可, 段学军, 王磊, 等. 基于“三生空间”的土地利用转型与生态环境效应——以长江三角洲核心区为例[J]. 地理科学, 2018, 38(1): 97−106. YANG Qingke, DUAN Xuejun, WANG Lei, et al. Land use transformation based on ecological-production-living spaces and associated eco-environment effects: a case study in the Yangtze River Delta [J]. Scientia Geographica Sinica, 2018, 38(1): 97−106. [21] 巩杰, 马学成, 张玲玲, 等. 基于InVEST模型的甘肃白龙江流域生境质量时空分异[J]. 水土保持研究, 2018, 25(3): 191−196. GONG Jie, MA Xuecheng, ZHANG Lingling, et al. Spatiotemporal variation of habitat quality in Bailongjiang watershed in Gansu based on InVEST model [J]. Research of Soil and Water Conservation, 2018, 25(3): 191−196. [22] 张学儒, 周杰, 李梦梅. 基于土地利用格局重建的区域生境质量时空变化分析[J]. 地理学报, 2020, 75(1): 160−178. ZHANG Xueru, ZHOU Jie, LI Mengmei. Analysis on spatial and temporal changes of regional habitat quality based on the spatial pattern reconstruction of land use [J]. Acta Geographica Sinica, 2020, 75(1): 160−178. [23] 李胜鹏, 柳建玲, 林津, 等. 基于1980—2018年土地利用变化的福建省生境质量时空演变[J]. 应用生态学报, 2020, 31(12): 4080−4090. LI Shengpeng, LIU Jianling, LIN Jin, et al. Spatial and temporal evolution of habitat quality in Fujian Province, China based on the land use change from 1980 to 2018 [J]. Chinese Journal of Applied Ecology, 2020, 31(12): 4080−4090. [24] 邓越, 蒋卫国, 王文杰, 等. 城市扩张导致京津冀区域生境质量下降[J]. 生态学报, 2018, 38(12): 4516−4525. DENG Yue, JIANG Weiguo, WANG Wenjie, et al. Urban expansion led to the degradation of habitat quality in the Beijing-Tianjin-Hebei Area [J]. Acta Ecologica Sinica, 2018, 38(12): 4516−4525. [25] 王向荣. 城市自然的十点倡导[J]. 中国园林, 2021, 37(12): 2−3. WANG Xiangrong. Ten points of advocacies of urban nature [J]. Chinese Landscape Architecture, 2021, 37(12): 2−3. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20240358
 点击查看大图
点击查看大图
计量
- 文章访问数: 1923
- HTML全文浏览量: 652
- PDF下载量: 65
- 被引次数: 0
