




-
对于降雨侵蚀力相当的同一地区来说,水力侵蚀的强弱主要取决于土壤本身特性及地形、植被等影响因子。土壤抗冲性是表征土壤抗蚀能力的重要指标,它指土壤抵抗径流的机械破坏和推动下移的能力,抗力来源于土粒间、微结构间的胶结力及其抵抗离散力[1]。朱显谟[2]研究表明: 土壤抗冲性取决于根系的缠绕和固结作用,这种作用使土体有较高的水稳结构和抗蚀强度,从而不易被径流带走。近年来,由于人口剧增,陡坡毁林毁草、垦荒耕种,破坏了植物根系对土体的固结作用,导致土壤抗冲性降低,是加剧全球性土壤侵蚀、生态环境恶化的一个重要内在原因[3]。辽宁西部的建平县人工沙棘Hippophae rhamnoides林面积为6.67万hm2,是全国乃至亚洲最大的人工沙棘林[4],对保持水土、改善生态环境、促进经济发展等方面起到了积极的作用。近年来,由于自然老化,过度干旱,病虫危害,经营管理不善等原因[5-6],该区沙棘死亡率达58.5%[4, 7],生态环境遭到严重破坏。然而关于沙棘林死亡对水土保持影响的定量评价及机制仍没有进行系统的研究。本研究通过样地测试,分析沙棘林林分退化对林地根系和土壤抗冲性的影响。
-
辽宁省建平县地处辽西低山丘陵区内,属北温带大陆季风气候区,多年平均气温为8.4 ℃,年平均降水量 450.0 mm,其中6-9月占70%,是辽宁省最严重的水土流失地区之一。
试点位于建平县建平镇胡家店村红金台小流域,土壤为黄土母质发育的褐土,水土保持树种主要有沙棘Hippophae rhamnoides,榆树Ulmus pumila,油松Pinus tabulaeformis,柠条Caragana korshinskii等,属辽西低丘区的典型小流域。该流域纯沙棘林有20年生以上的植株,出现逐步死亡的现象,取而代之是榆树的入侵,表现出近似演替的特征。
-
根据植被演替研究的通用方法——时空替代法[8-10],在空间上选择纯沙棘林(未死亡,F1),榆树入侵期(沙棘仍为优势种,F2),榆树发展期(沙榆势均力敌,F3),榆树优势期(有少量沙棘,F4),纯榆树林(F5)5个标准地,3次重复,分别作为沙棘死亡及近似演替的5个阶段,标准地尺寸为20 m × 20 m,各标准地平均坡度为7°,死亡的沙棘均被农民搬运移出。各标准地活立木数量见表 1。
表 1 各标准地活立木数量
Table 1. Numbers of living trees in sample plots
样地 沙棘/株 榆树/株 沙棘死亡率/% F1 172 3 — F2 143 22 16.86 F3 83 41 51.74 F4 49 59 71.51 F5 8 76 95.35 -
在每个标准地内,挖长2.0 m,宽2.0 m,深1.0 m的土壤剖面,对根系分布进行调查,通过土壤取样并对根系进行冲洗,利用游标卡尺将根系按直径大小分为d<1 mm,1≤d<2 mm,2≤d<5 mm,5≤d<10 mm和d≥l0 mm共5个径级,统计各径级根系的条数,以1 dm2中根的条数表示根系密度。
-
①针式抗冲仪:采用NLKC-90 针式土壤抗冲仪,在0~40 cm深度范围内按照10 cm间隔,在101.325 kPa压力(1个标准大气压)下,以0.7 mm直径的水柱冲击土层1 min,使其产生水蚀穴,每10个水蚀穴深(cm)与宽(cm)乘积均值的倒数,即为该土层的抗冲指数。②槽式抗冲仪:在0~40 cm深度范围内以10 cm间隔,将每个土壤剖面用15 cm × 30 cm × 10 cm的取样器在土壤剖面取土样,带回室内置于底部有少量水的盆内浸泡12 h(水面不能高于土表面),使土柱自然吸水达到饱和状态。利用NLKC-101槽式抗冲仪将土样坡度调整为10°,在101.325 kPa水压力下对土样进行冲刷,对泥沙过滤、烘干称量,将冲刷时间除以冲刷的土量,即冲刷单位质量的土所需时间,作为抗冲性度量指标(单位为s·g-1)。
-
从各径级的根系分布情况来看,径级为1≤d<2 mm,2≤d<5 mm,5 ≤d<10 mm和d≥l0 mm的根密度分布较小,因此将根系分为直径d≥1 mm和d<1 mm等2种径级进行分析。
-
经统计(详细结果未列出),直径d≥1 mm根密度随土层深度增加逐步减少,0~40 cm土层内递减较快,在40 cm以下土层递减较慢;不同沙棘死亡率下直径d≥1 mm的根量分布差异不显著(P>0.05)。
-
直径d<1 mm根密度的分布见图 1,在0~30 cm范围内,根密度随深度增大急剧减少,30~100 cm范围内逐渐减少并维持在极低水平,总根数的72%~80%分布在0~20 cm土层,20~40 cm土层内根系仅占总根数的10%~20%。这主要是由于该区缺水非常严重,且有愈演愈烈之势;加上该区为黄土状母质上发育的淋溶褐土,表土质地粗孔隙大,而表层以下为质地较为坚硬的黄土,结构不良,渗水性能差,极易形成超渗产流;同时该区地下水埋藏极深,沙棘根系无法吸收深层地下水,其生长主要靠天然降水。因此沙棘的根系主要位于浅层土壤中,随着深度增加,根系急剧减少,这也是沙棘出现大面积死亡的原因之一。分析表明,在地表下0~30 cm深度范围内,随着沙棘死亡率的提高,根径d<1 mm的根密度呈显著的降低趋势(P<0.05),而大于30 cm的区域,差异不显著(P>0.05),表明沙棘死亡会导致表层土壤的根系分布变得稀疏。若按照植被演替的规律,沙棘死亡和榆树生长意味着植被的正向演替,根密度会随之增大[11]。然而本试验却出现了相反的现象,这主要是由于沙棘林对浅层根系分布起决定作用,而榆树数量较少且根系分布相对较深,对浅层根系起次要作用。

图 1 各样地不同深度根径d<1mm 的根密度
Figure 1. Distribution tend of root number diameter less than 1 mm in different soil depth
-
针式土壤抗冲仪测定结果见表 2。结果表明:随着沙棘死亡率的提高,土壤抗冲性呈逐步降低趋势,纯沙棘林与其他样地差异为极显著(P<0.01),除榆树优势期外,其他3个样地之间的差异均达到显著水平(P<0.05),说明沙棘死亡对土壤的抗冲性具有显著的影响;并且随着土层深度的增大,抗冲性也逐步降低。这主要是由于表层根系分布密度大和枯枝落叶层较厚,虽然土壤较疏松,但根系对土壤的固持作用较强;随着深度增加,根径d<1 mm的根密度迅速减少,致使土壤缺乏根系对集中水流的缓冲作用,冲刷作用较强[12]。
表 2 各样地针式抗冲仪测定的抗冲性
Table 2. Anti-scouring abilities with needle-type instrument
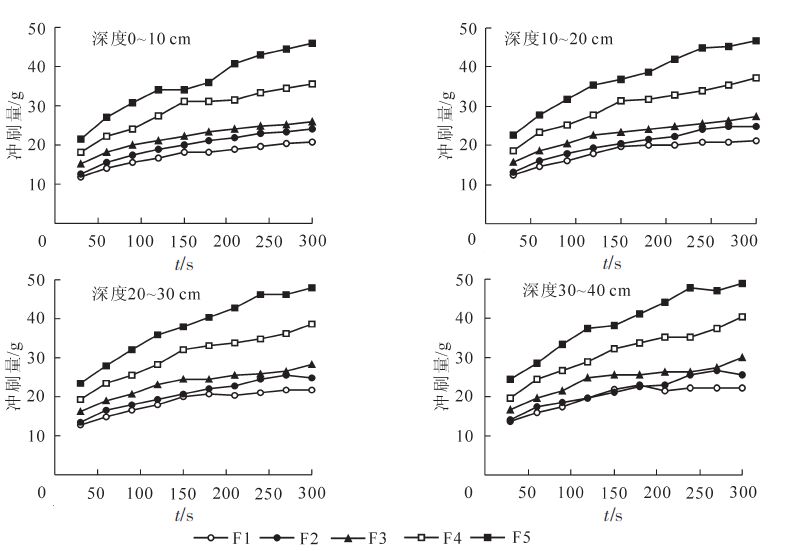
土层深度/cm 各样地针式抗冲仪测定的抗冲性/(g·cm-2mm-1) F1 F2 F3 F4 F5 0〜10 6.25±0.91 3 33±0.48 2.68±0.29 2 38±0.24 1 79±0.19 10〜20 4.00±0.75 2 98±0.35 2 14±0.27 1.89±0.21 1.56±0.20 20〜30 2.78±0.59 2 24±0.36 1 79±0.28 1.39±0.19 1.59±0.18 30〜40 2.04±0.34 1 76±0.27 1.59±0.16 1 21±0.13 1.11±0.19 平均 3.77±1.84 aA 2 58±0.71 bB 2.04±0.48 cBC 1 72±0.53 cdBC 1.51±0.29 dC 槽式抗冲仪测定的土壤冲刷量见图 2。从图 2可以看出:冲刷量随着时间的延续呈增加趋势,0~120 s之间冲刷量增加较快,随后增速变缓,至300 s时达到较平稳状态。同时随着沙棘死亡率的升高,土壤冲刷量逐步呈增加趋势,纯榆树林的冲刷量远高于其他样地,差异为极显著(P<0.01),榆树发展期与榆树入侵期之间差异不显著。通过计算单位冲刷量所需的时间,得到抗冲指数见表 3。

图 2 各样地土壤冲刷量随时间的变化
Figure 2. Relationship between soil scoured mass and time
表 3 各样地槽式抗冲仪测定的抗冲指数
Table 3. Anti-scouring abilities with trough-type instrument
深度/cm 槽式抗冲仪测定的抗冲指数/(s·g-1) F1 F2 F3 F4 F5 0〜10 14.42±1.06 12.39±1.15 11.56±1.26 8.44±1.27 6.52±0.68 10-20 14.13±0.98 12.15±1.09 10.90±1.64 8.04±1.23 6.40±0.72 20-30 13.86±1.20 12.01±1.35 10.64±1.34 7.77±1.12 6.27±0.59 30-40 13.59±2.01 11.77±1.27 10.04±1.28 7.40±1.03 6.15±0.67 平均 14.00±0.36 A 12.08±0.26 B 10.78±0.63 C 7.91±0.44 D 6.34±1.62 E 结果表明:无论是用水穴尺寸还是用冲刷量表示,随着沙棘的死亡和榆树生长,土壤抗冲性均呈逐步降低的趋势,这与周正朝等[11]植被演替增强土壤抗冲性的结论不一致。这主要由于土壤抗蚀性主要通过沙棘的根系起作用,随着沙棘的死亡,土壤中的根系数量减少,加上农民及时将死亡的沙棘移出,死亡的沙棘又无法变成腐殖质进一步保护土壤。虽然沙棘的死亡伴随着榆树的生长,单株乔木根系固结土壤和阻止径流的能力较强[13-14],然而水土流失的整体防治效果与植被结构密切相关,层次结构较好的植被对土壤的保护作用要优于单层植被[15];因此,密度较小的榆树林木对土壤的固结作用仅限于其根系周边,无法起到沙棘全面保护土壤的作用。另外,由于榆树根系较沙棘深,它更侧重于保护深层次的土壤,而土壤抗冲性主要体现在表层或近表层土壤,因此在这种情况下,榆树对土壤抗冲性的提高要逊于沙棘。
通过土壤抗冲性与根密度进行曲线拟合,表明抗冲性与根径d<1 mm根密度之间可用幂函数(y=axb)表示,与李勇等[12]的研究结论相类似。各样地参数a和b
见表 4。 表 4 土壤抗冲性(%)与根径d<1mm根密度(x)的幂函数(y=axb)参数
Table 4. Parameters for power functions between anti-scouring abilities and root density which diameter less than 1 mm
样地 针式抗冲仪 槽式抗冲仪 a b R2 P a b R2 P F1 0.53 0.40 0.99 0.003 12.68 0.022 0.96 0.008 F2 0.73 0.27 0.92 0.001 11.19 0.020 0.96 0.003 F3 1.00 0.15 0.98 0.005 8.68 0.050 0.95 0.009 F4 0.39 0.31 0.98 0.002 6.24 0.050 0.96 0.004 F5 0.84 0.14 0.92 0.010 5.82 0.020 0.96 0.020 -
通过对辽西建平县沙棘林死亡率的调查,并对不同死亡程度的林地进行土壤根系和土壤抗冲性的测定,研究沙棘林退化对水土流失的影响。结果表明:随着沙棘死亡率的提高,根径d<1 mm根密度逐步降低,沙棘死亡对较粗根系分布影响不显著;沙棘林具有较好的防治水土流失作用,随着沙棘活立木的减少,土壤抗蚀性呈逐步降低趋势,纯沙棘与其他时期差异为极显著;随着土层深度的增大,抗冲性逐步降低,土壤抗冲性与根径d<1 mm根系密度之间的关系可用幂函数表示。林地根系分布的减少和抗蚀性的减弱,水土流失使土壤结构产生破坏,因此土壤的水肥条件必然进一步恶化,从而加快沙棘的死亡,形成恶性循环。
近年来,当地林业和水土保持部门对人工沙棘林的退化现象非常重视,但一直仅就其林业方面的损失进行评估,对其引起的生态问题未进行系统的观测。本项研究从根系分布和土壤抗冲性入手,探讨沙棘退化对水土流失的影响,可以作为生态损失评估的基础。由于该区自然条件的恶劣性,加上沙棘林本身的老化现象,沙棘退化的趋势不可避免,需对其产生的影响进行全面评估,同时针对当地不良水分条件、土壤特性,以及稀疏乔木无法快速发挥水土保持效益的现状,采取措施抑制其退化速度或补植其他水土保持效果较好的灌木,将沙棘退化的生态影响降到最低。
Root system distribution and soil anti-scouring capability with Hippophae rhamnoides degradation
-
摘要: 近年来,辽宁建平县亚洲最大的人工沙棘Hippophae rhamnoides林呈现大面积死亡的现象。通过标准样地法调查沙棘的死亡程度,测定与对比不同沙棘死亡率下林地的根系密度和土壤抗冲性,研究沙棘林退化对水土流失的影响。结果表明:0~30 cm范围内,直径d<1 mm的根系密度随深度增大急剧减小,72%~80%根量分布在0~20 cm土层;0~30 cm范围内,随着沙棘死亡率的提高,d<1 mm根密度显著降低(P<0.05);随着深度增大,抗冲性也逐步降低,沙棘的死亡率对土壤抗冲性的影响亦达到显著水平(P<0.05),这主要是由根系密度的相应变化引起的;经曲线拟合,土壤抗冲性与d<1 mm根密度的关系可用幂函数表示。目前该区主要应通过采取有效措施降低沙棘林退化的生态影响。Abstract: In recent years,widespread death has occurred at the largest Asian artificial Sea Buckthorn(Hippophae rhamnoides)forest in Jianping,Liaoning. To evaluate the effects of Sea Buckthorn degradation on soil loss,the death rates of Sea Buckthorn were determined using standard plot methods with five treatments and three replications. Moreover,soil root density and soil anti-scouring capability were measured on site with soil profile methods and soil anti-scouring instruments,respectively. Results showed that soil root density for a root diameter less than 1 mm decreased at P<0.05 with an increase of soil depth within the 0-30 cm limit with 72%-80% in 0-20 cm. With a rise in the Sea Buckthorn death rate,the root density decreased significantly at P<0.05,and with an increase of soil depth, the soil anti-scouring capability decreased significantly at P<0.05. The relationship between soil anti-scouring capability and root density were fitted to power functions y=xb. The decrease of root density and soil anti-scouring capability meant damage of soil texture. Conversely,this will accelerate death rate of Sea Buckthorn,and vicious cycle format. Thus, effective measures,such as replanting shrubs,should be taken to reduce the harmful ecological effects of Sea Buckthorn degradation.
-
Key words:
- forest soil science /
- Hippophae rhamnoides /
- degradation /
- root density /
- soil anti-scouring ability /
- power function
-
图 1 各样地不同深度根径d<1mm 的根密度
Figure 1 Distribution tend of root number diameter less than 1 mm in different soil depth
表 1 各标准地活立木数量
Table 1. Numbers of living trees in sample plots
样地 沙棘/株 榆树/株 沙棘死亡率/% F1 172 3 — F2 143 22 16.86 F3 83 41 51.74 F4 49 59 71.51 F5 8 76 95.35  下载: 导出CSV
下载: 导出CSV
表 2 各样地针式抗冲仪测定的抗冲性
Table 2. Anti-scouring abilities with needle-type instrument
土层深度/cm 各样地针式抗冲仪测定的抗冲性/(g·cm-2mm-1) F1 F2 F3 F4 F5 0〜10 6.25±0.91 3 33±0.48 2.68±0.29 2 38±0.24 1 79±0.19 10〜20 4.00±0.75 2 98±0.35 2 14±0.27 1.89±0.21 1.56±0.20 20〜30 2.78±0.59 2 24±0.36 1 79±0.28 1.39±0.19 1.59±0.18 30〜40 2.04±0.34 1 76±0.27 1.59±0.16 1 21±0.13 1.11±0.19 平均 3.77±1.84 aA 2 58±0.71 bB 2.04±0.48 cBC 1 72±0.53 cdBC 1.51±0.29 dC
下载: 导出CSV
表 3 各样地槽式抗冲仪测定的抗冲指数
Table 3. Anti-scouring abilities with trough-type instrument
深度/cm 槽式抗冲仪测定的抗冲指数/(s·g-1) F1 F2 F3 F4 F5 0〜10 14.42±1.06 12.39±1.15 11.56±1.26 8.44±1.27 6.52±0.68 10-20 14.13±0.98 12.15±1.09 10.90±1.64 8.04±1.23 6.40±0.72 20-30 13.86±1.20 12.01±1.35 10.64±1.34 7.77±1.12 6.27±0.59 30-40 13.59±2.01 11.77±1.27 10.04±1.28 7.40±1.03 6.15±0.67 平均 14.00±0.36 A 12.08±0.26 B 10.78±0.63 C 7.91±0.44 D 6.34±1.62 E
下载: 导出CSV
表 4 土壤抗冲性(%)与根径d<1mm根密度(x)的幂函数(y=axb)参数
Table 4. Parameters for power functions between anti-scouring abilities and root density which diameter less than 1 mm
样地 针式抗冲仪 槽式抗冲仪 a b R2 P a b R2 P F1 0.53 0.40 0.99 0.003 12.68 0.022 0.96 0.008 F2 0.73 0.27 0.92 0.001 11.19 0.020 0.96 0.003 F3 1.00 0.15 0.98 0.005 8.68 0.050 0.95 0.009 F4 0.39 0.31 0.98 0.002 6.24 0.050 0.96 0.004 F5 0.84 0.14 0.92 0.010 5.82 0.020 0.96 0.020
下载: 导出CSV
-
[1] 查小春,贺秀斌. 土壤物理力学性质与侵蚀关系研究进展[J]. 水土保持研究,1999,6(2):98-104. ZHA Xiaochun,HE Xiubin. Progress on the study of soil erosion from soil physical property and erosive force[J]. Res Soil Water Convers,1999,6(2):98-104. [2] 朱显谟. 黄土高原植被因素对于水土流失的影响[J]. 土壤学报,1960,8(2):110-120. ZHU Xianmo. Effect of vegetation on soil and water loss in Loess Plateau[J]. Acta Pedol Sin,1960,8(2):110-120. [3] 李勇,徐晓琴,朱显谟,等. 植物根系与土壤抗冲性[J]. 水土保持学报,1993,7(3):11-18. LI Yong,XU Xiaoqin,ZHU Xianmo,et al. Plant roots and anti-scouring ability[J]. J Soil Water Conserv,1993,7(3):11-18. [4] 宫海志,王军. 建平县沙棘林大面积死亡成因分析及经营对策[J]. 中国水土保持,2003(4):17-18. GONG Haizhi,WANG Jun. Analysis on reasons of large-acreage death of Seabuckthorn forest and its operation strategies in Jianping County[J]. Chin Soil Water Conserv,2003(4):17-18. [5] 李树彬,党福江. 建平县沙棘林大面积死亡原因调查分析[J]. 水土保持科技情报,2001(6):4-6. LI Shubin,DANG Fujiang. Investigation and analysis on reasons of large-acreage death of Seabuckthorn forest in Jianping County[J]. Sci Technol Inform Soil Water Conserv,2001(6):4-6. [6] 惠兴学,张连翔,孔繁轼,等. 建平县沙棘林大面积死亡成因调查分析及对策[J]. 防护林科技,2002(2):53-55. HUI Xingxue, ZHANG Lianxiang,KONG Fanshi,et al. Investigation and analysis on reasons of large-acreage death of Seabuckthorn forest and its control in Jianping County[J]. Prot For Sci Technol,2002(2):53-55. [7] 张连翔,惠兴学,黄立华,等. 建平县沙棘林大面积死亡原因及其治理对策[J]. 沙棘,2002,15(3):26-29. ZHANG Lianxiang,HUI Xingxue,HUANG Lihua,et al. A large-acreage death of Seabuckthorn forest and its control in Jianping County[J]. Hippophae,2002,15(3):26-29. [8] 闫芊,何文珊,陆健健. 崇明东滩湿地植被演替过程中生物量与氮含量的时空变化[J]. 生态学杂志,2006,25(9):1019-1023. YAN Qian,HE Wenshan,LU Jianjian. Temporal and spatial changes of plant biomass and its N content during vegetation succession at Chongming Dongtan wetland[J]. Chin J Ecol,2006,25(9):1019-1023. [9] 宋洪涛,张劲峰,田昆,等. 滇西北亚高山地区黄背栎林植被演替过程中的林地土壤化学响应[J]. 西部林业科学,2007,36(2):65-70. SONG Hongtao,ZHANG Jinfeng,TIAN Kun,et al. Response of soil chemical properties to vegetation succession of Quercus pannosa forest in subalpine area of northwest Yunnan[J]. J West China For Sci,2007,36(2):65-70. [10] 王韵,王克林,邹冬生,等. 广西喀斯特地区植被演替对土壤质量的影响[J]. 水土保持学报,2007,21(6):130-134. WANG Yun,WANG Kelin,ZOU Dongsheng,et al. Effects of vegetation succession on soil quality in Karst region of Guangxi,China[J]. J Soil Water Conserv,2007,21(6):130-134. [11] 周正朝,上官周平. 子午岭次生林植被演替过程的土壤抗冲性[J]. 生态学报,2006,26(10):3270-3275. ZHOU Zhengchao,SHANGGUAN Zhouping. Soil anti-scouring ability during vegetation succession of Ziwuling secondary forest[J]. Acta Ecol Sin,2006,26(10):3270-3275. [12] 李勇,徐晓琴,朱显谟,等. 草类根系对土壤抗冲性的强化效应[J]. 土壤学报,1992,29(3):302-309. LI Yong,XU Xiaoqin,ZHU Xianmo,et al. Strengthening effects of grass root on soil anti-scouring ability[J]. Acta Pedol Sin,1992,29(3):302-309. [13] DILLAHA T A. Vegetative filter strips for agricultural nonpoint-source pollution control[J]. Tran ASAE,1989,32(3):513-519. [14] CHAUBEY I,EDWARDS D R,DANIEL T C,et al. Effectiveness of vegetative filter strips in controlling losses of surface-applied poultry litters constituents[J]. Tran ASAE,1995,38(6):1687-1692. [15] GONZALEZ H J C,RAVENTOS J,ECHEVARRIA M T. Comparison of sediment ratio curves for plants with different architectures[J]. Catena,1997,29(3/4):333-340. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2014.03.008
 点击查看大图
点击查看大图

计量
- 文章访问数: 4417
- HTML全文浏览量: 852
- PDF下载量: 524
- 被引次数: 0
