




-
近自然森林经营理论以抚育、间伐、择伐和天然更新等措施为主要技术特征,追求森林资源的永续利用[1]。近自然经营管理措施下森林土壤肥力的变化是评价该森林经营方式是否有利于森林可持续经营的重要基础。近自然经营对林下土壤肥力的影响国内外学者已有相关研究。张鼎华等[2]研究发现:近自然经营马尾松 Pinus massoniana 幼林的土壤养分含量增加、生物活性增强、土壤酶活性增加、土壤交换性能改善,并且立地条件差的变化幅度较大。吴瑶等[3]应用近自然森林技术对红松Pinus koraiensis林土壤影响的研究中发现:土壤有机碳、全氮、全磷、碱解氮、速效磷、速效钾的质量分数高于未经营林分。刘延滨等[4]对退化落叶松Larix gmelinii人工林近自然改造后发现:林分土壤微生物多样性增加,有效磷、有效钾增加。颜欢欢等[5]通过近自然与常规经营千年桐Vernicia montana林的对比发现,近自然经营的千年桐生态林土壤持水量、氮磷钾、有机质含量都高于常规千年桐林。但王新宇等[6]在水曲柳Fraxinus mandshurica和长白落叶松Larix olgansis人工林近自然经营中发现:近自然抚育后土壤容重、孔隙度、含水量无显著差异。徐庆祥等[7]也得出了相似结论。目前,关于近自然经营对土壤物理性质影响的研究存在一定差异,近自然经营对土壤肥力影响的研究侧重于土壤肥力某一方面的性质,没有综合各土壤肥力指标而考虑林分肥力变化。近自然经营不同措施对不同龄林马尾松林土壤肥力的影响还没有相关报道。本研究针对这些问题,选取了近自然不同经营措施马尾松林土壤肥力的物理、化学、生物学指标,以全面准确评价林分土壤肥力的变化,为近自然森林经营理论在中国的推广提供理论依据。
-
研究地点位于贵州省贵阳市开阳县近自然经营项目区(26°48′~27°22′N,106°45′~107°17′E, 平均海拔高度1 000~1 400 m)。地形以山地为主,生态环境良好,降雨充沛,属亚热带常绿阔叶林带,主要的植物属壳斗科Fagaceae、樟科Lauraceae、山茶科Theaceae 、冬青科Aquifoliaceae等,主要乔木有马尾松、华山松Pinus armandii、杉木Cunninghamia lanceolata、青冈Quercus glauca、白杨Populus tomentosa、光皮桦Betula luminifera、枫香Liquidambar formosana、樟树Cinnamomum camphora等;林下主要灌木有杜鹃Rhododendron simsii、马桑Coriaria nepalensis、火棘Pyracantha fortuneana、铁仔Myrsine africana等。林内湿度较大。近地层苔藓和地衣发达。项目区优势树种为马尾松,主要涉及幼龄林、中龄林、近熟林3个龄组,未进行近自然经营前林分郁闭度均达0.8 以上。林分土壤类型以黄壤、黄棕壤为主。近自然抚育起始时间为2010年,对象为9年生马尾松幼龄林:对林分进行修枝、补植、伐除,对目标树有竞争的乔木和目标树附近高大的灌木和草本进行清除,人工辅助调节林木水平分布和树种组成,改善林分空间,实现林分混交,不定期进行;近自然间伐在2013年开展,对象为18年生马尾松中龄林:对林分进行单株间伐,伐除对目标树有干扰和林分密集区域长势差的乔木,实施1次,强度为39.18%;近自然择伐在2013年实施,对象为22年生马尾松近熟林:对林分进行单株择伐,以目标树为核心,伐除竞争木及干扰木,无目标树区域调节林分密度,增加林中空隙,择伐“霸王树”,实施1次,强度为37.64%。研究区择伐和间伐林分约740 hm2,抚育约670 hm2。
-
2017年11月,依开阳县林班分布图,在开阳县近自然经营项目区分别针对幼龄林抚育、中龄林间伐、近熟林择伐设置立地条件相似的样地。各设置3组重复及对照,对照为项目区未进行任何经营的与相应龄组立地条件相似的马尾松林分。在每个林分中部设置20 m×20 m的大样方,并在每个大样方的对角线上选取3个20 cm×20 cm的小样方,采用环刀进行取土,取土厚度为20 cm。如此进行土壤样品采集,以充分全面了解林分土壤肥力情况。对采集的土壤样品各项土壤指标进行测定分析[8],pH值采用电位法;有机质质量分数采用重铬酸钾容量法;全氮质量分数采用混合盐消化碱扩散法;全磷采用铝锑抗比色法;全钾采用火焰光度计法;碱解氮采用碱解扩散法;速效磷采用碳酸氢钠浸提-钼锑抗比色法;速效钾采用火焰光度法;容重采用环刀法;含水量采用酒精燃烧法;总孔隙度采用环刀法;脲酶采用比色法;蔗糖酶采用3,5-二硝基水杨酸比色法;碱性磷酸酶采用磷酸苯二钠比色法。
-
由于各土壤肥力指标对土壤整体肥力水平的影响不同且数值存在量纲差距,所以会严重影响对土壤肥力评价和分析的准确性。为使评价更合理有效,基于模糊数学原理,构建土壤肥力指标的隶属度函数。根据植物效应曲线将隶属度函数分为2种类型[9]。相应函数表达式如下:
$$ f(x) = \left\{ {\begin{array}{*{20}{l}} {1.0,x \geqslant x_2}\\ {\dfrac{{0.9(x - x_1)}}{{x_2 - x_1}} + 0.1,x_1 \leqslant x \leqslant x_2}\\ {0.1,\;\;\;\;x < x_1} \end{array}} \right.\!\!\!\!{\text{;}} \quad f(x) = \left\{ {\begin{array}{*{20}{l}} {0.1,x < x_1\;{\text{或}}\;x \geqslant x_4}\\ {\dfrac{{0.9(x - x_1)}}{{x_2 - x_1}} + 0.1,x_1 \leqslant x \leqslant x_2}\\ {1.0,x_2 < x < x_3}\\ {1.0 - \dfrac{{0.9(x - x_3)}}{{x_4 - {x_3}}},x_3 < x \leqslant x_4} \end{array}} \right. \text{。} $$ (1) 式(1)中:x1、x2、x3、x4为土壤肥力指标在曲线中的转折点,通过公式计算出各土壤肥力指标的隶属度值,其值越大反映该指标水平越适宜马尾松生长,这些值在0.1~1.0之内。结合马尾松对于各项土壤肥力指标丰缺的生理特点,以及《全国第2次土壤普查暂行规程》的土壤养分含量分级和当地的具体实际,以及前人的研究情况[9-17],确定函数中转折点的相应取值如表1所示。
表 1 马尾松隶属度函数转折点取值
Table 1. Value of the turning point of the membership function of P. massoniana
转折点 有机质/(g·kg−1) 全氮/(g·kg−1) 碱解氮/(mg·kg−1) 全钾/(g·kg−1) 速效钾/(mg·kg−1) 全磷/(g·kg−1) 速效磷/(mg·kg−1) x1 6.00 0.50 30.00 5.00 30.00 0.20 3.00 x2 40.00 2.00 150.00 25.00 200.00 1.00 40.00 转折点 pH 脲酶/(μg·g−1) 蔗糖酶/(mg·g−1) 碱性磷酸酶/(mg·g−1) 容重/(g·cm−3) 含水量/% 总孔隙度/% x1 3.00 0.11 20.21 1.25 0.82 1.00 60.00 x2 3.50 1.28 39.88 1.85 1.00 17.00 70.00 x3 5.50 1.20 x4 7.50 1.40 -
采用SPSS 24.0统计分析软件对近自然经营与对照各土壤肥力指标进行独立样本t检验,以分析近自然经营与对照土壤肥力各项指标差异的显著程度。采用Excel 2016对消除量纲后的各指标做雷达图,以直观全面地了解研究区各土壤肥力指标变化的趋势及丰缺情况,结合SPSS 24对各项土壤肥力指标进行主成分分析,以综合评价土壤肥力。主成分分析提取了3个主成分(表2),每一个主成分都综合反映了原所有土壤肥力指标的大部分信息。通过各土壤类型主成分,利用公式计算各近自然经营措施土壤肥力综合评价得分。公式为:综合评价得分=W1Y1+W2Y2+W3Y3,其中:W1、W2、W3为各成分特征值权重进行归一化处理后的值,Y1、Y2、Y3为土壤肥力指标主成分。
表 2 土壤肥力指标主成分
Table 2. Main components of soil fertility index
土壤类型 Y1 Y2 Y3 间伐 1.510 174 4 0.226 733 5 0.914 857 4 间伐对照 0.934 324 4 −0.216 028 0 0.526 200 0 择伐 1.118 469 3 0.975 322 3 0.743 295 2 择伐对照 0.229 793 4 0.564 344 2 0.445 253 2 抚育 1.522 134 7 0.480 725 5 0.197 184 3 抚育对照 1.522 071 5 0.548 271 1 0.298 588 1 -
对林分而言,容重应在1.0~1.4为优[16]。由表3可知:间伐与择伐土壤容重都优于对照,其中择伐土壤容重显著大于对照,差异显著(P<0.05);抚育土壤容重小于对照,差异不显著(P>0.05)。间伐与择伐的含水量都大于对照,差异显著(P<0.05);抚育土壤含水量小于对照,差异不显著(P>0.05)。择伐与间伐的总孔隙度均小于对照,差异显著(P<0.05);抚育小于对照,差异不显著(P>0.05)。
表 3 近自然不同经营与对照的土壤物理指标比较
Table 3. Comparison of soil physical indicators between different near-natural management and controls
土壤类型 容重/(g·cm−3) 含水量/% 总孔隙度/% 间伐 1.00 16* 63* 间伐对照 0.97 13 69 择伐 1.01* 15* 61* 择伐对照 0.83 12 69 抚育 1.03 15 61 抚育对照 1.04 15 62 说明:用 t 检验法进行分析。*表示同列试验组与对 照组差异显著 (P<0.05) -
根据柳若安等[17]对马尾松林土壤酸度的研究,适宜马尾松生长的土壤pH为3.5~5.5。由表4可知:各经营措施与对照林分土壤酸度均处于优良状态,且差异不显著(P>0.05)。择伐与间伐的全氮、碱解氮、全钾、全磷、速效磷质量分数均大于对照,且差异显著(P<0.05)。土壤有机质质量分数近自然择伐大于对照,差异显著(P<0.05);间伐大于对照,差异不显著(P>0.05)。对于抚育,有机质、全氮、碱解氮、全钾、速效钾、全磷质量分数与对照相比,差异不显著(P>0.05),其中碱解氮和全钾差异显著(P<0.05)。
表 4 近自然不同经营管理与对照土壤化学指标的比较
Table 4. Comparison of soil chemical and fertility indicators between different near-natural management and control
土壤类型 有机质/
(g·kg−1)全氮/
(g·kg−1)碱解氮/
(mg·kg−1)全钾/
(mg·kg−1)速效钾/
(mg·kg−1)全磷/
(mg·kg−1)速效磷/
(g·kg−1)pH 间伐 22.70 0.61* 146.69* 447.90* 60.22* 320.73* 5.03* 4.56 间伐对照 22.38 0.46 79.91 408.87 58.30 296.56 4.10 4.55 择伐 19.36* 0.68* 119.01* 532.51* 59.88 200.50* 3.75* 4.65 择伐对照 11.89 0.32 79.94 386.70 59.91 140.94 2.46 4.64 抚育 21.99 0.77 114.32* 475.70* 59.98 245.60 5.40 4.60 抚育对照 22.19 0.71 126.74 505.67 59.90 245.73 5.21 4.63 说明:用t检验法进行分析。*表示同列试验组与对照组差异显著 (P<0.05) -
由表5可知:研究区择伐和间伐蔗糖酶和碱性磷酸酶质量分数高于对照,差异显著(P<0.05)。土壤脲酶质量分数间伐小于对照,差异不显著(P>0.05);择伐明显大于对照,差异显著(P<0.05)。抚育马尾松林分蔗糖酶质量分数低于对照,差异显著(P<0.05),碱性磷酸酶差异不显著(P>0.05)。
表 5 近自然不同经营管理与对照土壤生物指标的比较
Table 5. Comparison of soilbio-indicators between different near-natural management and control
土壤类型 脲酶/
(μg·g−1)蔗糖酶/
(mg·g−1)碱性磷酸酶/
(mg·g−1)间伐 1.16 36.42* 1.44* 间伐对照 1.25 34.36 1.28 择伐 0.93* 39.75* 1.83* 择伐对照 0.13 38.22 1.64 抚育 1.08* 20.23* 1.52 抚育对照 1.01 22.23 1.52 说明:用t检验法进行分析。*表示同列试验组与对照组差 异显著 (P<0.05) -
由图1可以看出:研究区采取择伐的经营措施后,土壤全氮、碱解氮、全钾、全磷、速效磷、脲酶、碱性磷酸酶质量分数都有明显的增长,土壤物理指标的变化趋势不明显。间伐的土壤碱解氮、全钾、全磷、速效磷、蔗糖酶质量分数有增长趋势,脲酶质量分数有减少趋势。抚育的碱解氮、全钾、速效磷、脲酶质量分数有减少趋势,蔗糖酶质量分数有增加趋势。间伐与择伐各土壤肥力指标所构成面积明显大于对照,抚育与对照各土壤肥力指标在雷达图中构成的面积相近,结合表6中间伐与择伐土壤肥力综合评价得分均高于对照,差异显著(P<0.05),抚育与对照得分差异不显著(P>0.05),可以断定近自然间伐与择伐土壤肥力优于对照,近自然抚育与对照土壤肥力相近。
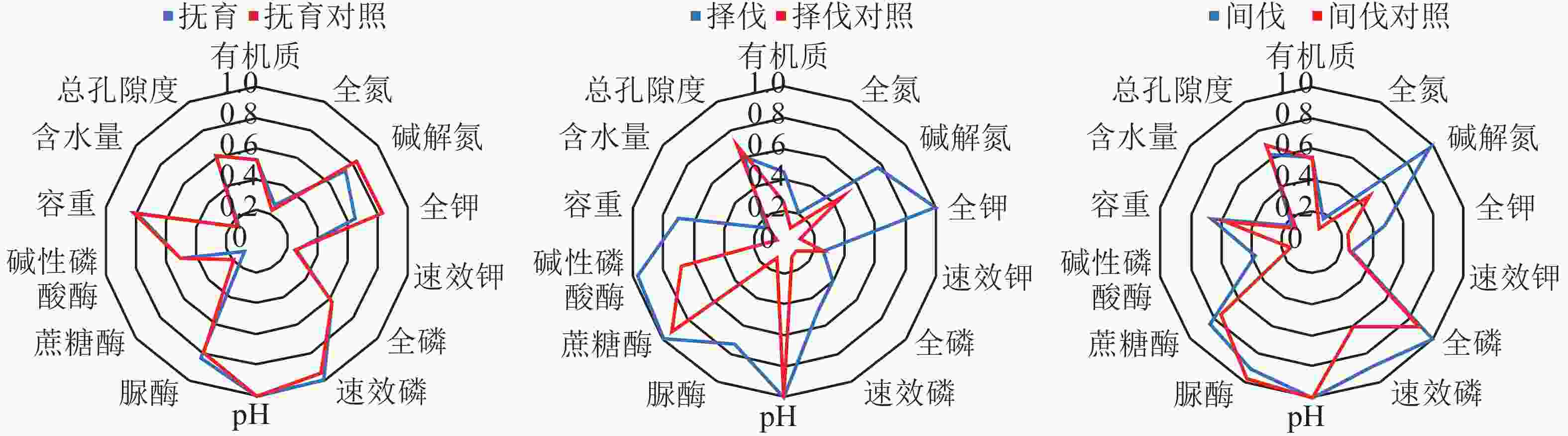
图 1 近自然与对照土壤肥力指标隶属度雷达图
Figure 1. Radar diagram of the membership degree of near-natural and control soil fertility indicators
表 6 近自然与对照主成分分析综合得分
Table 6. Comprehensive scores of near-natural and control principal component analysis
土壤类型 综合得分 间伐 1.07* 间伐对照 0.55 择伐 1.04* 择伐对照 0.35 抚育 1.09 抚育对照 1.12 说明:用t检验法进行分析。*表示同列试验组与对照组差 异显著 (P<0.05) -
研究区经过近自然经营后,近自然择伐的马尾松近熟林的土壤肥力发生了显著变化,主要体现在提高土壤养分和土壤生物酶活性上,其中土壤有机质、全氮、碱解氮、全钾、全磷、速效磷、脲酶、蔗糖酶、碱性磷酸酶质量分数均显著提升。近自然择伐对林分进行单株择伐,以目标树为核心,伐除竞争木及干扰木,无目标树区域调节林分密度,增加林中空隙,择伐“霸王树”。这些措施能够减少林分林木数量,增加林木生长空间,进而给幼小植株提供更多养分,同时增加林内光照和改善温度条件,进而增加低矮植物获得光照的强度和时间,调节林分整体结构,促进林分更新,加速了林内生物小循环,因此土壤化学和生物指标得到了提升。
近自然间伐的马尾松中龄林土壤肥力也发生了显著变化,其中土壤全氮、碱解氮、全钾、速效钾、全磷、脲酶、蔗糖酶、碱性磷酸酶、速效磷质量分数得到了显著提升。近自然间伐对林分进行单株间伐,伐除对目标树有干扰和林分密集区长势差的乔木,使得林分保留木生长空间扩大,促进了实施后几年林木生长,从而森林更新加快,林下生物多样性提高,加速了土壤物质循环和养分周转,因此提升了土壤肥力。吴瑶等[3]、刘延滨等[4]、包甜甜等[18]、刘俊钦[19]在近自然森林经营的研究中也得出了类似结论。
近自然抚育幼龄马尾松林土壤肥力变化不显著。在物理性质方面,近自然各经营措施对林分土壤的改良是间接作用,并没有直接改变土壤结构,因此土壤物理指标的变化不显著。这与王新宇等[6]、徐庆祥等[7]在近自然抚育后土壤容重、孔隙度、含水量等物理性质方面没有显著差异的研究结果相似。近自然抚育马尾松幼龄林土壤养分和生物酶质量分数变化不明显。本研究中,近自然抚育对林分进行修枝、伐除对目标树有竞争的乔木,对目标树附近高大的灌木和草本进行清除,抚育剩余物全面清理,每次对林分结构的调控影响较小。这些措施在研究区经营年限里对林分的影响可能还未体现。张鼎华等[2]、包甜甜等[18]在近自然幼林经营的研究中对修剪的枝干,割除的灌木和草本进行埋蔸,起到了提升土壤养分的作用,因此,对近自然抚育剩余物的不同处理方式可能对土壤养分有不同影响。另外,林分立地条件、林分年龄以及经营年限的不同也可能导致近自然抚育对土壤肥力的影响不同。近自然抚育对土壤肥力的影响还有待进一步研究。
整体而言,近自然不同经营管理措施确实能够改造林分结构,改善林境,促进森林更新,提高林下生物多样性,进而加速土壤物质循环和养分周转,影响土壤肥力的化学和生物指标,从而提高土壤养分和土壤生物酶活性,提高土壤肥力。土壤肥力的提升必然能够促进林分的生长,提高森林质量,这对林分的可持续经营有重要作用。
-
采用近自然择伐与近自然间伐等措施,马尾松林分土壤肥力得到了显著的改良,土壤肥力与未进行近自然经营的对照林分差异显著,其中近自然择伐效果最显著。近自然抚育对土壤肥力物理指标影响不显著。近自然间伐与择伐对马尾松林分土壤肥力的影响具有显著的促进作用,对马尾松林分土壤肥力的维护和提高有重要作用。近自然经营理论对贵阳市森林土壤肥力的提高和森林可持续经营有重要作用。
Soil fertility of Pinus massoniana forests under different near-natural management measures
-
摘要:
目的 研究近自然不同经营管理措施对马尾松Pinus massoniana林下土壤肥力的影响。 方法 在贵阳市近自然经营开阳项目区分别选取马尾松林近自然择伐、近自然间伐、近自然抚育及对照组,综合评价不同管理措施对马尾松林下土壤肥力的影响。 结果 近自然择伐的土壤有机质、全氮、碱解氮、全钾、全磷、速效磷、脲酶、碱性磷酸酶质量分数都高于对照,呈增加趋势,其中有机质、全氮、碱解氮、全钾、全磷、速效磷的质量分数差异显著(P<0.05);近自然间伐的土壤有机质、全氮、碱解氮、全钾、速效钾、全磷、速效磷、蔗糖酶质量分数都高于对照,呈增加趋势,其中全氮、碱解氮、全钾、速效钾、全磷、速效磷的质量分数差异显著(P<0.05),但脲酶质量分数呈减少趋势;近自然抚育的土壤碱解氮、全钾、速效磷、脲酶质量分数呈减少趋势。其中碱解氮和全钾差异显著(P<0.05),蔗糖酶质量分数有增加趋势。近自然间伐和择伐马尾松林分土壤肥力的综合评价得分与相应对照差异显著(P<0.05),抚育差异不显著(P>0.05);近自然不同管理措施对土壤物理指标的影响不显著(P>0.05)。 结论 近自然间伐与择伐对马尾松林分土壤肥力的影响具有显著的促进作用,对马尾松林分土壤肥力的维护和提高有重要意义;近自然抚育对林分影响不显著。图1表6参19 Abstract:Objective The research aims to study the effects of different near-natural management measures on soil fertility under Pinus massoniana forests. Method In Kaiyang near-natural management project area of Guiyang City, the near-natural selective cutting, near-natural thinning, near-natural tending, and the control group were selected to comprehensively evaluate the impact of different management measures on soil fertility under P. massoniana forests. Result The mass fractions of organic matter, soil total nitrogen, alkali-hydrolyzed nitrogen, total potassium, total phosphorus, available phosphorus, urease, alkaline phosphatase in near-natural selective cutting were higher than those of the control group, showing an up-going trend. The mass fractions of organic matter, total nitrogen, alkali-hydrolyzed nitrogen, total potassium, total phosphorus, and available phosphorus were significantly different(P<0.05). The mass fraction of soil organic matter, total nitrogen, alkali-hydrolyzed nitrogen, total potassium, available potassium, total phosphorus, available phosphorus, and sucrose in near-natural thinning was higher than that in the control group, showing an increasing trend, among which the mass fraction of total nitrogen, alkali-hydrolyzed nitrogen, total potassium, available potassium, total phosphorus and available phosphorus had significant differences (P<0.05), but the mass fraction of urease showed a decreasing trend. The mass fraction of alkali-hydrolyzed nitrogen, total potassium, available phosphorus and urease in near-natural tending showed a decreasing trend, among which the difference between alkali-hydrolyzed nitrogen and total potassium was significant (P<0.05), and the mass fraction of sucrose showed an increasing trend. The comprehensive evaluation scores of soil fertility in the P. massoniana stands with near-natural thinning and selective cutting were significantly different from those of the control group (P<0.05), but the difference in tending was not significant (P>0.05). The effect of different near-natural management measures on soil physical indexes was not significant (P>0.05). Conclusion Near-natural thinning and selective cutting can significantly promote soil fertility of P. massoniana forests, which is of great significance for maintenance and improvement of soil fertility of P. massoniana forests. The effect of near-natural tending on forest stands is not significant. The near-natural management theory plays an important role in improving forest soil fertility and sustainable forest management in Guiyang. [Ch, 1 fig. 6 tab. 19 ref.] -
图 1 近自然与对照土壤肥力指标隶属度雷达图
Figure 1 Radar diagram of the membership degree of near-natural and control soil fertility indicators
表 1 马尾松隶属度函数转折点取值
Table 1. Value of the turning point of the membership function of P. massoniana
转折点 有机质/(g·kg−1) 全氮/(g·kg−1) 碱解氮/(mg·kg−1) 全钾/(g·kg−1) 速效钾/(mg·kg−1) 全磷/(g·kg−1) 速效磷/(mg·kg−1) x1 6.00 0.50 30.00 5.00 30.00 0.20 3.00 x2 40.00 2.00 150.00 25.00 200.00 1.00 40.00 转折点 pH 脲酶/(μg·g−1) 蔗糖酶/(mg·g−1) 碱性磷酸酶/(mg·g−1) 容重/(g·cm−3) 含水量/% 总孔隙度/% x1 3.00 0.11 20.21 1.25 0.82 1.00 60.00 x2 3.50 1.28 39.88 1.85 1.00 17.00 70.00 x3 5.50 1.20 x4 7.50 1.40  下载: 导出CSV
下载: 导出CSV
表 2 土壤肥力指标主成分
Table 2. Main components of soil fertility index
土壤类型 Y1 Y2 Y3 间伐 1.510 174 4 0.226 733 5 0.914 857 4 间伐对照 0.934 324 4 −0.216 028 0 0.526 200 0 择伐 1.118 469 3 0.975 322 3 0.743 295 2 择伐对照 0.229 793 4 0.564 344 2 0.445 253 2 抚育 1.522 134 7 0.480 725 5 0.197 184 3 抚育对照 1.522 071 5 0.548 271 1 0.298 588 1
下载: 导出CSV
表 3 近自然不同经营与对照的土壤物理指标比较
Table 3. Comparison of soil physical indicators between different near-natural management and controls
土壤类型 容重/(g·cm−3) 含水量/% 总孔隙度/% 间伐 1.00 16* 63* 间伐对照 0.97 13 69 择伐 1.01* 15* 61* 择伐对照 0.83 12 69 抚育 1.03 15 61 抚育对照 1.04 15 62 说明:用 t 检验法进行分析。*表示同列试验组与对 照组差异显著 (P<0.05)
下载: 导出CSV
表 4 近自然不同经营管理与对照土壤化学指标的比较
Table 4. Comparison of soil chemical and fertility indicators between different near-natural management and control
土壤类型 有机质/
(g·kg−1)全氮/
(g·kg−1)碱解氮/
(mg·kg−1)全钾/
(mg·kg−1)速效钾/
(mg·kg−1)全磷/
(mg·kg−1)速效磷/
(g·kg−1)pH 间伐 22.70 0.61* 146.69* 447.90* 60.22* 320.73* 5.03* 4.56 间伐对照 22.38 0.46 79.91 408.87 58.30 296.56 4.10 4.55 择伐 19.36* 0.68* 119.01* 532.51* 59.88 200.50* 3.75* 4.65 择伐对照 11.89 0.32 79.94 386.70 59.91 140.94 2.46 4.64 抚育 21.99 0.77 114.32* 475.70* 59.98 245.60 5.40 4.60 抚育对照 22.19 0.71 126.74 505.67 59.90 245.73 5.21 4.63 说明:用t检验法进行分析。*表示同列试验组与对照组差异显著 (P<0.05)
下载: 导出CSV
表 5 近自然不同经营管理与对照土壤生物指标的比较
Table 5. Comparison of soilbio-indicators between different near-natural management and control
土壤类型 脲酶/
(μg·g−1)蔗糖酶/
(mg·g−1)碱性磷酸酶/
(mg·g−1)间伐 1.16 36.42* 1.44* 间伐对照 1.25 34.36 1.28 择伐 0.93* 39.75* 1.83* 择伐对照 0.13 38.22 1.64 抚育 1.08* 20.23* 1.52 抚育对照 1.01 22.23 1.52 说明:用t检验法进行分析。*表示同列试验组与对照组差 异显著 (P<0.05)
下载: 导出CSV
表 6 近自然与对照主成分分析综合得分
Table 6. Comprehensive scores of near-natural and control principal component analysis
土壤类型 综合得分 间伐 1.07* 间伐对照 0.55 择伐 1.04* 择伐对照 0.35 抚育 1.09 抚育对照 1.12 说明:用t检验法进行分析。*表示同列试验组与对照组差 异显著 (P<0.05)
下载: 导出CSV
-
[1] 贺志龙, 张芸香, 郭晋平. 我国近自然森林经营技术与效果评价研究进展[J]. 山西农业科学, 2017, 45(9): 1566 − 1570, 1582. HE Zhilong, ZHANG Yunxiang, GUO Jinping. Research progress on management technology and effect evaluation of near-natural forestry management in China [J]. Shanxi Agric Sci, 2017, 45(9): 1566 − 1570, 1582. [2] 张鼎华, 林卿. 近自然林业经营法在马尾松人工幼林经营中的应用[C]//中国农学会. 2001年生态农业与可持续发展国际研讨会论文集. 北京: 中国农业出版社, 2001: 277 − 282. [3] 吴瑶, 李凤日, 秦凯伦, 等. 近自然经营技术对红松林土壤化学性质的影响[J]. 东北林业大学学报, 2014, 42(1): 76 − 79. WU Yao, LI Fengri, QIN Kailun, et al. Impacts of close-to-nature management on soil chemical properties of Korean pine forest in northeastern China [J]. J Northeast For Univ, 2014, 42(1): 76 − 79. [4] 刘延滨, 王庆成, 王承义, 等. 退化落叶松人工林近自然化改造对土壤微生物及养分的影响[J]. 生态学杂志, 2012, 31(11): 2716 − 2722. LIU Yanbin, WANG Qingcheng, WANG Chengyi, et al. Effects of close-to-nature transformation of degenerated larch (Larix gmelinii) plantation on soil microbes and soil nutrients [J]. Chin J Ecol, 2012, 31(11): 2716 − 2722. [5] 颜欢欢, 吴承祯, 洪伟, 等. 千年桐生态林近自然经营对土壤酶活性的影响[J]. 西南林业大学学报, 2014, 34(3): 1 − 6. YAN Huanhuan, WU Chengzhen, HONG Wei, et al. Effects of intimate natural management of Aleurites montana eco-forests on activity of soil enzymes [J]. J Southwest For Univ, 2014, 34(3): 1 − 6. [6] 王新宇, 王庆成. 水曲柳落叶松人工林近自然化培育对林地土壤理化性质的影响[J]. 林业科学, 2008, 44(12): 21 − 27. WANG Xinyu, WANG Qingcheng. Effects of the close-to-nature cultivation of Larix olgensis and Fraxinus mandshurica on the soil physiochemical properties [J]. Sci Silv Sin, 2008, 44(12): 21 − 27. [7] 徐庆祥, 卫星, 王庆成, 等. 抚育间伐对兴安落叶松天然林生长和土壤理化性质的影响[J]. 森林工程, 2013, 29(3): 6 − 9. XU Qingxiang, WEIXing, WANG Qingcheng, et al. Impact of thinning on growth and soil properties of natural Larix gmelinii forest [J]. For Eng, 2013, 29(3): 6 − 9. [8] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000. [9] 王玲玲, 徐福利, 王渭玲, 等. 不同林龄华北落叶松人工林地土壤肥力评价[J]. 西南林业大学学报, 2016, 36(2): 17 − 24. WANG Lingling, XU Fuli, WANG Weiling, et al. Assessment of soil fertility in different aged Larix principis-rupprechtii plantation [J]. J Southwest For Univ, 2016, 36(2): 17 − 24. [10] LUO Mei. The Purpose and theoretic foundation of fertilizing in masson pine forest[C]// DAVID W. Proceedings of the Intelligent Information Technology Application Society Conference. Singapore: Agricultural and Natural Resources Engineering, 2011: 413 − 420. [11] 刘洪鹄, 赵玉明, 王秀颖, 等. 土壤肥力评价方法探讨[J]. 长江科学院院报, 2008, 25(3): 62 − 66. LIU Honghu, ZHAO Yuming, WANG Xiuying, et al. Discussion of evaluation methods on soil fertility [J]. J Yangtze River Sci Res Inst, 2008, 25(3): 62 − 66. [12] 王子龙, 付强, 姜秋香. 土壤肥力综合评价研究进展[J]. 农业系统科学与综合研究, 2007, 23(1): 15 − 18. WANG Zilong, FU Qiang, JIANG Qiuxiang. Research progress in the integrated evaluation of soil fertility [J]. Syst Sci Compr Studies Agric, 2007, 23(1): 15 − 18. [13] 姜春前, 徐庆, 姜培坤. 不同森林植被下土壤化学和生物化学肥力的综合评价[J]. 林业科学研究, 2002, 15(6): 700 − 705. JIANG Chunqian, XU Qing, JIANG Peikun. Integrated evaluation of soil chemical and biochemical fertility under different vegetations [J]. For Res, 2002, 15(6): 700 − 705. [14] 吕晓男, 陆允甫, 王人潮. 土壤肥力综合评价初步研究[J]. 浙江大学学报(农业与生命科学版), 1999, 25(4): 378 − 382. LÜ Xiaonan, LU Yunfu, WANG Renchao. Preliminary studies on the integrated evaluation of soil nutrient fertility [J]. J Zhejiang Univ Agric Life Sci, 1999, 25(4): 378 − 382. [15] 孙波, 张桃林, 赵其国. 我国东南丘陵山区土壤肥力的综合评价[J]. 土壤学报, 1995, 32(4): 362 − 369. SUN Bo, ZHANG Taolin, ZHAO Qiguo. Comprehensive evaluation of soil fertility in the hilly and mountainous region of southeastern China [J]. Acta Pedol Sin, 1995, 32(4): 362 − 369. [16] 王玲, 吕殿青, 覃志斌, 等. 岳麓山风景区坡面林地土壤理化特征评析[J]. 湖南师范大学(自然科学学报), 2011, 34(2): 84 − 88. WANG Ling, LÜ Dianqing, QIN Zhibin, et al. Review and analysis of soil physical and chemical characteristics on woodland slope in Yuelu Mountain [J]. J Nat Sci Hunan Norm Univ, 2011, 34(2): 84 − 88. [17] 柳若安, 刘厚田. 酸度和铝对马尾松生长的影响[J]. 植物学报, 1995, 37(2): 154 − 158. LIU Ruoan, LIU Houtian. Effect of acidity and aluminium on the growth of Pinus massoniana seedlings [J]. Acta Bot Sin, 1995, 37(2): 154 − 158. [18] 包甜甜, 张鼎华. “近自然抚育”对南方山地幼林地土壤磷有效性的影响[J]. 福建师范大学学报(自然科学版), 2019, 35(2): 97 − 103. BAO Tiantian, ZHANG Dinghua. Phosphorus availability of red soil and its relation with tree growth in mountainous region of southern China [J]. J Fujian Norm Univ Nat Sci Ed, 2019, 35(2): 97 − 103. [19] 刘俊钦. 杉木混交林近自然经营生长效果研究[J]. 乡村科技, 2018(14): 47 − 49. LIU Junqin. Study on the growth effect of natural management of Chinese fir mixed forest [J]. Village Sci Technol, 2018(14): 47 − 49. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190549
 点击查看大图
点击查看大图
计量
- 文章访问数: 2288
- HTML全文浏览量: 434
- PDF下载量: 69
- 被引次数: 0

