




 下载:
下载:



-
番茄Lycopersicon esculentum发育过程中果实积累的糖分组成和含量高低是评价果实品质的重要指标。光合作用产生的部分蔗糖经韧皮部卸载到果实,在蔗糖代谢相关酶的作用下形成糖组分结构。杨玉梅[1]报道:蔗糖合成酶(SS)能够调控蔗糖在韧皮部的卸载,当库器官与韧皮部达到一定的蔗糖浓度梯度后,蔗糖会被转运入细胞,且SS活性与库器官中蔗糖输入呈正相关。齐红岩等[2]以番茄‘辽园多丽’Lycopersicon esculentum ‘Liaoyuan Duoli’为研究对象,发现有一小部分蔗糖在组织内运输转移时被分解,到达果实内部后,进一步分解为葡萄糖和果糖。宋曼曼等[3]发现:不同时期番茄果实中各种糖的质量分数不同且不断变化,一般绿熟期果糖与葡萄糖质量分数相对较低,到成熟期达到峰值;绿熟期至转色期,蔗糖质量分数先增加后下降。刘以前等[4]的研究表明,在产量形成期,酸性转化酶(AI)和中性转化酶(NI)的活性较低,SS和蔗糖磷酸合成酶(SPS)的活性较高;在品质形成期,番茄果实以积累果糖为主,AI和NI的活性急剧升高,SS和SPS的活性很低。提高番茄果实的甜度是改善番茄品质的重要环节,而番茄果实的甜度取决于糖分积累中的蔗糖含量[5]。目前,对番茄生长发育过程中糖分的积累与蔗糖代谢相关酶活性变化的研究已较多,但关于营养液中添加不同盐类提高电导率(EC),对水培番茄果实成熟过程中糖组分变化与蔗糖代谢相关酶活性的影响研究较少。本研究探究了不同处理下番茄果实4种糖(蔗糖、葡萄糖、果糖和淀粉)的积累规律,目的在于为今后高品质高风味番茄栽培提供理论依据,也对营养液中添加不同盐类对水培番茄果实糖组分与蔗糖代谢相关酶活性的影响做了初步探索。
-
试验于2017年3月在宁夏贺兰园艺产业园科研开发区玻璃温室进行。供试番茄品种为‘粉太郎2号’ Lycopersicon esculentum ‘Fentailang 2’;使用不锈钢栽培槽(20.0 m × 35.5 cm × 10.0 cm)作为水培试验装置,底部铺设20.0 m × 35.0 cm的无纺布;使用岩棉块(8.5 cm × 8.5 cm × 6.5 cm)作为固定基质。定植后以泡沫板覆盖栽培槽,以减少营养液蒸发损失。
-
试验基础营养液配方:四水硝酸钙945.00 mg·L-1,硝酸钾809.00 mg·L-1,磷酸二氢铵153.00 mg·L-1,七水硫酸镁493.00 mg·L-1,其中微量元素铁3.00 mg·L-1,硼0.50 mg·L-1,锰0.50 mg·L-1,锌0.05 mg mg·L-1,铜0.02 mg·L-1,钼0.01 mg·L-1。电导率为0.24 S·m-1。试验共设计5个处理,对照组为基础营养液(ck),4个试验组分别在基础营养液中添加氯化钠(TA),氯化钾(TB),氯化钙(TC)和石膏(TD)。调节营养液电导率值为0.40 S·m-1。
试验为完全随机区组设计,设置小区3个·处理-1,定植番茄38株·小区-1。3月9日定植,7月13日拉秧,行距为158.0 cm,平均株距为17.4 cm。定植后4 d清水缓苗,再用试验基础营养液使幼苗适应2 d,之后用盐处理液浇灌。
栽培槽营养液pH值控制在pH 5.5~7.3,温度维持在18~25 ℃,24 h连续供液。循环使用1个月后彻底更新一次,深度根据外界环境与植株生长状况进行调整。
-
各处理标记生长良好的番茄植株12株,生长至绿熟期、转色期、成熟期时各摘取第2穗果中大小、发育程度基本一致的番茄果实1个,放入已装有大量冰袋的苯板箱中,运回实验室。解剖取样,每有4个果实解剖完毕,就将其各个部位分装至搅拌机中,搅拌成匀浆。以上操作均在4 ℃环境下进行。取样部位:果皮、果肉、心室隔壁及胶质胎座。
-
参照文献[3],用比色法测定蔗糖、葡萄糖、果糖及淀粉的质量分数。参照文献[2]和[6],测定蔗糖合成酶(sucrose synthase,SS),蔗糖磷酸合成酶(sucrose phosphate synthase,SPS),酸性转化酶(acid invertase,AI)和中性转化酶(neutral invertase,NI)的活性。
-
利用SAS 8.2统计软件和Excel 2010进行数据分析,采用最小显著差法(LSD)进行显著性差异分析。
-
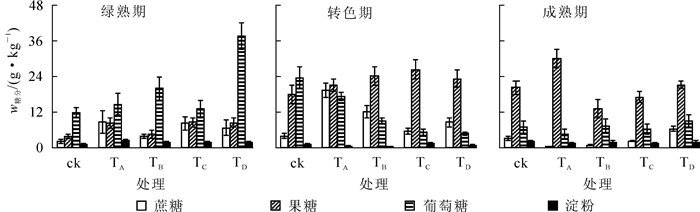
由图 1可知:绿熟期果皮几乎不含淀粉,成熟期果皮淀粉质量分数明显上升,说明果皮在成熟过程中不断积累淀粉。相比于转色期与成熟期,绿熟期各个处理均有极高的葡萄糖质量分数(质量分数为14.5~39.4 g·kg-1)和较低的果糖质量分数;到转色期,果皮中的葡萄糖质量分数明显下降,果糖质量分数明显上升(P<0.05);而对照中果糖质量分数增加相对较少,仅增加6.1 g·kg-1。可以推测,果皮从绿熟期到转色期积累了大量的果糖。营养液中添加盐类有利于转色期果皮果糖的积累。成熟果实的果皮蔗糖质量分数极低,添加盐类的处理均低于5.0 g·kg-1,其中TA,TB和TD处理成熟期果皮蔗糖质量分数明显低于对照(P<0.05),说明营养液中添加氯化钠、氯化钾或石膏会明显降低成熟期果皮的蔗糖质量分数;相比于成熟期,绿熟期与转色期果皮均有较高的蔗糖质量分数;对照果皮在绿熟期时蔗糖质量分数最低,各处理蔗糖质量分数明显高于对照(P<0.05),其中TA处理蔗糖质量分数为对照的8.7倍,表明营养液中添加不同盐类会明显提高绿熟期果皮的蔗糖质量分数,且添加氯化钠效果最为明显。所有处理的果皮在成熟期的4种糖的总质量分数均低于转色期,果实成熟时,TD处理下果皮4种糖(蔗糖、果糖、葡萄糖、淀粉)的总质量分数最高(41.5 g·kg-1)。TA处理成熟期果皮有较高的果糖质量分数,达20.7 g·kg-1,占4种糖总质量分数的64.09%。

图 1 营养液中添加不同盐类对水培番茄果皮糖组分的影响
Figure 1. Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit peel
-
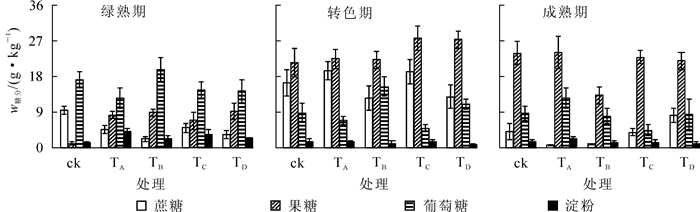
由图 2可知:果肉在3个时期都几乎不含淀粉,淀粉质量分数均小于2.5 g·kg-1。所有处理绿熟期果肉的糖以葡萄糖为主,TD处理在此时期葡萄糖质量分数极高,达37.6 g·kg-1,明显高于另外4个处理(P<0.05),说明营养液中添加石膏会提高此时期果肉的葡萄糖质量分数。所有处理果肉在转色期与成熟期主要含果糖。营养液中添加不同盐类会提高转色期果肉的果糖与蔗糖质量分数。对照果肉蔗糖质量分数在3个时期基本相当,而营养液中添加盐类的处理有一定程度的变化,TA与TB处理果肉蔗糖质量分数的变化较其他处理明显,且这2个处理成熟期果肉蔗糖质量分数极低,表明营养液中添加不同盐类或许会促进果肉在转色期积累蔗糖,而添加氯化钠或氯化钾会明显降低成熟期果肉的蔗糖质量分数。成熟期果实TD处理果肉4种糖的总质量分数最高(38.5 g·kg-1)。TA处理成熟期果肉的果糖质量分数极高,达30.0 g·kg-1,占4种糖总质量分数的83.10%。

图 2 营养液中添加不同盐类对水培番茄果肉糖组分的影响
Figure 2. Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit pulp
-
果实心室隔壁中4种糖的总质量分数在成熟过程中有明显的先上升后下降的趋势。由图 3可知:绿熟期果实心室隔壁主要含葡萄糖(14.2~19.7 g·kg-1),到转色期与成熟期则以果糖为主。果实心室隔壁在转色期有较高的蔗糖质量分数,相比于对照,所有处理在绿熟期果糖和淀粉质量分数均较高,蔗糖则较低,说明盐类会明显提高绿熟期果实心室隔壁的果糖与淀粉质量分数,显著降低蔗糖质量分数(P<0.05)。TA与TB处理下,成熟期果实心室隔壁蔗糖质量分数极低,表明氯化钠或氯化钾显著降低成熟果实心室隔壁的蔗糖质量分数。TA处理成熟期果实心室隔壁亦有极高的果糖质量分数,达24.1 g·kg-1,占4种糖总质量分数的60.86%。TB处理成熟期果实心室隔壁的4种糖的总质量分数(23.4 g·kg-1)明显低于其他处理(P<0.05),可以推测,氯化钾会降低成熟果实心室隔壁4种糖的总质量分数。

图 3 营养液中添加不同盐类对水培番茄果实心室隔壁糖组分的影响
Figure 3. Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit dissepiment
-
与对照相比,营养液中添加盐类的处理绿熟期果实胶质胎座葡萄糖质量分数占极高的比例(图 4),TA与TB处理下这一现象更为明显,说明营养液中添加不同盐类会改变绿熟期果实胶质胎座糖组分的占比结构,明显提高葡萄糖质量分数,降低果糖质量分数。TC与TD处理绿熟期果实胶质胎座4种糖的总质量分数明显低于其他3个处理,表明氯化钙或石膏会降低绿熟期果实胶质胎座的含糖量。转色期与成熟期的果实胶质胎座糖组分变化情况与果实果皮、果肉和心室隔壁相似。成熟期果实胶质胎座的4种糖总质量分数为TC处理最高(44.2 g·kg-1),说明营养液中添加氯化钙会在一定程度上提高成熟果实心室隔壁的糖总质量分数。

图 4 营养液中添加不同盐类对水培番茄果实胶质胎座糖组分的影响
Figure 4. Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit pectinic
-
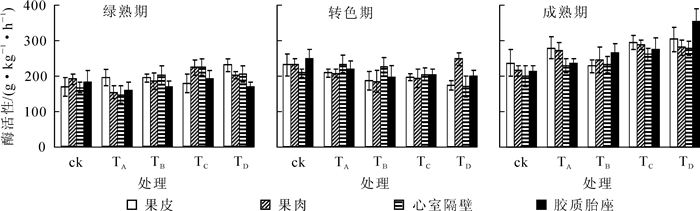
蔗糖合成酶(SS)是一种既能催化合成蔗糖,又能分解蔗糖的可逆酶。对SS合成活性的测定(图 5)可知:对照、TC和TD处理下番茄果实4个部位的SS活性从绿熟期至成熟期先上升后下降,在转色期达到最高,其次为成熟期。TC和TD处理果皮内的SS在各时期的活性均小于对照组,说明营养液中添加氯化钙或石膏会降低果皮SS的活性。TA处理与TB处理的果实果皮和心室隔壁内的SS活性变化类似,总体呈下降趋势,与另外3个处理不同,且绿熟期果实这2个部位的SS活性高于另外3个处理,说明营养液中添加氯化钠或氯化钾可能会改变这2个部位SS活性原有的变化趋势,并且会明显提高绿熟期果皮与心室隔壁内SS的活性。除TA外,其他处理果实果肉与胶质胎座内的SS活性的变化趋势一致,均先上升后下降,而TA处理果实4个部位的SS活性变化情况相似,均为绿熟期最高,转色期最低。由SS在转色期的表现可以推测,营养液中添加氯化钠会严重抑制转色期果实4个部位的SS活性。转色期果实4个部位的SS活性均为对照最高,说明营养液中添加不同盐类会在一定程度上抑制此时期SS活性。

图 5 不同处理下番茄果实不同部位蔗糖合成酶活性的变化
Figure 5. Variation of SS activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
-
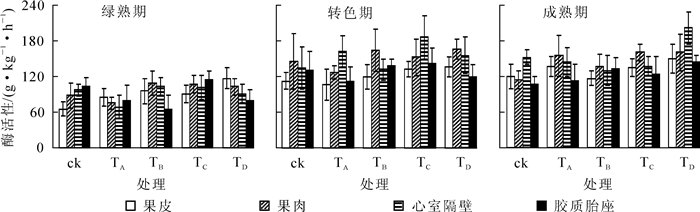
从图 6可知:除TA外,另4个处理果实4个部位的SPS活性变化趋势一致,自绿熟期至成熟期均先上升后下降。TB,TC和TD处理下,果实各部位SPS活性在转色期高于对照组,说明营养中添加氯化钾、氯化钙或石膏会提高转色期SPS活性;TA处理果实各部位的SPS活性变化趋势不一致,但活性变化不大,较稳定,且转色期心室隔壁SPS的活性低于其他处理,表明营养液中添加氯化钠会抑制心室隔壁在转色期的SPS活性。绿熟期果实果皮与果肉的SPS活性均表现为对照最低,可以推测营养液中添加盐类会提高绿熟期果皮与果肉的SPS活性。从成熟期果实SPS活性来看,营养液中添加石膏会提高成熟果实各部位的SPS活性。

图 6 不同处理下番茄果实不同部位蔗糖磷酸合成酶活性的变化
Figure 6. Variation of SPS activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
-
转化酶能够催化蔗糖分解成葡萄糖和果糖。转化酶有许多种类,一类为可溶性酸性转化酶(AI),等电点为酸性;另一类为中性转化酶(NI),等电点为中性[7]。由图 7可知:各个处理成熟期果实各部位的AI活性均高于绿熟期,且TA,TB,TC与TD处理成熟期果实各个部位的AI活性总体上高于对照,TD处理成熟期果实各部位的AI活性均高于其他处理,说明果实成熟时AI活性变大,营养液中添加盐类会提高成熟期果实AI的活性,且以石膏效果最为明显。而转色期时各个处理各部位的AI活性或变大或降低,仅对照处理果实4个部位AI活性呈现了一致的变化趋势。TD处理转色期果皮与心室隔壁AI活性低于其他处理,说明营养液中添加石膏会降低此时期这2个部位的AI活性。此外,营养液中添加氯化钙或石膏对AI活性变化影响较为一致。

图 7 不同处理下番茄果实不同部位酸性转化酶活性的变化
Figure 7. Variation of AI activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
-
由图 8可知:所有处理果实各部位中性转化酶活性转色期高于绿熟期,到成熟期时活性或继续变大或略有降低,而成熟期果实各部位的NI活性高于绿熟期。除TD处理外,不同处理果实各部位NI活性变化趋势较为一致,为倒Ⅴ型。成熟期,各处理组果实各部位的NI活性总体上高于对照,而TD处理高于其他处理,表明营养液中添加不同盐类会提高成熟果实NI的活性,且以石膏效果最为明显。

图 8 不同处理下番茄果实不同部位中性转化酶活性的变化
Figure 8. Variation of NI activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
-
果实各种糖分的积累是糖代谢相关酶共同作用的结果[1],果实中积累的糖分不仅决定果实的甜度,也是合成其他营养物质的原料[8],是果实品质形成的关键,因此可认为糖组分情况对果实风味和品质具有重要影响[2, 9-10]。营养液中添加不同盐类后,溶液电导率值变化,不同程度上影响番茄果实糖分的积累量与蔗糖代谢相关酶活性。本研究中,5个处理果实各部位4种糖的总质量分数为15.3~58.2 g·kg-1,转色期4种糖的总质量分数较高,成熟期总质量分数略有下降,糖组分的这一变化在心室隔壁尤为明显。果实由绿熟期至转色期积累大量的果糖;绿熟期果实各部位的糖以葡萄糖为主,而转色期与成熟期则以果糖为主;自绿熟期至成熟期,SS与SPS活性总体上表现为在转色期上升,成熟期下降的倒Ⅴ型趋势,且转色期活性远高于或略高于绿熟期,成熟期活性与绿熟期活性基本相当,甚至低于绿熟期;所有处理的成熟期果实各部位AI和NI活性均大于绿熟期,说明番茄果实各部位在转色期蔗糖合成较为活跃,在成熟期分解大量的蔗糖。基本解释了果实各部位在转色期含有较高的蔗糖质量分数,而到成熟期时又几乎不含蔗糖。在番茄果实分解蔗糖积累其他糖类的过程中,转化酶发挥着重要作用[9]。ISLAM等[11]报道表明,转化酶的活性在番茄果实成熟过程中不断增强,果实成熟时活性达到最高,所以成熟果实蔗糖分解最为强烈。另有研究表明,番茄发育至成熟阶段,果实AI与NI活性大幅上升,参与催化分解之前所积累的淀粉与蔗糖,产生葡萄糖与果糖[4, 12]。除个例外,本研究所进行的处理基本上不会改变果实原来的糖分变化趋势及蔗糖代谢相关酶活性的变化趋势,且研究所得结果与前人的研究结果基本相符。
有学者[1, 4, 9, 13-14]研究了糖与蔗糖代谢相关酶的相关性,发现番茄果实中可溶性总糖、果糖的质量分数与蔗糖降解酶的活性间存在很好相关性,而蔗糖质量分数却与SS和SPS的相关性很低,与AI和NI有相对较高的负相关性;果糖质量分数与SS和AI,NI的活性均为显著正相关;番茄果实中的可溶性总糖与SS的活性呈显著正相关[4]。罗海玲等[13]研究发现:厚皮甜瓜品种‘丰蜜二号’Cucumis melo ‘Fengmi 2’和‘好运52’ Cucumis melo ‘Haoyun 52’的还原糖质量分数与果肉NI活性呈显著负相关。此类结论定性地解释某类糖质量分数的上升或下降与某个酶的关联性,但不能明确说明某类糖在某段时间可能会积累或分解多少。本研究没有使用与样品实际糖质量分数相接近的糖液,仅能定性研究糖与酶的关系,不能对相关酶活性作出定量判断,因此在今后的研究中,需要对测定方法作进一步的改进,首先测定样品自身的糖组分质量分数,再根据组织的糖质量分数配制出相近浓度的糖液进行酶活性测定,以测得相对更可靠的蔗糖代谢相关酶活性,为能测算出糖净积累量或糖分解量奠定基础。这样则更有利于探究果实糖组分的变化规律,以期开发出新的高风味番茄栽培管理方法。
Carbohydrate composition and sucrose-metabolizing enzyme activities of hydroponic tomato fruit with salts added to nutrient solutions
-
摘要: 以营养液膜技术水培番茄‘粉太郎2号’ Lycopersicon esculentum ‘Fentailang 2’。测定果皮、果肉、心室隔壁和胶质胎座4个部位的糖质量分数与蔗糖代谢相关酶活性,研究营养液中添加不同盐类(氯化钠、氯化钾、氯化钙和石膏,电导率为0.40 S·m-1)对水培番茄不同生长期果实各部位糖组分和蔗糖代谢相关酶活性的影响。结果显示:转色期各部位4种糖(蔗糖、果糖、葡萄糖、淀粉)的总质量分数较高,成熟期质量分数略有下降;转色期与成熟期各部位以果糖为主,绿熟期以葡萄糖为主。各部位在转色期蔗糖合成较为活跃,在成熟期分解大量蔗糖。营养液中添加氯化钠或氯化钾明显降低成熟期果皮、果肉和心室隔壁的蔗糖质量分数。果实成熟过程中,蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性表现为在转色期上升,成熟期下降的倒Ⅴ型趋势。各处理成熟期果实各部位的酸性转化酶(AI)和中性转化酶(NI)活性均大于绿熟期。营养液中添加不同盐类在不同程度上抑制转色期果实SS活性,提高成熟期果实转化酶的活性;添加氯化钾、氯化钙或石膏提高转色期果实SPS活性,添加氯化钠或氯化钾改变果实某些部位SS活性原有的变化趋势。Abstract: To explore the effect of adding different salts to a nutrient solution (NaCl, KCl, CaCl2 or CaSO4·2H2O, the electric conductivity (EC) was controlled at 0.40 S·m-1) on carbohydrate composition and sucrose-metabolizing related enzyme activities of hydroponic tomato fruit during fruit development, the experimental design was totally randomized block, and this experiment cultivated Lycopersicon esculentum 'Fentailang 2' using the nutrient film technique (NFT), and twelve tomato plants were labeled with good growth in every treatment, one tomato fruit with the same size and development degree was selected from the second layers at different growth stages, and content of carbohydrates and sucrose-metabolizing related enzyme activities in four parts of the tomato fruit (peel, pulp, dissepiment, and pectinic) were determined. Significant difference analysis was conducted by least significant difference (LSD). The main results included a total mass fraction of four sugars (sucrose, fructose, glucose, and starch) in each part of the fruit that were relatively high at the color conversion stage and had a slight decline after maturity. Fructose was the main sugar in four parts of the fruit at the stage of color conversion and ripening, and glucose was the main sugar at the green ripening stage. More sucrose (P < 0.05) was accumulated in each part at the color conversion stage, and a large amount (P < 0.05)of sucrose was decomposed at the ripening stage. During fruit ripening, except some treatments sucrose synthase (SS) and sucrose phosphate synthase (SPS) activities rose at the color conversion stage and fell at the ripening stage in the shape of an inverted "V". For all treatments, acid invertase (AI) and neutral invertase (NI) activities in each part of ripened fruits were higher than those at the green ripening stage. Overall, adding NaCl or KCl to the nutrient solution could reduce (P < 0.05)the mass fraction of sucrose in the peel, pulp, and dissepiment of ripened fruits; adding different salts to the nutrient solution could inhibit the SS activity of the color conversion stage fruit in varying degrees and increase invertase activities of ripened fruit; adding KCl, CaCl2, or CaSO4·2H2O could increase (P < 0.05)the SPS activity of the color conversion stage fruit; and adding NaCl or KCl could change the original trend of SS activity in some parts of the fruit.
-
Key words:
- horticulture /
- salts /
- hydroponics /
- tomato fruit /
- carbohydrate composition /
- sucrose metabolism related enzymes
-
图 1 营养液中添加不同盐类对水培番茄果皮糖组分的影响
Figure 1 Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit peel
图 2 营养液中添加不同盐类对水培番茄果肉糖组分的影响
Figure 2 Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit pulp
图 3 营养液中添加不同盐类对水培番茄果实心室隔壁糖组分的影响
Figure 3 Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit dissepiment
图 4 营养液中添加不同盐类对水培番茄果实胶质胎座糖组分的影响
Figure 4 Effect of adding different salts to nutrient solution on carbohydrate component of hydroponic tomato fruit pectinic
图 5 不同处理下番茄果实不同部位蔗糖合成酶活性的变化
Figure 5 Variation of SS activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
图 6 不同处理下番茄果实不同部位蔗糖磷酸合成酶活性的变化
Figure 6 Variation of SPS activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
图 7 不同处理下番茄果实不同部位酸性转化酶活性的变化
Figure 7 Variation of AI activity in tomato fruit peel, pulp, dissepiment and pectinic under different treatments
-
[1] 杨玉梅.不同西瓜种质资源糖分积累规律与糖代谢相关酶关系的研究[D].杨凌: 西北农林科技大学, 2006. YANG Yumei. Study on Sugar Accumulation of Different Watermelon Germplasms and Correlation Between Sugar Metabolism and Related Enzymes[D]. Yangling: Northwest A & F University, 2006. [2] 齐红岩, 李天来, 张洁, 等.番茄果实发育过程中糖的变化与相关酶活性的关系[J].园艺学报, 2006, 33(2):294-299. QI Hongyan, LI Tianlai, ZHANG Jie, et al. Relationship between carbohydrate change and related enzymes activities during tomato fruit development[J]. Acta Hortic Sin, 2006, 33(2):294-299. [3] 宋曼曼, 韩广泉, 樊新民, 等.加工番茄果实糖分含量的变化[J].北方园艺, 2011(4):24-28. SONG Manman, HAN Guangquan, FAN Xinmin, et al. Study on sugar content changes of processing tomato fruit[J]. North Hortic, 2011(4):24-28. [4] 刘以前, 沈火林, 石正强.番茄果实生长发育过程中糖的代谢[J].华北农学报, 2006, 21(3):51-56. LIU Yiqian, SHEN Huolin, SHI Zhengqiang. The sugar metabolism in tomato developing fruits[J]. Acta Agric Boreal-Sin, 2006, 21(3):51-56. [5] 齐红岩, 李天来, 邹琳娜, 等.番茄果实不同发育阶段糖分组成和含量变化的研究初报[J].沈阳农业大学学报, 2001, 32(5):346-348. QI Hongyan, LI Tianlai, ZOU Linna, et al. Changes of composition and content of carbohydrate during tomato fruit development[J]. J Shenyang Agric Univ, 2001, 32(5):346-348. [6] 王永章, 张大鹏.乙烯对成熟期新红星苹果果实碳水化合物代谢的调控[J].园艺学报, 2000, 27(6):391-395. WANG Yongzhang, ZHANG Dapeng. Regulating effects of ethylene on carbohydrate metabolism in 'Starkrimson' apple fruit during the ripening period[J]. Acta Hortic Sin, 2000, 27(6):391-395. [7] 张明方, 李志凌.高等植物中与蔗糖代谢相关的酶[J].植物生理学通讯, 2002, 38(3):289-295. ZHANG Mingfang, LI Zhiling. Sucrose-metabolizing enzymes in higher plants[J]. Plant Physiol Commun, 2002, 38(3):289-295. [8] 秦巧平, 林飞凡, 张岚岚.枇杷果实糖酸积累的分子生理机制[J].浙江农林大学学报, 2012, 29(3):453-457. QIN Qiaoping, LIN Feifan, ZHANG Lanlan. Review of the studies on the accumulation mechanisms of sugar and organic acids in Eriobotrya japonica fruit[J]. J Zhejiang A & F Univ, 2012, 29(3):453-457. [9] 赵建华, 李浩霞, 尹跃, 等. 4种枸杞果实发育过程中糖积累与蔗糖代谢酶的关系[J].浙江农林大学学报, 2016, 33(6):1025-1032. ZHAO Jianhua, LI Haoxia, YIN Yue, et al. Sugar accumulation and sucrose-metabolizing enzyme activities in four Lycium species during fruit development[J]. J Zhejiang A & F Univ, 2016, 33(6):1025-1032. [10] 张永平, 乔永旭, 喻景权, 等.园艺植物果实糖积累的研究进展[J].中国农业科学, 2008, 41(4):1151-1157. ZHANG Yongping, QIAO Yongxu, YU Jingquan, et al. Progress of researches of sugar accumulation mechanism of horticultural plant fruits[J]. Sci Agric Sin, 2008, 41(4):1151-1157. [11] ISLAM S, MATSUI T, YOSHIDA Y. Carbohydrate content and the activities of sucrose synthase, sucrose phosphate synthase and acid invertase in different tomato cultivars during fruit development[J]. Sci Hortic, 1996, 65(2/3):125-136. [12] HO L C. The mechanism of assimilate partitioning and carbo-hydrate compartmentation in fruit in relation to the quality and yield of tomato[J]. J Exp Bot, 1996, 47(spec):1239-1243. [13] 罗海玲, 黄玉辉, 张曼, 等.转化酶与甜瓜果实膨大及糖分积累相关研究[J].南方农业学报, 2011, 42(4):384-387. LUO Hailing, HUANG Yuhui, ZHANG Man, et al. The relationship between invertase activity and sucrose accumulation during the development of fruits in melon[J]. J South Agric, 2011, 42(4):384-387. [14] 武红霞, 邢姗姗, 王松标, 等.'台农1号'芒果果实发育过程中的糖分积累与相关酶活性研究[J].西北植物学报, 2011, 31(9):1811-1815. WU Hongxia, XING Shanshan, WANG Songbiao, et al. Sugar accumulation and related enzyme activities in the developing mango fruits cv.'Tainong 1'[J]. Acta Bot Boreal-Occident Sin, 2011, 31(9):1811-1815. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2018.06.016
 点击查看大图
点击查看大图

计量
- 文章访问数: 3640
- HTML全文浏览量: 702
- PDF下载量: 496
- 被引次数: 0
