





-
森林群落的物种多样性和垂直、径阶结构反映了森林群落的形成、稳定性及演替规律[1-2]。开展濒危植物群落结构及物种多样性的研究,可以了解种群的数量动态及演变趋势,揭示珍稀植物的濒危过程及濒危机理[3],为濒危植物的就地和迁地保护提供理论基础。多脉铁木Ostrya multinervis属桦木科Betulaceae铁木属Ostrya植物,分布于湖北利川、湖南紫云山、四川东南部和贵州等地,分布区孤立局限,为中国特有种,具有树体高大,干形通直圆满,材质优良等特点[4]。浙江仅在文成县石垟林场和龙泉市凤阳山有记载,是《浙江珍稀濒危植物》收录的162种浙江省珍稀濒危的野生植物之一[5]。前人对多脉铁木的播种、扦插及嫁接繁育实验[4]和林分基本结构进行了初步研究[6],而对群落中不同植物的重要值、植物多样性指数、乔木层树种的垂直和径阶结构等方面尚未有报道。本研究采用标准样地法对浙江省文成石垟林场多脉铁木群落的区系组成、结构和演替规律等进行了研究,为该树种的就地和迁地保护提供理论基础。
-
研究区位于浙江省文成县石垟林场猴王谷景区倒臼源,多脉铁木分布中心的地理位置为27°50′N,119°50′E。该区属中亚热带季风性气候,年平均气温为12.8 ℃,最冷月(1月)平均气温为4.0 ℃,极端最低气温为-14.0 ℃,最热月(7月)平均气温为23.6 ℃,极端最高气温为37.3 ℃,年降水量为1 604 mm,年均相对湿度为84.0%,年日照时数为1 755 h,全年无霜期为288 d[7]。该区属天然常绿阔叶林,森林覆盖率达90%以上。多脉铁木群落分布区域的海拔为500~700 m,坡度为30°~35°,南坡,坡中部,地表有石块层覆盖,厚度为20~30 cm,石块大小为20~40 cm。土壤为发育于花岗岩的红壤土类,表层土壤(0~20 cm)理化性质为pH 4.6,有机碳为63.2 g·kg-1,碱解氮、有效磷、速效钾分别为145.2,21.8,187.0 mg·kg-1。
-
2017年8月上旬对多脉铁木群落进行全面调查。采用典型抽样法,以多脉铁木分布中心为起点,沿坡向在山体上坡至下坡地段,布设4个20 m × 20 m的样地,将每个样地划分为16个5 m× 5 m的小样方,开展分层调查。乔木层、灌木层和草本层的调查指标参见王丽敏等[2]方法。
-
根据多脉铁木群落的株高(H)特点,将乔木层的垂直结构划分为6个等级,间隔5 m,分别为3 m≤H<8 m,8 m≤H<13 m,13 m≤H<18 m,18 m≤H<23 m,23 m≤H<28 m,H≥28 m。乔木层径阶(D)结构划分为7个等级,相隔5 cm,分别为5 cm≤D<10 cm,10 cm≤D<15 cm,15 cm≤D<20 cm,20 cm≤D<25 cm,25 cm≤D<30 cm,30 cm≤D<35 cm,D≥35 cm。
-
不同植物的重要值、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数计算按郑昌龙等[8]方法进行;植物生活型谱统计参考高贤明等[9]方法进行;种子植物地理成分根据吴征镒等[10]的方法进行统计分析。
-
多脉铁木群落中共有维管束植物60种,隶属于40科52属。其中蕨类植物6科7属8种,裸子植物1科1属1种,被子植物33科44属51种。种类数量占优势的科由大到小依次为百合科Liliaceae(5属6种),壳斗科Fagaceae(4属4种),山茶科Theaceae(3属3种),槭树科(1属3种);大戟科Euphorbiaceae(2属2种),木通科Lardizabalaceae(2属2种),樟科Lauraceae(2属2种),鼠李科Rhamnaceae(2属2种),冬青科Aquifoliaceae(1属2种),莎草科Cyperaceae(1属2种),水龙骨科Polypodiaceae(1属2种),鳞毛蕨科Dryopteridaceae(1属2种)。含有单种的属占绝对优势,有28属,占总属数的53.8%。可见,多脉铁木群落科属组成复杂,植物种类丰富。
-
图 1表明:多脉铁木群落中,不同生活型植物种类所占比例依次为高位芽植物(41.7%)>地上芽植物(26.7%)>隐芽植物(20.0%)>地面芽植物(10.0%)>1年生植物(1.6%)。

图 1 多脉铁木群落不同生活型谱
Figure 1. Life-form spectrum of survival community of Ostrya multinervis
-
从表 1可知:多脉铁木群落中种子植物属泛热带分布的类型有9属,占总属数的20.0%,如菝葜属Smilax,紫金牛属Ardisia和冬青属Ilex等。其他依次为东亚分布类型有8属,占总属数的17.8%,如山麦冬属Liriope,木通属Akebia,和沿阶草属Ophiopogon等。北温带、东亚和北美洲间断分布类型均为7属,均占总属数的15.6%。该群落中没有植物分布的类型区有旧世界热带、热带亚洲至热带非洲、温带亚洲、地中海地区、西亚至中亚、中亚及中国特有等6个。
表 1 多脉铁木群落种子植物属的分布区
Table 1. Genera distribution types of seed plants in O. multinervis community
分布区类型 属数/个 占比/% 1 广布 2 4.4 2 泛热带 9 20.0 3 东亚及热带南美间断 4 8.9 4 旧世界热带 0 0.0 5 热带亚洲至热带大洋洲 1 2.2 6 热带亚洲至热带非洲 0 0.0 7 热带亚洲 6 13.3 8 北温带 7 15.6 9 东亚和北美洲间断 7 15.6 10 旧世界温带 1 2.2 11 温带亚洲 0 0.0 12 地中海地区、西亚至中亚 0 0.0 13 中亚 0 0.0 14 东亚 8 17.8 15 中国特有 0 0 合计 45 100 -
多脉铁木群落具有完整的垂直结构,具有乔木层、灌木层、草本层和层间植物。乔木层共有9种植物(表 2),多脉铁木的重要值最大,达62.158 6,是该群落的建群种,控制着整个群落的组成、结构和生境,依次分别是褐叶青冈Cyclobalanopsis stewardiana,小叶栎Quercus chenii,四照花Dendrobenthamia japonica var. chinensis,缺萼枫香Liquidambar acalycina,短尾柯Lithocarpus brevicaudatus,钩栲Castanopsis tibetana,尾叶冬青Ilex wilsonii,麂角杜鹃Rhododendron latoucheae。
表 2 多脉铁木群落主要植物的重要值
Table 2. Importance values of tree populations in Ostrya multinervis community
乔木层 重要值 灌木层 重要值 草本层 重要值 多脉铁木 62.158 6 披针叶茴香 15.673 5 里白 21.161 2 褐叶青冈 14.517 1 毛花连蕊茶 7.607 3 黑足鳞毛蕨 16.001 8 小叶栎 5.929 1 光叶石楠 6.849 2 麦冬 13.2102 四照花 4.000 7 长裂葛萝槭 5.129 1 求米草 6.789 8 缺萼枫香 3.169 7 红楠 4.786 1 三穗薹草 6.621 1 短尾柯 2.913 7 三尖杉 4.759 9 瘤足蕨 5.926 4 钩栲 2.567 6 猴欢喜 2.301 2 江南卷柏 5.479 5 尾叶冬青 0.980 9 大叶冬青 2.854 5 毛叶腹水草 4.588 5 麂角杜鹃 0.797 1 秀丽槭 1.865 6 大叶唐松草 3.942 2 青榨槭 1.857 1 延羽卵果蕨 3.168 9 灌木层发达,生长良好,平均高为1.5 m,盖度为40%。重要值排名前10位的优势植物如表 2所示。重要值最大的是披针叶茴香Illicium lanceolatum,其值为15.673 5,其次分别为毛花连蕊茶Chandleri elagans,光叶石楠Photinia glabra,长裂葛萝槭Acer grosseri,红楠Machilus thunbergii,三尖杉Cephalotaxus fortunei,猴欢喜Sloanea sinensis,大叶冬青Ilex latifolia,秀丽槭Acer elegantulum,青榨槭Acer davidii。
草本层平均高为0.4 m,盖度为20%。重要值排名前10位的优势草本植物见表 2。从表 2可知:重要值大小分别为里白Diplopterygium glaucum,黑足鳞毛蕨Dryopteris fuscipes,麦冬Ophiopogon japonicus,求米草Oplismenus undulatifolius,三穗薹草Carex tristachya,瘤足蕨Plagiogyria adnata,江南卷柏Selaginella moellendorfii,毛叶腹水草Veronicastrum villosulum,大叶唐松草Thalictrum faberi,延羽卵果蕨Phegopteris decursive-pinnata。
层间植物重要值排名前5位的有攀援星蕨Microsorum brachylepis,香花崖豆藤Callerya dielsiana,爬藤榕Ficus sarmentosa var. impressa,光叶菝葜Smilax glabra,络石Trachelospermum jasminoides。
-
植物物种多样性体现了某一区域的植物资源丰富程度,常用Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数来表征[11]。从表 3可知:Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数在各层的排列顺序均表现为灌木层>乔木层>草本层。灌木层物种数量相比乔木层更为丰富,主要原因是该层树种除了灌木外,还包括了乔木层树种中的幼苗、幼树等,如褐叶青冈、尾叶冬青。
表 3 群落乔木层、灌木层和草本层的物种多样性
Table 3. Community species diversity indices of layers in tree, shrub and herb
层次 Shannon-Wiener多样性指数 Simpson优势度指数 Pielou均匀度指数 乔木层 1.366 6 0.583 9 0.593 5 灌木层 3.002 3 0.929 7 0.844 4 草本层 2.245 5 0.846 9 0.749 5 -
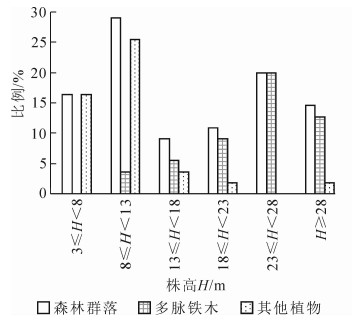
森林群落中多脉铁木的株数占整个乔木层树种总株数的50.9%,因此,将整个乔木层植物划分为两大类群:多脉铁木(1种)和其他植物(8种)。从图 2可知:建群种多脉铁木呈近正态分布的垂直结构,多集中于23 m≤H<28 m的高度范围内,占整个乔木层的20.0%;其他植物的树高低于多脉铁木的高度,多集中于8 m≤H<13 m的高度范围内,占整个乔木层的25.5%,呈明显的偏峰型结构。整个乔木层树高分布规律相对平均,6个高度层各自所占比例均超过10.0%,其中在8 m≤H<13 m和23 m≤H<28 m出现了2个高峰,分别占立木株数的29.1%和20.0%。

图 2 乔木层树种的树高分布
Figure 2. Tree height distribution of tree layer species

图 3 乔木层树种的径阶分布
Figure 3. Diameter distribution of tree layer species
-
与群落的垂直结构相似,多脉铁木的径阶结构呈现出近正态分布的特点(图 3),植物个体数的径阶(D)大多集中在20 cm≤D<25 cm,占整个乔木层的12.7%,其次是30 cm≤D<35 cm,占乔木层的10.9%;而其他植物和整个乔木层树种的径阶分布均为逆J字型,其形状与倒逆放置的J字相似。随着径阶的增大,其个体数量相对减少,即表现为小径阶的大峰和大径阶的小峰,以径阶为5 cm≤D<10 cm为最多,除多脉铁木外的其他树种占乔木层的27.3%,整个乔木层在该径阶所占的比例则高达30.9%;其次为10 cm≤D<15 cm的径阶,其他树种和整个乔木层的比例分别为21.8%和16.4%。
-
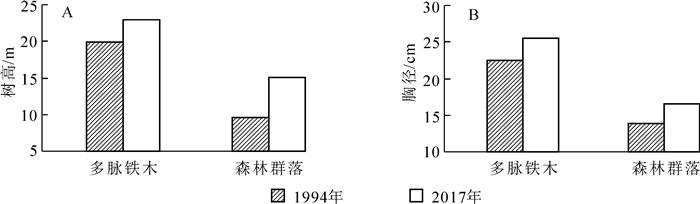
森林群落的垂直结构体现了不同树种的高度,反映了不同植物在群落中的分层结构和生态位[12]。多脉铁木群落是以多脉铁木为建群种组成的稳定性较高的森林群落,具有完整的乔木层、灌木层和草本层,共有维管植物60种,隶属于40科52属。乔木层为复层林,又可分为主林冠层、次林冠层和亚林冠层等3个层次。多脉铁木为阳性树种,居于林冠层之首,树高大于23 m的主林冠层,均以该树种为主,占乔木层总株数的34.5%;次林冠层株高为13~23 m,由多脉铁木、褐叶青冈、缺萼枫香、钩栲组成,占乔木层总株数的20.0%;株高小于13 m的亚林冠层,主要由褐叶青冈、多脉铁木、四照花、披针叶茴香等组成,占乔木层株数的45.5%。从图 4A可知:整个林分平均树高为15.1 m,其中多脉铁木平均树高为23.0 m,与1994年相比[6],整个乔木层和多脉铁木的树高分别增高了5.5,3.0 m,分别增加了57.8%和15.2%。

图 4 1994-2017年林分株高和胸径的变化
Figure 4. Changes of height and diameter during 1994-2017
群落的径阶分布主要有单峰型、间歇性、逆J字型3种类型[11],其中逆J字型是一种成熟稳定的群落结构[13]。该群落的特点是由不同物种一步步从原始生境演化更替而形成[14]。从图 3可知:多脉铁木群落径阶分布为逆J字型,从种群的发展趋势来看,该群落属于成熟稳定的发展阶段,林分平均胸径为16.5 cm,其中多脉铁木平均胸径25.5 cm,与1994年相比[6],整个乔木层和多脉铁木的胸径分别增粗了2.7,3.0 cm,分别增加了19.2%和13.1%(图 4B)。某一树种胸径大小可以反映其年龄水平[15],可以用径阶结构代替时间顺序关系,反映种群动态变化[16]。本研究表明:多脉铁木群落是由不同年龄大小的个体组成的异龄林,种群的年龄结构金字塔基本呈纺锤形(图 3),近似于正态分布,即中龄个体数量占种群总体数量的比例较高,而幼龄和老龄个体较少,表现为稳定的结构特征。相关研究也表明:年龄金字塔呈纺锤形的种群为稳定的群落结构[3]。
文成县石垟林场始建于1958年。20世纪60年代初,林场造林的树种仅仅局限于黄山松Pinus taiwanensis,柳杉Cryptomeria fortunei,杉木Cunninghamia lanceolata,木荷Schima superba,毛竹Phyllostachys edulis,而没有多脉铁木造林的记载[7]。选择的造林地往往是靠近村庄或林场周边土壤深厚、坡度平坦的低丘缓坡,因此在立地条件极差的猴王谷景区倒臼源的陡坡中是不可能营建人工林的,而且该区地表大量覆盖着20~30 cm厚度的大石块,土壤各层大石砾含量均超过50%。另外,限于当时的技术和交通条件,湖北、湖南、四川或贵州等地不可能生产多脉铁木,也不会将该苗木运输到文成县石垟林场进行营建。从多脉铁木的群落特征、物种的垂直和径阶结构、立地条件及科技水平等考虑,该群落并非于20世纪60年代初的人工造林而形成,而是以多脉铁木为建群种组成的阔叶混交林,是从原始生境演替而形成的稳定性较高的森林群落。
在自然条件下,种子向幼苗的转化是物种濒危的关键环节,没有足够数量的幼苗,种群就难以维持。种子产量低、品质差,种子向幼苗的转化率低,形成的幼苗数量少是频危植物存在的较普遍问题[17]。天目铁木Ostrya rehderiana,银杉Cathaya argyrophylla等珍稀濒危植物的种子不饱满、品质低,在野外环境下种子萌发率和成苗率均极低[18-19]。调查发现:研究区域内多脉铁木幼苗数量极少,仅在路边开阔地发现2株;多脉铁木种子的种皮坚硬、透水性差,吸水主要通过苞片网脉,种子不育、空瘪粒多,种子饱满率仅12%。因此种子品质差是多脉铁木走向濒危的内在因素。另外,林地表层有大量石块覆盖,种子较轻、有种翅,成熟后随风散布,很难进入土壤层,种子难以在土壤上着床,不能萌芽成苗,是造成多脉铁木濒危的关键外在因素。
针对多脉动铁木的濒危现状,可以从2个方面考虑该种群的保护和恢复策略:①从多脉铁木个体自身因素考虑,在加强对原生境及植株的保护与监测基础上,加大科研力度,开展多脉铁木生殖生态学研究,如种子萌发率、成苗率和高效繁育技术等,提高实生苗的数量,以扩大其种群分布范围。人工种植,引种栽培并加强推广,扩大其生态位。②从多脉铁木所处生境考虑,适度开展人为干预,搬动部分大石块,展露部分区域土壤,以保证多脉铁木种子顺利入土,促进种子萌发。在此基础上,间伐过密的非目标树种,适当扩大林窗,以降低森林郁闭度,增加林内光照,以利于幼苗的生长。另一方面应积极开展种子繁育、扦插育苗及快繁技术研究,扩大资源量,加强珍稀树种的保护。
Structural characteristics and species diversity for survival of the rare plant Ostrya multinervis
-
摘要: 以浙江省文成县石垟林场的多脉铁木Ostrya multinervis群落为对象,采用标准样地法研究了该群落植物种类、数量特征、物种多样性和群落的垂直和径阶结构。结果表明:该群落资源丰富,共有维管束植物60种(含变种),隶属于40科52属,其中蕨类植物6科7属8种,裸子植物1科1属1种,被子植物33科44属51种。多脉铁木群落垂直结构明显,可分为乔木层、灌木层和草本层,其中乔木层优势树种为多脉铁木,灌木层优势种为披针叶茴香Illicium lanceolatum,草本层优势种为里白Diplopterygium glaucum。群落中的生活型以高位芽植物数量最多,占41.7%。群落中灌木层植物最为丰富,Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数均大于草本层和乔木层。整个林分乔木层树种的树高分布相对均匀,径阶分布为逆J字型,多脉铁木单一树种的垂直和径阶结构均为近正态分布,多脉铁木群落具有较高的物种多样性和群落稳定性。幼苗个体缺乏是导致其濒危的重要原因之一。建议在目前封山育林保护基础上,加强多脉铁木高效繁育技术研究,并适当进行人工抚育,以保证其种群的正常更新。Abstract: Ostrya multinervis is an endangered species to China. To understand the O. multinervis plant community's structural characteristics and species diversity so as to protect this rare and endangered plant. A survey of the community structure and species diversity of O. multinervis was conducted through quadrat sampling at Shiyang Forest Farm, Wencheng County, Zhejiang Province. Using permanent plots and the technique of examining every individual, all trees in quadrats established in the community to quantify the community structural characteristics were identified and measured. Analysis included the Shannon-Wiener, Simpson, and Pielou indexes. Results showed 40 families, 52 genera, and 60 species of vascular plants with eight species of pteridophyte belonging to seven genera in six families, one species of gymnosperms belonging to one genus in one family, and 51 species of angiosperms belonging to 44 genera in 33 families. The vertical structure was composed of three layers:tree layer, shrub layer, and herb layer, in which O. multinervis was the dominant species in the tree layer, Illicium lanceolatum was the dominant species in the shrub layer, and Diplopterygium glaucum was the dominant species in the herb layer. Overall, phanerophytes accounted for the greatest number (41.7%). Species richness for the shrub layer was the largest of the three communities with the Shannon-Wiener, Simpson, and Pielou indexes larger than herb and tree layers(P < 0.05). Tree height distribution in the tree layer for whole trees was relatively uniform, the diameter order was an inverse J type species distribution, and tree height and diameter structure showed near normal distributions for O. multinervis. In summary, O. multinervis communities had a high species diversity and community stability, but were endangered by a lack of young individuals which could be overcome with closure of hillsides to facilitate afforestation, strengthening of research on highly efficient breeding technologies for O. multinervis, and implementation of artificial rearing measures to ensure a normal development of the population.
-
图 1 多脉铁木群落不同生活型谱
Figure 1 Life-form spectrum of survival community of Ostrya multinervis
表 1 多脉铁木群落种子植物属的分布区
Table 1. Genera distribution types of seed plants in O. multinervis community
分布区类型 属数/个 占比/% 1 广布 2 4.4 2 泛热带 9 20.0 3 东亚及热带南美间断 4 8.9 4 旧世界热带 0 0.0 5 热带亚洲至热带大洋洲 1 2.2 6 热带亚洲至热带非洲 0 0.0 7 热带亚洲 6 13.3 8 北温带 7 15.6 9 东亚和北美洲间断 7 15.6 10 旧世界温带 1 2.2 11 温带亚洲 0 0.0 12 地中海地区、西亚至中亚 0 0.0 13 中亚 0 0.0 14 东亚 8 17.8 15 中国特有 0 0 合计 45 100  下载: 导出CSV
下载: 导出CSV
表 2 多脉铁木群落主要植物的重要值
Table 2. Importance values of tree populations in Ostrya multinervis community
乔木层 重要值 灌木层 重要值 草本层 重要值 多脉铁木 62.158 6 披针叶茴香 15.673 5 里白 21.161 2 褐叶青冈 14.517 1 毛花连蕊茶 7.607 3 黑足鳞毛蕨 16.001 8 小叶栎 5.929 1 光叶石楠 6.849 2 麦冬 13.2102 四照花 4.000 7 长裂葛萝槭 5.129 1 求米草 6.789 8 缺萼枫香 3.169 7 红楠 4.786 1 三穗薹草 6.621 1 短尾柯 2.913 7 三尖杉 4.759 9 瘤足蕨 5.926 4 钩栲 2.567 6 猴欢喜 2.301 2 江南卷柏 5.479 5 尾叶冬青 0.980 9 大叶冬青 2.854 5 毛叶腹水草 4.588 5 麂角杜鹃 0.797 1 秀丽槭 1.865 6 大叶唐松草 3.942 2 青榨槭 1.857 1 延羽卵果蕨 3.168 9
下载: 导出CSV
表 3 群落乔木层、灌木层和草本层的物种多样性
Table 3. Community species diversity indices of layers in tree, shrub and herb
层次 Shannon-Wiener多样性指数 Simpson优势度指数 Pielou均匀度指数 乔木层 1.366 6 0.583 9 0.593 5 灌木层 3.002 3 0.929 7 0.844 4 草本层 2.245 5 0.846 9 0.749 5
下载: 导出CSV
-
[1] 马凯, 夏国华, 闫道良, 等.珍稀濒危植物堇叶紫金牛生存群落结构特征及物种多样性[J].浙江农林大学学报, 2012, 29(4):498-509. MA Kai, XIA Guohua, YAN Daoliang, et al. Structural characteristics and species diversity of the endangered plant Ardisia violacea[J]. J Zhejiang A&F Univ, 2012, 29(4):498-509. [2] 王丽敏, 缪心栋, 严彩霞, 等.浙江省小花花椒群落结构与物种多样性[J].浙江农林大学学报, 2013, 30(2):215-219. WANG Limin, MIAO Xindong, YAN Caixia, et al. Structure characteristics and species diversity of Zanthoxylum micranthum in Zhejiang Province[J]. J Zhejiang A&F Univ, 2013, 30(2):215-219. [3] 张婕, 上官铁梁, 段毅豪, 等.灵空山辽东栎种群年龄结构与动态[J].应用生态学报, 2014, 25(11):3125-3130. ZHANG Jie, SHANGGUAN Tieliang, DUAN Yihao, et al. Age structure and dynamics of Quercus wutaishanica population in Lingkong Mountain of Shanxi Province, China[J]. Chin J Appl Ecol, 2014, 25(11):3125-3130. [4] 张若蕙, 刘洪谔, 沈锡康, 等.多脉铁木家化栽培研究[J].浙江林学院学报, 1991, 8(2):167-173. ZHANG Ruohui, LIU Hong'e, SHEN Xikang, et al. Domestication of Ostrya multinervis[J]. J Zhejiang For Coll, 1991, 8(2):167-173. [5] 张若蕙, 楼炉焕, 李根有.浙江珍稀濒危植物[M].杭州:浙江科学技术出版社, 1994:76. [6] 朱昌乐, 叶正环, 胡立帅, 等.多脉铁木林调查分析[J].湖北林业科技, 1994(3):1-4. ZHU Changle, YE Zhenghuan, HU Lishuai, et al. Analysis of Ostrya multinervis forest survey[J]. J Hubei For Sci Technol, 1994(3):1-4. [7] 石垟林场志编篡委员会.石垟林场志[M].北京:北京艺术与科学电子出版社, 2012. [8] 郑昌龙, 王健铭, 李景文.北京松山国家级自然保护区不同林龄胡桃楸林下植物多样性[J].浙江农林大学学报, 2017, 34(5):825-832. ZHENG Changlong, WANG Jianming, LI Jingwen. Undergrowth diversity at different ages of Juglans mandshurica forests in the Songshan National Nature Reserve, Beijing[J]. J Zhejiang A&F Univ, 2017, 34(5):825-832. [9] 高贤明, 陈灵芝.植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J].植物学报, 1998, 40(6):553-559. GAO Xianming, CHEN Lingzhi. The revision of plant life-form system and an analysis of the life-form spectrum of forest plant in the warm temperate zone of China[J]. Acta Bot Sin, 1998, 40(6):553-559. [10] 吴征镒, 周浙昆, 孙喆, 等.种子植物分布区类型及其起源和分化[M].昆明:云南科技出版社, 2006:60-71. [11] 孟莹莹, 周莉, 周旺明, 等.长白山风倒区植被恢复26年后物种多样性变化特征[J].生态学报, 2015, 35(1):142-149. MENG Yingying, ZHOU Li, ZHOU Wangming, et al. Characteristics of plant species diversity in a wind throw area on Changbai Mountain after 26 years of natural recovery[J]. Acta Ecol Sin, 2015, 35(1):142-149. [12] 崔宁洁, 陈小红, 刘洋, 等.不同林龄马尾松人工林林下灌木和草本多样性[J].生态学报, 2014, 34(15):4313-4323. CUI Ningjie, CHEN Xiaohong, LIU Yang, et al. Shrub and herb diversity at different ages of Pinus massoniana plantation[J]. Acta Ecol Sin, 2014, 34(15):4313-4323. [13] 杨同辉, 王良衍.浙江天童舟山新木姜子群落结构及其更新类型[J].浙江林学院学报, 2010, 27(6):877-883. YANG Tonghui, WANG Liangyan. Community structure and regeneration types of Neolitsea sericea in Tiantong, Zhejiang Province[J]. J Zhejiang For Coll, 2010, 27(6):877-883. [14] BOSSUYT B, HONNAY O, HERMY M. Evidence for community assembly constraints during succession in dune slack plant communities[J]. Plant Ecol, 2005, 178(2):201-209. [15] 王磊, 孙启武, 郝朝运, 等.皖南山区南方红豆杉种群不同龄级立木的点格局分析[J].应用生态学报, 2010, 21(2):272-278. WANG Lei, SUN Qiwu, HAO Chaoyun, et al. Point pattern analysis of different age-class Taxus chinensis var. mairein dividuals in mountains area of southern Anhui Province[J]. Chin J Appl Ecol, 2010, 21(2):272-278. [16] 申仕康, 马海英, 王跃华, 等.濒危植物猪血木(Euryodendron excelsum H.T. Chang)自然种群结构及动态[J].生态学报, 2008, 28(5):2404-2412. SHEN Shikang, MA Haiying, WANG Yuehua, et al. The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang[J]. Acta Ecol Sin, 2008, 28(5):2404-2412. [17] 张文辉, 祖元刚, 刘国彬. 10种濒危植物的种群生态学特征及致危因素分析[J].生态学报, 2002, 22(9):1512-1520. ZHANG Wenhui, ZU Yuangang, LIU Guobin. Population ecological characteristics and analysis on endangered cause of ten endangered plant species[J]. Acta Ecol Sin, 2002, 22(9):1512-1520. [18] 乐笑玮, 崔敏燕, 杨淑贞, 等.濒危植物天目铁木种子休眠及萌发特征研究[J].华东师范大学学报(自然科学版), 2013(6):150-158. LE Xiaowei, CUI Minyan, YANG Shuzhen, et al. Characters on the seed dormancy and germination of an endangered species Ostrya rehderiana in Tianmu Mountain, China[J]. J East China Norm Univ Nat Sci, 2013(6):150-158. [19] 曹基武, 刘春林, 张斌, 等.珍稀植物银杉的种子萌发特性[J].生态学报, 2010, 30(15):4027-4034. CAO Jiwu, LIU Chunlin, ZHANG Bin, et al. Seed germination of endangered Cathaya argyrophylla Chun & Kuang[J]. Acta Ecol Sin, 2010, 30(15):4027-4034. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2019.01.005
 点击查看大图
点击查看大图

计量
- 文章访问数: 5320
- HTML全文浏览量: 1141
- PDF下载量: 175
- 被引次数: 0
