




-
近年来,由于生活污水、化肥农药等面源污染物质的排放,太湖水体富营养化日益突出,受到了社会广泛关注。有研究认为,过量使用化肥农药是造成太湖水质低下的主要原因之一,农田径流中氮磷流失量与肥料投入水平显著相关,施肥量远大于作物需求量,氮磷一直处于盈余状态[1]。在土壤-作物系统中,随降水径流和渗漏排出农田的氮素中有20%~25%是当季施用的氮素,而当季作物对磷肥的利用率只有5%~15%,约75%~90%的磷滞留在土壤中。长期过量施用磷肥会导致农田耕层土壤处于富磷状态,磷可通过地表径流等加速向水体迁移[2]。2010−2017年以来太湖水体总氮含量呈大幅度下降趋势,而总磷含量基本呈上升趋势[3]。河岸植被缓冲带是一个由土壤、水、植被等构成的生态系统,可通过植物吸收、土壤吸附等一系列物理、化学和生物等方式,阻止径流水中的磷进入水体[4],被认为是面源污染防治的最佳管理措施[5-6]。不同缓冲带截留磷的效率差异较大,截留功能受河岸带植物状况、宽度、坡度、土壤类型等影响[7]。国外对河岸植被缓冲带开展了较多的研究,主要以森林缓冲带为研究对象[8-9]。中国关于植被缓冲带的研究开展较晚,研究对象多以灌木、草本为主[10-12],而乔木缓冲带研究较少[13],且关于平缓坡度缓冲带的研究相对较少。本研究以太湖流域河岸人工林为研究对象,对比分析平缓坡地上不同宽度、植被组成和林龄河岸植被缓冲带对上游径流水中总磷和可溶性磷的去除差异,以期为太湖流域适宜河岸植被缓冲带的构建提供科学依据。
-
研究区位于江苏省宜兴市周铁镇沙塘港村(31°07′~31°37′N,119°31′~120°03′E)东部,太湖西部沿岸,农田下游,与太湖间相隔一个沿湖大堤。气候类型属亚热带季风气候,四季分明,雨量充沛,气候温和湿润,年均气温为15.7 ℃,无霜期为239 d,年均降水量为1 277.1 mm,多集中于夏季(6−8月)。土壤类型为中性重壤质黄土,土质均匀,土壤容重为1.38 g·cm−3。周边居民主要以农业为主,种植水稻Oryza sativa、小麦Triticum aestivum和油菜Brassica napus 等农作物。树种为太湖流域常见的中山杉Taxodium hybrid‘Zhongshanshan’和‘南林95’杨Populus × euramericana‘Nanlin 95’,林龄6 a,平均树高4.6 m,胸径5.2 cm,林下为自然更新的草本植物,以芦苇Phragmites australis为主。人工林样地共设置400、1 000和1 600株·hm−2等3个林分密度(表1)。
表 1 样地植被配置
Table 1. Allocation of vegetation in the experiment plots
样地设置 植被配置 林分密度/(株·hm−2) 样地设置 植被配置 林分密度/(株·hm−2) 样地1 荒地 0 样地5 中山杉林 1 000 样地2 中山杉林 400 样地6 ‘南林95’杨林 1 000 样地3 ‘南林95’杨林 400 样地7 中山杉林 1 600 样地4 ‘南林95’杨-中山杉混交林 1 000 样地8 ‘南林95’杨林 1 600 -
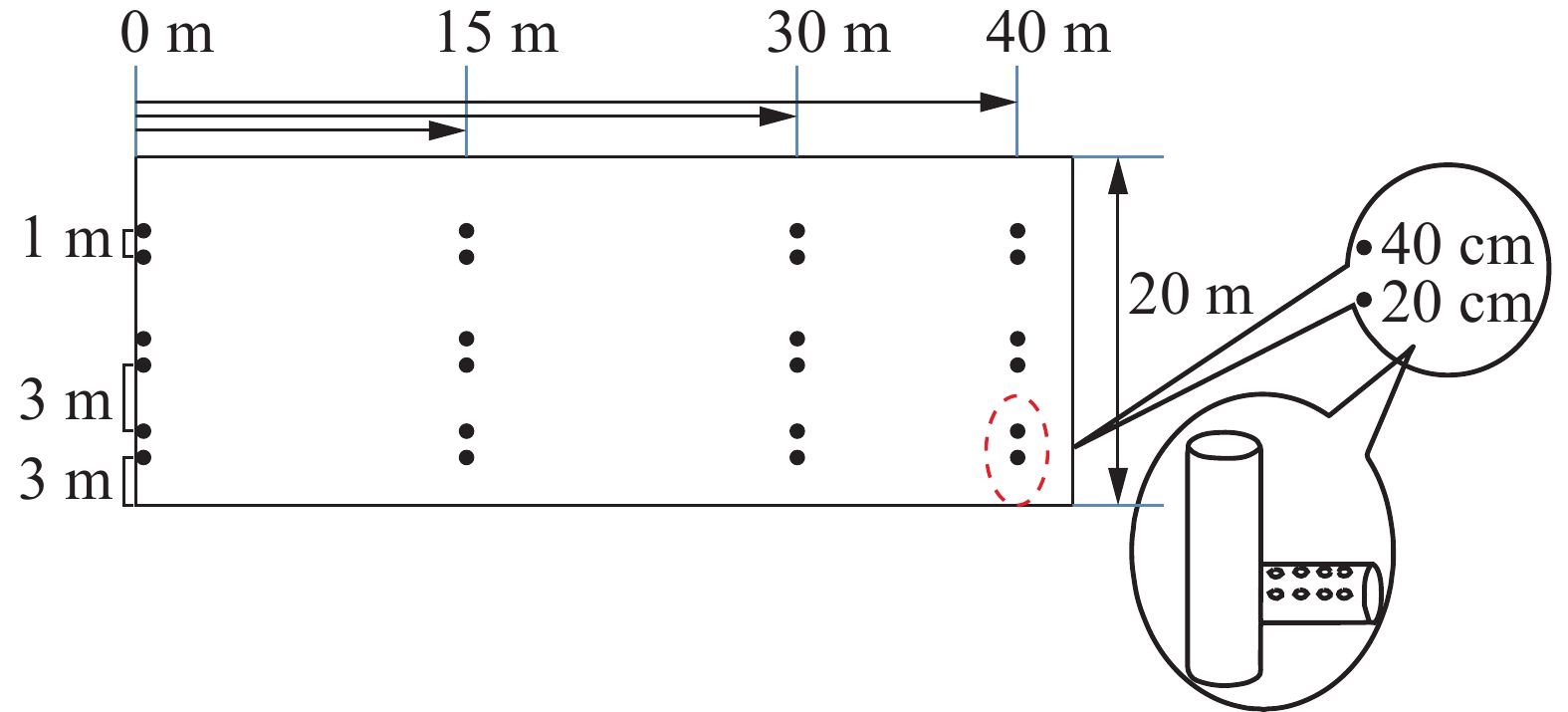
缓冲带分为8个小区(20 m×50 m),坡度比为1∶250。各样地平行排列,与地表径流方向垂直,每个小区相互间隔1 m,用宽60.0 cm、厚1.5 cm的胶合板分隔[14],以减少样地间干扰。在每块样地不同宽度处(沿径流方向分别距离样地起点15、30和40 m)分别埋设PVC淋溶管,收集径流水。每个宽度设3组淋溶管作为重复,每组2个深度(20和40 cm)。每个淋溶管间隔1 m,组与组间隔3 m(图1)。
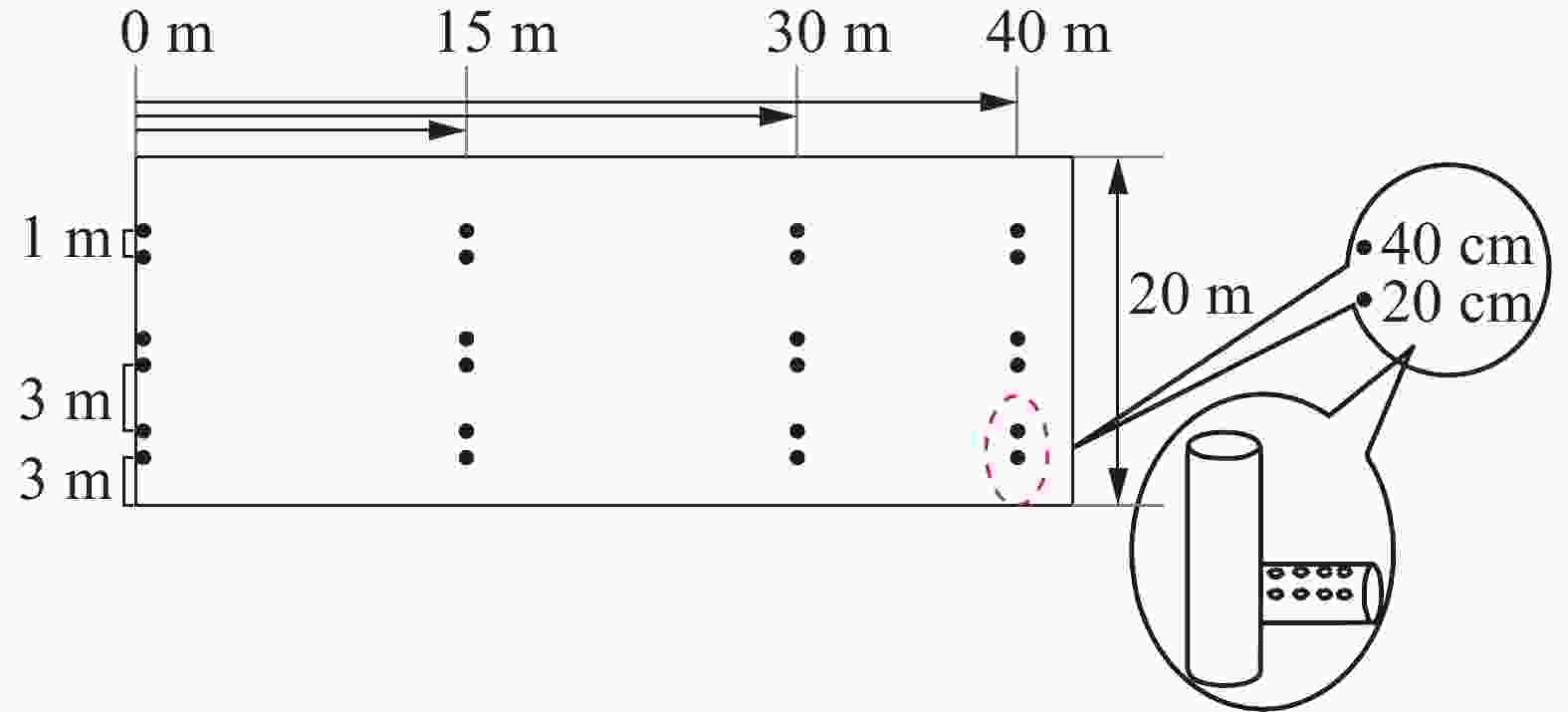
图 1 样地PVC集水管及其铺设示意图
Figure 1. Sketch map of PVC pipe and its laying
-
随径流流失是农田土壤中的磷进入水体的主要途径[2],因此在2014−2017年分别选择当地的雨季(4−9月)进行采样,利用降雨溶解肥料,模拟污染物进入植被缓冲带。在每个样地起始端0~0.5 m处进行施肥,施肥量参考当地农田施肥量,为1 200 kg·hm−2的复合肥(N∶P∶K为16∶8∶16)。在降雨前施肥,降雨结束后24 h内采集样品。用小型水泵抽取淋溶水,装入250 mL塑料瓶中,带回实验室,放入−4~0 ℃冰箱内保存,测定磷。每次采完水样,排空取水管内水,排空的水样在远离采样点之处倾倒。
采用钼锑抗-紫外分光光度法测定水样中总磷(TP)和可溶性磷(DP)[15]。其中待测DP的水样先经过0.45 μm滤膜抽滤预处理后,采用钼锑抗-紫外分光光度法进行测定[16]。
-
径流水中磷去除率计算公式为rP=(P0−Pi)/P0×100%。其中,rP为缓冲带不同宽度径流水中磷累计去除率;Pi为缓冲带i宽度处径流水中磷质量浓度(mg·L−1);P0为缓冲带起始处径流水中磷质量浓度(mg·L−1);i为宽度值(15、30和40 m)。
采用Microsoft Office 2016和SPSS 22.0进行数据处理与分析,所有数据均采用多次重复的平均值±标准误。
-
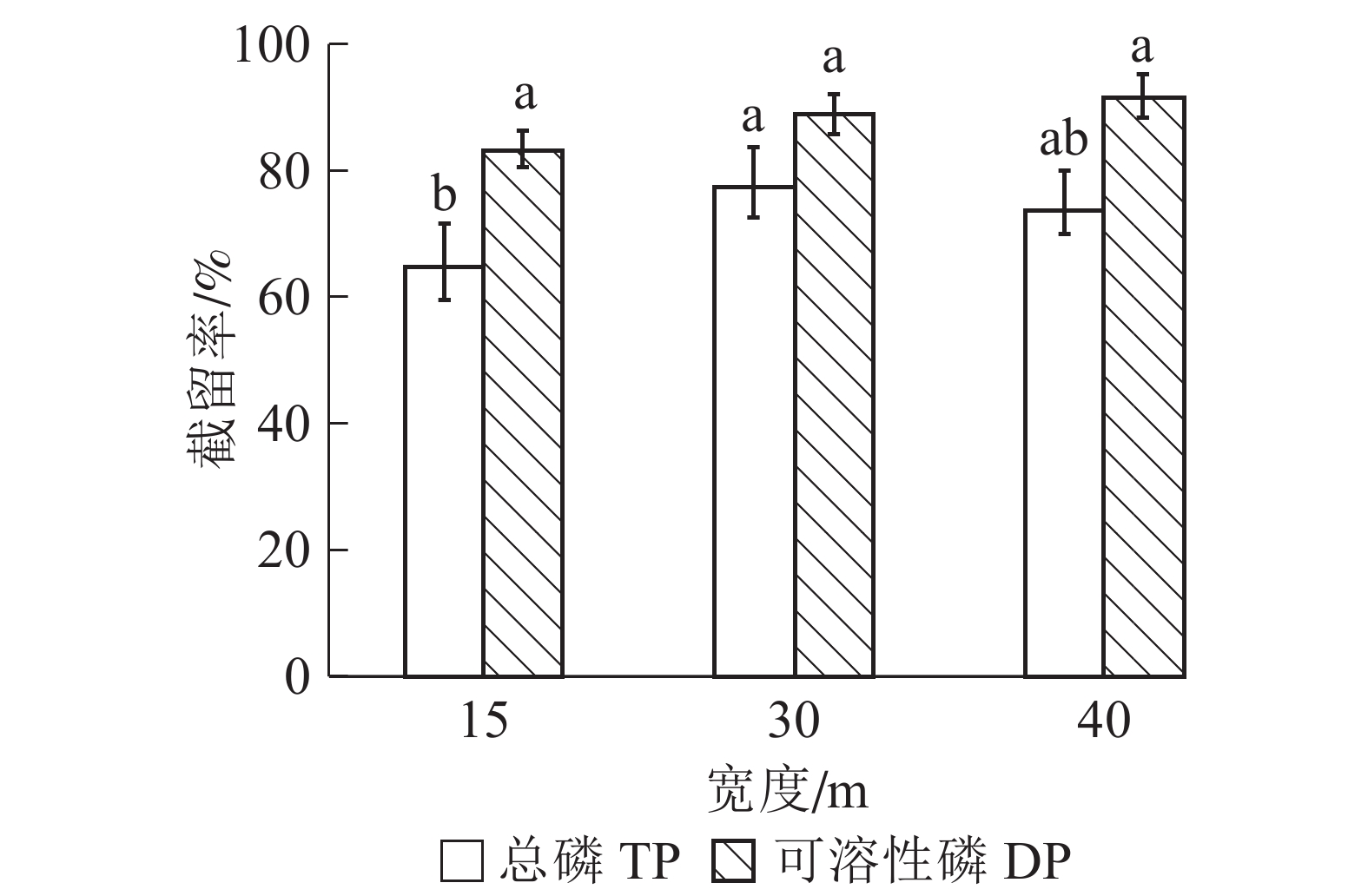
由图2可知:随宽度增加,缓冲带对径流水中总磷截留率呈先上升后下降趋势。在30 m宽度处截留率最高,为77.30%,其次是40 m和15 m,截留率分别下降了3.45%和12.54%。可溶性磷截留率较总磷高,且随缓冲带宽度增加逐渐提高,在40 m处到最大值,为91.50%。方差分析结果表明:15 m与30 m宽缓冲带之间对总磷的截留率差异显著(P<0.05),而30 m与40 m宽度之间差异不显著(P>0.05)。不同宽度缓冲带对可溶性磷的截留率差异不显著(P>0.05)。
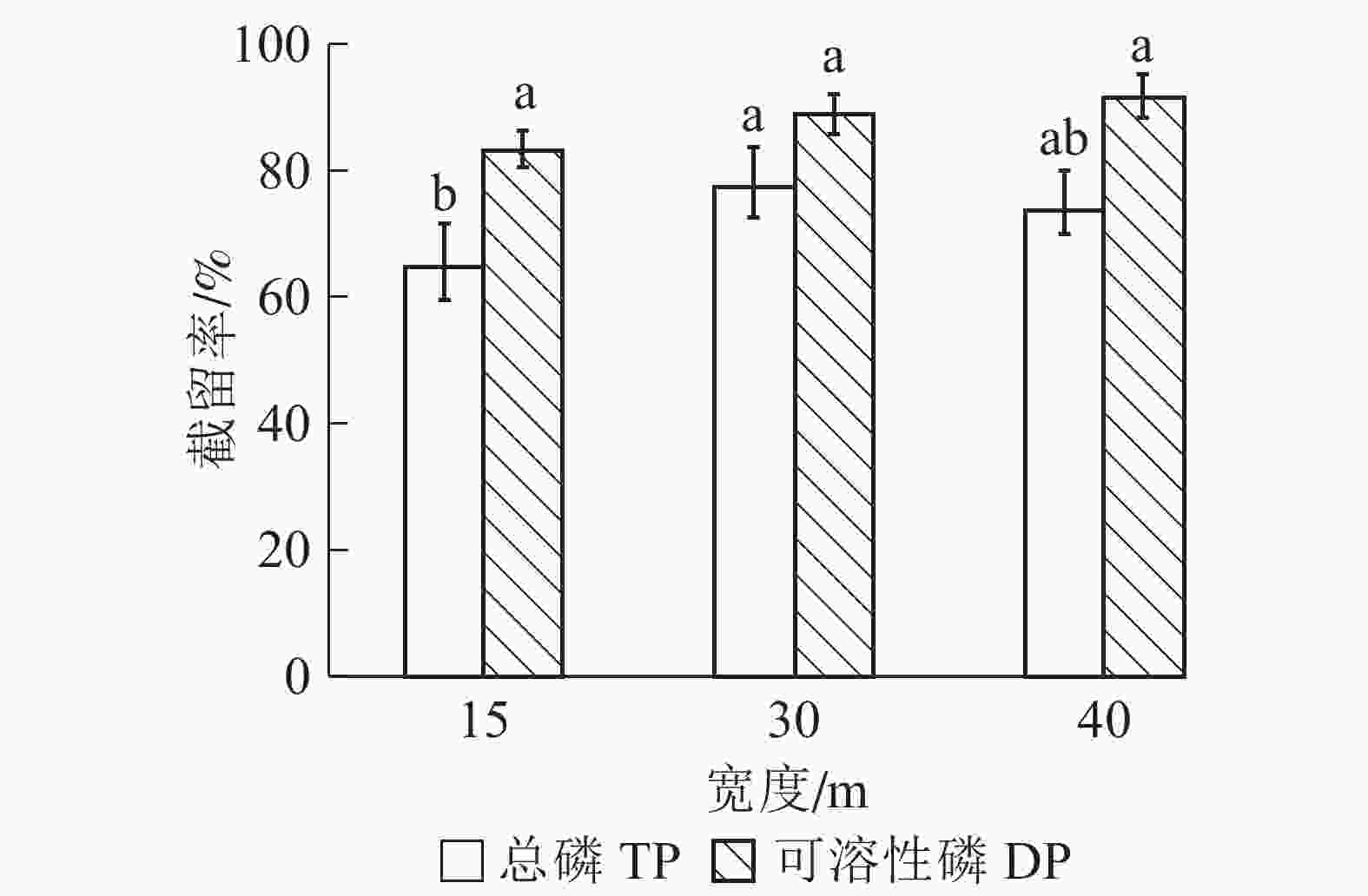
图 2 不同宽度缓冲带对径流水中总磷和可溶性磷截留效果
Figure 2. Removal rates of TP and DP by riparian buffer strips with different widths
-
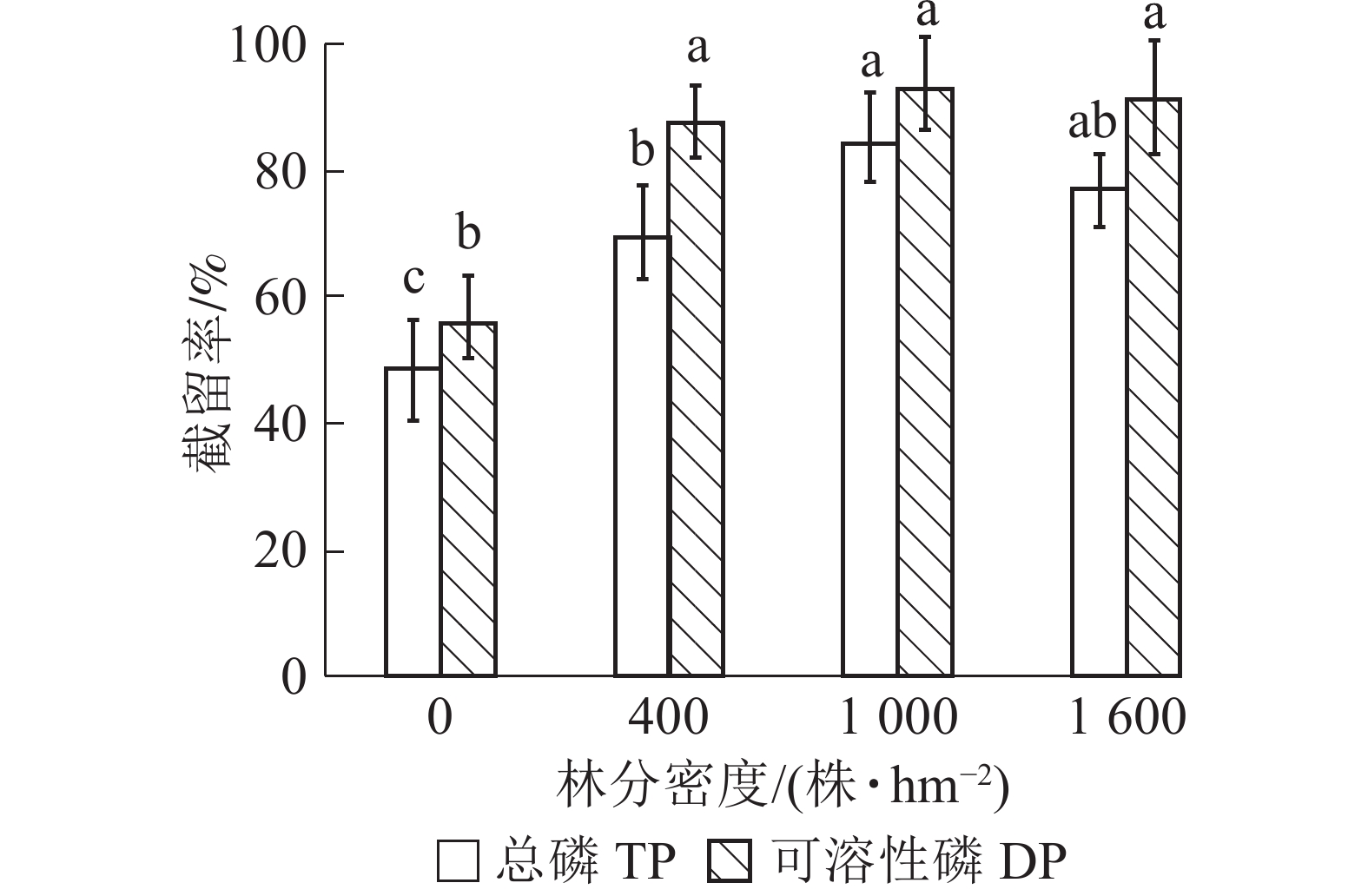
由图3可知:林地对径流水磷的截留率高于荒地。就不同林分密度缓冲带而言,对总磷和可溶性磷的截留效果从高到低依次为1 000、1 600、400株·hm−2,其中1 000株·hm−2缓冲带对总磷和可溶性磷的截留率分别为84.29%和93.25%,比1 600株·hm−2缓冲带分别增加了6.92%林分和2.04%。方差分析结果表明:400和1 000株·hm−2缓冲带对总磷截留效果差异显著(P<0.05),而不同林分密度缓冲带对可溶性磷的截留率差异不显著(P>0.05)。
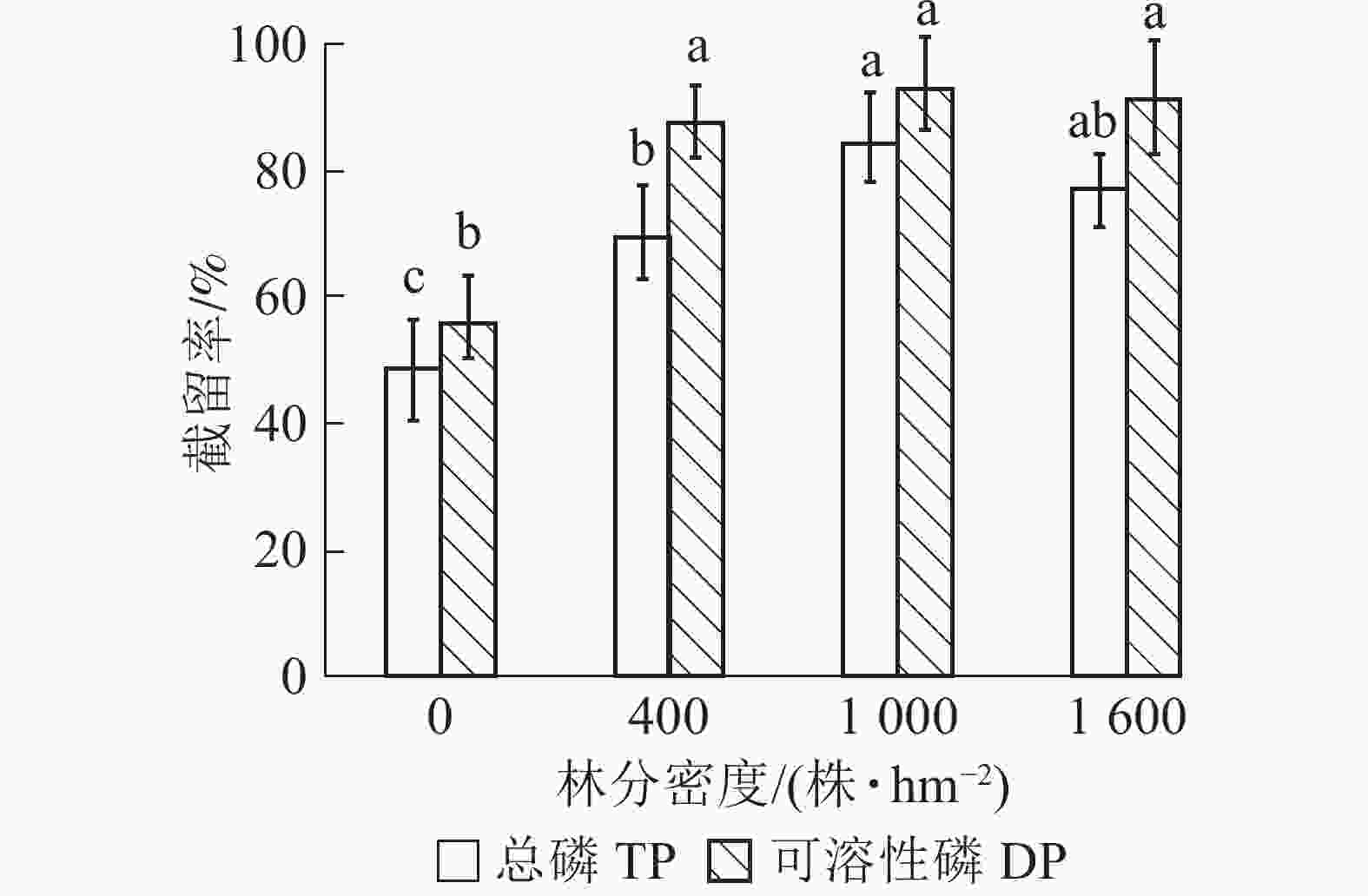
图 3 不同林分密度缓冲带对径流水中总磷和可溶性磷截留效果
Figure 3. Removal rates of TP and DP by riparian buffer strips with different densities
-
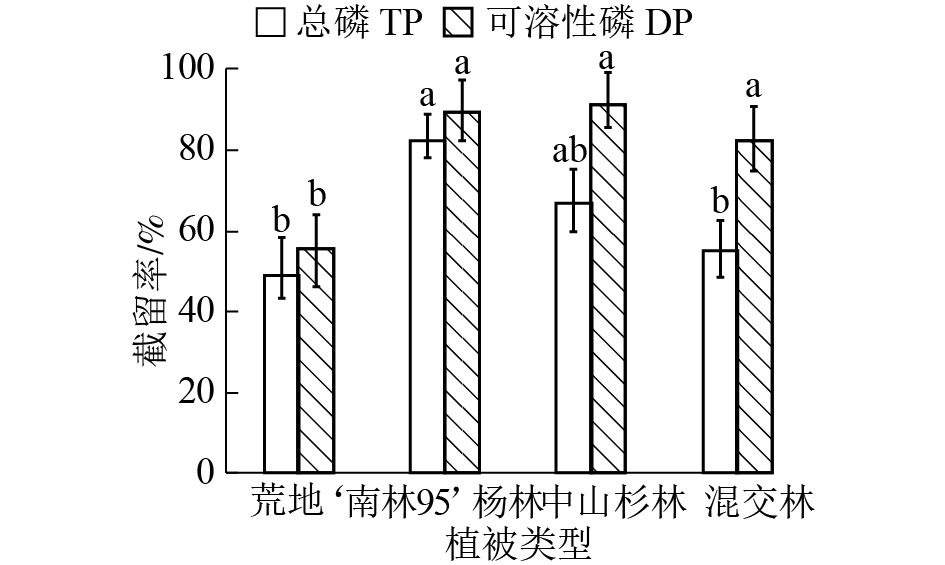
不同植被类型缓冲带对径流水中各形态磷截留效果不同(图4)。‘南林95’杨林缓冲带截留总磷效果较好,截留率为82.37%,中山杉林和混交林缓冲带对总磷的截留率较‘南林95’杨林分别低15.39%和27.10%。相比总磷,不同植被类型缓冲带对可溶性磷截留效果差异不显著(P>0.05)。其中,中山杉林缓冲带对可溶性磷截留率略高于‘南林95’杨林,两者截留率分别为91.28%和89.43%。
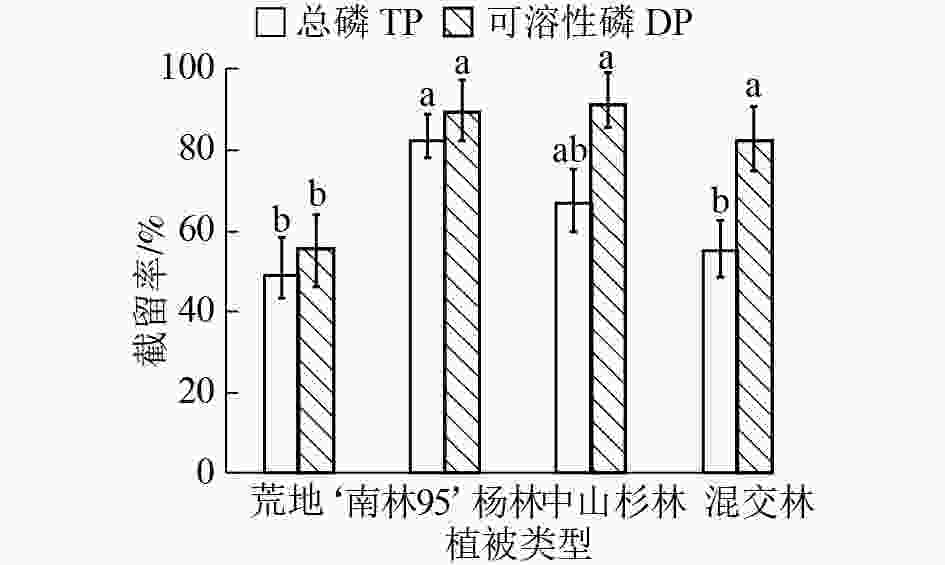
图 4 不同植被类型缓冲带对径流水中总磷和可溶性磷截留效果
Figure 4. Removal rates of TP and DP by riparian buffer strips with different plantations
-
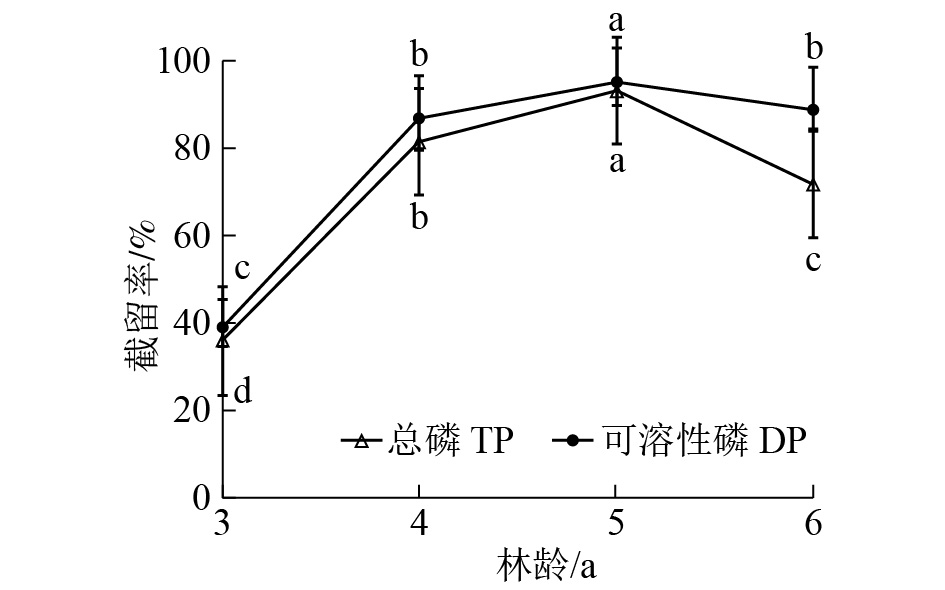
由图5可知:随林龄增长,缓冲带对径流水中总磷和可溶性磷的截留能力逐渐增强,在5年生时截留效果最好,总磷和可溶性磷截留率分别比3年生缓冲带高57.2%和56.37%。6年生缓冲带截留能力较5年生有所下降。单因素方差分析结果表明:不同林龄缓冲带对径流水中总磷和可溶性磷的截留率差异极显著(P<0.01),说明林龄对缓冲带截留磷效果有影响。
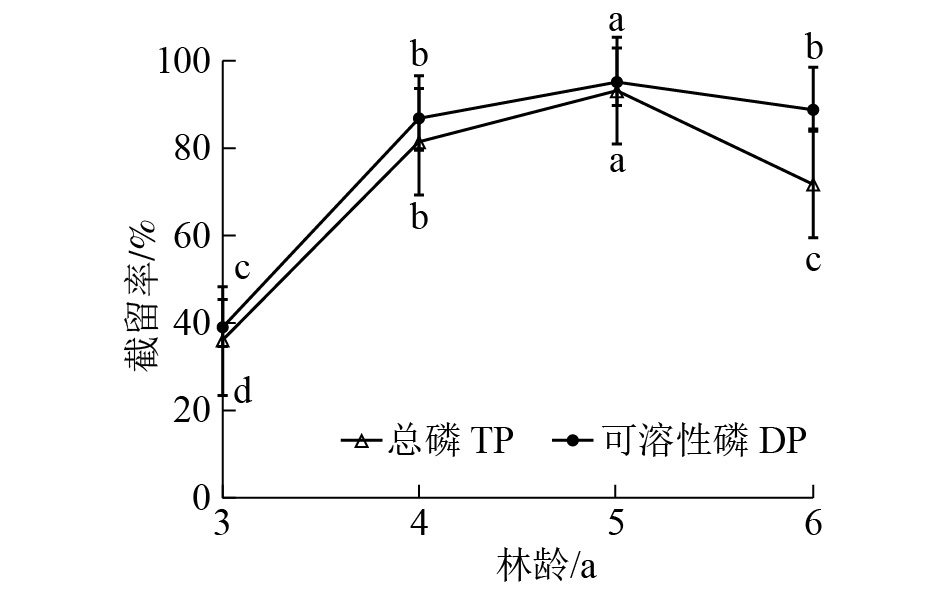
图 5 不同林龄缓冲带对径流水中总磷和可溶性磷截留效果
Figure 5. Dynamic of TP and DP removing by riparian buffer strips with different plantation ages
-
宽度影响缓冲带对磷等污染物的截留效率。研究发现宽度对河岸植被缓冲带生态功能的影响最大[17]。关于河岸缓冲带最佳宽度范围确定的研究较多,但迄今为止没有得到一致的结论[18-20]。何聪等[21]研究发现:缓冲带越宽去除污染物能力越强。WANYAMA等[22]发现缓冲带宽度和去污效果并不呈线性关系。随着宽度增加,单位宽度的缓冲带去除污染物效率不断降低[23]。本研究发现:缓冲带对总磷截留率呈先升高后降低的趋势,且前30 m宽的缓冲带对径流水中总磷截留率变化显著,30 m后变化不显著。总磷截留主要是通过物理拦截和植物吸附[24],可能是前30 m缓冲带截留了大部分磷素,导致被土壤和植物吸附的量减少,截留率下降。可溶性磷的截留率随缓冲带宽度增加而提高,但提高幅度不大,且变化不显著,可能是可溶性磷更依赖水流运动,比吸附在颗粒物上的磷更易通过缓冲带[25]。因此,30 m宽缓冲带就能满足截污要求,与前人的一些研究结果相似[26-28]。
河岸缓冲带对径流水中总磷和可溶性磷的截留效果存在差异。河岸缓冲带可拦截过滤大量附着在沉积物上的磷,尤其是颗粒态磷[29]。当径流水中溶解态磷浓度较高时,磷主要是经过土壤吸附、植物吸收等作用被拦截[7]。缓冲带还可通过减缓径流速度,促进磷的沉积和吸收[30]。研究表明河岸植被缓冲带对总磷的去除效果较可溶性磷好[31-32]。但YOUNG等[33]发现:21 m宽的河岸缓冲带对总磷和可溶性磷的去除率差异不显著。本研究中,缓冲带对径流水中可溶性磷的截留效果较总磷好。缓冲带对总磷和可溶性磷的去除效果不一致,可能是由于不同研究区土壤条件以及植被生长状况不同。
目前,国内外关于乔木缓冲带适宜密度的研究相对较少。JIN等[34]发现:植被缓冲带的密度影响其对地表污染物的截留效果。宋思铭[35]建立了河岸缓冲带植被密度-水质模型,认为河岸缓冲带最适林分密度为1 074株·hm−2。缓冲带拦截污染物能力受植被覆盖度影响[36]。林分密度小,截留污染物能力不足。林分密度大不仅影响草本植物生长,且乔木落叶易增加径流水中的氮磷含量,从而降低对面源氮磷的去除效果[37]。本研究中林分密度为1 000株·hm−2的河岸缓冲带对径流水中磷的截留效果较好,与朱颖等[14]的研究结论一致。
‘南林 95’杨林缓冲带截留径流水磷效果较中山杉好。‘南林 95’杨树为速生树种,能在短期内积累养分[38],且根系浅,主要分布于地表0~15 cm深度内,而中山杉是深根系树种,表层吸收少。有研究表明,根际土壤微生物数量比非根际多10~100倍,对根际微环境有显著影响[39]。当径流水中磷质量浓度过高时,‘南林 95’根际微生物可吸收过量的磷,对浅层土壤磷的拦截起着重要作用。混交林相比另外2种纯林截留率较低,可能是由于树种间他感作用造成不同树种吸收养分能力受到抑制。
本研究表明:随着林龄增加,缓冲带对磷截留率明显提高,5年生时截留率达90%以上。MANDER等[36]和SNYDER等[40]研究表明:缓冲带在林龄较小时截留污染物能力更强,认为幼龄植被生长需吸收大量的磷,土壤及土壤微生物的活动能力和吸附作用更好,保留更多的养分用于树木的生长,而林龄较大时缓冲带养分的输入和输出一般处于平衡状态,表现为对磷的需求量减少。
磷肥污染只是农业面源污染的一种类型,因此研究植被缓冲带功能还应考虑其他污染物质的影响。磷截留是个复杂的过程,地表径流中的磷素除了被乔木吸收截留外,林下的草本植物也起到一定作用[24],今后可增加这方面的研究,以深入分析缓冲带截留与吸收磷的过程和机制。
Effects of riparian plantation buffer strips on phosphorus interception in runoff water
-
摘要:
目的 含磷等农业面源污染物的大量排放,已成为太湖水体富营养化的主要原因之一。河岸植被缓冲带能通过吸收、截留,减少面源污染物进入水体。在平缓坡地上构建适宜的河岸植被缓冲带,研究河岸植被缓冲带对磷的截留效率,为减缓太湖农业面源污染提供科学依据。 方法 以太湖河岸人工林缓冲带为研究对象,分析缓冲带在不同宽度(15、30、40 m)、不同林分密度(400、1 000、1 600株·hm−2)、不同植物组成(‘南林95’杨Populus × euramericana‘Nanlin 95’林、中山杉Taxodium hybrid ‘Zhongshanshan’林、‘南林95’杨-中山杉混交林)以及不同林龄(3、4、5、6 a)方面对径流水中总磷和可溶性磷截留效果的差异,确定适宜的人工林缓冲带结构。 结果 30 m宽的河岸缓冲带可很好地截留径流水中磷素,总磷截留率在30 m处最高,为77.30%,缓冲带对可溶性磷的截留率随宽度增加而提高,截留率在不同宽度间差异不显著(P>0.05);对径流水中总磷和可溶性磷截留效果较好的河岸缓冲带的林分密度为1 000株·hm−2,截留率分别为84.29%和93.25%;杨树林缓冲带对径流水中总磷的截留率较高,达82.37%,中山杉林缓冲带对可溶性磷截留率略高于杨树林,不同植物组成缓冲带对可溶性磷的截留效果差异不显著(P>0.05),河岸缓冲带对径流水中可溶性磷的截留效果较总磷好;随林龄增加,缓冲带对径流水中磷的截留率呈上升趋势(P<0.01),5年生时截留率最高。 结论 30 m宽的河岸缓冲带基本能满足截污需求,且杨树林缓冲带截留污染物效果较好。缓冲带在林龄较小时截留磷能力较强。图5表1参40 Abstract:Objective The discharge of agricultural non-point source pollutants such as phosphorus has become one of the major causes for the eutrophication in Taihu Lake. Riparian vegetation buffer strips can reduce non-point source pollution from agricultural area into water through absorption and interception. The riparian vegetation strips were built on gentle slopes to study the efficiency of retaining phosphorus so as to provide a scientific basis for mitigating the pollution of agricultural non-point sources in Taihu Lake. Method Taking the buffer strip of plantation along the bank of Taihu Lake as the research object, the interception effects of riparian plantation buffer strips with different widths (15, 30, 40 m), densities (400, 1 000, 1 600 trees ·hm−2 ), plantation types (Populus × euramericana ‘Nanlin 95’ plantation, Taxodium hybrid ‘Zhongshanshan’ plantation and ‘Nanlin 95’-‘Zhongshanshan’ mixed plantation) and ages (3, 4, 5, 6 a) on total phosphorus (TP) and dissolved phosphorus (DP) in surface runoff water were studied to determine the suitable structure of vegetation buffer strips. Result The 30 m wide buffer strips had the optimal effect on intercepting phosphorus in runoff water, and the TP interception rate was the highest at 30 m, 77.30%. The interception rate of DP increased with the increase of buffer strips width but there existed no significant difference between different widths of buffer strips in terms of DP interception rate (P>0.05). The stand density of riparian buffer strip with good interception effect on TP and DP in runoff water was 1 000 trees ·hm−2, and the interception rate was 84.29% and 93.25%, respectively. The ‘Nanlin 95’ plantation could intercept 82.37% TP in runoff water while the interception rate of ‘Nanlin 95’- ‘Zhongshanshan’, the mixed plantation on DP was a little higher than that of ‘Nanlin 95’ plantation, but the interception rate of DP in the buffer strips of different plant compositions was not significant different (P>0.05). The effect of buffer strips on interception of DP in runoff water was better than that on TP. With the increase of plantation age, the interception rate of phosphorus in runoff water increased gradually (P<0.01), and reached the highest at the age of 5. Conclusion The 30 m wide buffer strips can meet the requirement of pollution interception, and the ‘Nanlin 95’ plantation buffer strips have good interception effect on phosphorus. Besides, the buffer strips have stronger phosphorus interception capacity at a young age. [Ch, 5 fig. 1 tab. 40 ref.] -
图 2 不同宽度缓冲带对径流水中总磷和可溶性磷截留效果
不同字母表示同一类型磷不同处理间差异显著(P<0.05)
Figure 2 Removal rates of TP and DP by riparian buffer strips with different widths
图 3 不同林分密度缓冲带对径流水中总磷和可溶性磷截留效果
不同字母表示同一类型磷不同处理间差异显著(P<0.05)
Figure 3 Removal rates of TP and DP by riparian buffer strips with different densities
图 4 不同植被类型缓冲带对径流水中总磷和可溶性磷截留效果
不同字母表示同一类型磷不同处理间差异显著(P<0.05)
Figure 4 Removal rates of TP and DP by riparian buffer strips with different plantations
图 5 不同林龄缓冲带对径流水中总磷和可溶性磷截留效果
不同字母表示同一类型磷不同处理间差异显著(P<0.05)
Figure 5 Dynamic of TP and DP removing by riparian buffer strips with different plantation ages
表 1 样地植被配置
Table 1. Allocation of vegetation in the experiment plots
样地设置 植被配置 林分密度/(株·hm−2) 样地设置 植被配置 林分密度/(株·hm−2) 样地1 荒地 0 样地5 中山杉林 1 000 样地2 中山杉林 400 样地6 ‘南林95’杨林 1 000 样地3 ‘南林95’杨林 400 样地7 中山杉林 1 600 样地4 ‘南林95’杨-中山杉混交林 1 000 样地8 ‘南林95’杨林 1 600  下载: 导出CSV
下载: 导出CSV
-
[1] 闫丽珍, 石敏俊, 王磊. 太湖流域农业面源污染及控制研究进展[J]. 中国人口·资源与环境, 2010, 20(1): 103 − 111. YAN Lizhen, SHI Minjun, WANG Lei. Research progress of agricultural non-point source pollution and control in Taihu Lake Basin [J]. China Popul Resour Environ, 2010, 20(1): 103 − 111. [2] 司友斌, 王慎强, 陈怀满. 农田氮、磷的流失与水体富营养化[J]. 土壤, 2000, 32(4): 188 − 193. SI Youbin, WANG Shenqiang, CHEN Huaiman. Loss of nitrogen and phosphorus in farmland and eutrophication of water [J]. Soil, 2000, 32(4): 188 − 193. [3] 王华, 陈华鑫, 徐兆安, 等. 2010−2017年太湖总磷浓度变化趋势分析及成因探讨[J]. 湖泊科学, 2019, 31(4): 919 − 929. WANG Hua, CHEN Huaxin, XU Zhaoan, et al. Trend analysis of total phosphorus concentration in Taihu Lake from 2010 to 2017 and discussion on its causes [J]. Lake Sci, 2019, 31(4): 919 − 929. [4] HOFFMANN C C, KJAERGAARD C, UUSI-KÄMPPÄ J, et al. Phosphorus retention in riparian buffers: review of their efficiency [J]. J Environ Qual, 2009, 38(5): 1942 − 1955. [5] MARC D, RICHARD H. Reduction in agricultural non-point source pollution in the first year following establishment of an integrated grass/tree filter strip system in southern Quebec (Canada) [J]. Agric Ecosyst Environ, 2009, 131(1): 85 − 97. [6] MESSER T L, BURCHELL M R I, GRABOW G L, et al. Groundwater nitrate reductions within upstream and downstream sections of a riparian buffer [J]. Ecol Eng, 2012, 47(5): 297 − 307. [7] 吴永波. 河岸植被缓冲带减缓农业面源污染研究进展[J]. 南京林业大学学报(自然科学版), 2015, 39(3): 143 − 148. WU Yongbo. Research progress on the riparian vegetation buffer strip functions on agricultural nonpoint source pollution reduction [J]. J Nanjing For Univ Nat Sci Ed, 2015, 39(3): 143 − 148. [8] RICHARDSON J S, NAIMAN R J, BISSON P A. How did fixed-width buffers become standard practice for protecting freshwaters and their riparian areas from forest harvest practices? [J]. Freshwater Sci, 2012, 31(1): 232 − 238. [9] VIRGÓS E. Relative value of riparian woodlands in landscapes with different forest cover for medium-sized Iberian carnivores [J]. Biodiversity Conserv, 2001, 10(7): 1039 − 1049. [10] 张广分. 潮白河上游河岸植被缓冲带对氮、磷去除效果研究[J]. 中国农学通报, 2013, 29(8): 189 − 194. ZHANG Guangfen. The effects of riparian vegetation buffer zones on nitrogen and phosphorus reduction in Chaobai River upstream [J]. Chin Agric Sci Bull, 2013, 29(8): 189 − 194. [11] 李萍萍, 崔波, 付为国, 等. 河岸带不同植被类型及宽度对污染物去除效果的影响[J]. 南京林业大学学报(自然科学版), 2013, 37(6): 47 − 52. LI Pingping, CUI Bo, FU Weiguo, et al. Effects of riparian buffer vegetation types and width on pollutant removal [J]. J Nanjing For Univ Nat Sci Ed, 2013, 37(6): 47 − 52. [12] 朱晓成, 吴永波, 余昱莹, 等. 太湖乔木林河岸植被缓冲带截留氮素效率[J]. 浙江农林大学学报, 2019, 36(3): 565 − 572. ZHU Xiaocheng,WU Yongbo,YU Yuying, et al. Removing nitrogen with trees planted in the riparian vegetation buffer strips of Taihu Lake [J]. J Zhejiang A&F Univ, 2019, 36(3): 565 − 572. [13] 王珊珊, 张帅, 王诗乐, 等. 河岸带乔木树种配置方式对面源污染物 ${\rm{NH}}_4^{+} $ [14] 朱颖, 吴永波, 李文霞, 等. 河岸人工林缓冲带截留磷素能力及适宜宽度[J]. 东北林业大学学报, 2016, 44(12): 31 − 36. ZHU Ying, WU Yongbo, LI Wenxia, et al. Phosphorus retention capacities and fitting width of different riparian plantation buffer strips [J]. J Northeast For Univ, 2016, 44(12): 31 − 36. [15] 雷立改, 马晓珍, 魏福祥, 等. 水中总氮、总磷测定方法的研究进展[J]. 河北工业科技, 2011, 28(1): 72 − 76. LEI Ligai, MA Xiaozhen, WEI Fuxiang, et al. Research progress of determination of total nitrogen and total phosphorus in seawater [J]. Hebei J Ind Sci Technol, 2011, 28(1): 72 − 76. [16] 高励珍, 郝茜, 张术杰. 分光光度法测定氯化稀土, 碳酸轻稀土中磷酸根量[J]. 稀土, 2016, 37(1): 116 − 120. GAO Lizhen, HAO Qian, ZHANG Shujie. Determination of phosphorus radicle content in rare earth chloride and light rare earth carbonate by phosphorus-antimonate-molybdenum blue spectrophotometric [J]. Chin Rare Earths, 2016, 37(1): 116 − 120. [17] 饶良懿, 崔建国. 河岸植被缓冲带生态水文功能研究进展[J]. 中国水土保持科学, 2008, 6(4): 121 − 128. RAO Liangyi, CUI Jianguo. Research advances on the eco-hydrological functions of riparian buffer [J]. Sci Soil Water Conserv, 2008, 6(4): 121 − 128. [18] 赵警卫, 胡彬. 河岸带植被对非点源氮、磷以及悬浮颗粒物的截留效应[J]. 水土保持通报, 2012, 32(4): 51 − 55. ZHAO Jingwei, HU Bin. Interception of non-point source N and P and suspended solid matter by riparian vegetation [J]. Bull Soil Water Conserv, 2012, 32(4): 51 − 55. [19] LEF K H, ISENHART T M, SCHULTZ R C, et al. Multispecies riparian buffers trap sediment and nutrients during rainfall simulations [J]. J Environ Qual, 2000, 29(4): 1200 − 1205. [20] DORIOZ J M, WANG D, POULENARD J, et al. The effect of grass buffer strips on phosphorus dynamics-a critical review and synthesis as a basis for application in agricultural landscapes in France [J]. Agric Ecosyst Environ, 2007, 117(1): 4 − 21. [21] 何聪, 刘璐嘉, 王苏胜, 等. 不同宽度草皮缓冲带对农田径流氮磷去除效果研究[J]. 水土保持研究, 2014, 21(4): 55 − 58. HE Cong, LIU Lujia, WANG Susheng, et al. Study on efficiency of sward buffer strips with different structures on removal of nitrogen and phosphorus in farmland runoff [J]. Res Soil Water Conserv, 2014, 21(4): 55 − 58. [22] WANYAMA J, KRISTOFF H, ISABIRYE M, et al. Evaluation of runoff and sediment trapping effectiveness of vegetative filter strips in the riparian zone of Lake Victoria [J]. Geophysic Res Abstr, 2011, 13: 48 − 63. [23] SCHMITT T J, DOSSKEY M G, HOAGLAND K D. Filter strip performance and processes for different vegetation, widths, and contaminants [J]. J Environ Qual, 1999, 28(5): 1479 − 1489. [24] MANKIN K R, NGANDU D M, BARDEN C J, et al. Grass-shrub riparian buffer removal of sediment, phosphorus, and nitrogen from simulated runoff 1 [J]. J Am Water Resour Assoc, 2007, 43(5): 1108 − 1116. [25] ALLAIRE S E, SYLVAIN C, LANGE S F, et al. Potential efficiency of riparian vegetated buffer strips in intercepting soluble compounds in the presence of subsurface preferential flows [J]. PLoS One, 2015, 10(7): e0131840. [26] CASTELLE A J, JOHNSON A W, CONOLLY C. Wetland and stream buffer size requirements: a review [J]. J Environ Qual, 1994, 23(5): 878 − 882. [27] 阎丽凤. 河岸缓冲带对氮、磷污染物的去除效果研究[D]. 阜新: 辽宁工程技术大学, 2011. YAN Lifeng. The Effects of Riparian and Phosphorus Reduction on Riparian Buffer Zone[D]. Fuxin: Liaoning Technology University, 2011. [28] SWEENEY A W, NEWBOLD J D. Streamside forest buffer width needed to protect stream water quality, habitat, and organisms: a literature review [J]. J Am Water Resour Assoc, 2014, 50(3): 560 − 584. [29] 钱进, 王超, 王沛芳, 等. 河湖滨岸缓冲带净污机理及适宜宽度研究进展[J]. 水科学进展, 2009, 20(1): 141 − 146. QIAN Jin, WANG Chao, WANG Peifang, et al. Research progresses in purification mechanism and fitting width of riparian buffer strip [J]. Adv Water Sci, 2009, 20(1): 141 − 146. [30] UUSI-KÄMPPÄ J. Phosphorus purification in buffer zones in cold climates [J]. Ecol Eng, 2005, 24(5): 491 − 502. [31] DANIELS R B, GILLIAM J W. Sediment and chemical load reduction by grass and riparian filters [J]. Soil Sci Soc Am J, 1996, 60(1): 246 − 251. [32] BARLING R D, MOORE I D. Role of buffer strips in management of waterway pollution: a review [J]. Environ Manage, 1994, 18(4): 543 − 558. [33] YONG R A, HUNTRODS T, ANDERSON W. Effectiveness of vegetated buffer strips in controlling pollution from feedlot runoff [J]. J Environ Qual, 1980(9): 483 − 487. [34] JIN C X, RÖMKENS M J M, GRIFFIOEN F. Estimation manning's roughness coefficient for shallow overland flow in non-submerged vegetative filter strips [J]. Trans ASAE, 2000, 43(6): 1459 − 1466. [35] 宋思铭. 河岸缓冲带净水效果及优化配置技术研究[D]. 北京: 北京林业大学, 2012. SONG Siming. Research on Water Purification Effect and Optimizing Allocation Technology of Riparian Buffer Strips[D]. Beijing: Beijing Forestry University, 2012. [36] MANDER U, KUUSEMETS V, LOHMUS K. Efficiency and dimensioning of riparian buffer zones in agricultural catchments [J]. Ecol Eng, 1997, 8(4): 299 − 324. [37] 郭蔚华, 徐灵华, 王柱. 桃花溪河岸生态建设中植物配置与面源氮磷削减[J]. 科技导报, 2010, 28(7): 50 − 54. GUO Weihua, XU Linghua, WANG Zhu. The relationship between the plant disposition and the reduction of nitrogen and phosphorus in the riparian ecological construction of Taohua River in Chongqing [J]. Sci Technol Rev, 2010, 28(7): 50 − 54. [38] FORTIER J, GAGNON D, TRUAX B, et al. Biomass and volume yield after 6 years in multiclonal hybrid poplar riparian buffer strips [J]. Biomass Bioenergy, 2010, 34(7): 1028 − 1040. [39] 王超, 尹炜, 贾海燕, 等. 滨岸带对河流生态系统的影响机制研究进展[J]. 生态科学, 2018, 37(3): 222 − 232. WANG Chao, YIN Wei, JIA Haiyan, et al. Review on the influence mechanism of riparian zone to the river ecosystem [J]. Ecol Sci, 2018, 37(3): 222 − 232. [40] SNYDER N J, MOSTAGHIMI S, BERRY D F, et al. Impact of riparian forest buffers on agricultural nonpoint source pollution [J]. J Am Water Resour Assoc, 1998, 34(2): 385 − 395. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190509
 点击查看大图
点击查看大图
计量
- 文章访问数: 2411
- HTML全文浏览量: 720
- PDF下载量: 265
- 被引次数: 0
