




-
樟子松Pinus sylvestris var. mongolica为松科Pinaceae松属Pinus常绿乔木,因其具有耐寒、耐干旱和对土壤要求不严等优良特性,被“三北”风沙区大规模引种栽植[1]。作为最早引种樟子松的地区,科尔沁沙地东南缘的章古台地区已形成了不同林龄樟子松共存的格局。然而,自20世纪90年代初以来,由于人为及多种自然因素的影响,章古台地区樟子松人工林出现了十分严重的衰退现象[2],并且该衰退现象已在全国多省的樟子松人工林中相继出现[3]。衰退现象的发生势必对“三北”风沙区防护林建设和沙地治理造成极大的损失和影响,因此必须加强对樟子松人工林衰退问题的研究。生态化学计量学是研究生物系统中多重化学元素平衡的科学[4],重点关注活有机体主要组成元素碳(C)、氮(N)、磷(P)的生态化学计量特征关系,是研究土壤养分循环与限制作用的重要工具。已有研究表明:碳、氮、磷是植物生长所必需的元素,耦合作用相较其他元素更强,与植物生长的关系十分密切[5]。土壤是林木生长的物质基础,而林木根际是林木和土壤进行物质、能量交换的场所,也是最强的生化活性区域[6]。根际是植物细根周围的土壤区域[7],该部分土壤受植物根系生长的调控,可从周围环境聚集养分,被称为根际土壤[8]。根际土壤中的根系分泌物是土壤有机碳的重要来源之一[9],植物通过根系改变植物生长过程中的土壤理化性质,影响土壤微生物的活动和群落结构组成[10]。因此,根际土壤能更确切地反映林木生长受土壤状况制约的程度,就樟子松林衰退问题,研究其根际与非根际土壤差异无疑是最直接有效的方法。目前,学者们对樟子松化学计量方面的研究主要为单一林龄下土壤与植物或不同林龄下单一组分的化学计量特征[11-14],以根际与非根际土壤为切入点,对不同林龄樟子松人工林根际与非根际土壤碳、氮、磷化学计量特征的研究尚未见报道。本研究以6个不同林龄(10、20、30、40、50和60 a)樟子松人工林为对象,分析林龄对根际与非根际土壤碳、氮、磷质量分数及化学计量特征的影响,为该地区的樟子松林培育、可持续经营及管理提供理论依据。
-
研究区位于辽宁省沙地治理与利用研究所章古台试验基地(42°39′~42°43′N,122°23′~122°33′E),是中国最早引种樟子松的地区,更是“三北”防护林重要的组成部分。该区地处中温带,属典型的亚湿润大陆性季风气候,年平均气温为5.5 ℃,1月平均气温为−16.3 ℃,7月平均气温为23.9 ℃;年平均降水量为450~550 mm,其中有近70%的降水集中在6−8月,年蒸发量可达1 300~1 800 mm;该地年平均风速为4.5 m·s−1,春冬季风尤烈,风速可达5.0 m·s−1。该研究区土壤主要为风沙土,植被多为抗旱性较强的沙生植物。代表性植物有樟子松、盐蒿Artemisia halo-dendron、小黄柳Salix gordejevii、榆树Ulmus pumila、大果榆Ulmus macrocarpa、中华委陵菜Potentilla anserina、中华隐子草Cleistogenes chinensis、胡枝子Lespedeza daurica、野古草Arundinella anomala和小白蒿Artemisia frigida等。
-
经过对研究区樟子松人工林的充分调查,选取土壤类型和立地条件基本一致的6个林龄(10、20、30、40、50和60 a)樟子松林样地,并保证样地在营造樟子松人工林前均为固定沙地。在每个林龄的样地内分别设置3块20 m×20 m的样方,在样方内进行每木检尺,记录株高、胸径、冠幅,并选择3株平均木作为标准木。样地基本情况见表1。于2020年6月对标准木根际与非根际土壤进行采集。根际土样采集方法为抖落法[15],在树冠投影范围内,对根系分布较为密集的0~20 cm土层进行挖掘,仔细挑拣出其中的植物根系,先轻轻抖落不含根系的大块土壤,然后将附着在根系表面2 mm厚度的土壤作为根际土壤,抖落入自封袋,为根际土壤样品。同时,用土钻在每个样方内按“S”型采集0~20 cm土层根系外部土壤样品5个,并保证5个钻孔在树冠投影内、外均有分布,将土样充分混合均匀后带回实验室,为非根际土壤样品。对根际与非根际土壤样品进行风干处理,并剔除石头等杂物,研磨并过0.25 mm的网筛后待测。
表 1 样地基本情况
Table 1. Basic information of the sample plots
样地
编号林分
年龄/a平均树
高/m平均胸
径/cm东西平均
冠幅/m南北平均
冠幅/m土壤容重/
(g·cm−3)1 10 3.28 7.50 2.59 2.42 1.67 2 20 7.80 13.31 3.60 4.21 1.64 3 30 9.25 16.69 4.01 4.10 1.60 4 40 11.27 20.92 4.30 4.69 1.56 5 50 11.01 19.61 4.59 4.72 1.60 6 60 13.04 20.59 4.62 4.92 1.62 -
土壤有机碳的测定采用重铬酸钾氧化-外加热法,土壤全氮的测定采用凯氏定氮法,土壤全磷的测定采用酸溶-钼锑抗比色法[16]。根际效应采用根际土和非根际土各相应养分指标的比值表示,>1为根际正效应,<1为根际负效应。
-
利用SPSS 22.0分析数据,其中,林龄、根际对土壤碳、氮、磷质量分数及生态化学计量比的影响采用双因素方差分析;数据显著性检验采用Tukey法;相关系数和决定系数分别通过Pearson相关分析和线性回归分析确定;用Excel 2017作图表。
-
由表2可知:根际与非根际土壤有机碳质量分数范围分别为6.67~12.93和5.96~11.16 g·kg−1,且随樟子松林龄的增加而增大,在营林时间为60 a时达到峰值。根际全氮质量分数范围为0.23~0.41 g·kg−1,随樟子松林龄的增加先逐渐增高,40 a近熟林时达到峰值,后呈下降趋势。非根际土壤全氮质量分数范围为0.11~0.36 g·kg−1,总体随林龄增加持续增大,其峰值出现在60 a过熟林。根际与非根际土壤全磷质量分数范围分别为0.19~0.25和0.17~0.20 g·kg−1,随樟子松林龄的增加呈现出先增加后降低的变化趋势,在40 a近熟林时达到峰值。本研究区不同林龄樟子松根际土壤碳、氮、磷质量分数均高于非根际土壤。其中,各林龄的根际土壤有机碳质量分数均显著高于非根际土壤(P<0.05);根际土壤全氮质量分数在20~50 a时显著高于非根际土壤(P<0.05),其余林龄根际土壤全氮质量分数与非根际土壤差异并不显著(P>0.05);根际土壤全磷质量分数在10~50 a时与非根际土壤差异显著(P<0.05)。由双因素方差分析可知(表3):林龄、根际以及两者之间的交互作用,均对土壤碳、氮、磷质量分数有极显著影响(P<0.01)。
表 2 樟子松根际与非根际土壤碳、氮、磷质量分数及其生态化学计量比
Table 2. Content of C, N, P in rhizosphere and non-rhizosphere soil of P. sylvestris var. mongolica and its ecological stoichiometric ratio
林龄/a 土壤类型 有机碳/(g·kg−1) 全氮/(g·kg−1) 全磷/(g·kg−1) C∶N C∶P N∶P 10 根际 6.67±0.28 Ad 0.23±0.02 Ac 0.19±0.01 Ae 29.13±2.19 Bb 35.07±0.08 Ad 1.23±0.10 Ad 非根际 5.96±0.08 Be 0.11±0.09 Ad 0.17±0.01 Be 59.48±22.34 Aa 35.63±0.09 Ae 0.65±0.53 Bd 20 根际 7.16±0.26 Ac 0.26±0.01 Ac 0.22±0.01 Ac 27.55±0.60 Bbc 33.09±0.14 Ae 1.20±0.03 Ad 非根际 5.96±0.33 Be 0.17±0.01 Bcd 0.19±0.01 Bd 35.06±0.13 Ab 32.12±1.27 Af 0.92±0.04 Bcd 30 根际 9.89±0.04 Ab 0.36±0.03 Ab 0.24±0.01 Aa 27.59±2.19 Bbc 41.21±0.14 Bc 1.50±0.11 Ac 非根际 8.85±0.29 Bc 0.26±0.01 Bb 0.20±0.01 Bab 34.04±0.20 Ab 44.24±1.04 Ac 1.30±0.04 Bbc 40 根际 10.17±0.12 Ab 0.41±0.01 Aa 0.25±0.02 Aa 24.81±0.56 Bc 41.13±0.22 Ac 1.66±0.03 Ab 非根际 8.31±0.16 Bd 0.24±0.01 Bbc 0.20±0.01 Ba 34.65±0.78 Ab 41.18±1.16 Ad 1.19±0.06 Bbc 50 根际 12.79±0.22 Aa 0.36±0.02 Ab 0.23±0.01 Ab 35.60±1.92 Aa 56.73±0.37 Ab 1.60±0.08 Abc 非根际 10.73±0.24 Bb 0.30±0.02 Bab 0.20±0.01 Bb 35.84±1.59 Ab 54.64±0.72 Bb 1.53±0.09 Aab 60 根际 12.93±0.33 Aa 0.38±0.01 Aab 0.20±0.0 1 Ad 34.04±0.81 Aa 65.26±1.03 Aa 1.92±0.02 Aa 非根际 11.16±0.15 Ba 0.36±0.03 Aa 0.19±0.01 Ac 31.12±2.18 Ab 58.46±0.26 Ba 1.88±0.12 Aa 说明:不同大写字母表示同一林龄在不同土壤类型间差异显著(P<0.05),不同小写字母表示同一土壤类型在不同林龄间差异显著 (P<0.05) 表 3 土壤碳、氮、磷质量分数及其生态化学计量比方差分析
Table 3. Soil C, N, P content and its ecological stoichiometric ratio variance analysis
指标 F 有机碳 全氮 全磷 C∶N C∶P N∶P 林龄(A) 1 310.94** 81.92** 299.20** 4.05** 1 603.44** 76.69** 根际(B) 680.28** 180.92** 1 220.84** 15.23** 23.10** 70.28** AB交互作用 15.53** 9.09** 46.38** 4.71** 32.26** 7.39** 说明:*表示影响显著(P<0.05),**表示影响极显著(P<0.01) 樟子松人工林根际土壤C∶N、C∶P,非根际土壤C∶P均随林龄的增加先降后升,非根际土壤C∶N随林龄增加呈先降后升再降低的变化趋势。根际与非根际土壤N∶P随林龄的增加持续上升。樟子松人工林根际与非根际土壤碳、氮、磷生态化学计量比之间的大小关系在不同林龄略有差异。根际与非根际土壤C∶N分别为24.81~35.60、31.12~59.48,根际与非根际土壤C∶P分别为33.09~65.26、32.12~58.46。在营林时间为10~40 a时,非根际土壤C∶N、C∶P均高于根际土壤,其中非根际土壤C∶N与根际土壤存在显著差异(P<0.05)。50~60 a时,根际土壤C∶N、C∶P均高于非根际,但不存在显著差异(P>0.05)。根际与非根际土壤N∶P分别为1.20~1.92、0.65~1.88,根际土壤N∶P始终高于非根际,且在10~40 a时差异显著(P<0.05)。
-
如图1所示:土壤有机碳、全氮、全磷根际效应值均大于1,呈正效应,其中根际效应值最大的是土壤全氮。樟子松人工林有机碳、全磷根际效应值随林龄的增加整体呈先升后降趋势,均在40 a近熟林时达到峰值。全氮根际效应值随林龄增加呈现出先降后升再降低的变化趋势,在40 a近熟林时最大,在60 a过熟林时降低到最小值。总体上看,土壤有机碳、全氮、全磷在40 a近熟林时根际效应最高,60 a过熟林时根际效应最低,土壤全氮受根际效应的影响最大。
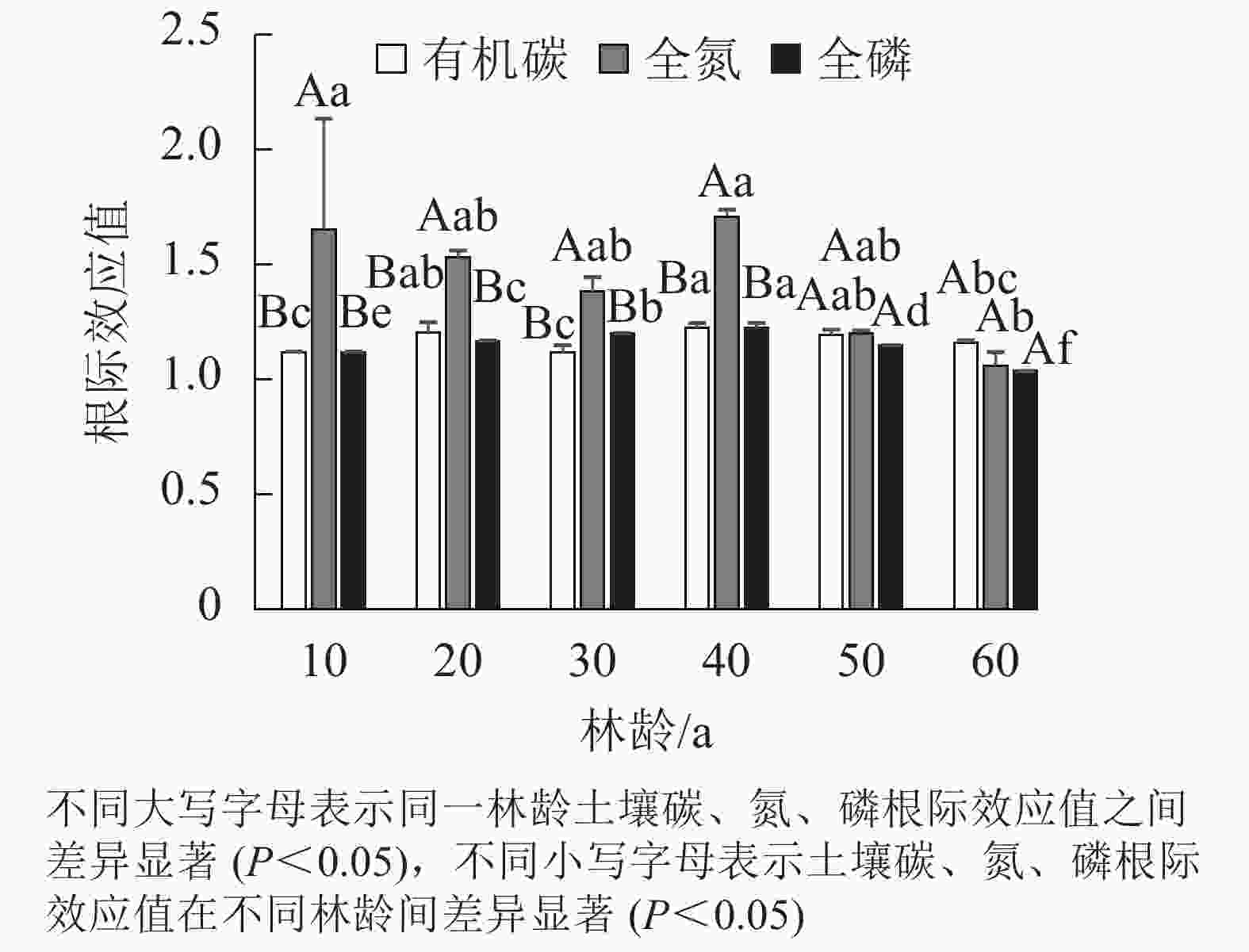
图 1 不同林龄樟子松土壤碳、氮、磷根际效应值
Figure 1. Soil C, N, P rhizosphere effect values of P. sylvestris var. mongolica with different stand ages
-
如表4所示:土壤有机碳对土壤C∶N、土壤全磷对土壤C∶ P的决定系数(R2)均低于0.08,土壤有机碳、全氮分别只能决定土壤C∶N、C∶P变化的7% (R2=0.07)、1%(R2=0.01),解释程度较低,表征效果较差。除土壤有机碳与土壤C∶N、土壤全磷与土壤C∶P之间的相关性不显著外(P>0.05),土壤有机碳、全氮和全磷之间均存在极显著的相关性(P<0.01)。从不同土壤类型的角度分析,由表5可知:根际土壤中,C∶P与C∶N、N∶P相关性较高(P<0.01),C∶N与N∶P之间无显著相关性(P>0.05)。非根际土壤的N∶P分别与C∶N、C∶P相关性极显著(P<0.01),C∶N与C∶P之间相关性不显著(P>0.05)。
表 4 土壤碳、氮、磷质量分数对生态化学计量比的解释程度及相关关系
Table 4. Soil C, N, P content and its ecological stoichiometric ratio correlation
项目 R2 有机碳 全氮 全磷 C∶N 0.07 (0.13) 0.45 (<0.01) C∶P 0.84 (<0.01) 0.01 (0.71) N∶P 0.99 (<0.01) 0.21 (<0.01) 说明:R2为决定系数,值越大解释程度越高。括号内为P值, P<0.05表示显著相关,P<0.01表示极显著相关 表 5 根际生态化学计量比间相关关系和非根际生态化学计量比间相关关系
Table 5. Correlation between rhizosphere ecological stoichiometric ratio and non-rhizosphere ecological stoichiometric ratio
项目 根际土壤 非根际土壤 C∶N C∶P N∶P C∶N C∶P N∶P C∶N 1 1 C∶P 0.77** 1 −0.38 1 N∶P 0.32 0.85** 1 −0.78** 0.85** 1 说明:*表示显著相关(P<0.05),**表示极显著相关(P<0.01) -
土壤碳、氮、磷是植物生长的必需元素。本研究中,各林龄樟子松人工林根际与非根际土壤碳、氮、磷质量分数的平均值(9.22、0.29、0.20 g·kg−1)均远低于全国平均水平(11.12、1.06、0.65 g·kg−1)[17],根据全国第2次土壤普查养分分组标准,研究区土壤碳、氮、磷质量分数分别为4级(缺乏)、6级(极缺乏)和5级(很缺乏)状态,可见该地区土壤碳、氮、磷极为贫瘠。林龄可以改变植物的生理生态特征、林分结构和生产力[18]。本研究中,林龄、根际以及二者之间的交互作用,对土壤碳、氮、磷质量分数均具有显著影响。从樟子松林龄的角度来看,根际与非根际土壤有机碳质量分数均随林龄的增加不断提高,造成此现象的原因是随着林龄的增加,林木枯枝落叶层逐渐增厚,微生物分解转化的有机碳不断增多,形成碳积累[19]。土壤全氮、全磷总体上随林龄先增大,40 a后有所降低。其原因在于樟子松人工林在10~30 a时处于幼龄林和中龄林,其生长发育对土壤氮、磷消耗较多,而枯枝落叶层积累较少,对土壤养分的返还相对不足,因此,土壤氮、磷质量分数相对较低。40 a樟子松趋近成熟期,其生长发育趋于稳定,对土壤氮、磷消耗降低,而此时枯枝落叶层的增厚增加了土壤养分的返还量,因而氮、磷质量分数最高。50 a成熟林后,樟子松人工林土壤相对酸化[20],土壤真菌病害增多,土壤微生物活性降低,导致枯落物转化为土壤氮、磷养分的效率也随之降低。同时,土壤积累的氮、磷养分被樟子松持续消耗,土壤氮、磷质量分数开始降低。
从根际与非根际的角度进行分析,不同林龄樟子松根际土壤有机碳、全氮、全磷质量分数均高于非根际,表现为根际正效应,其原因在于根系环境有利于土壤有机碳、全氮、全磷的积累。根系分泌物、脱落的根毛和根表皮细胞是影响根际有机碳的重要因素[21],其在微生物共同作用下形成的养分循环机制,为根际土壤提供了丰富的碳源,是导致根际土壤有机碳高于非根际土壤的决定性因素。天然林地土壤氮主要来自植物残体的返还[22]。有研究表明:樟子松的根际可以为微生物提供营养基质,增强微生物的活性[23],微生物通过对根系残茬和枯枝落叶层的分解,提升根际土壤的氮、磷质量分数,对氮、磷富集程度较高,表现为根际正效应。土壤有机碳、全氮、全磷根际效应均在40 a时达到峰值,其原因在于40 a的樟子松人工林趋近成熟,其养分利用率相对较低,而枯枝落叶层对养分的返还较高,细根周转和微生物代谢致使大量有机物在根际土壤富集,根际效应值持续升高。60 a过熟林时,樟子松根系出现衰退,不能为微生物提供足够营养基质,微生物活性的降低导致分解效率下降,根际效应值近一步降低。
本研究中,林龄、根际以及二者之间的交互作用,对土壤碳、氮、磷生态化学计量比均具有显著影响。土壤C∶N被认为是反映土壤氮素矿化能力的标志,可在一定程度上指示凋落物与根系残茬对土壤碳、氮的积累程度[24]。本研究中各林龄根际与非根际土壤的C∶N平均值均远高于全国(11.90)和全球(13.33)[25]。决定系数可知:土壤有机碳对土壤C∶N解释率(R2=0.07)低于土壤全氮(R2=0.45),土壤C∶N主要受到土壤全氮的影响,因此,可以推断樟子松人工林各林龄根际与非根际土壤均受到氮限制。其中,60 a过熟林的根际与非根际土壤C∶N最大,意味着该阶段受氮限制更严重。土壤C∶P不但对土壤磷的有效性具有指示作用[26],还可以衡量在微生物作用下土壤有机质释放或固持磷的潜力[27]。本研究除60 a过熟林根际土壤外,各林龄土壤C∶P均低于全国平均水平(61.00)[25],60 a过熟林根际土壤C∶P高于全国平均水平。然而决定系数结果表明:土壤全磷对C∶P的解释率较低(R2=0.01),表征效果较差,因此无法表明60 a樟子松人工林根际土壤受到磷限制,需要进一步的研究加以证实。土壤N∶P虽然不能较好地反映生态系统限制水平,但可间接作为养分限制和供给水平的有效预测指标[28]。各林龄根际与非根际土壤N∶P均远低于全国水平(5.20)[25],同时,土壤全氮对土壤N∶P的解释率(R2=0.99)要远高于全磷(R2=0.21),表明相对于磷,氮是更重要的限制因子,也再次证实樟子松整个生长过程中始终受氮的限制。与此同时,从根际、非根际生态化学计量比相关性分析结果来看,根际土壤C∶N与C∶P相关关系极显著,而非根际土壤C∶N与C∶P相关关系不显著,土壤生态化学计量比可以反映土壤养分元素的限制性,因而本结果表明:相较于非根际土壤,根际土壤氮、磷限制具有协同性,二者中任一养分限制易引起另一限制的发生。
-
辽西北沙地樟子松人工林土壤养分贫瘠,根际土壤有机碳、全氮、全磷质量分数均高于非根际土壤,根系对养分的富集与平衡性维持作用明显。林龄、根际以及二者之间的交互作用对土壤碳、氮、磷质量分数影响显著。从生态化学计量学角度分析,各林龄樟子松人工林根际与非根际土壤均表现为氮限制,其中,60 a过熟林时氮限制更为强烈。相较于非根际土壤,根际土壤氮、磷限制具有协同性。林龄、根际以及二者之间的交互作用对土壤碳、氮、磷生态化学计量比具有显著影响。由土壤碳、氮、磷质量分数及其生态化学计量比之间的决定系数可知:樟子松人工林土壤C∶N主要受到土壤全氮的影响,土壤全磷对土壤C∶P的表征效果较差,土壤C∶P主要受土壤有机碳的影响,土壤N∶P受土壤全氮影响大于全磷。从土壤养分质量分数和生态化学计量比综合考虑,建议对辽西北沙地樟子松人工林合理施用氮肥、引入固氮植物以解除氮限制,并针对根际土壤氮、磷限制具有协同性的特点适时补充磷肥。
Ecological stoichiometric characteristics of rhizosphere and non-rhizosphere soil of Pinus sylvestris var. mongolica plantations at different ages
-
摘要:
目的 探究辽西北沙地不同林龄樟子松Pinus sylvestris var. mongolica人工林根际与非根际土壤碳(C)、氮(N)、磷(P)质量分数及生态化学计量特征关系,为该地区的樟子松林培育、经营及管理提供理论依据。 方法 采用时空互代的方法,在辽西北章古台地区选取6个林龄(10、20、30、40、50和60 a)的樟子松人工林作为研究对象,分析各林龄下根际与非根际土壤碳、氮、磷质量分数及化学计量比的差异和影响因素。 结果 辽西北沙地樟子松人工林土壤贫瘠,根际土壤碳、氮、磷质量分数均高于非根际土壤,根系对养分的富集与平衡维持作用明显。林龄、根际以及二者之间的交互作用,对土壤碳、氮、磷质量分数及其生态化学计量比影响显著。樟子松人工林土壤C∶N主要受到土壤全氮的影响,土壤C∶P主要受土壤有机碳的影响,土壤N∶P受土壤全氮的影响大于全磷。各林龄樟子松人工林土壤C∶N均远高于全国平均水平,表现为氮限制,其中60 a过熟林氮限制更为强烈。樟子松人工林根际土壤氮、磷限制存在一定程度的协同性。 结论 各林龄樟子松生长均受到氮限制,相较于根际土壤,非根际土壤氮更为缺乏。在森林经营过程中,应充分考虑根际与非根际土壤的差异性,建议对辽西北沙地樟子松人工林施用氮肥、引入固氮植物以解除氮限制,并注意根系磷肥的补充。图1表5参28 Abstract:Objective The purpose is to explore the relationship between C, N, and P contents in rhizosphere and non-rhizosphere soil and ecological stoichiometric characteristics of Pinus sylvestris var. mongolica plantations at different ages in sandy land of northwest Liaoning Province, so as to provide theoretical basis for cultivation and management of P. sylvestris var. mongolica plantations in this area. Method Using space-temporal exchange method, six P. sylvestris var. mongolica plantations (10, 20, 30, 40, 50 and 60 a) were selected in Zhanggutai area to analyze the differences and influencing factors of C, N, P contents and stoichiometric ratio between rhizosphere and non-rhizosphere soil of different stand ages. Result The soil of P. sylvestris var. mongolica plantation in the sandy land of northwest Liaoning was poor, and the contents of C, N, and P in rhizosphere soil were higher than those in non-rhizosphere soil. The root system had a significant effect on nutrient enrichment and balance maintenance. Stand age, rhizosphere, and their interaction had significant effects on soil C, N, P contents and their ecological stoichiometry. Soil C∶N of the P. sylvestris var. mongolica plantation was mainly affected by total N, soil C∶P was mainly affected by soil organic carbon, and soil N∶P was more affected by total N than total P. The soil C∶N ratio of P. sylvestris var. mongolica plantation at all ages was much higher than that of the national average level, which was manifested as N limitation, especially in the 60 a over mature forest. The limitation of N and P in rhizosphere soil of P. sylvestris var. mongolica plantation was synergistic to some extent. Conclusion The growth of P. sylvestris var. mongolica at all ages is restricted by N, and non-rhizosphere soil is more deficient in N compared with rhizosphere soil. In forest management, the difference between rhizosphere and non-rhizosphere soil should be fully considered. It is suggested that N fertilizer and N-fixing plants should be applied and introduced to P. sylvestris var. mongolica plantations in sandy land of northwest Liaoning to relieve N limitation, and P fertilizer should be added to the root system. [Ch, 1 fig. 5 tab. 28 ref.] -
图 1 不同林龄樟子松土壤碳、氮、磷根际效应值
Figure 1 Soil C, N, P rhizosphere effect values of P. sylvestris var. mongolica with different stand ages
表 1 样地基本情况
Table 1. Basic information of the sample plots
样地
编号林分
年龄/a平均树
高/m平均胸
径/cm东西平均
冠幅/m南北平均
冠幅/m土壤容重/
(g·cm−3)1 10 3.28 7.50 2.59 2.42 1.67 2 20 7.80 13.31 3.60 4.21 1.64 3 30 9.25 16.69 4.01 4.10 1.60 4 40 11.27 20.92 4.30 4.69 1.56 5 50 11.01 19.61 4.59 4.72 1.60 6 60 13.04 20.59 4.62 4.92 1.62  下载: 导出CSV
下载: 导出CSV
表 2 樟子松根际与非根际土壤碳、氮、磷质量分数及其生态化学计量比
Table 2. Content of C, N, P in rhizosphere and non-rhizosphere soil of P. sylvestris var. mongolica and its ecological stoichiometric ratio
林龄/a 土壤类型 有机碳/(g·kg−1) 全氮/(g·kg−1) 全磷/(g·kg−1) C∶N C∶P N∶P 10 根际 6.67±0.28 Ad 0.23±0.02 Ac 0.19±0.01 Ae 29.13±2.19 Bb 35.07±0.08 Ad 1.23±0.10 Ad 非根际 5.96±0.08 Be 0.11±0.09 Ad 0.17±0.01 Be 59.48±22.34 Aa 35.63±0.09 Ae 0.65±0.53 Bd 20 根际 7.16±0.26 Ac 0.26±0.01 Ac 0.22±0.01 Ac 27.55±0.60 Bbc 33.09±0.14 Ae 1.20±0.03 Ad 非根际 5.96±0.33 Be 0.17±0.01 Bcd 0.19±0.01 Bd 35.06±0.13 Ab 32.12±1.27 Af 0.92±0.04 Bcd 30 根际 9.89±0.04 Ab 0.36±0.03 Ab 0.24±0.01 Aa 27.59±2.19 Bbc 41.21±0.14 Bc 1.50±0.11 Ac 非根际 8.85±0.29 Bc 0.26±0.01 Bb 0.20±0.01 Bab 34.04±0.20 Ab 44.24±1.04 Ac 1.30±0.04 Bbc 40 根际 10.17±0.12 Ab 0.41±0.01 Aa 0.25±0.02 Aa 24.81±0.56 Bc 41.13±0.22 Ac 1.66±0.03 Ab 非根际 8.31±0.16 Bd 0.24±0.01 Bbc 0.20±0.01 Ba 34.65±0.78 Ab 41.18±1.16 Ad 1.19±0.06 Bbc 50 根际 12.79±0.22 Aa 0.36±0.02 Ab 0.23±0.01 Ab 35.60±1.92 Aa 56.73±0.37 Ab 1.60±0.08 Abc 非根际 10.73±0.24 Bb 0.30±0.02 Bab 0.20±0.01 Bb 35.84±1.59 Ab 54.64±0.72 Bb 1.53±0.09 Aab 60 根际 12.93±0.33 Aa 0.38±0.01 Aab 0.20±0.0 1 Ad 34.04±0.81 Aa 65.26±1.03 Aa 1.92±0.02 Aa 非根际 11.16±0.15 Ba 0.36±0.03 Aa 0.19±0.01 Ac 31.12±2.18 Ab 58.46±0.26 Ba 1.88±0.12 Aa 说明:不同大写字母表示同一林龄在不同土壤类型间差异显著(P<0.05),不同小写字母表示同一土壤类型在不同林龄间差异显著 (P<0.05)
下载: 导出CSV
表 3 土壤碳、氮、磷质量分数及其生态化学计量比方差分析
Table 3. Soil C, N, P content and its ecological stoichiometric ratio variance analysis
指标 F 有机碳 全氮 全磷 C∶N C∶P N∶P 林龄(A) 1 310.94** 81.92** 299.20** 4.05** 1 603.44** 76.69** 根际(B) 680.28** 180.92** 1 220.84** 15.23** 23.10** 70.28** AB交互作用 15.53** 9.09** 46.38** 4.71** 32.26** 7.39** 说明:*表示影响显著(P<0.05),**表示影响极显著(P<0.01)
下载: 导出CSV
表 4 土壤碳、氮、磷质量分数对生态化学计量比的解释程度及相关关系
Table 4. Soil C, N, P content and its ecological stoichiometric ratio correlation
项目 R2 有机碳 全氮 全磷 C∶N 0.07 (0.13) 0.45 (<0.01) C∶P 0.84 (<0.01) 0.01 (0.71) N∶P 0.99 (<0.01) 0.21 (<0.01) 说明:R2为决定系数,值越大解释程度越高。括号内为P值, P<0.05表示显著相关,P<0.01表示极显著相关
下载: 导出CSV
表 5 根际生态化学计量比间相关关系和非根际生态化学计量比间相关关系
Table 5. Correlation between rhizosphere ecological stoichiometric ratio and non-rhizosphere ecological stoichiometric ratio
项目 根际土壤 非根际土壤 C∶N C∶P N∶P C∶N C∶P N∶P C∶N 1 1 C∶P 0.77** 1 −0.38 1 N∶P 0.32 0.85** 1 −0.78** 0.85** 1 说明:*表示显著相关(P<0.05),**表示极显著相关(P<0.01)
下载: 导出CSV
-
[1] 朱教君, 康宏樟, 许美玲. 科尔沁沙地南缘樟子松(Pinus sylvestris var. mongolica)人工林天然更新障碍[J]. 生态学报, 2007, 27(10): 4086 − 4095. ZHU Jiaojun, KANG Hongzhang, XU Meiling. Natural regeneration barriers of Pinus sylvestris var. mongolica plantations in southern Keerqin Sandy Land, China [J]. Acta Ecol Sin, 2007, 27(10): 4086 − 4095. [2] 吴祥云, 姜凤岐, 李晓丹, 等. 樟子松人工固沙林衰退的规律和原因[J]. 应用生态学报, 2004, 15(12): 2225 − 2228. WU Xiangyun, JIANG Fengqi, LI Xiaodan, et al. Decline regularity and causes of Pinus sylvestris var. mongolica plantation on sandy land [J]. Chin J Appl Ecol, 2004, 15(12): 2225 − 2228. [3] 宋立宁, 朱教君, 郑晓. 基于沙地樟子松人工林衰退机制的营林方案[J]. 生态学杂志, 2017, 36(11): 3249 − 3256. SONG Lining, ZHU Jiaojun, ZHENG Xiao. Forestation and management scheme of Pinus sylvestris var. mongolica plantations in sandy lands based on their decline mechanisms [J]. Chin J Ecol, 2017, 36(11): 3249 − 3256. [4] HESSEN D. Stoichiometry in food webs: Lotka revisted [J]. Oikos, 1997, 7(9): 95 − 200. [5] 陈军强, 张蕊, 侯尧宸, 等. 亚高山草甸植物群落物种多样性与群落C、N、P生态化学计量的关系[J]. 植物生态学报, 2013, 37(11): 979 − 987. CHEN Junqiang, ZHANG Rui, HOU Yaochen, et al. Relationships between species diversity and C, N and P ecological stoichiometry in plant communities of sub-alpine meadow [J]. Chin J Plant Ecol, 2013, 37(11): 979 − 987. [6] 梅杰, 周国英. 不同林龄马尾松林根际与非根际土壤微生物、酶活性及养分特征[J]. 中南林业科技大学学报, 2011, 31(4): 46 − 49. MEI Jie, ZHOU Guoying. Study of rhizosphere and non-rhizosphere microbial, enzyme activity and nutrients element content of soil in different stand ages Pinus massoniana forest [J]. J Cent South Univ For Technol, 2011, 31(4): 46 − 49. [7] FRASER T D, LYNCH D H, GAIERO J, et al. Quantification of bacterial non-specific acid (PhoC) and alkaline (PhoD) phosphatase genes in bulk and rhizosphere soil from organically managed soybean fields [J]. Appl Soil Ecol, 2017, 111: 48 − 56. [8] BALAKRISHNAN B, SAHU B K, RANISHREE J K, et al. Assessment of heavy metal concentrations and associated resistant bacterial communities in bulk and rhizosphere soil of Avicennia marina of Pichavaram mangrove, India [J]. Environ Earth Sci, 2017, 76(1): 58. doi: 10.1007/s12665-016-6378-7. [9] ANGST G, INGRID K, KIRFEL K, et al. Spatial distribution and chemical composition of soil organic matter fractions in rhizosphere and non-rhizosphere soil under European beech (Fagus sylvatica L.) [J]. Geoderma, 2016, 264: 179 − 187. [10] BIRD J A, HERMAN D J, FIRESTONE M K. Rhizosphere priming of soil organic matter by bacterial groups in a grassland soil [J]. Soil Biol Biochem, 2011, 43(4): 718 − 725. [11] 王凯, 沈潮, 宋立宁, 等. 持续干旱下沙地樟子松幼苗C、N、P化学计量变化规律[J]. 生态学杂志, 2020, 39(7): 2175 − 2184. WANG Kai, SHEN Chao, SONG Lining, et al. Variations in C, N and P stoichiometry of Pinus sylvestris var. mongolica seedlings under continuous drought [J]. Chin J Ecol, 2020, 39(7): 2175 − 2184. [12] 王凯, 赵成姣, 张日升, 等. 不同密度樟子松人工林土壤碳氮磷化学计量特征[J]. 生态学杂志, 2020, 39(3): 741 − 748. WANG Kai, ZHAO Chengjiao, ZHANG Risheng, et al. Soil carbon, nitrogen and phosphorus stoichiometry of Pinus sylvestris var. mongolica plantations with different densities [J]. Chin J Ecol, 2020, 39(3): 741 − 748. [13] 赵姗宇, 黎锦涛, 孙学凯, 等. 樟子松人工林原产地与不同自然降水梯度引种地土壤和植物叶片生态化学计量特征[J]. 生态学报, 2018, 38(20): 7189 − 7197. ZHAO Shanyu, LI Jintao, SUN Xuekai, et al. Responses of soil and plant stoichiometric characteristics along rainfall gradients in Mongolian pine plantations in native and introduced regions [J]. Acta Ecol Sin, 2018, 38(20): 7189 − 7197. [14] 淑敏, 姜涛, 王东丽, 等. 科尔沁沙地不同林龄樟子松人工林土壤生态化学计量特征[J]. 干旱区研究, 2018, 35(4): 789 − 795. SHU Min, JIANG Tao, WANG Dongli, et al. Soil Ecological stoichiometry under the planted of Pinus sylvestris var. mongolica forests with different stand ages in the Horqin Sandy Land [J]. Arid Zone Res, 2018, 35(4): 789 − 795. [15] PHILLIPS R P, FAHEY T J. The influence of soil fertility on rhizosphere effects in northern hardwood forest soils [J]. Soil Sci Soc Am J, 2008, 72(2): 453 − 461. [16] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2005. [17] 胡启武, 聂兰琴, 郑艳明, 等. 沙化程度和林龄对湿地松叶片及林下土壤C、N、P化学计量特征影响[J]. 生态学报, 2014, 34(9): 2246 − 2255. HU Qiwu, NIE Lanqin, ZHENG Yanming, et al. Effects of desertification intensity and stand age on leaf and soil carbon, nitrogen and phosphorus stoichiometry in Pinus elliottii plantation [J]. Acta Ecol Sin, 2014, 34(9): 2246 − 2255. [18] 李玉新, 赵忠, 陈金泉. 不同林龄人工沙棘林结构与林下物种多样性研究[J]. 西北植物学报, 2010, 30(4): 776 − 785. LI Yuxin, ZHAO Zhong, CHEN Jinquan. Seabuckthorn plantation structure at different ages and its understory species diversity [J]. Acta Bot Boreal-Occident Sin, 2010, 30(4): 776 − 785. [19] 马月婷, 张丽静, 杜明新, 等. 不同种植年限白沙蒿对根际土壤营养元素的影响[J]. 草业科学, 2014, 31(2): 224 − 231. MA Yueting, ZHANG Lijing, DU Mingxin, et al. Effects of different ages Artemisia sphaerocephala on the content of nutrient elements in rhizosphere soil [J]. Pratacult Sci, 2014, 31(2): 224 − 231. [20] 于德良, 雷泽勇, 赵国军, 等. 土壤酶活性对沙地樟子松人工林衰退的响应[J]. 环境化学, 2019, 38(1): 97 − 105. YU Deliang, LEI Zeyong, ZHAO Guojun. et al. Response of soil enzyme activity to the decline of Pinus sylvestris var. mongolica plantations on sand land [J]. Environ Chem, 2019, 38(1): 97 − 105. [21] 朱秋莲, 邢肖毅, 程曼, 等. 宁南山区典型植物根际与非根际土壤碳、氮形态[J]. 应用生态学报, 2013, 24(4): 983 − 988. ZHU Qiulian, XING Xiaoyi, CHENG Man, et al. Concentrations of different carbon and nitrogen fractions in rhizosphere and non-rhizosphere soils of typical plant species in mountainous area of southern Ningxia, Northwest China [J]. Chin J Appl Ecol, 2013, 24(4): 983 − 988. [22] 徐华山, 赵同谦, 贺玉晓, 等. 滨河湿地不同植被对农业非点源氮污染的控制效果[J]. 生态学报, 2010, 30(21): 5759 − 5768. XU Huashan, ZHAO Tongqian, HE Yuxiao, et al. Effect of different vegetation types on agricultural non-point nitrogen pollution in riparian wetlands [J]. Acta Ecol Sin, 2010, 30(21): 5759 − 5768. [23] 赵琼, 曾德慧, 于占源, 等. 沙地樟子松人工林土壤磷素转化的根际效应[J]. 应用生态学报, 2006, 17(8): 1377 − 1381. ZHAO Qiong, ZENG Dehui, YU Zhanyuan, et al. Rhizosphere effects of Pinus sylvestris var. mongolica on soil phosphorus transformation [J]. Chin J Appl Ecol, 2006, 17(8): 1377 − 1381. [24] 张良侠, 樊江文, 张文彦, 等. 京津风沙源治理工程对草地土壤有机碳库的影响: 以内蒙古锡林郭勒盟为例[J]. 应用生态学报, 2014, 25(2): 374 − 380. ZHANG Liangxia, FAN Jiangwen, ZHANG Wenyan, et al. Impact of the Beijing and Tianjin Sand Source Control Project on the grassland soil organic carbon storage: a case study of Xilingol League, Inner Mongolia, China [J]. Chin J Appl Ecol, 2014, 25(2): 374 − 380. [25] 程滨, 赵永军, 张文广, 等. 生态化学计量学研究进展[J]. 生态学报, 2010, 30(6): 1628 − 1637. CHENG Bin, ZHAO Yongjun, ZHANG Wenguang, et al. The research advances and prospect of ecological stoichiometry [J]. Acta Ecol Sin, 2010, 30(6): 1628 − 1637. [26] 朱仁欢, 李玮, 郑子成, 等. 退耕植茶地土壤碳氮磷生态化学计量学特征[J]. 浙江农林大学学报, 2016, 33(4): 612 − 619. ZHU Renhuan, LI Wei, ZHENG Zicheng, et al. Ecological stoichiometry of soil C, N, and P for returning farmland to tea plantations [J]. J Zhejiang A&F Univ, 2016, 33(4): 612 − 619. [27] 孙超. 基于生态化学计量学的草地退化研究[D]. 长春: 吉林大学, 2012. SUN Chao. Research on Grassland Degradation based on Ecological Stoichiometry[D]. Changchun: Jilin University, 2012. [28] 曹娟, 闫文德, 项文化, 等. 湖南会同3个林龄杉木人工林土壤碳、氮、磷化学计量特征[J]. 林业科学, 2015, 51(7): 1 − 8. CAO Juan, YAN Wende, XIANG Wenhua, et al. Stoichiometry characterization of soil C, N and P of Chinese fir plantations at three different ages in Huitong, Hunan Province, China [J]. Sci Silv Sin, 2015, 51(7): 1 − 8. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210211
 点击查看大图
点击查看大图

计量
- 文章访问数: 1672
- HTML全文浏览量: 532
- PDF下载量: 45
- 被引次数: 0
