





-
1980−2010年间,中国陆地区域氮沉降平均增加了7.9 kg·hm−2,是世界上最大的活性氮排放国之一[1-2]。在森林生态系统中,氮沉降一部分被冠层截留,另一部分进入土壤[3],直接或间接影响地上地下生物地球化学循环和生物多样性等[4]。土壤氮素有效性对植物生长至关重要,植物能够利用氮素进行代谢,但植物吸收的大部分土壤氮则是经矿化作用形成的无机氮,只能吸收小部分有机氮[5]。氮矿化是土壤氮循环的关键过程,通过调控土壤养分供给等物质循环和能量流动影响生态系统生产力和功能[6-7]。因此,研究氮沉降背景下森林生态系统无机氮动态对森林管理与经营具有重要意义。研究认为:氮输入对氮素转化速率的影响包括促进[8]、抑制[9]和无影响[10]。中国陆地大多数生态系统处于氮限制状态,受碳氮耦合作用影响,少量氮输入能够增加陆地生态系统碳汇[11-12],对大气二氧化碳(CO2)升高等全球变化形成负反馈。随大气氮沉降持续增加,过量氮输入使得土壤氮饱和或过饱和[13],导致土壤酸化[14],并引发一系列生态后果[15]。土壤氮素转化受到微生物特征、气候因子与土壤理化性质等生物与非生物因素共同调控[7, 16],但不同生态系统氮素转化速率的驱动因素具有差异性[17]。研究表明:杉木Cunninghamia lanceolata对氮沉降响应较为敏感[18],但由于其生长快、经济价值高,成为中国南方亚热带地区重要的造林树种之一[19-20]。本研究以福建省沙县官庄国有林场的杉木人工林为研究对象,设置氮添加梯度进行氮沉降模拟试验,并利用野外原位培养与实验室分析相结合,旨在探究土壤无机氮与氮素转化对不同氮添加和季节变化的响应,为研究区杉木人工林土壤健康管理与可持续经营提供科学指导。
-
研究区福建省沙县官庄国有林场(26°30′N,117°43′E)地处亚热带季风气候区,平均海拔约200 m,无霜期约271 d。该地区雨热同期,夏季气温高,且降雨量大。年平均气温约19.6 ℃,最热月为7月(28.8 ℃),最冷月为1月(10.5 ℃)。年平均降水量约1 660.0 mm,降水量最大月为6月(230.2 mm),最小月为12月(31.6 mm);各季节土壤含水量为23.6%~27.8%。研究区杉木人工林土壤为红壤,林下植被稀疏,主要植物有五节芒Miscanthus floridulus、乌毛蕨Blechnum orientale、芒萁Dicranopteris pedata和菝葜Smilax china等。
-
参照文献[13]设置12个林分和土壤立地条件相似的20 m×20 m样方,为减少不同样方施肥处理的相互干扰[8],在每个样方内划定15 m×15 m的核心区和外围缓冲区。2003年末进行的样方本底调查发现:研究区杉木林龄为12 a,密度约1 700株·hm−2,平均树高约12.0 m,平均胸径约16.1 cm。土壤pH约4.68,全氮质量分数为1.21 g·kg−1,硝态氮、铵态氮、有效磷质量分数分别为14.86、22.09和5.22 mg·kg−1[21]。2004年初,依据氮沉降背景值,选择尿素为氮源模拟氮沉降;设置4个梯度的氮添加处理:对照(N0,0 g·m−2·a−1)、低氮(N1,6 g·m−2·a−1)、中氮(N2,12 g·m−2·a−1)和高氮(N3,24 g·m−2·a−1),每处理3个重复;按处理要求将尿素溶于20 L水,于每月初用喷雾器均匀喷洒在各样方中。
利用PVC管盖法原位研究土壤氮矿化速率[22],该方法切断了植物根系并阻断水气交换,可能会对微生物活动产生影响,但有研究[23]证明对原位氮矿化影响相对较小,因此可用于原位估计森林土壤氮矿化。分别于2013年6月、9月、12月和2014年3月,在每个样方内选3个土壤性质基本相同的点,将3个长为15.0 cm、直径为3.5 cm的PVC管打入各土壤表层10 cm,标记为P0(上下管口镂空都不做处理,打入后即取土)、P1(上管口用保鲜膜封住,培养3个月后取土)和P2(上下管口镂空不做处理,培养3个月后取土)。将取回的土样带回实验室过筛、混匀后,测定并分析土壤pH、有机质、铵态氮、硝态氮等理化性质。
-
于2013年6月起按季度进行为期1 a的土壤pH、有机质、铵态氮、硝态氮等指标测定。理化性质测定方法参考文献[23]和[24],氮素转化速率计算方法参考文献[25]。公式为:
$$ A = B + C{\text{;}}\;\;\;\;\;\;\;\;\;\;\;\; $$ (1) $$ {R_{\rm{m}}} = ({A_1} - {A_0})/3{\text{;}} $$ (2) $$ {R_{\rm{n}}} = ({B_1} - {B_0})/3{\text{;}}\;\; $$ (3) $$ {R_{\rm{l}}} = ({A_1} - {A_2})/3{\text{。}}\;\; $$ (4) 式(1)~(4)中:A为无机氮质量分数,B为硝态氮质量分数,C为铵态氮质量分数;A1为P1土样的无机氮质量分数,A0为P0土样的无机氮质量分数,B1为P1土样的硝态氮质量分数,B0为P0土样的硝态氮质量分数,A2为P2土样中的无机氮质量分数;Rm、Rn分别为净氮矿化速率、硝化速率,Rl为淋溶速率。质量分数单位为mg·kg−1,速率单位为mg·kg−1·月−1。
-
采用双因素方差分析法(two-way ANOVAs)检验季节和氮添加对土壤无机氮库质量分数和氮素转化速率的影响,用最小差异显著法(LSD)检验处理间的显著性,用R 4.0.4进行数据分析,用Excel 2016绘图。
-
由表1可知:氮添加对土壤pH、有机质、全氮质量分数和碳氮比(C/N)影响显著,不同水平氮处理下,土壤pH、有机质与碳氮比显著降低(P<0.05),全氮质量分数显著提高(P<0.05)。具体表现为:在所有处理中,土壤均为酸性,土壤pH 3.85~4.28。有机质质量分数为25.32~30.57 g·kg−1,全氮质量分数为1.35~1.67 g·kg−1,碳氮比为8.9~13.3。此外,无机氮质量分数为39.41~76.17 mg·kg−1,其中铵态氮质量分数为27.14~44.51 mg·kg−1,高于硝态氮(12.27~31.67 mg·kg−1)。
表 1 不同氮添加处理土壤主要理化和养分性质
Table 1. Main soil physicochemical and nutrient properties under different N addition
处理 pH 有机质/(g·kg−1) 全氮/(g·kg−1) C/N 铵态氮/(mg·kg−1) N0 4.28±0.19 a 30.57±2.89 a 1.35±0.14 c 13.3±2.0 a 27.14±4.58 d N1 4.14±0.24 b 27.13±3.07 b 1.57±0.17 b 10.1±1.4 b 32.19±6.56 c N2 4.00±0.22 c 26.82±3.19 b 1.62±0.19 ab 9.7±1.7 bc 39.13±6.35 b N3 3.85±0.16 d 25.32±3.34 b 1.67±0.18 a 8.9±1.7 cd 44.51±6.25 a 处理 铵态氮比例/% 硝态氮/(mg·kg−1) 硝态氮比例/% 铵态氮/硝态氮 无机氮/(mg·kg−1) N0 68.9 12.27±2.51 d 31.1 2.2 39.41±4.79 d N1 63.0 18.67±3.76 c 37.0 1.7 50.86±7.55 c N2 63.0 23.51±5.49 b 37.0 1.7 62.64±6.78 b N3 58.4 31.67±5.12 a 41.6 1.4 76.17±8.26 a 说明:数值为平均值±标准误,同列不同字母表示差异显著(P<0.05),铵态氮比例为铵态氮占无机氮比例,硝态氮比例为硝态氮占 无机氮比例 -
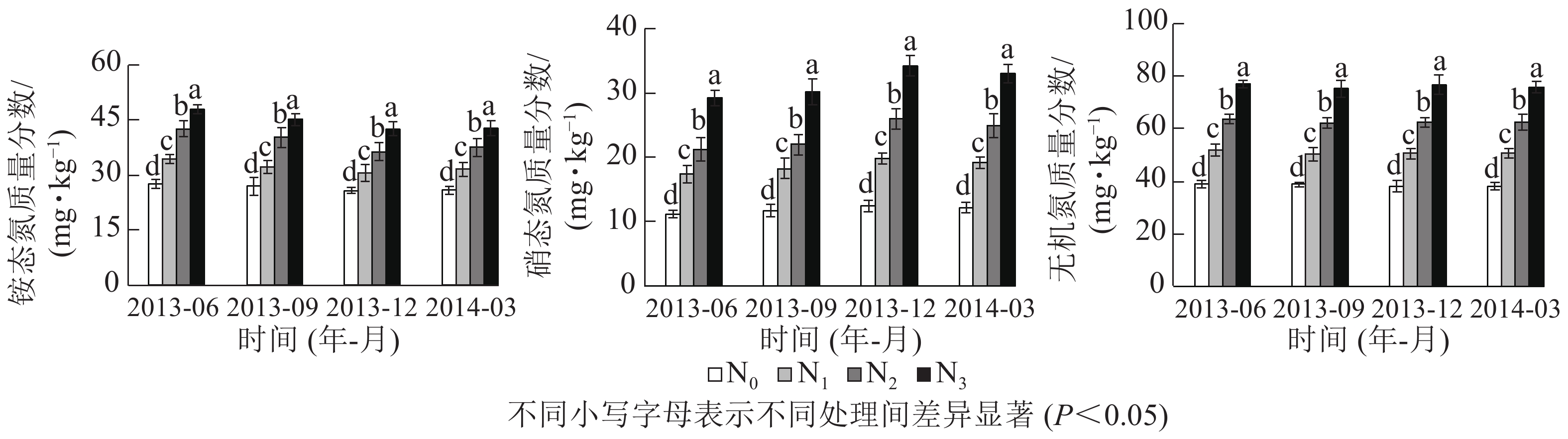
由表1可知:铵态氮、硝态氮与无机氮质量分数随氮添加梯度显著增加,从小到大依次为N0、N1、N2、N3。铵态氮占无机氮的比例(58.4%~68.9%)高于硝态氮(31.1%~41.6%),但铵态氮/硝态氮随氮添加梯度增加而逐渐降低。相对于N0,N1、N2和N3处理下无机氮年平均质量分数分别显著增加了29.1%、58.9%和93.3%,铵态氮为18.6%、44.1%和64.0%,硝态氮为52.2%、91.6%、158.1%,硝态氮质量分数增加幅度大于铵态氮。另外,季节对铵态氮和硝态氮质量分数影响显著,但对总无机氮质量分数影响不显著,且氮添加与季节无交互作用(P>0.05,表2)。在各季节间,夏秋2季铵态氮质量分数略高于冬春2季,而硝态氮则相反,无机氮质量分数在季节间变化不大(图1)。
表 2 氮添加和季节影响氮质量分数的双因素方差分析
Table 2. Effects of N addition, season, and their interactions on N mass fraction as indicated by two-way ANOVAs
处理 P 铵态氮 硝态氮 无机氮 氮添加 0.000* 0.000* 0.000* 季节 0.015* 0.005* 0.889 氮添加×季节 0.994 0.944 0.999 说明:*表示处理间差异显著 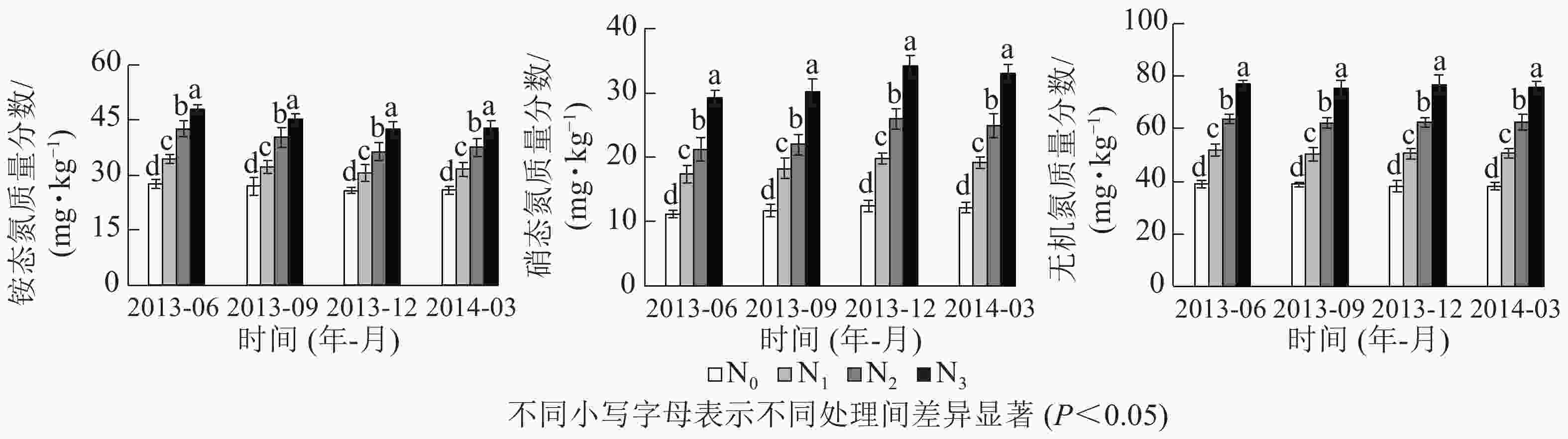
图 1 0~10 cm土层铵态氮、硝态氮和无机氮质量分数
Figure 1. Mass fractions of ammonium, nitrate and inorganic N in 0−10 cm soil layer
-
双因素方差分析和LSD多重比较表明:氮添加和季节处理对Rm、Rn和Rl的影响显著(P<0.05),且季节和处理对净氮矿化有交互影响(P<0.05,表3)。在氮添加方面,N3显著促进了氮矿化,各处理下土壤净氮矿化速率从大到小依次为N3、N2、N1、N0。硝化与淋溶速率对氮添加的响应趋势与净氮矿化具有相似性。此外,在季节方面,冬季净氮矿化为负值,表现为氮固定,而冬季的淋溶也为负。不同季节土壤氮素转化速率不同,但所有处理下均表现为夏季高、冬季低,秋季逐渐降低和春季逐渐回升的规律。不同季节氮素转化速率从大到小依次为夏季、秋季、春季、冬季(图2)。
表 3 氮添加和季节影响氮素转化速率的双因素方差分析
Table 3. Effects of N addition, season, and their interactions on N transformation rates as indicated by two-way ANOVAs
处理 P 矿化速率 硝化速率 淋溶速率 氮添加 0.000* 0.001* 0.042* 季节 0.000* 0.000* 0.000* 氮水平×季节 0.012* 0.693 0.955 说明:*表示处理间差异显著 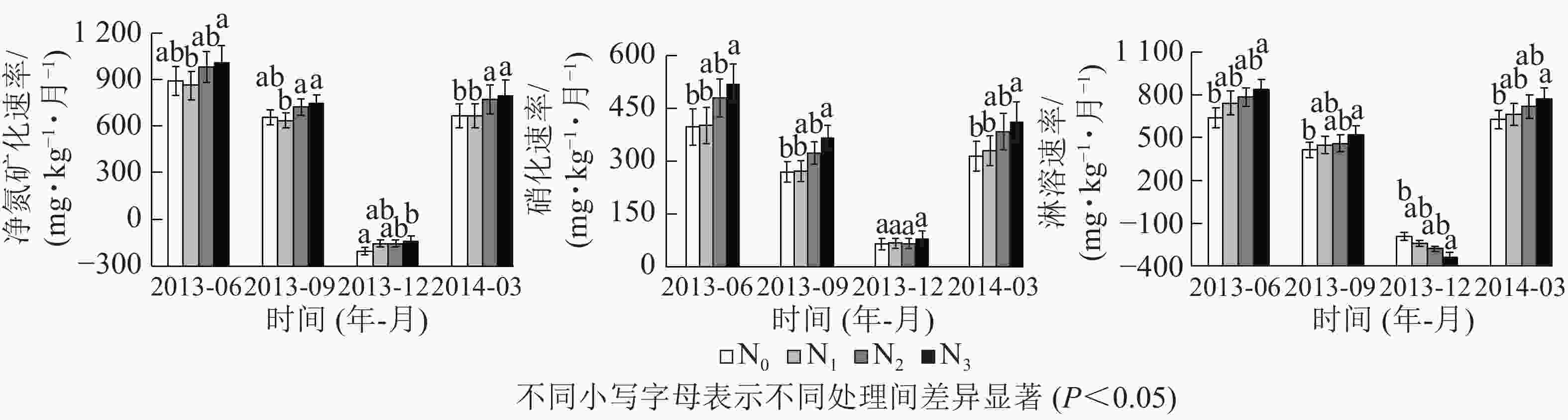
图 2 0~10 cm土层土壤净氮矿化速率、硝化速率和淋溶速率
Figure 2. Soil N mineralization, nitrification and leaching rates in the 0−10 cm soil layer
-
土壤中全氮主要包括有机氮和无机氮,无机氮大部分由铵态氮和硝态氮组成[26]。本研究发现:氮添加显著增加了无机氮及组分质量分数,与NIE等[8]研究结果相似;表明外源氮输入能够增加土壤氮养分,原因可能是氮添加为氮矿化提供了充足氮源,提高了硝化细菌等微生物多样性及活性,增强了微生物对有机质的分解与吸收,产生相对较多的铵态氮与硝态氮[27]。本研究中土壤无机氮以铵态氮形式为主,与CHEN等[13]在鼎湖山阔叶林土壤中发现铵态氮占优势的结果一致。土壤pH可能是造成无机氮差异的重要因素,因为研究区土壤pH为3.85~4.28,远低于硝化细菌适宜土壤pH(6.7~8.3)[28]。酸性土壤条件下,硝化细菌活性弱,施加的氮素(尿素)主要被脲酶转化为铵态氮,造成铵态氮在土壤中累积[29];同时由于硝酸根带负电荷,更容易淋失[25],进一步导致硝态氮质量分数降低。本研究发现:铵态氮/硝态氮随氮添加的升高而降低,这可能是由于外源氮添加对微生物活性产生激发效应,并随氮添加效应增强[30]。氮素施加激发了处于休眠或具有潜在活性的微生物增长,增加了微生物对能耗较小的铵态氮利用[26],从而导致铵态氮在无机氮中的比例下降。
氮沉降通过影响土壤氮素储量进而影响土壤氮矿化过程,而该过程受非生物因素和生物因素共同调控[7]。研究区为酸性土,土壤pH随施肥量增加显著降低[4]。一般认为,土壤酸化通过降低硝化细菌等微生物多样性与活性抑制氮矿化作用[6]。本研究发现:较低的土壤pH下净氮矿化速率更高,与李一凡等[30]在重庆缙云山常绿阔叶林发现土壤铵化、硝化速率等随土壤pH增加而下降的结果一致。推测原因是,长期氮添加下,与氮矿化相关的微生物适应了酸性环境,而较高的土壤pH下有机氮素转化受到抑制[31]。另外,碳氮比也在氮素转化过程中也扮演重要作用,充足氮素条件下,微生物处于碳限制,导致氮素转化速率通常与碳氮比变化相反[5]。本研究发现:随氮添加升高碳氮比显著下降。然而,土壤氮矿化对氮添加响应具有阈值,过高的氮输入也会降低微生物活性,导致产生的胞外酶下降,反而降低土壤氮矿化速率,使得土壤生态系统达到氮饱和或过饱和[32]。本研究暂未发现这种现象,一方面是因为研究区杉木人工林母质气候等非生物环境和微生物群落组成和多样性等生物条件不同,而不同生态系统中氮矿化对不同氮沉降水平响应存在差异性[7, 24]。另一方面,研究区杉木林种植于1992年,“S”型曲线生长的杉木林仍处在速生阶段,对氮素等养分的需求较高[33]。
土壤硝化作用是氮循环的关键过程,能够改变土壤氮的有效性及促进反硝化作用,并刺激氧化亚氮(N2O)的排放,该过程受到土壤性质和气候特征等因素所影响[16]。本研究发现:硝化速率、淋溶速率对氮添加的响应规律与净氮矿化速率具有相似性,这表明土壤中各种氮素转化通量关系密切[23]。由于有机氮矿化为硝化过程提供更多的基质,矿化产生的一部分铵态氮不稳定,经异养硝化立即转变为硝态氮[34],因而硝化速率也随氮添加梯度增大。本研究利用PVC管盖法测定了原位最大潜在淋溶速率[22-23],结果发现:高氮处理显著促进淋溶速率,造成了更多无机氮淋失,这与王甜等[35]在三峡库区秭归县人工种植的柑橘Citrus reticulata园中发现的结果相似。淋溶成分以硝态氮为主,会通过径流、渗透和反硝化还原为N2O等途径流失[36]。本研究中,硝态氮随氮添加梯度在无机氮中占比逐步增大,增加了潜在淋溶风险。
-
气温和降水是影响无机氮及其组分的最主要气候因子[37-38]。本研究发现:季节变化对铵态氮、硝态氮影响显著,这与肖好燕等[39]对亚热带地区天然林、格氏栲Castanopsis kawakamii人工林和杉木人工林土壤矿质氮研究结果一致。在本研究中土壤铵态氮质量分数夏秋高于冬春,而硝态氮则正好相反。原因一方面是夏秋季杉木林正处于生长季,会吸收大量硝态氮从而减少硝态氮累积[38]。另一方面,夏秋季降雨量大,带负电荷的硝态氮淋失加剧[25];而在雨量小的冬春季,铵态氮在硝化微生物作用下转化为硝态氮,使得硝态氮累积,铵态氮减少。因此,表现出2种无机氮组分在夏秋季和冬春季相反的季节格局。
研究发现季节也对氮素转化速率影响显著,均呈现夏季高,冬季低的季节动态。通常不同季节造成土壤温湿度产生差异,进而影响微生物的丰度、多样性和活性等,改变与氮素转化相关酶的分泌,对土壤氮素循环过程产生影响[25]。在本研究中,各季节土壤含水率相差不大(23.22%~27.33%),但土壤温度夏季明显高于冬季[40],推测该研究区人工杉木林土壤氮素转化季节动态主要驱动因子为温度。因此,在全球变暖背景下,需要进一步监测氮循环对变暖的响应。
-
本研究利用野外原位试验与实验室分析相结合,探究了氮添加梯度和季节变化对人工杉木林土壤氮素转化的影响。研究发现高氮添加显著增强了氮素转化速率,土壤无机氮养分质量分数随氮添加梯度增加而增大,有利于杉木吸收和生长,但同时由于氮素淋溶增加,加大了土壤无机氮损失的风险,影响土壤养分维持和土壤健康。因此,在未来氮沉降与全球变暖的背景下,需要继续监测杉木人工林土壤氮养分库和转化速率,为杉木人工林土壤健康管理和可持续经营提供科学指导。
Response of soil N mineralization to long-term N addition and season in Cunninghamia lanceolata plantation
-
摘要:
目的 探讨长期氮沉降和季节变化对杉木Cunninghamia lanceolata人工林无机氮及氮素转化速率的影响。 方法 以福建省三明市沙县官庄国有林场亚热带人工杉木林为研究对象,开展长期(10 a)氮添加梯度(对照:N0;低氮:N1;中氮:N2;高氮:N3)野外控制试验,通过野外原位培养方法测定氮添加对净氮矿化、硝化和淋溶的影响。 结果 ①氮添加显著提高了铵态氮(NH4 +-N)、硝态氮(NO3 −-N) 和总无机氮质量分数,从大到小均依次为N3、N2、N1、N0,且铵态氮质量分数均高于硝态氮。②氮素转化速率随氮添加梯度而增大,高氮显著促进了氮素转化(P<0.05)。③季节显著影响氮素转化(P<0.05),净氮矿化速率、硝化速率与淋溶速率均表现为夏季高、冬季低的季节动态。 结论 氮添加显著增加了土壤无机氮与氮素转化速率,土壤pH、碳氮比(C/N)和土壤温度可能是研究区氮添加驱动氮素转化的主要因子。在杉木人工林的经营与管理中需要更多关注土壤养分和氮素转化速率对外源氮输入的响应。图2表3参40 Abstract:Objective With an experiment conducted Guanzhuang National Forestry Farm, Sha County, Sanming City of Fujian Province, this study is aimed to investigate the effects of long-term nitrogen (N) deposition and seasonal change on inorganic N and N transformation rates in the Cunninghamia lanceolata plantation. Method Besides a long-term (10-year-long) N addition experiment with four gradients (control: N0, low: N1, medium: N2, high: N3), the cultivation experiment in situ was conducted to determinate the N mineralization, nitrification and leaching rates in response to N addition. Result (1) N addition significantly increased the mass fractions of ammonium N (NH4 +-N), nitrate N (NO3 −-N) and total inorganic N, showing the trend of N3, N2, N1 and N0, and the ammonium N was higher than nitrate N. (2) N transformation rates increased with the N addition gradients, while high N addition significantly promoted the N transformation rates. (3) Seasonal changes significantly affected N transformation rates, and the N mineralization, nitrification and leaching rates were higher in summer and lower in winter. Conclusion N addition significantly increased soil inorganic N and N transformation rates whereas soil pH, C/N ratio and temperature may be the factors of N addition driving soil inorganic N and N transformation rates in the C. lanceolata plantation . Therefore, closer attention should be paid to the response of soil nutrients and N transformation rates to exogenous N input in the management of C. lanceolata plantation. [Ch, 2 fig. 3 tab. 40 ref.] -
Key words:
- N mineralization /
- nitrification /
- N leaching /
- global change /
- soil nutrients
-
图 1 0~10 cm土层铵态氮、硝态氮和无机氮质量分数
Figure 1 Mass fractions of ammonium, nitrate and inorganic N in 0−10 cm soil layer
图 2 0~10 cm土层土壤净氮矿化速率、硝化速率和淋溶速率
Figure 2 Soil N mineralization, nitrification and leaching rates in the 0−10 cm soil layer
表 1 不同氮添加处理土壤主要理化和养分性质
Table 1. Main soil physicochemical and nutrient properties under different N addition
处理 pH 有机质/(g·kg−1) 全氮/(g·kg−1) C/N 铵态氮/(mg·kg−1) N0 4.28±0.19 a 30.57±2.89 a 1.35±0.14 c 13.3±2.0 a 27.14±4.58 d N1 4.14±0.24 b 27.13±3.07 b 1.57±0.17 b 10.1±1.4 b 32.19±6.56 c N2 4.00±0.22 c 26.82±3.19 b 1.62±0.19 ab 9.7±1.7 bc 39.13±6.35 b N3 3.85±0.16 d 25.32±3.34 b 1.67±0.18 a 8.9±1.7 cd 44.51±6.25 a 处理 铵态氮比例/% 硝态氮/(mg·kg−1) 硝态氮比例/% 铵态氮/硝态氮 无机氮/(mg·kg−1) N0 68.9 12.27±2.51 d 31.1 2.2 39.41±4.79 d N1 63.0 18.67±3.76 c 37.0 1.7 50.86±7.55 c N2 63.0 23.51±5.49 b 37.0 1.7 62.64±6.78 b N3 58.4 31.67±5.12 a 41.6 1.4 76.17±8.26 a 说明:数值为平均值±标准误,同列不同字母表示差异显著(P<0.05),铵态氮比例为铵态氮占无机氮比例,硝态氮比例为硝态氮占 无机氮比例  下载: 导出CSV
下载: 导出CSV
表 2 氮添加和季节影响氮质量分数的双因素方差分析
Table 2. Effects of N addition, season, and their interactions on N mass fraction as indicated by two-way ANOVAs
处理 P 铵态氮 硝态氮 无机氮 氮添加 0.000* 0.000* 0.000* 季节 0.015* 0.005* 0.889 氮添加×季节 0.994 0.944 0.999 说明:*表示处理间差异显著
下载: 导出CSV
表 3 氮添加和季节影响氮素转化速率的双因素方差分析
Table 3. Effects of N addition, season, and their interactions on N transformation rates as indicated by two-way ANOVAs
处理 P 矿化速率 硝化速率 淋溶速率 氮添加 0.000* 0.001* 0.042* 季节 0.000* 0.000* 0.000* 氮水平×季节 0.012* 0.693 0.955 说明:*表示处理间差异显著
下载: 导出CSV
-
[1] LIU Xuejun, ZHANG Ying, HAN Wenxuan, et al. Enhanced nitrogen deposition over China [J]. Nature, 2013, 494(7438): 459 − 462. [2] GALLOWAY J N, TOWNSEND A R, ERISMAN J W, et al. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions [J]. Science, 2008, 320(5878): 889 − 892. [3] LIU Shengjie, BEHM J E, WAN Shiqiang, et al. Effects of canopy nitrogen addition on soil fauna and litter decomposition rate in a temperate forest and a subtropical forest [J]. Geoderma, 2021, 382: 114703. doi:10.1016/j.geoderma.2020.114703. [4] WU Jianping, LIU Wenfei, ZHANG Weixin, et al. Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest [J]. Appl Soil Ecol, 2019, 142: 43 − 51. [5] LI Zhaolei, ZENG Zhaoqi, TIAN Dashuan, et al. Global variations and controlling factors of soil nitrogen turnover rate [J]. Earth Sci Rev, 2020, 207: 103250. doi: 10.1016/j.earscirev.2020.103250. [6] LI Zhaolei, TIAN Dashuan, WANG Bingxue, et al. Microbes drive global soil nitrogen mineralization and availability [J]. Global Change Biol, 2019, 25(3): 1078 − 1088. [7] 左倩倩, 王邵军. 生物与非生物因素对森林土壤氮矿化的调控机制[J]. 浙江农林大学学报, 2021, 38(3): 613 − 623. ZUO Qianqian, WANG Shaojun. Regulation mechanism of biological and abiotic factors on the nitrogen mineralization of forest soil [J]. J Zhejiang A&F Univ, 2021, 38(3): 613 − 623. [8] NIE Yanxia, HAN Xiaoge, CHEN Jie, et al. The simulated N deposition accelerates net N mineralization and nitrification in a tropical forest soil [J]. Biogeosciences, 2019, 16(21): 4277 − 4291. [9] CORRE M D, VELDKAMP E, ARNOLD J, et al. Impact of elevated N input on soil N cycling and losses in old-growth lowland and montane forests in Panama [J]. Ecology, 2010, 91(6): 1715 − 1729. [10] SHARON J H, PAMELA A M. Nutrient status of tropical rain forests influences soil N dynamics after N additions [J]. Ecol Monogr, 2003, 73(1): 107 − 129. [11] YAN Guoyong, XING Yajuan, WANG Jianyu, et al. Sequestration of atmospheric CO2 in boreal forest carbon pools in Northeastern China: effects of nitrogen deposition [J]. Agric For Meteorol, 2018, 248: 70 − 81. [12] LU Xiaofei, HOU Enqing, GUO Jieyun, et al. Nitrogen addition stimulates soil aggregation and enhances carbon storage in terrestrial ecosystems of China: a meta-analysis [J]. Global Change Biol, 2021, 27(12): 2780 − 2792. [13] CHEN Hao, GURMESA G A, ZHANG Wei, et al. Nitrogen saturation in humid tropical forests after 6 years of nitrogen and phosphorus addition: hypothesis testing [J]. Funct Ecol, 2016, 30(2): 305 − 313. [14] HÖGBERG P, FAN Houbao, QUIST M, et al. Tree growth and soil acidification in response to 30 years of experimental nitrogen loading on boreal forest [J]. Global Change Biol, 2006, 12(3): 489 − 499. [15] ÅGREN G I, BOSATTA E. Nitrogen saturation of terrestrial ecosystems [J]. Environ Pollut, 1988, 54(3/4): 185 − 197. [16] LI Zhaolei, ZENG Zhaoqi, TIAN Dashuan, et al. Global patterns and controlling factors of soil nitrification rate [J]. Global Change Biol, 2020, 26(7): 4147 − 4157. [17] YANG W H, RYALS R A, CUSACK D F, et al. Cross-biome assessment of gross soil nitrogen cycling in California ecosystems [J]. Soil Biol Biochem, 2017, 107: 144 − 155. [18] WEI Xiaohua, BLANCO J A, JIANG Hong, et al. Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems [J]. Sci Total Environ, 2012, 416: 351 − 361. [19] 林雨萱, 哀建国, 宋新章, 等. 模拟氮沉降和磷添加对杉木林土壤呼吸的影响[J]. 浙江农林大学学报, 2021, 38(3): 494 − 501. LIN Yuxuan, AI Jianguo, SONG Xinzhang, et al. Effects of simulated nitrogen deposition and phosphorus addition on soil respiration in Cunninghamia lanceolata forest [J]. J Zhejiang A&F Univ, 2021, 38(3): 494 − 501. [20] 聂二旗, 张心昱, 郑国砥, 等. 氮磷添加对杉木林土壤碳氮矿化速率及酶动力学特征的影响[J]. 生态学报, 2018, 38(2): 615 − 623. NIE Erqi, ZHANG Xinyu, ZHENG Guodi, et al. Effects of nitrogen and phosphorus addition on soil carbon and nitrogen mineralization rate and enzyme kinetics characteristics of Cunninghamia lanceolata forest [J]. Acta Ecol Sin, 2018, 38(2): 615 − 623. [21] 蔡乾坤, 段洪浪, 刘文飞, 等. 氮沉降对杉木林土壤有效氮和磷含量的影响[J]. 森林与环境学报, 2016, 36(3): 342 − 348. CAI Qiankun, DUAN Honglang, LIU Wenfei, et al. Effects of nitrogen deposition on soil available N and P contents in Cunninghamia lanceolata forest [J]. J For Environ, 2016, 36(3): 342 − 348. [22] RAISON R J, CONNELL M J, KHANNA P K. Methodology for studying fluxes of soil mineral-N in situ [J]. Soil Biol Biochem, 1987, 19(5): 521 − 530. [23] 刘振林, 刘广生, 蒋乃华, 等. 土壤理化分析与土壤剖面描述 [M]. 北京: 中国标准出版社, 1996. [24] WANG Faming, ZHU Weixing, XIA Hanping, et al. Nitrogen mineralization and leaching in the early stages of a subtropical reforestation in Southern China [J]. Restor Ecol, 2010, 18: 313 − 322. [25] CHEN Jie, XIAO Guoliang, KUZYAKOV Y, et al. Soil nitrogen transformation responses to seasonal precipitation changes are regulated by changes in functional microbial abundance in a subtropical forest [J]. Biogeosciences, 2017, 14(9): 2513 − 2525. [26] 王帅, 张永发, 任常琦, 等. 早期不同施氮量下橡胶园土壤中无机氮的运移 [J]. 热带作物学报, 2021. doi: 46.1019.S.20210607.1108.002. WANG Shuai, ZHANG Yongfa, REN Changqi, et al. Inorganic nitrogen transport in rubber plantation soil under different nitrogen application rates at early stage [J]. J Trop Crop, 2021. doi: 46.1019.S.20210607.1108.002. [27] 胡星云, 孙志高, 张党玉, 等. 外源氮输入对黄河口碱蓬湿地土壤碳氮含量动态的影响[J]. 水土保持学报, 2017, 31(6): 204 − 211. HU Xingyun, SUN Zhigao, ZHANG Dangyu, et al. Effects of exogenous nitrogen input on soil carbon and nitrogen content dynamics of Suaeda salsa wetland in the Yellow River Estuary [J]. J Soil Water Conserv, 2017, 31(6): 204 − 211. [28] 黄孝肖, 陈重军, 张蕊, 等. 厌氧氨氧化与反硝化耦合反应研究进展[J]. 应用生态学报, 2012, 23(3): 849 − 856. HUANG Xiaoxiao, CHEN Zhongjun, ZHANG Rui, et al. Research progress of anammox and denitrification coupling reaction [J]. Chin J Appl Ecol, 2012, 23(3): 849 − 856. [29] 刘少文, 殷敏, 褚光, 等. 土壤氮激发效应及其微生物机理研究进展[J]. 中国水稻科学, 2019, 33(4): 303 − 312. LIU Shaowen, YIN Min, CHU Guang, et al. Research progress on soil nitrogen stimulation effect and microbial mechanism [J]. Rice Sci China, 2019, 33(4): 303 − 312. [30] 李一凡, 王玉杰, 王彬, 等. 西南酸雨区重庆缙云山常绿阔叶林土壤氮矿化特征[J]. 林业科学, 2019, 55(6): 1 − 12. LI Yifan, WANG Yujie, WANG Bin, et al. Soil nitrogen mineralization in an evergreen broad-leaved forest of Jinyun Mountain, southwest China [J]. Sci Silv Sin, 2019, 55(6): 1 − 12. [31] KEMMITT S, WRIGHT D, GOULDING K, et al. pH regulation of carbon and nitrogen dynamics in two agricultural soils [J]. Soil Biol Biochem, 2006, 38(5): 898 − 911. [32] CHENG Yi, WANG Jing, GE Zhiwei, et al. Background nitrogen deposition controls the effects of experimental nitrogen addition on soil gross N transformations in forest ecosystems [J]. Biogeochemistry, 2020, 151(2/3): 335 − 341. [33] 杨传桃. 不同年龄杉木林分结构与生长过程 [D]. 长沙: 中南林业科技大学, 2019. YANG Chuantao. The Structure and Growth Process of Chinese Fir Plantation at Different Stand Ages [D]. Changsha: Central South University of Forestry and Technology, 2019. [34] BUTLER S M, MELILLO J M, JOHNSON J E, et al. Soil warming alters nitrogen cycling in a New England forest: implications for ecosystem function and structure [J]. Oecologia, 2012, 168(3): 819 − 828. [35] 王甜, 黄志霖, 曾立雄, 等. 不同施肥处理对三峡库区柑橘园土壤氮磷淋失影响[J]. 水土保持学报, 2018, 32(5): 53 − 57. WANG Tian, HUANG Zhilin, ZENG Lixiong, et al. Effects of different fertilization treatments on soil nitrogen and phosphorus leaching in Citrus reticulata Blanco orchards in the Three Gorges Reservoir Region [J]. J Soil Water Conserv, 2018, 32(5): 53 − 57. [36] CHEN Zhe, LIN Shan, YAO Zhisheng, et al. Enhanced nitrogen cycling and N2O loss in water-saving ground cover rice production systems (GCRPS) [J]. Soil Biol Biochem, 2018, 121: 77 − 86. [37] SONG Yanyu, SONG Changchun, HOU Aixin, et al. Effects of temperature and root additions on soil carbon and nitrogen mineralization in a predominantly permafrost peatland [J]. Catena, 2018, 165: 381 − 389. [38] HISHI T, URAKAWA R, TASHIRO N, et al. Seasonality of factors controlling N mineralization rates among slope positions and aspects in cool-temperate deciduous natural forests and larch plantations [J]. Biol Fert Soils, 2014, 50(2): 343 − 356. [39] 肖好燕, 刘宝, 余再鹏, 等. 亚热带不同林分土壤矿质氮库及氮矿化速率的季节动态[J]. 应用生态学报, 2017, 28(3): 730 − 738. XIAO Haoyan, LIU Bao, YU Zaipeng, et al. Seasonal dynamics of soil mineral nitrogen pools and nitrogen mineralization rates in a subtropical forest [J]. Chin J Appl Ecol, 2017, 28(3): 730 − 738. [40] FAN Houbao, WU Jianping, LIU Wenfei, et al. Nitrogen deposition promotes ecosystem carbon accumulation by reducing soil carbon emission in a subtropical forest [J]. Plant Soil, 2014, 379: 361 − 371. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210403
 点击查看大图
点击查看大图
计量
- 文章访问数: 2135
- HTML全文浏览量: 491
- PDF下载量: 88
- 被引次数: 0




