






-
土壤微生物参与森林生态系统中很多生态过程[1]。土壤细菌群落是土壤养分循环的参与者,在有机质的矿化、无机元素的转化以及土壤碳氮的积累等过程中发挥着重要作用[2-5];土壤真菌通过产生多种胞外酶,可以有效分解各类有机质,将死去的有机物转化为生物量、二氧化碳和有机酸,从而调节碳和养分的平衡[6-8]。
气候变暖以及氮沉降对土壤微生物群落的结构、多样性和丰富度有显著影响[9]。山地海拔梯度作为模拟气候变化的有效方法[10],其特点是在较短的地理距离上发生巨大的生物与非生物变化[11],例如温度和湿度等气候要素[1],因此,海拔梯度可以探究土壤微生物群落对气候变化的响应[12]。研究表明:土壤微生物多样性随海拔的变化有多种模式,包括下降、增加、中海拔峰值和极端海拔峰值等[13-15]。这是由于影响微生物群落变化的因子多种多样。LOOBY等[13]在土壤微生物与海拔关系的研究综述中阐明:气候和土壤(尤其是pH)是驱动土壤微生物群落变化的主要机制。在对亚热带高山区域的研究中,赵盼盼等[16]研究表明:温度与可溶性有机氮(DON)对土壤微生物群落结构变化起主导作用;孟苗婧等[17]发现:土壤理化因子是海拔梯度上土壤微生物群落产生差异的主导因子。此外,植物群落对土壤特性有重要影响,从而改变土壤微生物的海拔分布[18]。因此,气候因素、土壤理化性质和植物群落均影响土壤微生物多样性和群落组成[19]。
目前,亚热带土壤微生物群落结构对海拔梯度响应的研究,主要针对细菌、真菌等大类的变化,而对不同分类水平下具体类群的关注相对较少。例如,在福建省戴云山自然保护区的研究表明:黄山松Pinus taiwanensis林土壤细菌/真菌比随海拔升高显著下降[16];对武夷山主要植被类型土壤的研究表明:真菌数量随海拔升高而降低[20]。针对浙江凤阳山针阔混交林,孟苗婧等[17]利用磷脂脂肪酸法发现革兰氏阴性细菌丰度随海拔升高而增大,并且这种变化与土壤理化性质有关。这些大类群的变化并不能很好地表征土壤细菌和真菌在门水平乃至属水平的变化特征,因此,本研究通过高通量测序,探索凤阳山常绿阔叶林土壤微生物群落在不同海拔梯度下的变化规律,以增加对土壤微生物群落组成和多样性的了解。
-
研究区位于浙江省丽水市龙泉市凤阳山自然保护区(27°46′~27°58′N,119°06′~119°15′E),面积约1.52万hm²,属中亚热带温暖湿润气候区。保护区年降水量为2 400 mm,年蒸发量为1 100 mm,年平均气温为12.3 ℃。主要天然植被为常绿阔叶林与针叶林,常绿阔叶林的优势属为木荷属Schima、润楠属Machilus、石栎属Lithocarpus、栲属Castanopsis、青冈属Cyclobalanopsis。土壤类型以红壤、黄壤为主,800 m以下为红壤,800 m以上为黄壤[17, 21-22]。
-
选取4个不同海拔梯度下坡度、坡向、地上植被特征基本一致的常绿阔叶林地。海拔选择以土壤类型分界和凤阳山常绿阔叶林海拔分布范围为依据,分别为低海拔343 m (EG1)、中海拔765 m (EG2)、高海拔1364 m (EG3)、高海拔1 611 m (EG4)。不同海拔的林分主要树种及郁闭度见表1。去除表面凋落物后,在每个海拔内设立10 m×10 m的样地3个,间隔10 m以上,在每个样地内沿对角线采集混合土壤样品,用于土壤理化性质的测定(表2)。考虑到土壤微生物异质性可能更强,在相同区域内按5 m×5 m,间隔10 m采集4个混合土样,−80 ℃冰箱保存,用于土壤微生物的高通量测序分析。由于本研究没有直接在研究地内监测温度,故采用凤阳山森林生态长期定位站设置的不同海拔梯度(241、600、900、1 364 m)温度监测数据进行拟合得到。年平均气温(y)随海拔(x)变化关系为y=−0.0063x+21.592 (R2=0.9942)。拟合得出4个海拔(EG1、EG2、EG3、EG4)的年平均气温分别为19.43、16.77、13.15、11.44 ℃。
表 1 不同海拔林分特征
Table 1. General Situation of the trees at different altitudes
海拔编号 树种组成 郁闭度/% EG1 木荷Schima superba、甜槠Castanopsis eyrei、青冈Cyclobalanopsis glauca、石栎Lithocarpus glaber、
檵木Loropetalum chinense等91 EG2 木荷、甜槠、青冈、檵木、山鸡椒Litsea cubeba等 87 EG3 木荷、甜槠、青冈、杨桐Cleyera japonica、马银花Rhododendron ovatum等 92 EG4 甜槠、青冈、马银花、尖叶山茶Camellia cuspidata、麂角杜鹃Rhododendron latoucheae等 90 表 2 不同海拔土壤理化性质
Table 2. Physical and chemical properties of soil at different altitudes
海拔编号 pH 土壤湿度/% 总碳/(mg·g−1) 全氮/(mg·g−1) 全磷/(mg·g−1) 全钾/(mg·g−1) EG1 4.86±0.03 a 15.54±6.41 c 103.97±7.60 a 3.57±0.35 b 0.21±0.04 c 19.63±2.58 a EG2 4.93±0.18 a 43.45±19.89 b 160.53±96.86 a 9.43±4.65 a 0.51±0.12 b 14.13±1.37 b EG3 4.67±0.07 a 38.41±7.81 bc 117.23±25.94 a 7.13±1.12 ab 0.45±0.16 b 11.43±1.03 b EG4 4.73±0.21 a 71.36±14.42 a 125.97±45.76 a 9.63±3.09 a 0.73±0.05 a 14.37±1.58 b 说明:数值为平均值±标准差(n=3),不同小写字母表示不同海拔间差异显著(P<0.05) -
pH采用电位法(水土比为2.5∶1.0)通过pH计测定;土壤湿度采用环刀法测定;土壤总碳、全氮采用德国Element公司的元素分析仪测定(动态燃烧法);土壤全磷采用酸溶-钼锑抗比色法,通过分光光度计测定;土壤全钾采用氢氧化钠熔融,火焰光度计测定[23]。
-
采用土壤DNA试剂盒(试剂盒名称为MagPure Soil DNA KF Kit),从0.25 g土壤样品中提取DNA,利用琼脂糖凝胶电泳和NanoDrop 2000检测DNA浓度。以基因组DNA为模板,根据测序区域的选择,使用带 barcode 的特异引物,Takara公司的TksGflex DNA Polymerase进行聚合酶链式反应(PCR)。细菌16SrDNAV3-V4扩增引物为343F-5′-TACGGRAGGCAGCAG-3′和798R-5′- AGGGTATCTAATCCT-3′,真菌ITS扩增引物为ITS1F-5′-CTTGGTCATTTAGAGGAAGTAA-3′和ITS2-5′- GCTGCGTTCTTCATCGATGC-3′。测序委托中国上海欧易生物医学科技有限公司Illumina MiSeq平台进行。将获得的原始序列用一系列软件Trimmonatic[24]、Flash[25]和QIIME中的split_libraries[26]处理,得到优化序列,并使用 UCHIME软件[27]去除嵌合体;利用Vsearch软件[28],生成操作分类单元(OTUs),细菌、真菌相似度均为97%。
-
首先,对所有数据进行正态分布检验和方差齐性检验,结果均符合正态分布。对于土壤理化性质和微生物多样性指数的数据,使用SPSS软件进行单因素方差分析(one-way ANOVA),确定不同海拔下土壤理化性质和土壤微生物多样性的差异(LSD最小显著差异法,显著性水平为P<0.05),其中土壤微生物覆盖度(goods_coverage,Cdepth)指数表示测序的深度。其计算方法为:Cdepth=1−n1/N,其中:n1为只含有1条序列的 OTU 数目;N为抽样中出现的总的序列数;对于门和属水平上各类群相对丰度的数据,利用Pearson相关分析来确定海拔与门和属水平上微生物群落的相关性;对于界、门、纲、目、科、属、种水平上土壤微生物相对丰度的数据,利用组间群落差异[LDA Effect Size(LefSe)]分析 (LefSe:
http://huttenhower.sph.harvard.edu/lefse/ 。设置标准:LDA大于2,P<0.05)来研究不同海拔梯度下土壤微生物群落的差异类群[29],对于OTU水平上土壤微生物序列数的数据,使用Canoco 5.0进行主坐标分析(PCoA)。由于微生物和土壤理化性质样品数量不一致,无法一一对应进行关联分析,故通过Bootstrap重采样方法,将每个海拔土壤微生物(n=4)与土壤理化性质(n=3)进行两两组合(12种可能),从12种可能中随机抽取3个作为重复(n=3),4个海拔共采集12个样品;重采样通过Matlab进行1000次,每1次都计算土壤微生物与土壤理化性质的Pearson相关系数,得到相关系数的频率分布,最终计算得到平均相关系数。对照Pearson相关系数显著性检验表中0.05显著性水平下相关系数的临界值,采用双侧检验判断显著性。 -
由图1A可知:细菌群落的OTU总数为8408个,共享OTU数 2828个。其中343、765、1364、1611 m特有OTU个数分别占到细菌OTU总数的7.67%、5.00%、6.24%、6.89%。两两比较中,中低海拔即343与765 m共有OTU数最多,为4593个。海拔变化同样引起土壤真菌群落OTU数量变化。如图1B所示:检测出的真菌群落OTU总数少于细菌群落,为3985个。不同海拔真菌群落共有的OTU占到真菌群落总数的4.19%。无论是细菌群落或真菌群落,中低海拔OTU数均高于高海拔;两两比较中,343与765 m共有的OTU数最多。这一结果表明343与765 m样地OTU组成相似。
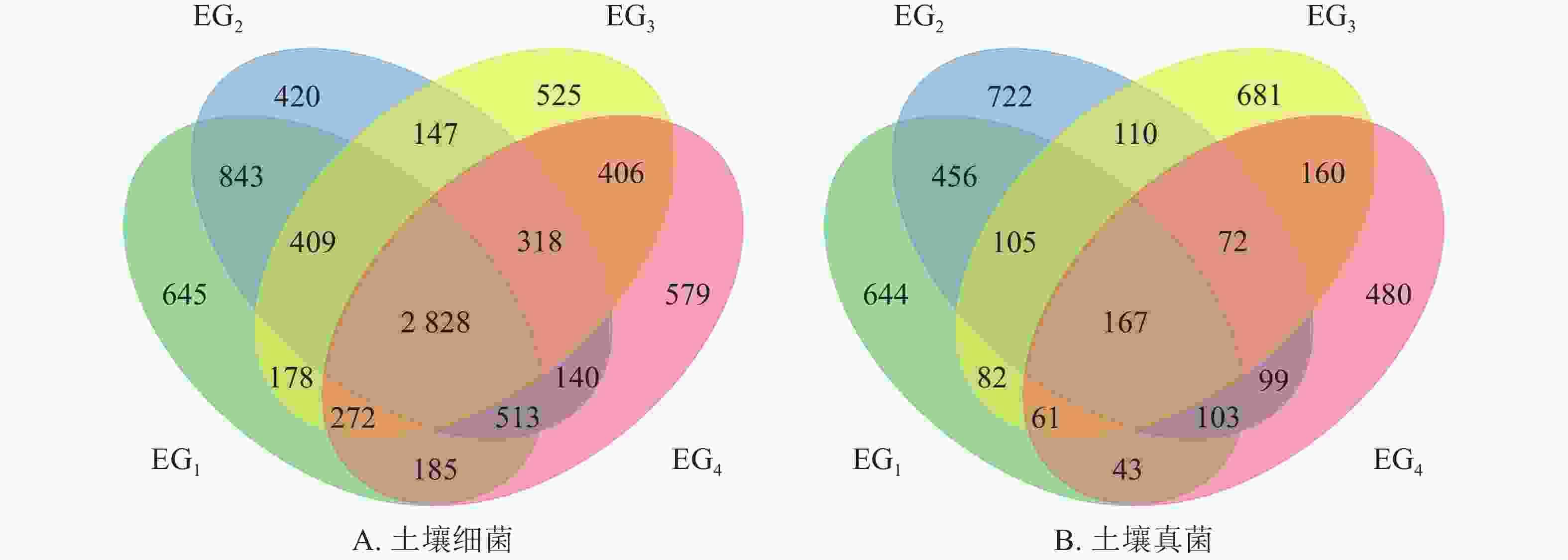
图 1 不同海拔土壤细菌(A)和真菌(B)群落维恩图
Figure 1. Venn diagram showing the unique and shared OTUs at different elevations in bacterial (A) and fungal (B) communities
-
Chao 1指数与Shannon指数分别反映了微生物群落的丰富度与多样性。细菌群落的多样性分析发现:中低海拔Chao 1指数显著高于高海拔1364与1611 m,呈现出沿海拔增高逐渐减小的趋势(P<0.05)(表3)。Shannon指数在343 m处最高,显著高于765 m处(P<0.05)。真菌群落的多样性结果显示:高海拔Chao 1指数显著低于中低海拔(P<0.05),Shannon指数在343 m处最低,且显著低于765与1364 m处(P<0.05)。细菌群落与真菌群落覆盖度指数均在0.97以上,且中低海拔呈现较低的水平(P<0.05),这说明测序结果可靠。
表 3 不同海拔土壤微生物群落多样性
Table 3. Soil bacterial diversity indices at different altitudes
细菌 真菌 海拔编号 Chao 1指数 Shannon指数 覆盖度 海拔编号 Chao 1指数 Shannon指数 覆盖度 EG1 4506.81±41.85 a 9.2622±0.246 8 a 0.9773±0.000 6 b EG1 783.12±36.26 a 3.6031±0.6633 b 0.9911±0.0006 b EG2 4255.62±295.74 b 8.8781±0.214 4 b 0.9780±0.001 7 b EG2 807.96±115.36 a 5.0242±0.6876 a 0.9911±0.0013 b EG3 3688.80±100.61 c 9.0043±0.231 9 ab 0.9824±0.000 5 a EG3 729.86±52.44 ab 4.6790±0.4236 a 0.9921±0.0002 ab EG4 3791.13±110.41 c 8.9974±0.176 5 ab 0.9815±0.000 8 a EG4 653.00±67.50 b 4.5171±0.6495 ab 0.9928±0.0008 a 说明:数值为平均值±标准差(n = 4),不同小写字母表示不同海拔间差异显著(P<0.05) -
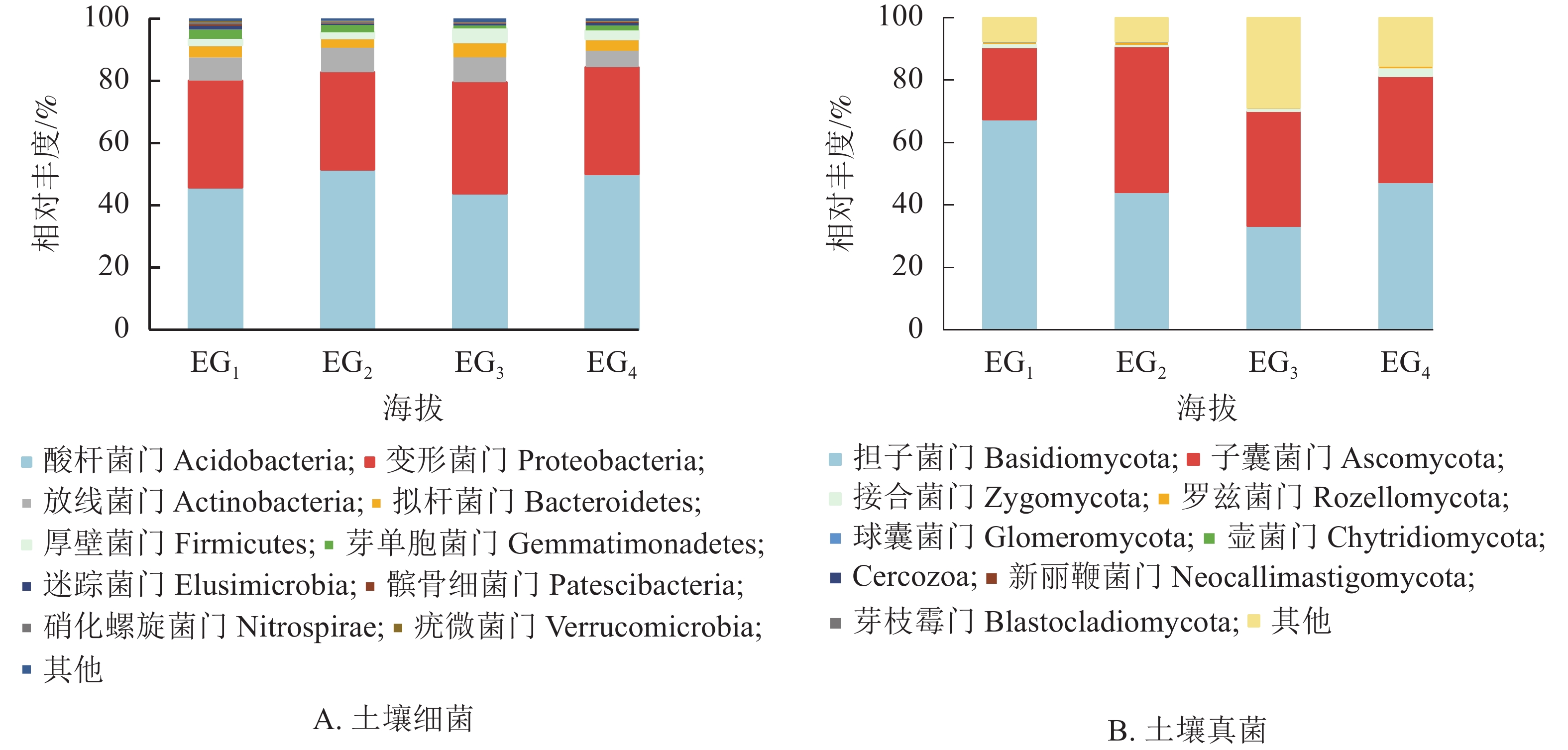
通过测序发现,4个海拔土壤细菌群落共检测出33个门。图2A为门水平上相对丰度前10位的细菌群落。分析显示:凤阳山常绿阔叶林优势菌群为酸杆菌门Acidobacteria、变形菌门Proteobacteria和放线菌门Actinobacteria。其中,酸杆菌门相对丰度高达43.77%~51.55%,是丰度最高的类群,其次是变形菌门(31.18%~35.77%)、放线菌门(5.24%~7.99%)。Pearson相关性分析发现:芽单胞菌门Gemmatimonadetes、硝化螺旋菌门Nitrospirae、疣微菌门Verrucomicrobia与海拔呈极显著负相关(P<0.01),厚壁菌门Firmicutes与海拔呈显著正相关(P<0.05) (表4)。
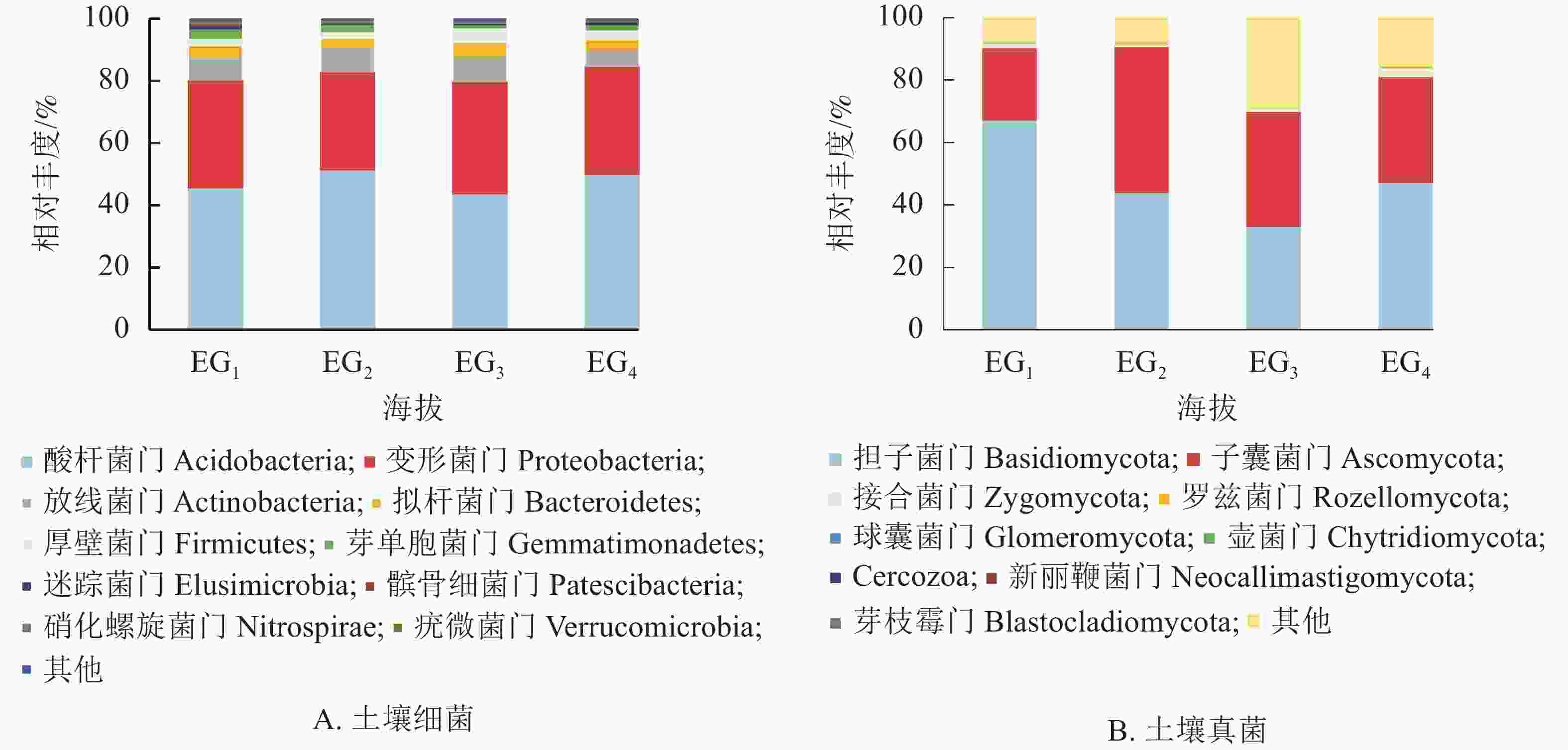
图 2 不同海拔门水平上土壤细菌(A)和真菌(B)群落相对丰度
Figure 2. Relative abundance of bacteria (A) and fungi (B) phylum at different altitudes
表 4 海拔与微生物门Pearson相关性系数
Table 4. Pearson correlation coefficient between altitude and soil bacterial phylum
细菌门 相关性系数 真菌门 相关性系数 细菌门 相关性系数 真菌门 相关性系数 酸杆菌门 0.032 担子菌门 −0.402 芽单胞菌门 −0.817** 壶菌门 −0.209 变形菌门 0.234 子囊菌门 0.173 迷踪菌门 −0.260 Cercozoa −0.191 放线菌门 −0.417 接合菌门 0.437 髌骨细菌门 −0.490 新丽鞭菌门 −0.233 拟杆菌门 0.269 罗兹菌门 −0.308 硝化螺旋菌门 −0.723** 芽枝霉门 −0.236 厚壁菌门 0.525* 球囊菌门 −0.036 疣微菌门 −0.705** 说明:*表示在0.05水平(双侧)上显著相关;**表示在0.01水平(双侧)上显著相关 图2B显示了所有鉴定出的已分类的门水平上的真菌群落。大部分类群隶属担子菌门Basidiomycota (33.16%~67.35%)与子囊菌门Ascomycota (22.98%~46.78%)。343 m处担子菌门最为丰富,相对丰度高达65.07%。子囊菌门是中高海拔相对丰度最高的真菌类群。除上述优势菌群,相对丰度高于1%的为343与1611 m的接合菌门Zygomycota。
-
细菌群落共检测出708个属。图3A展示了相对丰度前15位的细菌属,占21.61%~24.52%。Candidatus_Solibacter相对丰度最高,在343、765、1 611 m 超过5%,Bryobacter在1 611 m最高,Acidibacter在各海拔相对丰度为2.25%~3.01%。相关性分析(表5)显示:Acidibacter、Gemmatimonas、Sphingomonas、MND1与海拔呈显著负相关(P<0.05)。
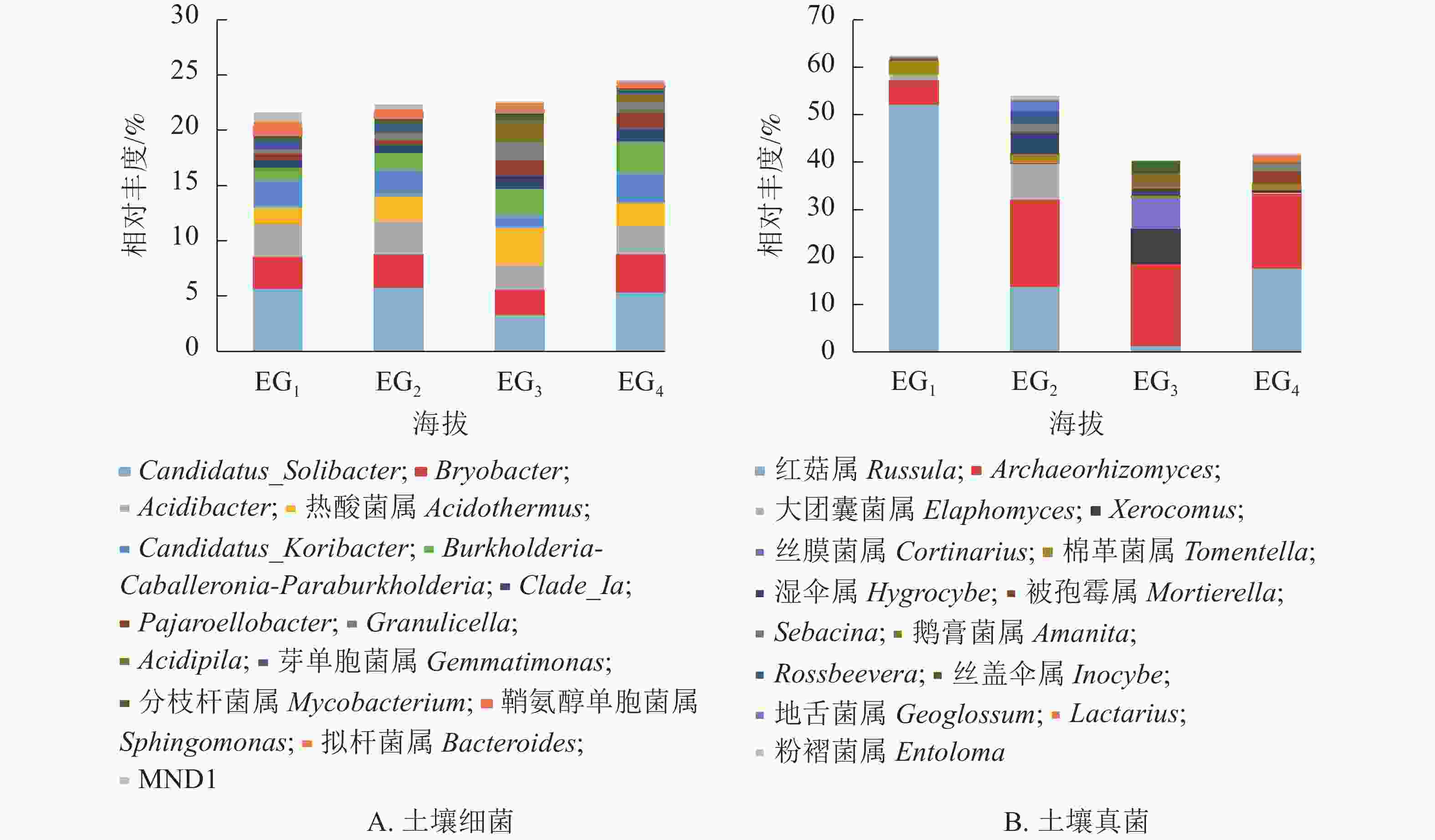
图 3 不同海拔属水平上土壤细菌(A)和真菌(B)群落相对丰度
Figure 3. Relative abundance of bacteria (A) and fungi (B) genus at different altitudes
表 5 海拔与微生物属Pearson相关性系数
Table 5. Pearson correlation coefficient between altitude and soil bacterial genus
细菌属 真菌属 细菌属 真菌属 属名 相关系数 属名 相关系数 属名 相关系数 属名 相关系数 Candidatus_Solibacter −0.458 红菇属Russula −0.645** Granulicella 0.719** Sebacina 0.197 Bryobacter 0.088 Archaeorhizomyces 0.290 Acidipila 0.639** 鹅膏菌属Amanita 0.250 Acidibacter −0.520* 大团囊菌属Elaphomyces −0.234 芽单胞菌属Gemmatimonas −0.734** Rossbeevera −0.127 热酸菌属Acidothermus 0.514* Xerocomus 0.200 分枝杆菌属Mycobacterium 0.035 丝盖伞属Inocybe 0.190 Candidatus_Koribacter −0.178 丝膜菌属Cortinarius 0.182 鞘氨醇单胞菌属Sphingomonas −0.778** 地舌菌属Geoglossum −0.194 Burkholderia-Caballeronia-
Paraburkholderia0.648** 棉革菌属Tomentella −0.259 拟杆菌属Bacteroides 0.224 Lactarius 0.311 Clade_Ia 0.472 湿伞属Hygrocybe −0.097 MND1 −0.743** 粉褶菌属Entoloma −0.209 Pajaroellobacter 0.740** 被孢霉属Morlierella 0.702** 说明:*表示在0.05水平(双侧)上显著相关;**表示在0.01水平(双侧)上显著相关 真菌群落共检测出384个属。图3B展示了相对丰度前15位的真菌属。红菇属Russula在343 m检出较多,相对丰度高达52.30%,中高海拔红菇属相对丰度均低于20%。Archeaorhizomyces相对丰度仅次于红菇属,在中高海拔大量存在。相关性分析(表5)显示:红菇属与海拔呈显著负相关,被孢霉属Morlierella与海拔呈显著正相关(P<0.05)。
-
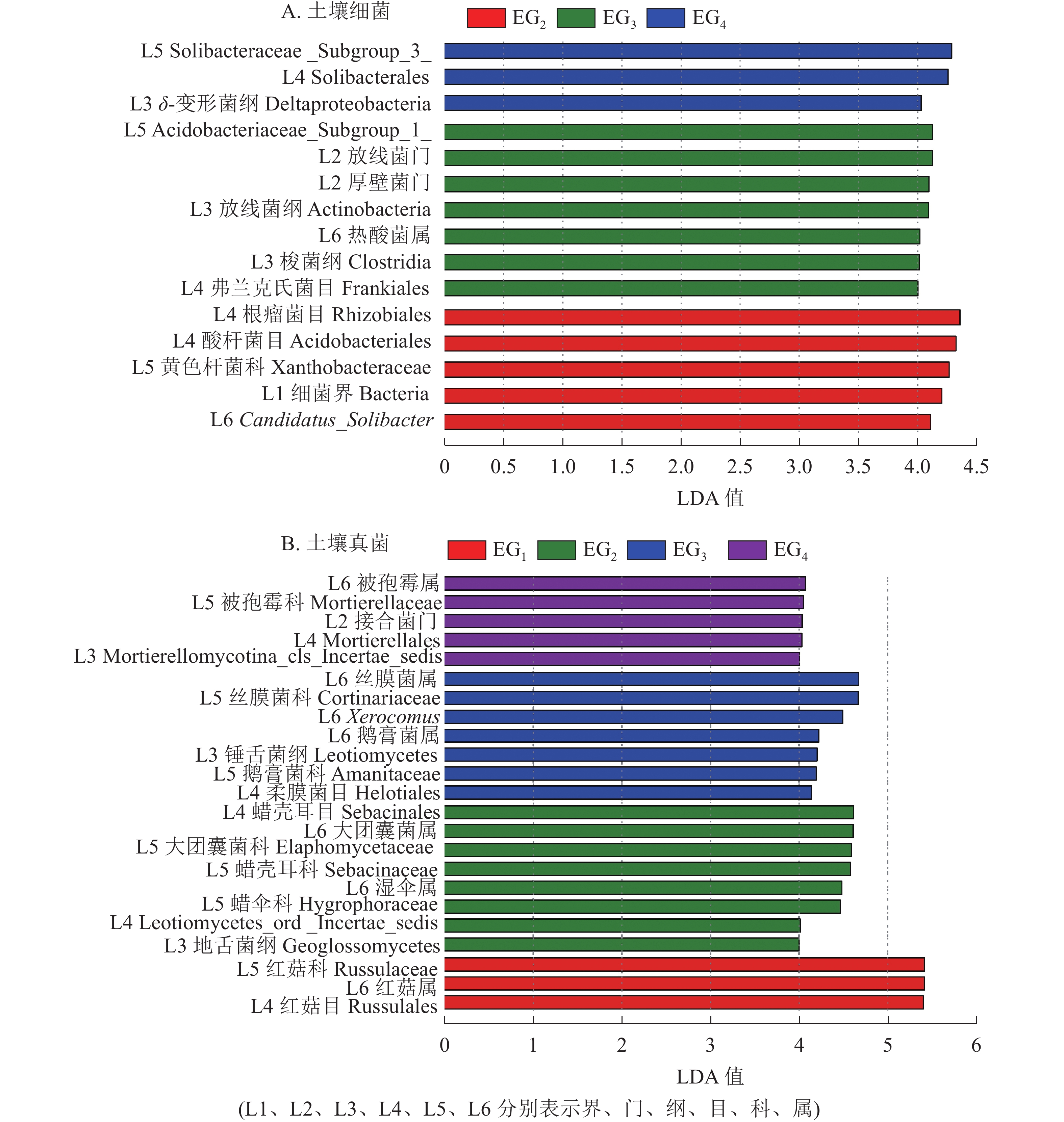
为进一步了解不同海拔土壤微生物组成的差异性,对不同分类水平上差异类群进行分析。除去未鉴定的物种,根据LefSe的结果显示:土壤细菌群落共有15个差异类群。其中1 364 m具有最多的差异类群,包括厚壁菌门、 放线菌门、放线菌纲Actinobacteria、梭菌纲Clostridia、热酸菌属等类群。765 m处存在5个差异类群,包括细菌界Bacteria、Candidatus_Solibacter等类群。δ-变形菌纲Deltaproteobacteria是1 611 m的差异类群(图4A)。
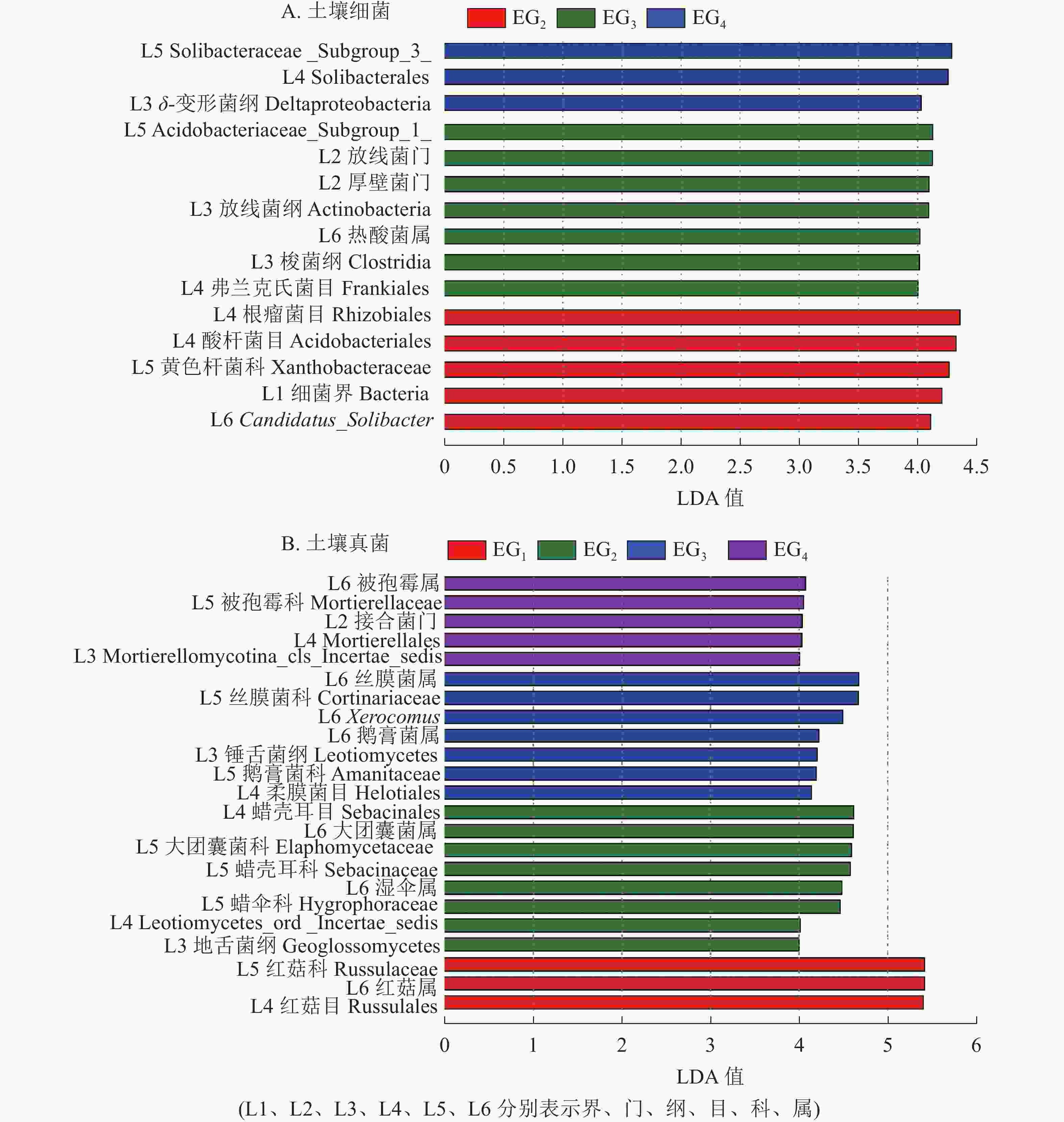
图 4 不同海拔土壤细菌(A)和真菌(B)群落LefSe分析
Figure 4. LefSe analysis showing the significant differences at different bacteria (A) and fungi (B) taxonomic levels
土壤真菌群落共有23个差异类群。343 m处具有最少的差异类群,包括红菇目Russulales、红菇科Russulaceae、红菇属。765 m包括地舌菌纲Geoglossomyeetes、湿伞属、大团囊菌属等差异类群;1 364 m包括锤舌菌纲Leotiomyeetes、鹅膏菌属、Xerocomus、丝膜菌属等差异类群。1 611 m包括接合菌门、被孢霉属等差异类群(图4B)。
-
采用主坐标分析(PCoA)法,基于杰卡德距离(binaryjaccard)算法,根据抽平后的OTU丰度分析不同海拔间微生物群落结构差异性,以二维图的形式呈现。由图5A显示:海拔引起了微生物群落的变化(P<0.05)。在土壤细菌群落中,同一海拔高度内样本间的细菌群落组成相对接近。其次,中低海拔细菌群落高度相似位于第1主轴(PC1)的负轴上,高海拔细菌群落位于PC1正轴上。
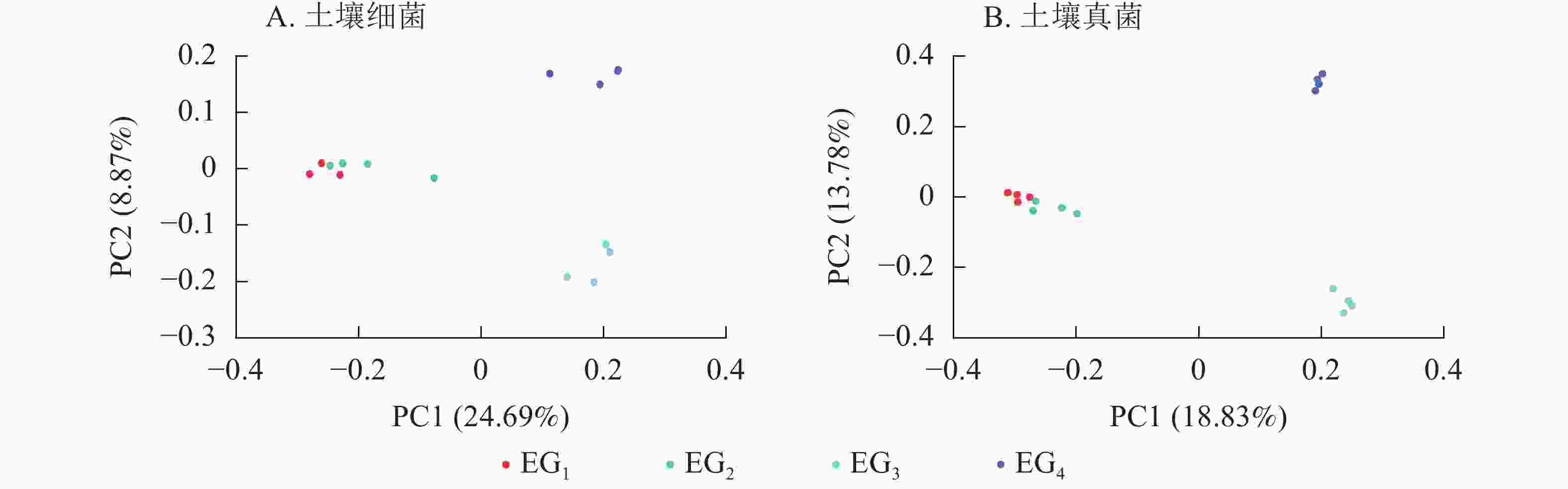
图 5 不同海拔OTU水平上土壤细菌(A)和真菌(B)群落PCoA分析
Figure 5. PCoA analysis showing the first two principal coordinates at different altitudes in bacterial (A) and fungal (B) communities
对土壤真菌群落的PCoA分析中显示了与细菌群落基本一致的结果(图5B),343与765 m各样本集中于PC1轴的负轴上,1 364与1 611 m各样本集中于PC1正轴上,343与765 m真菌群落相似度高,高海拔与中低海拔微生物多样性相似度低。
-
将土壤理化性质指标与微生物多样性、微生物群落结构[各样点在PC1、第2主轴(PC2)上的得分]进行Pearson相关性分析。结果(表6)表明:Chao 1指数与温度呈显著正相关(细菌的相关系数为0.886,真菌的相关系数为0.603,P<0.05)。微生物群落结构的相关性分析发现,PC1轴与温度相关性最强(细菌的相关系数为−0.944,真菌的相关系数为−0.939),呈显著负相关(P<0.05)。同时,细菌、真菌群落的PC1轴也与土壤全磷、全钾、pH显著相关(P<0.05)。
表 6 土壤理化性质与土壤微生物的Pearson相关性分析
Table 6. Pearson correlation between soil physical and chemical properties and soil microorganism
指标 细菌多样性 细菌群落结构 真菌多样性 真菌群落结构 Chao 1 Shannon PC1 PC2 Chao 1 Shannon PC1 PC2 温度 0.886* 0.320 −0.944* −0.190 0.603* −0.377 −0.939* −0.241 总碳 −0.041 −0.208 −0.010 0.057 0.048 0.242 −0.026 0.033 全氮 −0.412 −0.415 0.380 0.262 −0.215 0.478 0.349 0.249 全磷 −0.588* −0.394 0.618* 0.485 −0.448 0.435 0.581* 0.498 全钾 0.719* 0.427 −0.656* 0.325 0.207 −0.555 −0.671* 0.316 土壤湿度 −0.560 −0.360 0.600* 0.513 −0.461 0.382 0.562 0.530 pH 0.527 −0.016 −0.585* 0.176 0.343 −0.026 −0.616* 0.121 说明:*表示在0.05水平(双侧)上显著相关 -
本研究通过高通量测序对浙江凤阳山不同海拔常绿阔叶林的细菌和真菌群落进行调查和分析,以探究海拔对土壤微生物分布的影响。α多样性分析结果显示:海拔343 m细菌群落具有较高的丰富度指数,随着海拔的上升,丰富度指数下降,高海拔地区细菌群落丰富度指数显著低于低海拔地区,但多样性指数在低海拔与高海拔间并无显著差异。细菌群落的丰富度受海拔形成的地理梯度影响较大。SHEN等[30]在研究长白山不同海拔冻原生态系统土壤细菌群落的多样性格局时发现:细菌群落丰富度随海拔上升线性下降,安前东等[31]在西藏的研究也得出了一致的结果。海拔与年平均气温和年降水量高度相关,因此土壤细菌群落受气候的影响大于受地球化学或土壤结构因素的影响[32]。尽管如此,也不能排除其他因素对细菌丰富度的影响,比如土壤碳氮含量、pH等土壤理化因子可能是造成这种差异性格局的主要因素[30-31]。本研究中,温度与丰富度指数相关性系数为0.886,是导致细菌群落丰富度沿海拔垂直分布的主要影响因子,而全磷、全钾也与细菌丰富度指数有一定的联系。此外,在部分研究中,并未发现细菌群落丰富度的垂直梯度模式。如贺婧等[33]发现:太白山与朱雀山北坡的土壤细菌群落Chao 1指数无明显规律。与细菌群落不同的是,真菌群落丰富度指数虽然存在沿海拔上升而下降的趋势,但只在1 611 m处显著下降,因此真菌群落丰富度指数与温度的相关性弱于细菌群落。目前,大部分研究集中于对土壤细菌群落的海拔格局研究,关于真菌群落的相关报道较少。WANG等[34]发现:土壤真菌丰富度与海拔呈显著负相关。李敏等[35]研究指出:真菌丰富度指数与多样性指数在海拔1500 m时较低,但在海拔1800 m时较高,表明海拔对真菌多样性格局的影响尚不明确。
除土壤微生物群落的多样性格局,本研究还关注群落中特定类群的变化。门水平上,细菌群落以酸杆菌门、变形菌门和放线菌门为主。这些类群在植物多糖物质降解过程中起到了关键性作用[36]。属水平上,凤阳山常绿阔叶林的优势属为Candidatus_Solibacter、Bryobacter、Acidibacter。其中,Candidatus_Solibacter与Bryobacter能够有效地利用碳源,与有机质相关[37]。在不同的分类水平上,不同海拔细菌群落相对丰度存在显著差异。LefSe分析显示:细菌群落在1 364 m具有最多的差异类群,343 m处不存在差异类群。细菌群落某一特定类群丰度并没有在343 m显著增加,但对于一些类群,如硝化螺旋菌门、Acidibacter等,与海拔呈显著负相关(P<0.05)。这些个体类群与海拔之间的关系可能有助于整体细菌群落的海拔格局的形成[30]。真菌群落的优势菌群为担子菌门与子囊菌门。两者在刘子涵等[38]、薛晓敏等[39]的研究中也具有较高的丰度,是全球范围内最主要的土壤真菌门[14]。作为腐生菌,担子菌门和子囊菌门能够有效地分解植物残体[40]。根据LefSe的分析结果,虽然真菌群落比细菌群落具有更多差异类群,但是差异类群集中在分类水平较低的科、属上,且属水平上的相关性分析显示仅有2个属呈现垂直梯度的变化,因此真菌群落组成差异较大但具体类群沿海拔梯度垂直分布的特征不明显。
PCoA分析表明:无论是细菌群落还是真菌群落,海拔变化引起了土壤微生物群落组成的变化。343与765 m群落结构相似,且中低海拔与高海拔间存在明显分界,这与凤阳山土壤类型分界一致。厉桂香等[15]将其归结为微生物群落具有海拔分异性。本研究结果表明:343与765 m处具有更多共享的OTU数目,从另一角度佐证了微生物群落的海拔分异性特征。细菌和真菌群落结构在PC1轴上与温度相关性最强。由于海拔343 m处土壤理化性质与其他海拔差异较大,当去除343 m的数据进一步做相关性分析发现,PC1轴上,温度的影响并没有改变,但土壤理化性质与PC1轴的相关性明显减弱,土壤全磷、全钾、湿度、pH不再与PC1轴显著相关;PC2轴上,土壤理化性质的影响进一步增强,土壤全磷、全钾、湿度从弱相关变为显著相关(去除343 m后,细菌群落全磷、全钾、湿度与PC2轴相关系数绝对值均高于0.710,真菌群落绝对值均高于0.678)。这说明土壤微生物群落结构在PC1轴上主要受到温度的调控,而在PC2轴上由土壤理化性质起主导作用。在增温实验中,也同样验证了温度对微生物群落结构的重要性[41]。因此,温度是影响土壤微生物群落结构变化的主要因子,其次为土壤理化性质。
-
浙江凤阳山常绿阔叶林土壤微生物群落存在沿海拔梯度的变化规律,丰富度指数与相关性分析表明,细菌群落比真菌群落的垂直分布格局更明显。尽管真菌群落OTU数量少于细菌群落,但真菌群落具有更多的差异类群。此外,微生物的海拔效应表明,温度变化对土壤微生物群落空间结构的形成起着重要作用。
Soil microbial characteristics of evergreen broad-leaved forest at different altitudes in Fengyang Mountain, Zhejiang Province
-
摘要:
目的 探明浙江凤阳山地带性植被常绿阔叶林土壤微生物群落特征,明确土壤微生物群落沿海拔梯度的变化规律及影响微生物群落结构和多样性的主要因子。 方法 采集海拔343、765、1 364、1 611 m处土壤样品,利用高通量测序技术,探究土壤微生物群落与海拔的关系。 结果 凤阳山细菌操作分类单元 (OTU)数量多于真菌,中低海拔(343和765 m)共有的OTU数目较多;Chao 1指数随海拔上升呈下降趋势,Shannon指数没有明显的变化规律。在门水平上,细菌群落优势类群为酸杆菌门Acidobacteria (43.77%~51.55%)、变形菌门Proteobacteria (31.18%~35.77%)和放线菌门Actinobacteria (5.24%~7.99%);真菌群落优势类群为担子菌门Basidiomycota (33.16%~67.35%)和子囊菌门Ascomycota(22.98%~46.78%)。相对丰度前10 位的细菌门中,芽单胞菌门Gemmatimonadetes、硝化螺旋菌门Nitrospirae、疣微菌门Verrucomicrobia与海拔呈极显著负相关(P<0.01)。真菌群落不存在与海拔相关的微生物门,而LefSe线性判别分析结果显示:真菌群落具有更多的差异类群。此外,主坐标分析显示:土壤微生物群落以765 m为界,存在海拔分异性特征,且第1主轴与温度、土壤全磷、土壤全钾、土壤pH显著相关(P<0.05)。 结论 海拔变化引起了凤阳山土壤微生物群落特征的变化,并且温度是最主要的驱动因子。图5表6参41 Abstract:Objective The objective is to explore the characteristics of soil microbial community of zonal vegetation in evergreen broad-leaved forest in Fengyang Mountain, Zhejiang Province, and to clarify the changes of soil microbial community along the elevation gradient and the main factors affecting structure and diversity of microbial community. Method Soil samples were collected at 343, 765, 1364 and 1611 m above sea level. High-throughput sequencing technology was used to explore the relationship between soil microbial community and altitude. Result The number of OTU of bacteria was more than that of fungi, and middle and low altitudes (343 and 765 m) displayed more OTUs. Chao 1 index decreased with the increase of altitude, while Shannon index had no obvious trend. The dominant taxa of bacteria at the phylum level were Acidobacteria (43.77%−51.55%), Proteobacteria (31.18%−35.77%) and Actinobacteria, (5.24%−7.99%), while the dominant groups of fungal community were Basidiomycota (33.16%−67.35%) and Ascomycota (22.98%−46.78%). Among the top 10 bacterial phyla in relative abundance, Gemmatimonadetes, Nitrospirae and Verrucomicrobia were significantly negatively correlated with altitude (P<0.01). There were no altitudinal taxa in the fungal community at the phylum level. LefSe (LDA Effect Size) analysis exhibited more different taxa in the fungal community. In addition, PCoA showed that the soil microbial community had the characteristics of altitudinal differentiation bounded by 765 m, and the first axis of this PcoA (PC1) was significantly correlated with temperature, total phosphorus, total kalium and pH(P<0.05). Conclusion The change of altitude leads to the change of soil microbial community characteristics in Fengyang Mountain, and temperature is the main driving factor. [Ch, 5 fig. 6 tab. 41 ref.] -
Key words:
- soil bacteria /
- soil fungi /
- altitude /
- high throughput sequencing /
- Fengyang Mountain
-
图 1 不同海拔土壤细菌(A)和真菌(B)群落维恩图
Figure 1 Venn diagram showing the unique and shared OTUs at different elevations in bacterial (A) and fungal (B) communities
图 2 不同海拔门水平上土壤细菌(A)和真菌(B)群落相对丰度
Figure 2 Relative abundance of bacteria (A) and fungi (B) phylum at different altitudes
图 3 不同海拔属水平上土壤细菌(A)和真菌(B)群落相对丰度
Figure 3 Relative abundance of bacteria (A) and fungi (B) genus at different altitudes
图 4 不同海拔土壤细菌(A)和真菌(B)群落LefSe分析
Figure 4 LefSe analysis showing the significant differences at different bacteria (A) and fungi (B) taxonomic levels
图 5 不同海拔OTU水平上土壤细菌(A)和真菌(B)群落PCoA分析
Figure 5 PCoA analysis showing the first two principal coordinates at different altitudes in bacterial (A) and fungal (B) communities
表 1 不同海拔林分特征
Table 1. General Situation of the trees at different altitudes
海拔编号 树种组成 郁闭度/% EG1 木荷Schima superba、甜槠Castanopsis eyrei、青冈Cyclobalanopsis glauca、石栎Lithocarpus glaber、
檵木Loropetalum chinense等91 EG2 木荷、甜槠、青冈、檵木、山鸡椒Litsea cubeba等 87 EG3 木荷、甜槠、青冈、杨桐Cleyera japonica、马银花Rhododendron ovatum等 92 EG4 甜槠、青冈、马银花、尖叶山茶Camellia cuspidata、麂角杜鹃Rhododendron latoucheae等 90  下载: 导出CSV
下载: 导出CSV
表 2 不同海拔土壤理化性质
Table 2. Physical and chemical properties of soil at different altitudes
海拔编号 pH 土壤湿度/% 总碳/(mg·g−1) 全氮/(mg·g−1) 全磷/(mg·g−1) 全钾/(mg·g−1) EG1 4.86±0.03 a 15.54±6.41 c 103.97±7.60 a 3.57±0.35 b 0.21±0.04 c 19.63±2.58 a EG2 4.93±0.18 a 43.45±19.89 b 160.53±96.86 a 9.43±4.65 a 0.51±0.12 b 14.13±1.37 b EG3 4.67±0.07 a 38.41±7.81 bc 117.23±25.94 a 7.13±1.12 ab 0.45±0.16 b 11.43±1.03 b EG4 4.73±0.21 a 71.36±14.42 a 125.97±45.76 a 9.63±3.09 a 0.73±0.05 a 14.37±1.58 b 说明:数值为平均值±标准差(n=3),不同小写字母表示不同海拔间差异显著(P<0.05)
下载: 导出CSV
表 3 不同海拔土壤微生物群落多样性
Table 3. Soil bacterial diversity indices at different altitudes
细菌 真菌 海拔编号 Chao 1指数 Shannon指数 覆盖度 海拔编号 Chao 1指数 Shannon指数 覆盖度 EG1 4506.81±41.85 a 9.2622±0.246 8 a 0.9773±0.000 6 b EG1 783.12±36.26 a 3.6031±0.6633 b 0.9911±0.0006 b EG2 4255.62±295.74 b 8.8781±0.214 4 b 0.9780±0.001 7 b EG2 807.96±115.36 a 5.0242±0.6876 a 0.9911±0.0013 b EG3 3688.80±100.61 c 9.0043±0.231 9 ab 0.9824±0.000 5 a EG3 729.86±52.44 ab 4.6790±0.4236 a 0.9921±0.0002 ab EG4 3791.13±110.41 c 8.9974±0.176 5 ab 0.9815±0.000 8 a EG4 653.00±67.50 b 4.5171±0.6495 ab 0.9928±0.0008 a 说明:数值为平均值±标准差(n = 4),不同小写字母表示不同海拔间差异显著(P<0.05)
下载: 导出CSV
表 4 海拔与微生物门Pearson相关性系数
Table 4. Pearson correlation coefficient between altitude and soil bacterial phylum
细菌门 相关性系数 真菌门 相关性系数 细菌门 相关性系数 真菌门 相关性系数 酸杆菌门 0.032 担子菌门 −0.402 芽单胞菌门 −0.817** 壶菌门 −0.209 变形菌门 0.234 子囊菌门 0.173 迷踪菌门 −0.260 Cercozoa −0.191 放线菌门 −0.417 接合菌门 0.437 髌骨细菌门 −0.490 新丽鞭菌门 −0.233 拟杆菌门 0.269 罗兹菌门 −0.308 硝化螺旋菌门 −0.723** 芽枝霉门 −0.236 厚壁菌门 0.525* 球囊菌门 −0.036 疣微菌门 −0.705** 说明:*表示在0.05水平(双侧)上显著相关;**表示在0.01水平(双侧)上显著相关
下载: 导出CSV
表 5 海拔与微生物属Pearson相关性系数
Table 5. Pearson correlation coefficient between altitude and soil bacterial genus
细菌属 真菌属 细菌属 真菌属 属名 相关系数 属名 相关系数 属名 相关系数 属名 相关系数 Candidatus_Solibacter −0.458 红菇属Russula −0.645** Granulicella 0.719** Sebacina 0.197 Bryobacter 0.088 Archaeorhizomyces 0.290 Acidipila 0.639** 鹅膏菌属Amanita 0.250 Acidibacter −0.520* 大团囊菌属Elaphomyces −0.234 芽单胞菌属Gemmatimonas −0.734** Rossbeevera −0.127 热酸菌属Acidothermus 0.514* Xerocomus 0.200 分枝杆菌属Mycobacterium 0.035 丝盖伞属Inocybe 0.190 Candidatus_Koribacter −0.178 丝膜菌属Cortinarius 0.182 鞘氨醇单胞菌属Sphingomonas −0.778** 地舌菌属Geoglossum −0.194 Burkholderia-Caballeronia-
Paraburkholderia0.648** 棉革菌属Tomentella −0.259 拟杆菌属Bacteroides 0.224 Lactarius 0.311 Clade_Ia 0.472 湿伞属Hygrocybe −0.097 MND1 −0.743** 粉褶菌属Entoloma −0.209 Pajaroellobacter 0.740** 被孢霉属Morlierella 0.702** 说明:*表示在0.05水平(双侧)上显著相关;**表示在0.01水平(双侧)上显著相关
下载: 导出CSV
表 6 土壤理化性质与土壤微生物的Pearson相关性分析
Table 6. Pearson correlation between soil physical and chemical properties and soil microorganism
指标 细菌多样性 细菌群落结构 真菌多样性 真菌群落结构 Chao 1 Shannon PC1 PC2 Chao 1 Shannon PC1 PC2 温度 0.886* 0.320 −0.944* −0.190 0.603* −0.377 −0.939* −0.241 总碳 −0.041 −0.208 −0.010 0.057 0.048 0.242 −0.026 0.033 全氮 −0.412 −0.415 0.380 0.262 −0.215 0.478 0.349 0.249 全磷 −0.588* −0.394 0.618* 0.485 −0.448 0.435 0.581* 0.498 全钾 0.719* 0.427 −0.656* 0.325 0.207 −0.555 −0.671* 0.316 土壤湿度 −0.560 −0.360 0.600* 0.513 −0.461 0.382 0.562 0.530 pH 0.527 −0.016 −0.585* 0.176 0.343 −0.026 −0.616* 0.121 说明:*表示在0.05水平(双侧)上显著相关
下载: 导出CSV
-
[1] CHANG E H, CHEN T H, TIAN G L, et al. The effect of altitudinal gradient on soil microbial community activity and structure in moso bamboo plantations [J]. Appl Soil Ecol, 2016, 98: 213 − 220. [2] MA Yuhua, FENG Chun, WANG Zhaocheng, et al. Restoration in degraded subtropical broadleaved forests induces changes in soil bacterial communities [J/OL]. Global Ecol Conserv, 2021, 30: e01775[2021-11-20]. doi: 10.1016/j.gecco.2021.e01775. [3] SHAOPengshuai, LIANG Chao, RUBERT-NASON K, et al. Secondary successional forests undergo tightly-coupled changes in soil microbial community structure and soil organic matter [J]. Soil Biol Biochem, 2019, 128: 56 − 65. [4] NELSON M B, MARTINY A C, MARTINY J B H. Global biogeography of microbial nitrogen-cycling traits in soil [J]. Proc Natl Acad Sci, 2016, 113(29): 8033 − 8040. [5] WARING B G, AVERILL C, HAWKES C V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: insights from meta-analysis and theoretical models [J]. Ecol Lett, 2013, 16(7): 887 − 894. [6] SUN Junming, IRZYKOWSKI W, JEDRYCZKA M, et al. Analysis of the genetic structure of Sclerotinia sclerotiorum (Lib. ) de Bary populations from different regions and host plants by random amplified polymorphic DNA markers [J]. J Integr Plant Biol, 2005, 47(4): 385 − 395. [7] ŽIFČÁKOVÁ L, VĚTROVSKÝ T, HOWE A, et al. Microbial activity in forest soil reflects the changes in ecosystem properties between summer and winter [J]. Environ Microbiol, 2016, 18(1): 288 − 301. [8] FRAC M, HANNULA S E, BELKA M, et al. Fungal biodiversity and their role in soil health [J/OL]. Front Microbiol, 2018, 9: 707[2021-11-20]. doi:10.3389/fmicb.2018.00707. [9] MARGESIN R, NIKLINSKA M A. Editorial: elevation gradients: microbial indicators of climate change? [J/OL]. Front Microbiol, 2019, 10: 2405[2021-11-20]. doi:10.3389/fmicb.2019.02405. [10] SILES J A, CAJTHAML T, FILIPOVÁ A, et al. Altitudinal, seasonal and interannual shifts in microbial communities and chemical composition of soil organic matter in Alpine forest soils [J]. Soil Biol Biochem, 2017, 112: 1 − 13. [11] WU Jiejun, ANDERSON B J, BUCKLEY H L, et al. Aspect has a greater impact on alpine soil bacterial community structure than elevation [J/OL]. FEMS Microbiol Ecol, 2017, 93(5): fiw253[2021-11-20]. doi: 10.1093/femsec/fix032. [12] COLLINS H P, CAVIGELLI M A. Soil microbial community characteristics along an elevation gradient in the Laguna Mountains of Southern California [J]. Soil Biol Biochem, 2003, 35(8): 1027 − 1037. [13] LOOBY C I, MARTIN P H. Diversity and function of soil microbes on montane gradients: the state of knowledge in a changing world [J/OL]. FEMS Microbiol Ecol, 2020, 96(9): fiaa122[2021-11-20]. doi: 10.1093/femsec/fiaa122. [14] 周煜杰, 贾夏, 赵永华, 等. 森林生态系统土壤真菌群落及其影响因素研究进展[J]. 生态环境学报, 2020, 29(8): 1703 − 1712. ZHOU Yujie, JIA Xia, ZHAO Yonghua, et al. A review on soil fungal community and its affectingfactors in forest ecosystem [J]. Ecol Environ Sci, 2020, 29(8): 1703 − 1712. [15] 厉桂香, 马克明. 土壤微生物多样性海拔格局研究进展[J]. 生态学报, 2018, 38(5): 1521 − 1529. LI Guixiang, MA Keming. Progress in the study of elevational patterns of soil microbial diversity [J]. Acta Ecol Sin, 2018, 38(5): 1521 − 1529. [16] 赵盼盼, 周嘉聪, 林开淼, 等. 海拔梯度变化对中亚热带黄山松土壤微生物生物量和群落结构的影响[J]. 生态学报, 2019, 39(6): 2215 − 2225. ZHAO Panpan, ZHOU Jiacong, LIN Kaimiao, et al. Effect of different altitudes on soil microbial biomass and community structure of Pinus taiwanensis forest in mid-subtropical zone [J]. Acta Ecol Sin, 2019, 39(6): 2215 − 2225. [17] 孟苗婧, 郭晓平, 张金池, 等. 海拔变化对凤阳山针阔混交林地土壤微生物群落的影响[J]. 生态学报, 2018, 38(19): 7057 − 7065. MENG Miaojing, GUO Xiaoping, ZHANG Jinchi, et al. Effects of altitude on soil microbial community in Fengyang Mountain coniferous and broad-leaved forest [J]. Acta Ecol Sin, 2018, 38(19): 7057 − 7065. [18] LI Guixiang, XU Guorui, SHEN Congcong, et al. Contrasting elevational diversity patterns for soil bacteria between two ecosystems divided by the treeline [J]. Sci China Life Sci, 2016, 59(11): 1177 − 1186. [19] DAI Zhongmin, ZANG Huadong, CHEN Jie, et al. Metagenomic insights into soil microbial communities involved in carbon cycling along an elevation climosequences [J]. Environ Microbiol, 2021, 23(8): 4631 − 4645. [20] 金裕华. 武夷山不同海拔土壤微生物多样性的变化特征[D]. 南京: 南京林业大学, 2012. JIN Yuhua. Variations of Soil Microbial Diversity along an Elevation Gradient in the Wuyi Mountains [D]. Nanjing: Nanjing Forestry University, 2012. [21] 丁炳扬, 陈根荣, 程秋波, 等. 浙江凤阳山自然保护区种子植物区系的统计分析[J]. 云南植物研究, 2000, 22(1): 27 − 37. DING Bingyang, CHEN Genrong, CHENG Qiubo, et al. A floristic statistics and analyses of seed plants of Fengyangshan Nature Reserve in Zhejiang Province [J]. Acta Bot Yunnan, 2000, 22(1): 27 − 37. [22] 徐筱芃. 浙江凤阳山常绿阔叶林树种多样性及其影响因素研究[D]. 南京: 南京林业大学, 2017. XU Xiaopeng. Study on Plant Diversity and Its Influence Factors of the Evergreen Broad-leaved Forest in Fengyang Mountain [D]. Nanjing: Nanjing Forestry University, 2017. [23] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Rukun. The Analysis Method of Soil Agricultural Chemistry[M]. Beijing: China Agricultural Science and Technology Press, 2000. [24] BOLGER A M, LOHSE M, USADEL B. Trimmomatic: a flexible trimmer for Illumina sequence data [J]. Bioinformatics, 2014, 30(15): 2114 − 2120. [25] MAGOČ T, SALZBERG S L. FLASH: fast length adjustment of short reads to improve genome assemblies [J]. Bioinformatics, 2011, 27(21): 2957 − 2963. [26] CAPORASO J G, KUCZYNSKI J, STOMBAUGH J, et al. QIIME allows analysis of high-throughput community sequencing data [J]. Nat Methods, 2010, 7(5): 335 − 336. [27] EDGAR R C, HAAS B J, CLEMENTE J C, et al. UCHIME improves sensitivity and speed of chimera detection [J]. Bioinformatics, 2011, 27(16): 2194 − 2200. [28] ROGNES T, FLOU T, NICHOLS B, et al. VSEARCH: a versatile open source tool for metagenomics [J/OL]. Peer J, 2016, 4: 2409v1[2021-11-20]. doi: 10.7287/peerj. preprints. 2409v1. [29] SEGATA N, IZARD J, WALDRON L, et al. Metagenomic biomarker discovery and explanation [J/OL]. Genome Biol, 2011, 12(6): R60 [2021-11-20]. https://doi.org/10.1186/gb-2011-12-6-r60. [30] SHEN Congcong, NI Yingying, LIANG Wenjun, et al. Distinct soil bacterial communities along a small-scale elevational gradient in alpine tundra [J/OL]. Front Microbiol, 2015, 6: 582[2021-11-20]. doi: 10.3389/fmicb.2015.00582. [31] 安前东, 徐梦, 张旭博, 等. 西藏色季拉山垂直植被带土壤细菌群落组成及功能潜势[J]. 应用生态学报, 2021, 32(6): 2147 − 2157. AN Qiandong, XU Meng, ZHANG Xubo, et al. Soil bacterial community composition and functional potentials along the vertical vegetation transect on Mount Segrila, Tibet, China [J]. Chin J Appl Ecol, 2021, 32(6): 2147 − 2157. [32] SINGH D, LEE-CRUZ L, KIM W S, et al. Strong elevational trends in soil bacterial community composition on Mt. Halla, South Korea [J]. Soil Biol Biochem, 2014, 68: 140 − 149. [33] 贺婧, 闫冰, 李俊生, 等. 秦岭中段北坡不同海拔土壤中细菌群落的分布特征及区域差异比较[J]. 环境科学研究, 2019, 32(8): 1374 − 1383. HE Jing, YAN Bing, LI Junsheng, et al. Altitude distribution patterns and regional differences of soil bacterial community in northern slopes in the Middle Qinling Mountains [J]. Res Environ Sci, 2019, 32(8): 1374 − 1383. [34] WANG Juntao, ZHENG Yuanming, HU Hangwei, et al. Soil pH determines the alpha diversity but not beta diversity of soil fungal community along altitude in a typical Tibetan forest ecosystem [J]. J Soil Sediment, 2015, 15(5): 1224 − 1232. [35] 李敏, 闫伟. 海拔对乌拉山油松根围真菌群落结构的影响[J]. 菌物学报, 2019, 38(11): 1992 − 2006. LI Min, YAN Wei. Effects of altitude on rhizosphere fungal community structure of Pinus tabulaeformis in Wula Mountain, China [J]. Mycosystema, 2019, 38(11): 1992 − 2006. [36] KANOKRATANA P, UENGWETWANIT T, RATTANACHOMSRI U, et al. Insights into the phylogeny and metabolic potential of a primary tropical peat swamp forest microbial community by metagenomic analysis [J]. Microb Ecol, 2011, 61(3): 518 − 528. [37] 杜思瑶, 于淼, 刘芳华, 等. 设施种植模式对土壤细菌多样性及群落结构的影响[J]. 中国生态农业学报, 2017, 25(11): 1615 − 1625. DU Siyao, YU Miao, LIU Fanghua, et al. Effect of facility management regimes on soil bacterial diversity and community structure [J]. Chin J Eco-Agric, 2017, 25(11): 1615 − 1625. [38] 刘子涵, 黄方园, 黎景来, 等. 覆盖模式对旱作农田土壤微生物多样性及群落结构的影响[J]. 生态学报, 2021, 41(7): 2750 − 2760. LIU Zihan, HUANG Fangyuan, LI Jinglai, et al. Effects of farmland and mulching patterns on soil microbial diversity and community structure in dryland [J]. Acta Ecol Sin, 2021, 41(7): 2750 − 2760. [39] 薛晓敏, 王来平, 韩雪平, 等. 不同树盘覆盖对矮砧苹果园土壤微生物群落结构和多样性的影响[J]. 生态学报, 2021, 41(4): 1528 − 1536. XUE Xiaomun, WANG Laiping, HAN Xueping, et al. Effects of different tree disk mulching on soil microbial community structure and diversity in dwarfing rootstock apple orchard [J]. Acta Ecol Sin, 2021, 41(4): 1528 − 1536. [40] BASTIDA F, TORRES I F, MORENO J L, et al. The active microbial diversity drives ecosystem multifunctionality and is physiologically related to carbon availability in Mediterranean semi-arid soils [J]. Mol Ecol, 2016, 25(18): 4660 − 4673. [41] 陈岳民, 高金涛, 熊德成, 等. 土壤增温对中亚热带杉木幼林土壤微生物群落结构和有效氮的影响[J]. 亚热带资源与环境学报, 2016, 11(4): 1 − 8. CHEN Yuemin, GAO Jintao, XIONG Decheng, et al. Effects of soil warming on soil microbial community structure and soil available nitrogen in subtropical young Chinese fir plantation [J]. J Subtrop Resour Environ, 2016, 11(4): 1 − 8. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210820
 点击查看大图
点击查看大图

计量
- 文章访问数: 1683
- HTML全文浏览量: 484
- PDF下载量: 79
- 被引次数: 0
