





-
松材线虫Bursaphelenchus xylophilus病在亚洲和欧洲造成严重的生态和经济损失[1-2],伊氏杀线虫真菌Esteya vermicola (EV菌)是松材线虫的内寄生真菌,产生的新月形孢子能侵染并杀死松材线虫,在松材线虫的生物防治方面具有良好的应用前景[3]。同时,EV菌亦可侵染拟松材线虫B. mucronatus、水稻干尖线虫Aphelenchoides besseyi等线虫。研究表明:EV真菌胞内存在共生细菌[4],共生细菌对真菌的生物学和生态学具有重要的作用,如Rhizopus microsporus胞内的共生细菌Burkholderia能产生植物毒素根霉素,然后由宿主真菌分泌到环境中,导致水稻枯萎病[5]。菌丝内的共生微生物(细菌或病毒)是重要的能直接或间接改变真菌和宿主关系的互作生物[6-8],提高宿主真菌的生态适应性[9-10]。存在共生细菌的真菌对碳氮源如何响应,其代谢产物发生何种变化,目前尚未见相关报道。
代谢组学是系统生物学研究中非常重要的一个环节,旨在研究生物体或组织,甚至单个细胞的全部小分子代谢物成分及其动态变化[11]。通过对代谢物质的分析,可以从生物样本中检测并筛选出具有重要生物学意义和显著差异的代谢物质,并以此为基础研究生物体的代谢过程和变化机制[12]。对EV菌在不同碳、氮培养条件下代谢物的变化进行分析,明确显著差异代谢物,特别是重要的信号分子,不仅为EV菌的应用提供重要理论基础,而且对深入研究内共生细菌在EV菌的功能亦具有重要意义。
-
伊氏杀线虫真菌Esteya vermicola CBS115803购于荷兰菌物保存中心。
-
碳培养基为24 g 马铃薯葡萄糖肉汤培养基(PDB,Becton, Dickinson and Company,美国)和10 g琼脂溶于1 L双蒸水。氮培养基为5 g酵母粉(OXOID公司,英国)和10 g琼脂溶于1 L双蒸水。
-
Nano高分辨液质联用分析仪QE (Thermo Q-Exactive,德国);赛多利斯BSA124S电子天平;真空浓缩仪(Labogene MaixVac Alpha, 丹麦);Sigma 3-30KS高速离心机,KQ5 200DE型数控超声波清洗器(昆山市超声仪器有限公司,中国);色谱级甲醇(Fisher,美国)。
-
配制碳、氮固体培养基,培养基凝固后放入已灭菌的尼龙膜(孔径3 μm,赛多利斯),接种EV菌后于25 ℃培养7 d。将长满菌丝和孢子的尼龙膜用无菌镊子取出,用超纯水冲洗尼龙膜,再用无菌的吸水纸吸掉尼龙膜接触培养基面的水分,而后将菌丝和孢子刮入2 mL圆底灭菌干燥离心管,装样品前后分别称量离心管的质量,迅速将样品管放入液氮中淬灭后于−80 ℃保存。每处理设置5次生物学重复,编号依次为C1~C5和N1~N5。加入冷冻洁净无菌的直径为3 mm的钢珠2个,液氮冷冻条件下使用莱驰RETSCH MM 400研磨2 min。将研磨好的样品管置于干冰上,加入2 mL预冷的体积分数为80%甲醇于−80 ℃冰箱静置1 h,4 ℃、14 000 g离心20 min后将上清液转移至另一相同体积离心管,真空浓缩干燥样品后于−80 ℃保存。用300 μL体积分数为90%甲醇超声复溶并经0.45 μm的滤膜过滤,完成亲水代谢产物提取。
-
采用非靶标的高效液相色谱-质谱联用技术(HPLC-MS)对样品进行测定和代谢物识别。色谱柱:Waters ACQUITY UPLC®(2.1 mm×100.0 mm,1.7 μm)。流动相:0.1%甲酸-水溶液(A),0.1%甲酸-乙腈(B)。洗脱条件:0~0.5 min,95%B;0.5~7.0 min,95%~65% B;7.0~8.0 min,65%~40% B;8.0~9.0 min,40%B;9.0~9.1 min,40%~95%B;9.1~12.0 min,95%B。质谱参数条件:鞘气,310 275 Pa;辅助气,103 425 Pa;喷雾电压,4 000 V (正)/3 500 V (负);离子传输管温度:350 ℃。在采集软件(Xcalibur 4.0.27,Thermo)的控制下,采用一级质谱全扫描(全MS)结合自动触发二级质谱扫描(MS/MS)模式,使用阴、阳离子模式2种电离方式采集质谱数据。运行时间:0~18 min。分辨率:全MS,170000;MS/MS,17500。
-
对原始数据进行基线过滤、峰识别、积分、保留时间校正、峰对齐和归一化,最终得到1个保留时间、质荷比和代谢物信号(峰面积)的数据矩阵。将处理后的代谢物信号与一级和二级数据库进行匹配,将其中二级得分<30的化合物可信予以剔除。分析时10份样品共插入3个质量对照(QC)样本以考察整个分析过程中仪器的稳定性,QC样本每种化合物的峰面积的变异系数若>30%,则予以剔除。未检出值设定为0,缺失值由其余重复样品的均值填补。
-
由Metaboanalyst 4.0在线软件(https://www.metaboanalyst.ca/MetaboAnalyst/home.xhtml)完成。利用对数转换和pareto缩放对各代谢物相对含量进行标准化处理。使用主成分分析法(PCA)和偏最小二乘法判别法(PLS-DA)评估不同处理的分组情况。结合以下2个标准对2种培养基的阴阳离子模式的差异化合物进行筛选:经FDR (false discovery rate)校正的t检验的P<0.05;正交偏最小二乘判别分析(OPLS-DA)中变量投影重要性(variable influence on projection,VIP)得分>1。
-
代谢通路分析由Metaboanalyst 4.0 (
https://www.metaboanalyst.ca/MetaboAnalyst/upload/PathUploadView.xhtml )中的代谢通路分析模块(pathway analysis module)完成。该分析将选定的差异代谢物与KEGG (Kyoto encyclopedia of genes and genomes)数据库(2019年10月)和人类代谢组数据库(HMDB)进行匹配,然后选择酵母Saccharomyces cerevisiae的代谢通路库作为参照进行富集分析和拓扑分析。富集分析中采用参数超几何法(hypergeometric test)检验每个代谢通路的显著性(P<0.05);拓扑分析中差异显著性检验的方法为相对介数中心性(relative-betweeness centrality)。 -
阳离子模式下鉴定到的化合物多于阴离子模式下鉴定的化合物。阴、阳离子模式分别检测出279和461种化合物,其中74种为2种模式共同检出,共计666种化合物。通过质量控制过滤处理,阴、阳离子模式分别有176和362种化合物,其中2种模式共同含有40种化合物,共计498种化合物。数据总的缺失值的比例平均为0.74%:阴离子模式下,碳和氮培养基下的代谢物缺失率均为0.23%;阳离子模式下,碳培养基下的缺失率为2.38%,高于氮培养基0.11%。
-
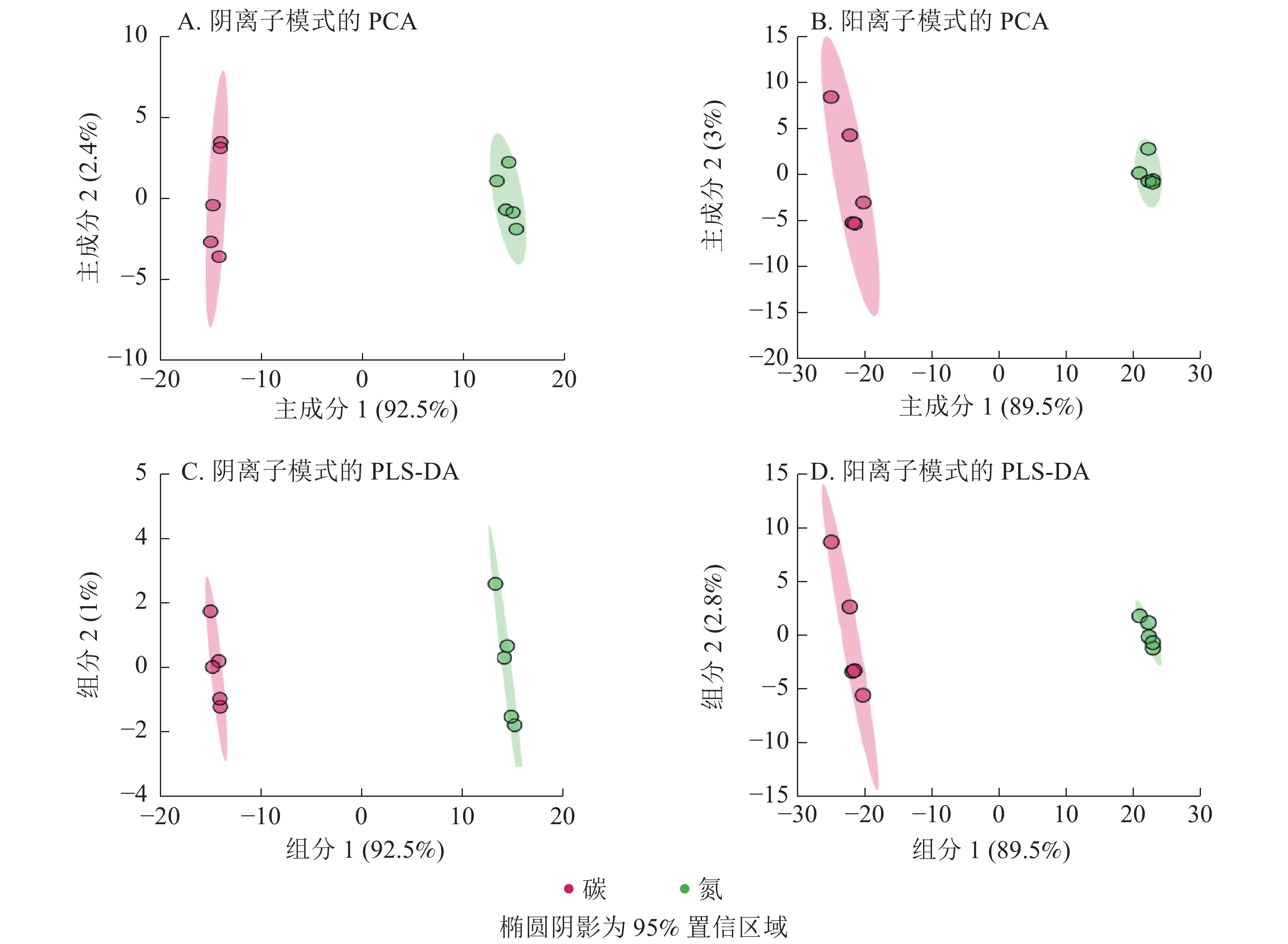
阴离子和阳离子模式下EV菌菌丝体代谢物的主成分分析得分图(图1A~B)显示:前2个主成分分别解释了94.9% (主成分1为92.5%,主成分2为2.4%)和92.5% (主成分1为89.5%,主成分2为3.0%)的变异。2种离子模式下EV菌代谢谱在碳、氮培养基培养后差异大,分组明显。偏最小二乘法判别法的分组结果(图1C~D)与主成分分析法结果一致。
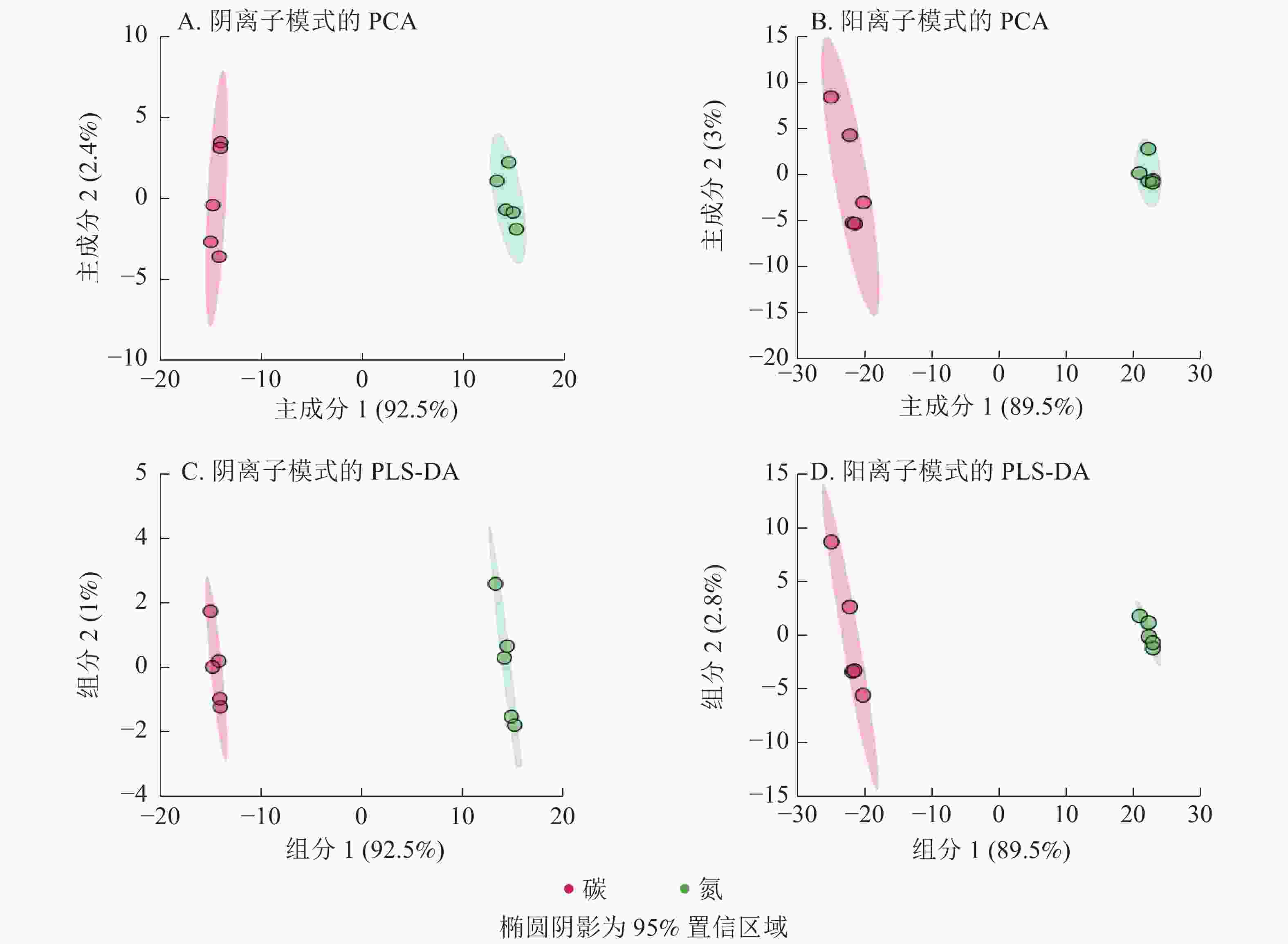
图 1 碳、氮培养条件下EV菌代谢物的PCA和PLS-DA分析
Figure 1. PCA and PLS-DA of EV metabolites under carbon and nitrogen culture conditions
-
采用火山图的形式进行差异化合物展示,见图2。t检验差异显著(P<0.05)的化合物共有444种,占总数的89.2%;阴离子和阳离子模式分别有162和310种,28种为2种模式共有。VIP>1的化合物共有469种,占总数的94.2%;阴离子和阳离子模式分别有167和334种,32种为2种模式共有。利用VIP>1筛选出的化合物包含t检验法的筛选结果,表明后者更严格,且更符合统计要求。
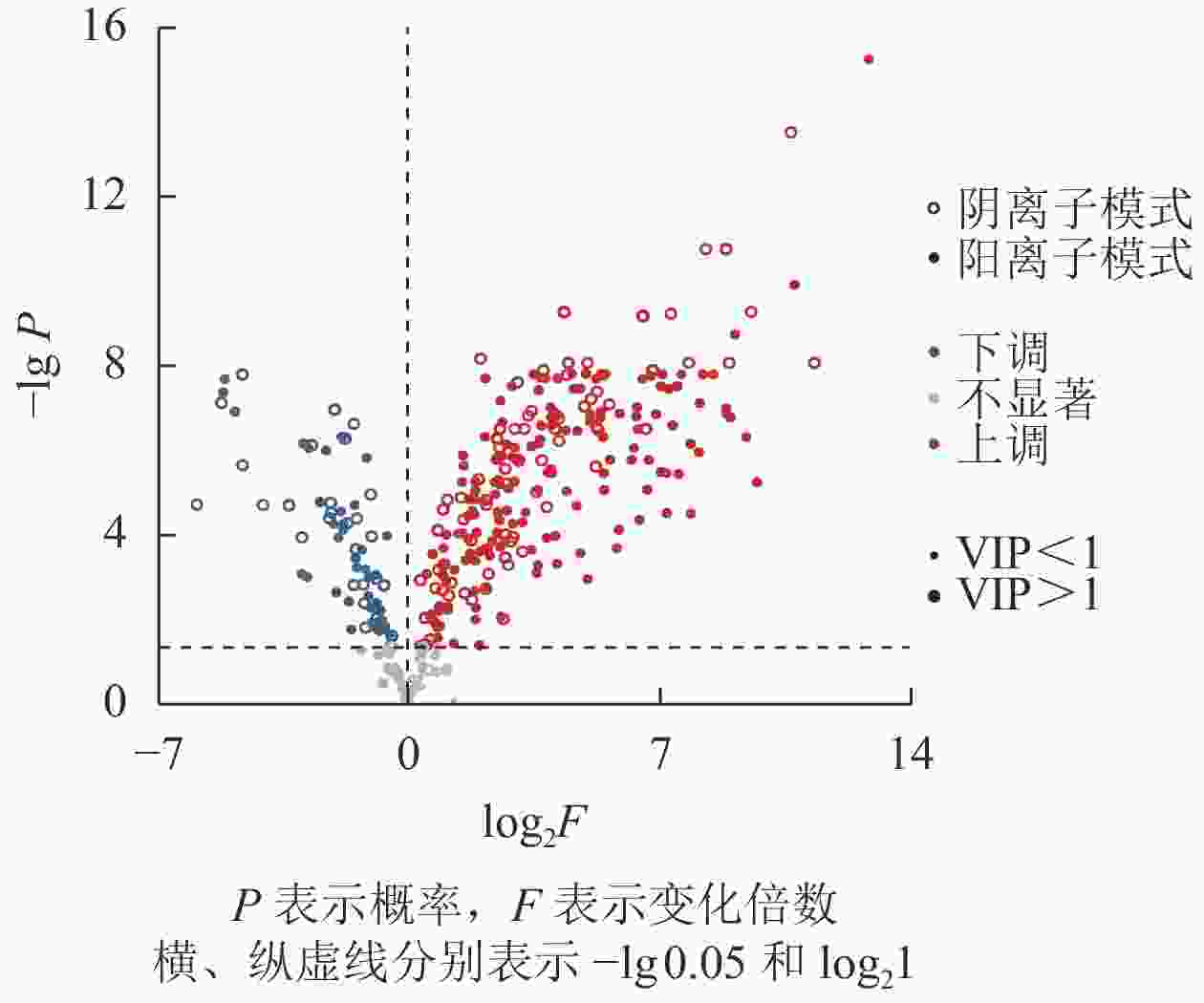
图 2 碳、氮培养条件下EV菌的差异代谢物
Figure 2. Differential metabolites of EV bacteria under carbon and nitrogen culture conditions
以碳为对照组,在氮组中上调代谢物的数量是下调的3.4倍,分别为342和102种;上调和下调代谢物分别有309和86种匹配到HMDB数据库,有159和71种匹配到KEGG数据库。磷酸胍基乙酸酯和对甲酚硫酸盐是在氮培养下大量产生的特有代谢物,尿囊素、光色素、吲哚和海藻糖等是代谢物在氮培养条件下显著上调(表1)。
表 1 部分差异化合物
Table 1. Part of significantly different metabolites
化合物名称 质荷比 保留时间/min 二级数据库得分 变化倍数 P KEGG编号 磷酸胍基乙酸酯 198.03 9.86 62 12.46 1.13×10−13 C03166 对甲酚硫酸盐 187.01 6.17 53 12.41 8.88×10−15 邻氨基苯甲酸酯 136.04 2.00 50 9.93 1.67×10−8 C00108 尿酸 190.05 1.29 40 8.26 6.26×10−7 C01717 尼古丁 163.12 4.68 44 5.51 1.14×10−6 C16150 4-羟基-2-喹啉羧酸 188.03 5.76 30 5.48 9.62×10−8 C01717 烟酸 124.04 3.50 41 5.17 1.71×10−7 C00253 尿囊素 159.05 5.46 50 5.09 2.53×10−4 C01551 吲哚丙烯酸 188.07 7.98 98 4.20 4.68×10−8 2-吡咯烷酮 86.06 3.79 41 4.10 2.57×10−5 C11118 咪唑乙酸 127.05 9.62 69 3.54 1.56×10−7 C02835 组胺 112.09 5.74 45 3.34 2.79×10−8 C00388 谷胱甘肽 306.08 1.02 58 3.26 5.97×10−4 C00051 光色素 243.09 1.58 70 3.00 2.79×10−4 C01727 肌肽 227.11 12.66 55 2.40 3.30×10−5 C00386 甜菜碱醛 102.09 6.73 68 2.25 9.68×10−3 C00576 吲哚 118.07 8.32 57 2.12 4.40×10−3 C00463 苹果酸 133.01 0.90 43 1.90 2.86×10−5 C00711 甜菜碱 118.09 8.30 51 1.88 2.79×10−8 C00719 胍基乙酸d 118.06 8.26 38 1.87 1.19×10−3 C00581 葫芦巴碱 138.06 9.05 38 1.59 1.71×10−7 C01004 海藻糖 341.11 1.04 78 1.34 6.16×10−5 C01083 左旋肉碱 162.11 9.31 72 1.34 2.97×10−6 C00318 邻乙酰左旋肉碱 204.12 8.54 73 1.32 1.69×10−6 C02571 茶碱 181.07 2.24 55 −1.64 6.07×10−7 C07130 二乙醇胺 106.09 8.16 45 −1.73 1.42×10−4 C06772 去甲肾上腺素 170.08 5.83 57 −2.40 1.52×10−5 C00547 阿魏酸盐 193.05 5.36 32 −3.58 2.87×10−5 C01494 核糖醇 151.06 1.01 46 −4.08 3.13×10−8 C00474 甲基咪唑乙酸 141.07 9.52 41 −4.51 2.90×10−8 C05828 赤藓糖醇 121.05 1.04 75 −5.20 2.80×10−5 C00503 -
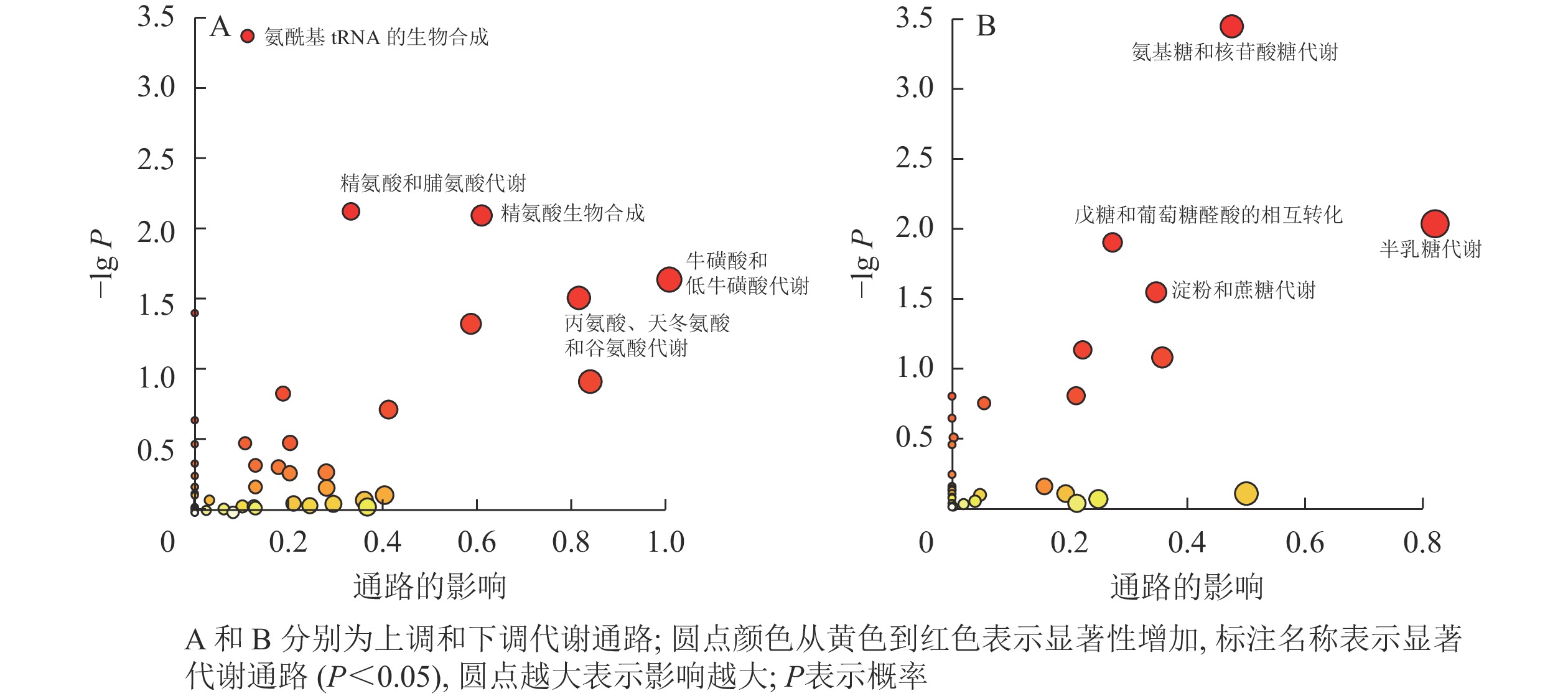
由表2和图3可见:利用KEGG数据库对氮培养中上调和下调的显著差异代谢产物进行富集分析。上调代谢产物主要富集到氨基酸代谢通路,包括氨酰基tRNA的生物合成,精氨酸和脯氨酸代谢,精氨酸生物合成,牛磺酸和低牛磺酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢。下调的显著差异代谢产物主要涉及糖类代谢通路,包括氨基糖和核苷酸糖代谢、半乳糖代谢、戊糖和葡萄糖醛酸酯的相互转化、淀粉和蔗糖代谢。
表 2 碳、氮培养条件下EV菌差异代谢物富集到的KEGG通路
Table 2. Enriched KEGG pathways by differential metabolites of EV under carbon and nitrogen culture conditions
代谢通路编号 代谢通路名称 匹配情况 P 影响大小 上调 sce00970 氨酰基tRNA的生物合成 18/46 4.38×10−4 0.11 sce00330 精氨酸和脯氨酸代谢 10/25 7.53×10−3 0.33 sce00220 精氨酸生物合成 8/18 8.05×10−3 0.60 sce00430 牛磺酸和低牛磺酸代谢 4/7 2.28×10−2 1.00 sce00250 丙氨酸、天冬氨酸和谷氨酸代谢 8/22 3.06×10−2 0.81 下调 sce00520 氨基糖和核苷酸糖代谢 8/24 3.41×10−4 0.48 sce00052 半乳糖代谢 5/17 8.98×10−3 0.82 sce00040 戊糖和葡萄糖醛酸的相互转化 4/12 1.22×10−2 0.27 sce00500 淀粉和蔗糖代谢 4/15 2.79×10−2 0.35 说明:匹配情况为匹配化合物个数/通路化合物总数 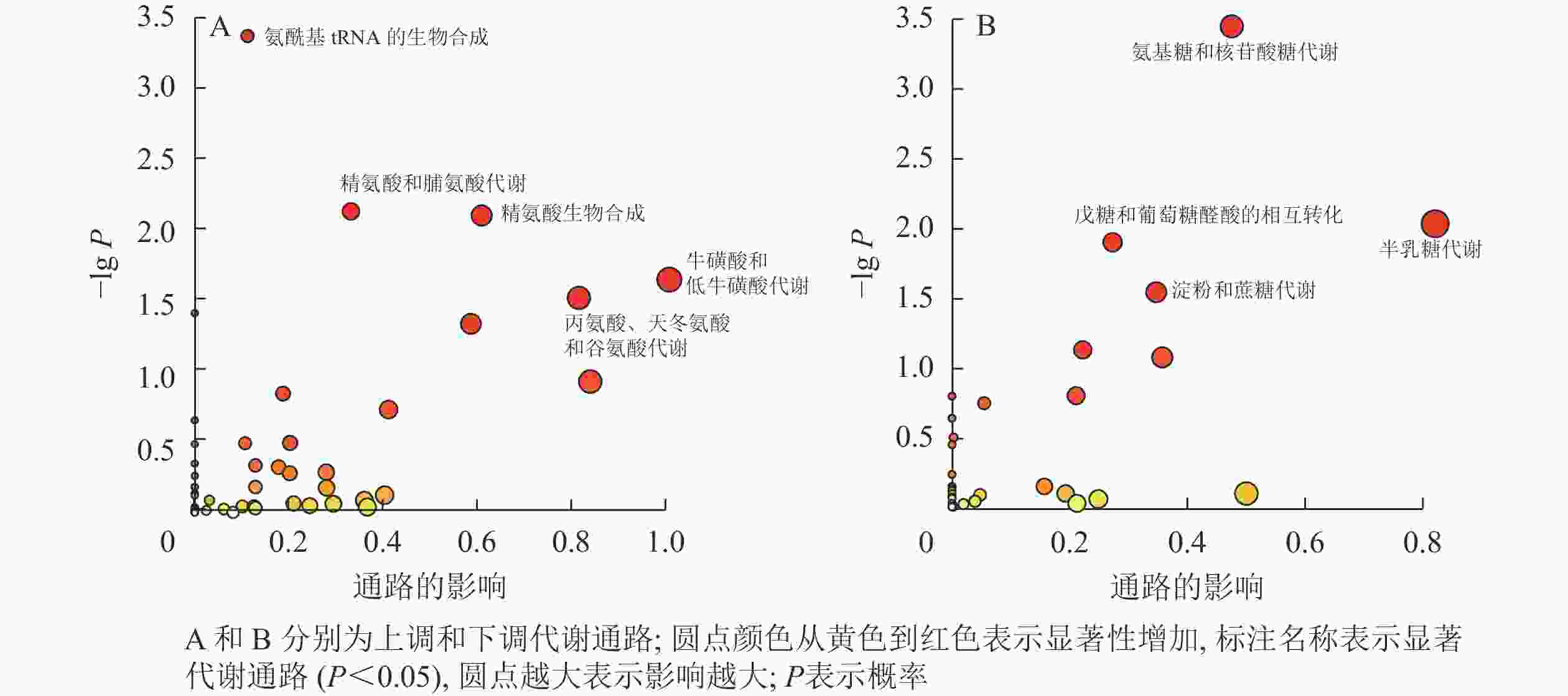
图 3 差异代谢产物富集到的显著代谢通路
Figure 3. Significant metabolic pathways enriched by differential metabolites
-
本研究在代谢组水平探究含内共生细菌的生防真菌EV菌对不同碳氮培养基的化学响应,鉴定到一些差异显著的化合物,尤其是在氮源培养基中特有的代谢产物,并定位到重要代谢途径上。磷酸胍基乙酸酯和对甲酚硫酸盐是在氮培养下EV菌大量产生的特有代谢物,尿囊素、光色素、吲哚和海藻糖等是氮培养条件下显著上调的代谢物。对甲酚硫酸盐是细菌代谢酪氨酸的产物[13]。已有研究证实:拟杆菌科Bacteroidaceae、双歧杆菌科Bifidobacteriaceae、梭菌科Clostridiaceae、肠杆菌科Enterobacteriaceae、肠球菌科Enterococcaceae、优杆菌科Eubacteriaceae、梭杆菌科Fusobacteriaceae、毛螺菌科Lachnospiraceae、乳杆菌科Lactobacillaceae、紫单胞菌科Porphyromonadaceae、葡萄球菌科Staphylococcaceae、疣微菌科Ruminococcaceae和韦荣氏菌科Veillonellaceae等细菌是对甲酚硫酸盐的产生菌[14]。已有研究表明:细菌也可以降解对甲酚[15],真菌Trichosporon cutaneum和Aspergillus fumigatus也可以利用对甲酚作为碳源[16-17]。因此推测:对甲酚硫酸盐可能是在高氮源培养下,由于碳源严重缺乏,EV菌胞内共生细菌代谢酪氨酸产生对甲酚或对甲酚硫酸盐,从而将其做为碳源进一步利用。
多种革兰氏阳性细菌和革兰氏阴性细菌(迄今为止共有85种)都会产生大量的吲哚。作为细胞间信号分子,吲哚控制细菌生理的多个方面,例如孢子形成、质粒稳定性、耐药性、生物膜形成和吲哚产生细菌的毒力[18]。在细菌中,吲哚由氨基酸色氨酸的降解产物产生。吲哚是细菌Ⅲ型分泌系统表达的信号分子[19]。EV真菌细菌共生体可产生吲哚,根据上述目前的研究只在细菌发现吲哚的产生,因此我们推测胞内细菌可能是吲哚的产生者,细菌可以利用吲哚这一信号分子协调两者行为,以在真菌胞内生存。
前期研究表明:EV菌基因组中含有比其他杀线虫真菌,如Arthrobotrys oligospora、Dactylellina haptotyla、Drechmeria coniospora更多的尿囊素转运蛋白[20]。此外,EV菌胞内的共生细菌属于假单孢属Pseudomonas[6]。尿囊素是腺嘌呤和鸟嘌呤代谢的中间产物,尿囊素可以被某些真菌、细菌和植物用作碳和氮的来源,尿囊素或尿囊素通路的衍生物可能有助于提高真菌向宿主植物提供氮的能力[21]。如在一些外生菌根真菌中,已鉴定出尿囊素/尿囊酸转运蛋白[21-22]。研究发现:假单孢属的细菌和酵母Saccharomyces cerevisiae也可以降解尿囊素[23-24]。在氮源充足的氮培养条件下,尿囊素代谢产物约是碳培养条件下的32倍,因此,尿囊素很可能是EV菌供给胞内共生细菌氮源的一种形式。
海藻糖在自然界中分布广泛,包括细菌、真菌、植物、无脊椎动物和哺乳动物。由于其特殊的物理特性,海藻糖能够保护细胞的完整性免受各种环境损害和营养限制。细菌可以使用海藻糖作为碳和能量的唯一来源,一些分枝杆菌中海藻糖可作为细胞壁的结构成分,而酵母细胞在很大程度上不能以海藻糖为碳源生长[25]。在根瘤菌-豆科Leguminosae植物共生期间,海藻糖在根瘤发育过程中被储存在根瘤中,并成为细菌的主要碳水化合物,而与所供应的碳源和氮源类型无关[26]。内源性海藻糖在碳缺乏或在给定培养基中碳源耗尽后被动员。海藻糖很可能是EV共生细菌的一种碳源,在碳源缺乏时作为储备能源(氮培养下产生的海藻糖是碳培养下的2.5倍)。
光色素是核黄素的光敏分解产物。关于光色素的酶促转化路径已在假单胞菌等细菌中进行研究[27]。已有研究认为:光色素是细菌群感效应的信号分子[28],是能影响植物生长的细菌信号分子[29],同时也是共生的信号分子[30-31]。不同的营养条件(如氮和磷)可改变细菌产生光色素的浓度[30]。本研究使用的氮源是有机氮源。在该氮培养下产生的光色素是碳培养下的8倍,氮培养条件下光色素的产量远高于碳培养条件,不同于前人报道的高浓度硝酸盐降低光色素的产量[30]。
差异最显著的代谢物多数未能富集到显著的代谢通路,原因在于这些化合物的代谢通路尚未研究透彻,或者它们是EV菌的特异化合物,尚未包括在参照数据库中。无论以真菌还是细菌数据库为参照,富集到的显著代谢通路基本一致,均与氨基酸和糖类代谢相关,而这些通路是真菌和细菌共有的,无法区分内共生细菌对其真菌宿主代谢的影响。笔者曾试图利用多种抗生素去除内共生细菌以解决该问题,并未获得成功,需要更深入的研究。
Metabolome of nematicidal fungus Esteya vermicola in carbon and nitrogen culture
-
摘要:
目的 比较伊氏杀线虫真菌Esteya vermicola (EV菌)在碳、氮营养源培养下的代谢差异,并找到重要代谢物或信号分子。 方法 选取培养真菌的碳培养基[主要为马铃薯葡萄糖肉汤培养基(PDB)]和培养细菌的氮培养基(主要为酵母粉),将EV菌在2种培养基上25 ℃条件下培养7 d,收获菌丝体并提取代谢产物。采用非靶标的高效液相色谱-质谱联用技术(HPLC-MS),在阴、阳离子模式下对代谢物组分进行分析和鉴定,并分析差异显著代谢物的代谢通路。 结果 共得到498种代谢物,阴、阳离子模式分别有176和362种,其中2种模式共同含有40种。差异显著的代谢物共有444种,占总数的89.2%,其中阴、阳离子模式分别有162和310种,有28种为2种模式共有。主成分和偏最小二乘判别分析均可使碳、氮培养条件下的代谢物聚为不同的簇并显著分离。氮培养条件下,磷酸胍基乙酸酯和对甲酚硫酸盐是大量产生且特有的代谢物;重要代谢物尿囊素、光色素、吲哚和海藻糖产量显著上调。通路分析将显著上调和下调的代谢物分别富集到氨基酸和糖类代谢相关的代谢通路。 结论 EV菌在碳培养和氮培养条件下呈现明显的代谢差异,代谢通路主要涉及糖类和氨基酸代谢。重要代谢物可为EV菌的高效培养和应用提供基础。图3表2参31 Abstract:Objective The objective is to compare the metabolic differences of the nematode-killing fungus Esteya vermicola (EV) cultured in carbon and nitrogen nutrient sources and to identify key metabolites or signal molecules. Method The carbon medium (mainly composed of PDB) for culturing fungi and the nitrogen medium (mainly composed of yeast powder) for culturing bacteria were selected. EV bacteria were cultured on two kinds of culture media at 25 ℃ for 7 days. The mycelia were harvested and the metabolites were extracted. Non-target high performance liquid chromatography-mass spectrometry (HPLC-MS) was used to analyze and identify metabolite components in both positive and negative ion modes. The metabolic pathways of metabolites with significant differences were analyzed. Result A total of 498 metabolites were identified, including 176 negative and 362 positive ion modes and 40 metabolites in both modes. There were 444 metabolites with significant differences, accounting for 89.2% of the total, among which 162 were negative and 310 were positive, and 28 were common to the two modes. Both principal component analysis and partial least square discriminant analysis could cluster the metabolites into different clusters and separate them significantly in carbon and nitrogen culture. In nitrogen culture, guanidine phosphate acetate and p-cresol sulfate were abundant and unique metabolites, and the yield of allantoin, photopigment, indole, and trehalose were significantly up-regulated. Pathway analysis enriched the significantly up-regulated and down-regulated metabolites into the metabolic pathways related to amino acid and carbohydrate metabolism, respectively. Conclusion EV bacteria showed significant metabolic differences in carbon and nitrogen culture. The metabolic pathway mainly involves carbohydrate and amino acid metabolism. The important metabolites will provide a theoretical basis for efficient culture and application of EV. [Ch, 3 fig. 2 tab. 31 ref.] -
Key words:
- biocontrol fungi /
- Esteya vermicola /
- metabonomics /
- LC-MS
-
图 1 碳、氮培养条件下EV菌代谢物的PCA和PLS-DA分析
Figure 1 PCA and PLS-DA of EV metabolites under carbon and nitrogen culture conditions
图 2 碳、氮培养条件下EV菌的差异代谢物
Figure 2 Differential metabolites of EV bacteria under carbon and nitrogen culture conditions
图 3 差异代谢产物富集到的显著代谢通路
Figure 3 Significant metabolic pathways enriched by differential metabolites
表 1 部分差异化合物
Table 1. Part of significantly different metabolites
化合物名称 质荷比 保留时间/min 二级数据库得分 变化倍数 P KEGG编号 磷酸胍基乙酸酯 198.03 9.86 62 12.46 1.13×10−13 C03166 对甲酚硫酸盐 187.01 6.17 53 12.41 8.88×10−15 邻氨基苯甲酸酯 136.04 2.00 50 9.93 1.67×10−8 C00108 尿酸 190.05 1.29 40 8.26 6.26×10−7 C01717 尼古丁 163.12 4.68 44 5.51 1.14×10−6 C16150 4-羟基-2-喹啉羧酸 188.03 5.76 30 5.48 9.62×10−8 C01717 烟酸 124.04 3.50 41 5.17 1.71×10−7 C00253 尿囊素 159.05 5.46 50 5.09 2.53×10−4 C01551 吲哚丙烯酸 188.07 7.98 98 4.20 4.68×10−8 2-吡咯烷酮 86.06 3.79 41 4.10 2.57×10−5 C11118 咪唑乙酸 127.05 9.62 69 3.54 1.56×10−7 C02835 组胺 112.09 5.74 45 3.34 2.79×10−8 C00388 谷胱甘肽 306.08 1.02 58 3.26 5.97×10−4 C00051 光色素 243.09 1.58 70 3.00 2.79×10−4 C01727 肌肽 227.11 12.66 55 2.40 3.30×10−5 C00386 甜菜碱醛 102.09 6.73 68 2.25 9.68×10−3 C00576 吲哚 118.07 8.32 57 2.12 4.40×10−3 C00463 苹果酸 133.01 0.90 43 1.90 2.86×10−5 C00711 甜菜碱 118.09 8.30 51 1.88 2.79×10−8 C00719 胍基乙酸d 118.06 8.26 38 1.87 1.19×10−3 C00581 葫芦巴碱 138.06 9.05 38 1.59 1.71×10−7 C01004 海藻糖 341.11 1.04 78 1.34 6.16×10−5 C01083 左旋肉碱 162.11 9.31 72 1.34 2.97×10−6 C00318 邻乙酰左旋肉碱 204.12 8.54 73 1.32 1.69×10−6 C02571 茶碱 181.07 2.24 55 −1.64 6.07×10−7 C07130 二乙醇胺 106.09 8.16 45 −1.73 1.42×10−4 C06772 去甲肾上腺素 170.08 5.83 57 −2.40 1.52×10−5 C00547 阿魏酸盐 193.05 5.36 32 −3.58 2.87×10−5 C01494 核糖醇 151.06 1.01 46 −4.08 3.13×10−8 C00474 甲基咪唑乙酸 141.07 9.52 41 −4.51 2.90×10−8 C05828 赤藓糖醇 121.05 1.04 75 −5.20 2.80×10−5 C00503  下载: 导出CSV
下载: 导出CSV
表 2 碳、氮培养条件下EV菌差异代谢物富集到的KEGG通路
Table 2. Enriched KEGG pathways by differential metabolites of EV under carbon and nitrogen culture conditions
代谢通路编号 代谢通路名称 匹配情况 P 影响大小 上调 sce00970 氨酰基tRNA的生物合成 18/46 4.38×10−4 0.11 sce00330 精氨酸和脯氨酸代谢 10/25 7.53×10−3 0.33 sce00220 精氨酸生物合成 8/18 8.05×10−3 0.60 sce00430 牛磺酸和低牛磺酸代谢 4/7 2.28×10−2 1.00 sce00250 丙氨酸、天冬氨酸和谷氨酸代谢 8/22 3.06×10−2 0.81 下调 sce00520 氨基糖和核苷酸糖代谢 8/24 3.41×10−4 0.48 sce00052 半乳糖代谢 5/17 8.98×10−3 0.82 sce00040 戊糖和葡萄糖醛酸的相互转化 4/12 1.22×10−2 0.27 sce00500 淀粉和蔗糖代谢 4/15 2.79×10−2 0.35 说明:匹配情况为匹配化合物个数/通路化合物总数
下载: 导出CSV
-
[1] FUTAI K. Pine wood nematode, Bursaphelenchus xylophilus [J]. Ann Rev Phytopathol, 2013, 51(1): 61 − 83. [2] VICENTE C, ESPADA M, VIEIRA P, et al. Pine wilt disease: a threat to european forestry [J]. Eur J Plant Pathol, 2012, 133(1): 89 − 99. [3] LIOU J Y, SHIH J Y, TZEAN S S. Esteya, a new nematophagous genus from Taiwan, attacking the pinewood nematode (Bursaphelenchus xylophilus) [J]. Mycol Res, 1999, 103(2): 242 − 248. [4] WANG Ruizhen, DONG Leiming, CHEN Yuequ, et al. Esteya Vermicola, a nematophagous fungus attacking the pine wood nematode, harbors a bacterial endosymbiont affiliated with Gammaproteobacteria [J]. Microbes Environ, 2017, 32(3): 201 − 209. [5] SCHERLACH K, BUSCH B, LACKNER G, et al. Symbiotic cooperation in the biosynthesis of a phytotoxin [J]. Angewandte Chemie, 2012, 51(38): 9615 − 9618. [6] HOFFMAN M T, GUNATILAKA M K, WIJERATNE K, et al. Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte[J/OL]. PLoS One, 2013, 8(9): e73132[2021-11-23]. doi: 10.1371/journal.pone.0073132. [7] ROOSSINCK M J. Metagenomics of plant and fungal viruses reveals an abundance of persistent lifestyles[J/OL]. Front Microbiol, 2015, 5[2021-11-23]. doi: 10.3389/fmicb.2014.00767. [8] SALVIOLI A, GHIGNONE S, NOVERO M, et al. Symbiosis with an endobacterium increases the fitness of a mycorrhizal fungus, raising its bioenergetic potential [J]. ISME J, 2016, 10(1): 130 − 144. [9] LUMINI E, BIANCIOTTO V, JARGEAT P, et al. Presymbiotic growth and sporal morphology are affected in the arbuscular mycorrhizal fungus Gigaspora margarita cured of its endobacteria [J]. Cell Microbiol, 2007, 9(7): 1716 − 1729. [10] SALVIOLI A, CHIAPELLO M, FONTAINE J, et al. Endobacteria affect the metabolic profile of their host Gigaspora margarita, an arbuscular mycorrhizal fungus [J]. Environ Microbiol, 2010, 12(8): 2083 − 2095. [11] FIEHN O, KOPKA J, DÖRMANN P, et al. Metabolite profiling for plant functional genomics [J]. Nat Biotechnol, 2000, 18(11): 1157 − 1161. [12] NICHOLSON J K, LINDON J C, HOLMES E. ‘Metabonomics’: understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data [J]. Xenobiotica, 1999, 29(11): 1181 − 1189. [13] D’ARI L, BARKER H A. p-Cresol formation by cell-free extracts of Clostridium difficile [J]. Arch Microbiol, 1985, 143(3): 311 − 312. [14] GRYP T, VANHOLDER R, VANEECHOUTTE M, et al. p-Cresyl sulfate [J/OL]. Toxins, 2017, 9(2): 52[2021-12-01]. doi: 10.3390/toxins9020052. [15] BOSSERT I D, YOUNG L Y. Anaerobic oxidation of p-cresol by a denitrifying bacterium [J]. Appl Environ Microbiol, 1986, 52(5): 1117 − 1122. [16] JONES K H, TRUDGILL P W, HOPPER D J. Metabolism of p-cresol by the fungus Aspergillus fumigatus [J]. Appl Environ Microbiol, 1993, 59(4): 1125 − 1130. [17] POWLOWSKI J B, DAGLEY S. β-Ketoadipate pathway in Trichosporon cutaneum modified for methyl-substituted metabolites [J]. J Bacteriol, 1985, 163(3): 1126 − 1135. [18] LEE J H, LEE J. Indole as an intercellular signal in microbial communities [J]. FEMS Microbiol Rev, 2010, 34(4): 426 − 444. [19] HIRAKAWA H, KODAMA T, TAKUMI-KOBAYASHI A, et al. Secreted indole serves as a signal for expression of type III secretion system translocators in enterohaemorrhagic Escherichia coli O157: H7 [J]. Microbiology, 2009, 155(2): 541 − 550. [20] WANG Ruizhen, DONG Leiming, RAN Qinghua, et al. Comparative genomic analyses reveal the features for adaptation to nematodes in fungi [J]. DNA Res, 2018, 25(3): 245 − 256. [21] LARSEN P E, SREEDASYAM A, TRIVEDI G, et al. Using next generation transcriptome sequencing to predict an ectomycorrhizal metabolome[J/OL]. BMC Syst Biol, 2011, 5(1): 70[2021-11-23]. doi: 10.1186/1752-0509-5-70. [22] RINEAU F, SHAH F, SMITS M M, et al. Carbon availability triggers the decomposition of plant litter and assimilation of nitrogen by an ectomycorrhizal fungus [J]. ISME J, 2013, 7(10): 2010 − 2022. [23] CAMPBELL L L. The mechanism of allantoin degradation by a Pseudomonas [J]. J Bacteriol, 1954, 68(5): 598 − 603. [24] TRIJBELS F, VOGELS G D. Degradation of allantoin by Pseudomonas acidovorans [J]. Biochim Biophys Acta, 1966, 113(2): 292 − 301. [25] ARGÜELLES J C. Physiological roles of trehalose in bacteria and yeasts: a comparative analysis [J]. Arch Microbiol, 2000, 174(4): 217 − 224. [26] SALMINEN S O, STREETER J G. Enzymes of \begin{document}$ \alpha $\end{document} \begin{document}$ \alpha $\end{document} [27] FOSTER J W, YANAGITA T. A bacterial riboflavin hydrolase [J]. J Biol Chem, 1956, 221(2): 593 − 607. [28] RAJAMANI S, BAUER W D, ROBINSON J B, et al. The vitamin riboflavin and its derivative lumichrome activate the LasR bacterial quorum-sensing receptor [J]. Mol Plant-Microb Interactions, 2008, 21(9): 1184 − 1192. [29] DAKORA F D. Lumichrome: A Bacterial Signal Molecule Influencing Plant Growth, in: Biological Nitrogen Fixation[M]. Hoboken: John Wiley & Sons, Inc, 2015: 389 − 396. [30] KANU S A, DAKORA F D. Effect of N and P nutrition on extracellular secretion of lumichrome, riboflavin and indole acetic acid by N2-fixing bacteria and endophytes isolated from Psoralea nodules [J]. Symbiosis, 2012, 57(1): 15 − 22. [31] PHILLIPS D A, JOSEPH C M, YANG G P, et al. Identification of lumichrome as a Sinorhizobium enhancer of alfalfa root respiration and shoot growth [J]. Proc Natl Acad Sci U S A, 1999, 96(22): 12275 − 12280. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210828
 点击查看大图
点击查看大图
计量
- 文章访问数: 1721
- HTML全文浏览量: 418
- PDF下载量: 180
- 被引次数: 0
