




-
随着全球碳排放的增长,森林火灾频发,森林资源可持续发展受到严重威胁,自然生态平衡遭到破坏,防御和控制森林火灾也成为国际社会关注的焦点[1]。火是陆地生态系统演化的重要驱动因素,在生态系统的进化与发展中十分重要[2]。过度禁火或频繁的林火干扰都不利于森林生态系统修复与更新,在一定程度上影响生态系统的物质循环和能量流动,导致生态系统进化环节缺失[3]。森林长期禁火导致林下可燃物不断蓄积,林分质量变差,还增加林地内火灾风险。与森林火灾不同,计划烧除是针对适宜林种,在规定范围用低强度地表火烧除林区内可燃物,达到降低火险、提高森林防火能力的技术性措施[4]。周期性计划烧除能清除林内杂物,减少病虫害,增强土壤碳稳定性,促进森林的生长与更新,对森林群落演替和保护生态环境具有重要作用[5−6]。云南松Pinus yunnanensis作为西南地区的特有物种,在西南地区分布广泛,生态适应幅度很广,是区域内荒山造林的先锋树种。云南松富含油脂,林冠下地表凋落物极易燃烧,大部分的云南松生活在森林火灾高发区[7]。从20世纪90年代起,云南省多个地区已经大面积开展计划烧除工作,森林防火效果较好[8]。尽管计划烧除的强度较低,但是低强度地表火会使林地内凋落物蓄积量减少,植被根系烧毁,降雨直接冲刷裸露地表,不利于土壤团聚体的形成,势必会影响土壤抗侵蚀和抗冲刷能力。有研究发现:周期性计划烧除还会使土壤表层团粒结构改变,可蚀性增强[9],促进坡面径流泥沙产生[10],甚至影响土壤养分稳定性,对土壤养分环境产生长期影响[11]。
计划烧除作为生态系统管理的有效手段,最早在加拿大、澳大利亚等林业发达国家实施运用,技术和体系较为成熟。国外学者在探究适宜的计划烧除制度方面进行了研究,同时持续关注计划烧除对土壤养分的影响,监测空气质量、森林生态环境等[12−14]。国内计划烧除工作起步较晚,研究内容集中在计划烧除对森林防火的积极作用以及计划烧除对森林生态系统、群落结构的影响等[15−16]。国内外关于火烧对土壤理化性质[17]、土壤微生物[18]的影响已有报道,但集中在林火发生的频次和强度不固定的火灾区,在指定树种、周期性计划烧除对土壤侵蚀方面的影响报道较少。探究计划烧除对林内土壤抗蚀和抗冲性能的影响,对明晰低强度火干扰下土壤侵蚀的内在机制显得尤为重要。本研究以期揭示计划烧除对土壤侵蚀、森林生态环境的影响,明确计划烧除在林地管理中的作用,并为森林的可持续经营以及优化林区计划烧除制度提供可靠的科学依据。
-
研究区位于滇中高原玉溪市新平县南部的照壁山,海拔为1 990~2 050 m,是典型的高原山地丘陵地貌。代表性土壤为红壤,是云南省亚热带北部与亚热带南部的过渡地带。多年平均降水量为1 050 mm,最高气温为32.0 ℃,最低气温为1.0 ℃,年平均气温为15.0 ℃。
-
在照壁山上对该区域内林分类型、地质地貌及自然条件进行系统调查,选择立地条件相似,平均树龄为30 a且长势较好的云南松林地为研究对象。在防火通道2侧对称布设计划烧除与未烧除样地。林区内灌木主要有木荷Schima superba、杜鹃Rhododendron simsii等,草本主要有紫茎泽兰Ageratina adenophora、贯众Cyrtomium fortunei、毛茛Ranunculus japonicus、白茅Imperata cylindrica等。从20世纪90年代末期起,该区域制订了相对完善的计划烧除制度,并于每年1—2月开展计划烧除工作,至今20余年从未间断。2021年1月实施了该年度森林防火计划烧除工作,计划烧除火焰高度1.5 m,属低强度火烧。在计划烧除和未烧除2块大样地内分别设置20 m×20 m标准样地3块,于2021年7月1—4日采集土壤样品。
-
按0~10、10~20、20~40和40~60 cm土层分层采样,测定土壤团聚体相关指标,每个样品设置3个重复且均大于500 g。土壤团聚体采用干筛和湿筛法分级,按照粒径>5.00、5.00~2.00、2.00~1.00、1.00~0.50、0.50~0.25和<0.25 mm的分级标准分别称量,并计算各粒级质量百分比,土壤抗蚀指数采用静水崩解法测定。
平均质量直径(DMW)、平均几何直径(DMG)[19]计算公式如下:
$$ \begin{gathered} {{D_{\rm{MW}}}} = \sum\limits_{i = 1}^n {{w_i}} {x_i};\; {D_{\rm{MG}}}=\text{exp}\left(\frac{\displaystyle \sum \limits _{i=1}^{n}{w}_{i} \mathrm{ln}{x}_{i}} {{\displaystyle \sum \limits _{i=1}^{n}{w}_{i}}}\right)。 \end{gathered} $$ 其中:wi为第i粒径团聚体组分的质量占土壤总质量的质量百分比(%);xi为第i粒径团聚体组分的平均直径(mm)。
-
在计划烧除和未烧除2块大样地内设置的3块标准样地中,分别设置3个50 cm×50 cm的样方,收集样方中地表凋落物分解层和半分解层,带回实验室即刻称量。采用室内浸泡法,测定凋落物自然含水率、蓄积量以及持水性能[20]等指标。
-
土壤抗冲性采用原状土冲刷水槽法测定[21]。在收集完凋落物的样方中,用自制20 cm×10 cm×5 cm取样器取原状土样,取完后用保鲜膜和泡沫袋将土样包裹,避免运输过程中对原状土样的破坏。实验前将土样浸水24 h至土壤样品饱和,沥去多余重力水后置于1.2 m×0.1 m冲刷槽内。设计冲刷坡度为20°,冲刷流量为2 L·min−1,冲刷时间固定为10 min,隔1 min取1次浑水样,共取10次。试验结束后,将浑水样静置过滤,并烘干称泥沙质量。土壤抗冲系数计算公式如下:CAS=(Qt)/M。其中:CAS为土壤抗冲系数(L·g−1);M为冲走的土样质量(g);t为冲刷时间(min);Q为冲刷流量(L·min−1)。
-
将冲刷试验结束后取样器内剩余土样置于0.25 mm土筛中反复冲洗,洗出根系后于65 ℃条件下烘干,称量,用于计算根长密度[DR=N/(LWH)]和根系生物量[BR=Mr/(LWH)][22]。其中:N为取样器中根系总长度(cm),L、W、H分别为取样器的长、宽、高(cm),Mr为根系干质量(g)。
-
采用Excel和SPSS 21.0 进行数据统计分析(试验数据均采用上限排除法进行统计),并用 Origin 9.0 绘制图形。
-
土壤团聚体质量百分比差异直接影响土壤孔隙度的大小,导致土壤结构和功能发生变化。计划烧除后地表凋落物和土壤环境受到影响,团聚体组分特征发生变化。如图1所示:干筛后各土层土壤团聚体均以粒径>5.00 mm团聚体占比最大,粒径0.25~0.50 mm团聚体占比最小;随土层深度增加,粒径>5.00 mm团聚体所占比例减小,粒径<0.25 mm团聚体占比增大。计划烧除后,0~40 cm土层粒径>5.00、2.00~5.00 mm团聚体比例增加,粒径<0.25 mm团聚体比例减小;其中0~10 cm土层变化较明显。未烧除样地0~10 cm土层土壤粒径>5.00、2.00~5.00和<0.25 mm团聚体含量分别为24.73%、14.70%和18.13%,计划烧除样地为28.62%、15.95%和13.58%。
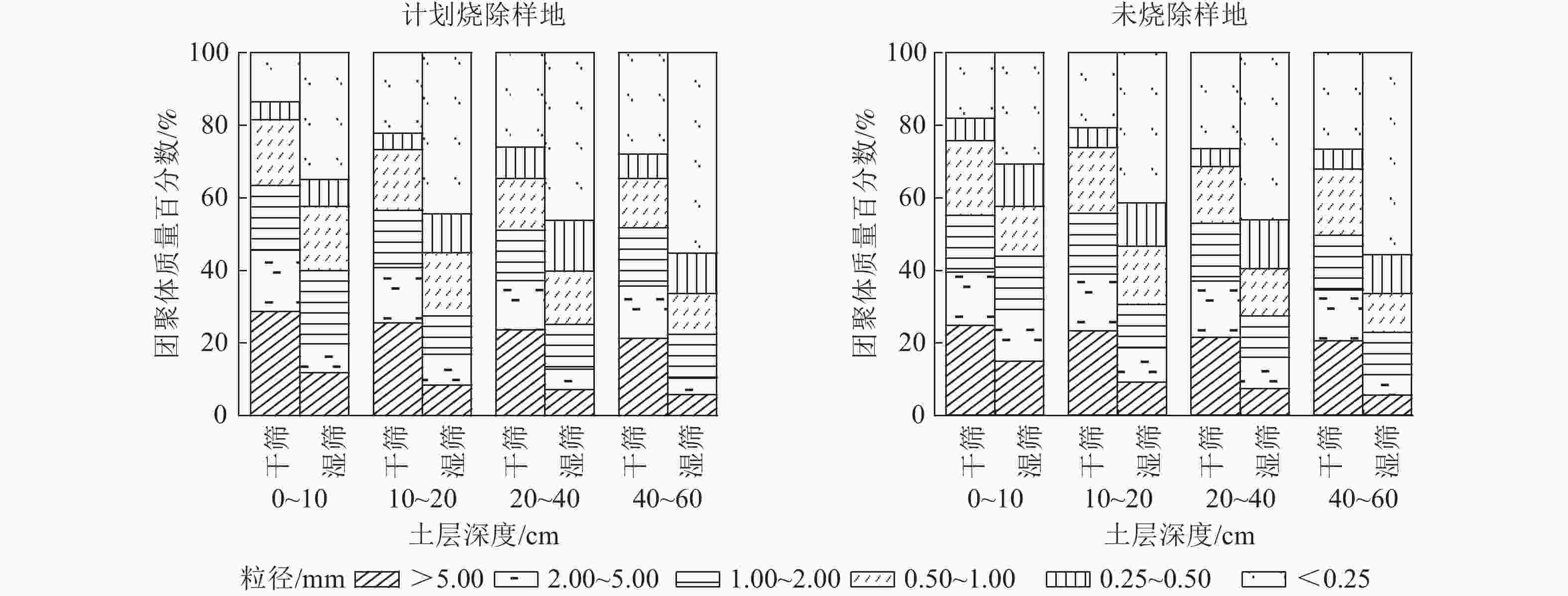
图 1 土壤团聚体分布特征
Figure 1. Distribution characteristics of soil aggregates
图1显示:湿筛后各土层土壤团聚体均以粒径<0.25 mm占比最大。随着土层深度增加,粒径>5.00和2.00~5.00 mm团聚体所占比例减小,粒径0.25~0.50和<0.25 mm团聚体占比增大。未烧除样地0~10、10~20和20~40 cm土层土壤粒径>2.00 mm土壤团聚体含量分别为29.17%、18.57%和16.01%,计划烧除样地为19.79%、16.80%和12.82%。表明计划烧除后林地内土壤0~40 cm土层粒径>2.00 mm土壤机械稳定性团聚体含量增加,水稳性团聚体含量减少,40~60 cm土层各粒级团聚体含量无明显变化。
-
在团聚体组分特征的基础上,平均质量直径和平均几何直径也是评价土壤团聚体大小和结构稳定性的重要指标。一般来说平均质量直径和平均几何直径越大,土体结构稳定性越好[23]。土壤团聚体稳定性特征见表1。
表 1 土壤团聚体稳定性特征
Table 1. Soil gathering body stability characteristics
土层深度/cm 计划烧除样地 未烧除样地 平均质量直径/mm 平均几何直径/mm 平均质量直径/mm 平均几何直径/mm 干筛 湿筛 干筛 湿筛 干筛 湿筛 干筛 湿筛 0~10 3.18±0.09 A 1.67±0.11 B 1.61±0.05 A 0.65±0.02 B 2.80±0.15 B 2.02±0.10 A 1.28±0.07 B 0.77±0.07 A 10~20 2.86±0.08 B 1.30±0.11 C 1.24±0.07 B 0.47±0.03 C 2.73±0.04 C 1.41±0.00 B 1.22±0.08 B 0.51±0.01 C 20~40 2.63±0.03 C 1.14±0.03 D 1.04±0.02 C 0.42±0.01 D 2.56±0.13 D 1.23±0.05 C 1.05±0.05 C 0.44±0.01 D 40~60 2.50±0.07 E 0.97±0.01 E 0.99±0.04 C 0.34±0.01 E 2.45±0.06 E 0.99±0.01 E 0.97±0.06 C 0.34±0.01 E 说明:数值为平均值±标准差。同行数值后不同大写字母表示不同处理间差异显著(P<0.05) 表1表明:随着土层深度增加,平均质量直径和平均几何直径逐渐减小。干筛处理下,计划烧除使0~40 cm土层平均质量直径和0~10 cm土层平均几何直径显著增大(P<0.05),40~60 cm土层平均质量直径和平均几何直径无显著变化。而湿筛处理下,计划烧除使0~40 cm土层平均质量直径和0~10 cm土层平均几何直径显著减小(P<0.05),40~60 cm土层平均质量直径和平均几何直径无显著差异,说明计划烧除使土壤团聚度降低,土体结构遇水稳定性减弱。
-
团聚体结构破坏率是表征土壤团聚体稳定性的重要指标。团聚体结构破坏率越小,土壤结构越稳定,抗侵蚀能力越好。土壤团聚体结构破坏率及抗蚀指数见表2。
表 2 土壤团聚体结构破坏率及抗蚀指数
Table 2. Destruction rate and corrosion index of soil cluster structure
土层深
度/cm计划烧除样地 未烧除样地 粒径>0.25 mm土壤团聚体比例/% 团聚体结构
破坏率/%抗蚀指
数/%粒径>0.25 mm土壤团聚体比例/% 团聚体结构
破坏率/%抗蚀指
数/%干筛 湿筛 干筛 湿筛 0~10 86.42±0.97 A 65.07±0.98 B 21.35±1.84 A 64.44±2.95 B 81.87±2.20 B 69.24±2.82 A 12.62±2.83 B 73.78±5.69 A 10~20 77.82±3.28 C 55.61±1.54 D 22.21±2.46 A 52.89±4.91 C 79.26±3.28 C 58.52±1.62 C 20.74±2.35 A 58.67±6.53 B 20~40 73.90±0.25 D 53.80±0.64 D 20.10±0.77 A 37.38±8.02 D 73.62±2.14 D 53.90±0.69 D 19.73±1.45 A 38.67±8.22 D 40~60 73.04±0.68 D 43.75±1.04 E 29.29±0.73 C 27.56±5.48 E 73.36±1.78 D 44.14±0.61 E 29.22±1.17 C 28.00±5.66 E 说明:数值为平均值±标准差。同行数值后不同大写字母表示不同处理间差异显著(P<0.05) 表2表明:计划烧除后0~10 cm土层团聚体结构破坏率显著增大(P<0.05),40~60 cm土层团聚体结构破坏率无显著差异。采用静水崩解法,能较为直观地分析土壤抗水蚀的能力大小。火烧后,0~20 cm土层抗蚀指数显著减小(P<0.05),随土层深度的增加,土粒抵御静水崩解的能力有减弱的趋势,但在40~60 cm土层基本无影响。火烧后林内植被覆盖度降低,使土壤性质和功能发生变化,土壤团聚体遇水更容易崩散,表层团聚体结构破坏率显著增大。可见计划烧除使土壤表层抗蚀性减小,随着土层深度的增加这种影响逐渐变小。
-
林下凋落物的覆盖能很大程度地减缓降雨对土壤的冲刷。计划烧除后林下草本植物以及地表凋落物全部化为灰烬,灌木生长受到影响,地表裸露面积增加。本研究对多年连续计划烧除后地表凋落物蓄积量及持水性能进行测定,结果见表3。
表 3 凋落物蓄积量及持水性能
Table 3. Destiny storage volume and water holding performance
样地类型 凋落物层 凋落物蓄积量/
(t·hm−2)最大持水量/
(t·hm−2)最大持水率/% 最大拦蓄量/
(t·hm−2)最大拦蓄率/% 有效拦蓄率/% 计划烧除样地 未分解层 2.95±0.18 c 4.80±0.64 c 163.34±24.23 d 3.71±0.64 c 126.27±23.48 d 89.19±22.81 d 半分解层 0.87±0.05 d 2.55±0.11 d 294.26±19.16 b 2.00±0.11 d 230.49±17.70 b 166.72±16.88 b 合计 3.82±0.23 7.35±0.75 193.13±22.79 5.71±0.75 149.98±21.97 106.83±21.25 未烧除样地 未分解层 5.91±0.16 a 13.90±0.20 a 235.27±7.89 c 11.19±0.35 a 189.38±10.05 c 143.50±12.21 c 半分解层 3.43±0.21 b 11.67±0.43 b 340.48±8.04 a 9.77±0.35 b 285.12±9.07 a 229.76±12.96 a 合计 9.35±0.20 26.55±1.30 283.93±9.21 21.93±1.24 234.52±9.71 185.12±10.23 说明:数值为平均值±标准差。同列数值后不同小写字母表示2种样地类型凋落物蓄积量存在显著差异(P<0.05) 由表3可以看出:经多年连续计划烧除后,林内地表蓄积量仅为3.82 t·hm−2,未进行计划烧除的对照样地地表蓄积量为9.35 t·hm−2,是计划烧除样地的2.45倍。凋落物组分变化导致凋落物层持水性能也随之改变,未烧除样地最大持水率、最大拦蓄率、有效拦蓄率均高于计划烧除样地,分别是计划烧除样地的1.47、1.90和2.15倍;分层来看,2种样地类型半分解层蓄积量均低于未分解层,但半分解层最大持水率、最大拦蓄率、有效拦蓄率均高于未分解层。表明计划烧除后凋落物蓄积量显著减少,持水能力下降。
-
在土壤表层,植物根系分布较广,主要来源于灌木和草本植物。计划烧除后土壤根长密度及生物量见表4。表4表明:在表层土壤根系分布中,计划烧除和未烧除样地<0.50 mm径级的根生物量及根长密度均大于>2.00、0.50~2.00 mm径级,均以<0.50 mm径级根系为主。计划烧除后林地<0.50 mm根系占总生物量百分比显著降低(P<0.05)。可见低强度地表火对细根影响较大,一些较粗根系能抵御低强度火烧或在火烧后逐渐恢复生长。
表 4 根长密度及生物量
Table 4. Root long density and biomass
根系径级/mm 计划烧除样地 未烧除样地 根长密度/
(×10−3cm·cm−3)根系生物量/
(×10−3g·cm−3)占根总生物量百分比/% 根长密度/
(×10−3cm·cm−3)根系生物量/
(×10−3g·cm−3)占根总生物量百分比/% >2.00 9.70±1.10 e 0.28±0.02 e 35.20±4.14 b 29.17±1.86 d 0.76±0.08 b 24.64±2.51 c 0.50~2.00 24.73±2.50 d 0.20±0.03 e 24.82±3.32 c 46.10±4.17 c 0.46±0.04 c 14.77±1.38 d <0.50 59.33±7.41 b 0.33±0.03 d 39.99±1.37 b 162.80±14.62 a 1.89±0.07 a 60.59±2.31 a 说明:数值为平均值±标准差。同列数值后不同小写字母表示不同根系径级存在显著差异(P<0.05) -
细根在土壤缠绕固结方面发挥重要作用。林火干扰后大量细根烧毁,土壤抗冲性能变化。对烧除前后泥沙流失量及抗冲系数进行配对t检验结果见表5。由表5可见:随着冲刷时间的延长,计划烧除和未烧除样地含沙量总体上呈下降趋势;冲刷过程前3 min,计划烧除样地产沙量较大,计划烧除样地产沙量显著大于未烧除样地(P<0.05);3 min后的冲刷产沙量均处于低水平平稳状态,无显著差异;烧除前后抗冲系数也有显著差异(P<0.05),未烧除样地抗冲系数是计划烧除样地的4.09倍,表明多年连续计划火烧使土壤抗冲性能显著降低。
表 5 计划烧除前后泥沙流失量及抗冲系数t检验
Table 5. Detection of sediment loss and impact coefficient t test before and after planning
样地类型 因素 不同冲刷历时泥沙流失量/g 抗冲系数/(L·g−1) 1 2 3 4 5 6 7 8 9 10 min 计划烧除样地 均值 1.561 0.236 0.101 0.079 0.058 0.041 0.037 0.031 0.024 0.022 9.217 标准差 0.159 0.064 0.012 0.028 0.014 0.019 0.019 0.018 0.018 0.014 0.908 未烧除样地 均值 0.264 0.060 0.032 0.031 0.031 0.022 0.021 0.017 0.013 0.011 37.496 标准差 0.098 0.006 0.015 0.002 0.012 0.002 0.001 0.002 0.004 0.003 4.739 显著性(双尾)P值 0.007 0.007 0.011 0.126 0.117 0.287 0.291 0.284 0.456 0.264 0.013 说明:当显著性P<0.05时,表示计划烧除前后泥沙流失量及抗冲系数有显著差异 林下凋落物的覆盖度和土壤内植物根系分布状况与土壤抗冲性能密切相关,土壤抗冲性与凋落物及根系参数相关性见表6。由表6可知:土壤抗冲系数与凋落物和根系指标均呈正相关关系,其中土壤抗冲系数与凋落物蓄积量、最大持水率、最大拦蓄率、有效拦蓄率、根长密度、根系生物量呈极显著正相关(P<0.01),与自然含水率呈显著正相关 (P<0.05)。因此,林内凋落物蓄积量越大,水分含量越高,越能有效减少地表径流的产生,减轻降雨对土壤的直接侵蚀。同时植被根系在土壤中交错穿插,够显著提髙土壤的抗冲性能,减少水土流失。长期计划烧除使凋落物无法蓄积,根系损毁,是导致抗冲性能大幅下降的主要原因。
表 6 土壤抗冲系数与凋落物及根系参数相关性
Table 6. Correlation between soil anti-rinse coefficients and litter and root parameters
参数 凋落物 根系 凋落物蓄积量 自然含水率 最大持水率 最大拦蓄率 有效拦蓄率 根长密度 根系生物量 土壤抗冲系数 0.965** 0.875* 0.952** 0.954** 0.936** 0.976** 0.970** 说明:*表示显著相关 (P<0.05);**表示极显著相关 (P<0.01) -
土壤团聚体是土壤的基本结构单位,对维持土体结构稳定有重要作用,是评价土壤抗蚀性的指标。林火干扰后,燃烧剩余灰烬堵塞土壤孔隙,从而增加土壤大团聚体。本研究显示:土壤表层粒径>2.00 mm机械稳定性团聚体增加,这与前人研究结果一致[24]。地表覆盖物的拦蓄作用以及植物根系的胶结作用能有效减缓降雨雨滴击溅造成的土壤侵蚀,提高水稳性团聚体含量,增强土体结构稳定性,减少土壤侵蚀。火烧后地表裸露,不利于大团聚体的形成,土壤抗蚀性能受到影响。不同火烧强度对土壤抗蚀性的影响不相同。大部分研究认为:林火干扰对多种林型的影响趋势基本一致,火烧后表层凋落物和植物细根被烧毁,土壤黏粒、粉粒结构减少,同时土壤微生物活动受到抑制,有机质含量减少,不利于土壤团粒结构的形成。随着火烧强度增加,土壤的抗蚀性下降[25−26]。也有研究认为:只有高强度的火烧才会使土壤团聚体破坏严重,中低强度火干扰对土壤团聚体影响不大,还能使土壤团聚体周围形成一层疏水薄膜,减缓土壤的润湿速率,提高土壤团聚体的抗水蚀能力[27]。本研究显示:在进行周期性计划烧除后,虽是低强度的地表火,但土壤表层水稳性团聚体减少,团聚体结构破坏率增大,随着土层深度的增加,影响逐渐减弱,这可能与火烧的频率有关。已有报道显示:低强度火烧形成的隔水层在2 a后消失[28]。在本研究中长期的林火干扰可能不利于土壤疏水层的形成,并且使土壤的结构和功能发生变化,再加上表层土壤失去凋落物的拦蓄作用,降雨发生时土壤更易遭到侵蚀。
-
土壤抗冲性变化是多种因素共同作用的结果。林下凋落物对土壤抗冲性的影响分为直接和间接2个部分:直接影响是凋落物层通过对降雨的截持来减轻降雨动能对土壤的侵蚀,间接影响是凋落物在分解过程中,微生物活性增强,产生大量有机质及胶结物质增强土体稳定性,通过改良土壤质地来减缓水流对土壤的冲刷作用[29−30]。众多研究表明:土壤凋落物覆盖下土壤容重和土壤孔隙度降低[31],土壤水文性质得到有效改良,输沙能力随凋落物蓄积量增加而递减[32],且凋落物半分解层的水土保持与水源涵养功能优于未分解层。本研究显示:周期性计划烧除将林地内凋落物烧毁,短期内无法大量蓄积,同时火烧后土壤微生物活动受到抑制,不利于凋落物分解及养分归还,凋落物层稀松,导致计划烧除样地地表大面积裸露,难以抵御降雨和水流的直接冲击与破坏,抗冲系数显著增大。这与陈璐等[33]对大兴安岭火烧迹地土壤抗冲性研究结果基本一致。根系通过对土壤的覆盖与固持,能有效防止土粒崩散,增强土壤结构稳定性,改善土壤理化性质,抑制水土流失[34−35],对土壤抗冲性有重要的积极作用。研究表明:林火干扰后,浅层土壤中的细根系燃烧殆尽,部分粗根系也因林火高温难以存活,无法起到固结土壤的作用,特别是重度火烧迹地由于林地裸露和雨水冲击,土壤形成抗水层,表面板结,孔隙度和渗透率降低,土壤结构严重退化,土壤物理性质受到长期影响[36]。也有研究认为:死亡的根系腐化会产生大量的胶结物质,促进土壤团聚体的形成,增强土壤的抗侵蚀能力[37]。本研究发现:与高强度森林火灾不同,计划烧除只将地表凋落物、草本植物及部分灌木烧毁,对云南松生长影响不大。低强度火烧后细根和部分粗根系被烧毁,但仍有一部分存活根系和新生根系,对土壤仍然起着一定的固持作用。本研究是在2021年年度计划烧除结束半年后采样,部分灌木和草本植物恢复生长,在一定程度上减轻了火烧对土壤抗冲性的影响。但长期林火干扰使林内灌草覆盖度远低于未烧除样地,地表裸露面积增加,土壤中根系含量减少,根系间网络串联、根土黏结和生物化学作用减弱,微生物对烧死根系分解速率减缓,可能使其无法产生胶结物质。同时,计划烧除导致降雨发生时雨水与土壤直接接触,土壤抗水蚀能力下降,大量表层土壤易被冲走,存在土壤侵蚀风险。
-
在云南松林地内开展周期性计划烧除后,大量燃烧剩余灰烬堵塞土壤孔隙,使土壤变得紧实。干筛处理下,计划烧除样地粒径>2.00 mm团聚体高于未烧除样地,团聚体平均质量直径、团聚体平均几何直径也显著增大;湿筛处理下,计划烧除样地粒径<0.25 mm团聚体较未烧除样地有所增加,团聚体平均质量直径、团聚体平均几何直径、土壤抗蚀指数均显著低于未烧除样地,土壤团聚体结构破坏率增大,土壤抗水蚀性能减弱。随着土层深度的增加,火干扰对土壤抗蚀性的影响逐渐减弱,对深层土壤基本无影响。
高频率、低强度的地表火对林地内凋落物和植被根系有一定的影响,地表凋落物持水性能减弱,土壤中根系减少,土壤失去凋落物的覆盖和植被根系的固持作用,土壤抗冲性能显著低于未烧除样地。
Effect of prescribed burning on soil anti-erodibility and anti-scourability of Pinus yunnanensis forest
-
摘要:
目的 计划烧除是森林预防火灾的重要措施之一。探究计划烧除对云南松Pinus yunnanensis林地土壤抗蚀性和抗冲性的影响,为森林的可持续经营以及优化林区计划烧除作业提供可靠的科学依据。 方法 以滇中高原云南松林计划烧除样地为研究对象,通过野外调查取样与室内实验分析相结合的方法,系统研究了计划烧除对云南松林土壤抗蚀性和抗冲性的影响。 结果 ①周期性计划烧除后,土壤0~40 cm土层中粒径>2.00 mm的机械稳定性团聚体含量增加,粒径>2.00 mm的水稳性团聚体含量减少,水稳性团聚体平均质量直径和平均几何直径减小,土壤团聚体结构破坏率增大,抗蚀指数减小,40~60 cm土层中土壤抗蚀性指标无明显变化。②计划烧除后地表凋落物蓄积量减小,凋落物持水性能与拦蓄能力减弱,植物根系被烧毁。周期性计划烧除后凋落物和根系减少是林地内土壤抗冲系数减小的主要原因。 结论 计划烧除使土壤表层抗水蚀能力下降。随着土层深度增加,计划烧除对土壤抗蚀性的影响逐渐减小,火烧后地表凋落物和植被根系减少,抗冲性能减弱。图1表6参37 Abstract:Objective Prescribed burning is an essential measure for forest fire prevention. The purpose of this study is to explore the impact of prescribed burning on soil anti-erodibility and anti-scourability of Pinus yunnanensis forest, so as to provide scientific basis for sustainable forest management and optimization of prescribed burning in forest areas. Method The sample plots of P. yunnanensis forest in central Yunnan Plateau were selected as the research objects. The effects of prescribed burning on soil anti-erodibility and anti-scourability of P. yunnanensis forest were systematically studied by combining field investigation and sampling with indoor experimental analysis. Result (1) After prescribed burning, the content of mechanically stable aggregates(>2.00 mm) in 0−40 cm soil layer increased and the content of waterstable aggregates (>2.00 mm) decreased. The mean mass diameter and mean geometric diameter of the water stable aggregates decreased. The damage rate of soil aggregate structure increased, and the erosion resistance index decreased. The soil erosion resistance index of 40−60 cm soil layer did not change significantly. (2)After the prescribed burning, the litter accumulation decreased and the water holding capacity and storage capacity of the litter weakened, and the plant root system was burned. The decrease of litter and root system after periodic burning was the main reason for the decrease of soil erosion resistance coefficient in forest land. Conclusion Prescribed burning reduces water erosion resistance of soil surface. With the increase of soil depth, the impact of planned burning on soil erosion resistance gradually decreases. After prescribed burning, the surface litter and vegetation roots decrease and the erosion resistance weakens. [Ch, 1 fig. 6 tab. 37 ref.] -
Key words:
- prescribed burning /
- Pinus yunnanensis forest /
- anti-erodibility /
- anti-scourability
-
表 1 土壤团聚体稳定性特征
Table 1. Soil gathering body stability characteristics
土层深度/cm 计划烧除样地 未烧除样地 平均质量直径/mm 平均几何直径/mm 平均质量直径/mm 平均几何直径/mm 干筛 湿筛 干筛 湿筛 干筛 湿筛 干筛 湿筛 0~10 3.18±0.09 A 1.67±0.11 B 1.61±0.05 A 0.65±0.02 B 2.80±0.15 B 2.02±0.10 A 1.28±0.07 B 0.77±0.07 A 10~20 2.86±0.08 B 1.30±0.11 C 1.24±0.07 B 0.47±0.03 C 2.73±0.04 C 1.41±0.00 B 1.22±0.08 B 0.51±0.01 C 20~40 2.63±0.03 C 1.14±0.03 D 1.04±0.02 C 0.42±0.01 D 2.56±0.13 D 1.23±0.05 C 1.05±0.05 C 0.44±0.01 D 40~60 2.50±0.07 E 0.97±0.01 E 0.99±0.04 C 0.34±0.01 E 2.45±0.06 E 0.99±0.01 E 0.97±0.06 C 0.34±0.01 E 说明:数值为平均值±标准差。同行数值后不同大写字母表示不同处理间差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 土壤团聚体结构破坏率及抗蚀指数
Table 2. Destruction rate and corrosion index of soil cluster structure
土层深
度/cm计划烧除样地 未烧除样地 粒径>0.25 mm土壤团聚体比例/% 团聚体结构
破坏率/%抗蚀指
数/%粒径>0.25 mm土壤团聚体比例/% 团聚体结构
破坏率/%抗蚀指
数/%干筛 湿筛 干筛 湿筛 0~10 86.42±0.97 A 65.07±0.98 B 21.35±1.84 A 64.44±2.95 B 81.87±2.20 B 69.24±2.82 A 12.62±2.83 B 73.78±5.69 A 10~20 77.82±3.28 C 55.61±1.54 D 22.21±2.46 A 52.89±4.91 C 79.26±3.28 C 58.52±1.62 C 20.74±2.35 A 58.67±6.53 B 20~40 73.90±0.25 D 53.80±0.64 D 20.10±0.77 A 37.38±8.02 D 73.62±2.14 D 53.90±0.69 D 19.73±1.45 A 38.67±8.22 D 40~60 73.04±0.68 D 43.75±1.04 E 29.29±0.73 C 27.56±5.48 E 73.36±1.78 D 44.14±0.61 E 29.22±1.17 C 28.00±5.66 E 说明:数值为平均值±标准差。同行数值后不同大写字母表示不同处理间差异显著(P<0.05)
下载: 导出CSV
表 3 凋落物蓄积量及持水性能
Table 3. Destiny storage volume and water holding performance
样地类型 凋落物层 凋落物蓄积量/
(t·hm−2)最大持水量/
(t·hm−2)最大持水率/% 最大拦蓄量/
(t·hm−2)最大拦蓄率/% 有效拦蓄率/% 计划烧除样地 未分解层 2.95±0.18 c 4.80±0.64 c 163.34±24.23 d 3.71±0.64 c 126.27±23.48 d 89.19±22.81 d 半分解层 0.87±0.05 d 2.55±0.11 d 294.26±19.16 b 2.00±0.11 d 230.49±17.70 b 166.72±16.88 b 合计 3.82±0.23 7.35±0.75 193.13±22.79 5.71±0.75 149.98±21.97 106.83±21.25 未烧除样地 未分解层 5.91±0.16 a 13.90±0.20 a 235.27±7.89 c 11.19±0.35 a 189.38±10.05 c 143.50±12.21 c 半分解层 3.43±0.21 b 11.67±0.43 b 340.48±8.04 a 9.77±0.35 b 285.12±9.07 a 229.76±12.96 a 合计 9.35±0.20 26.55±1.30 283.93±9.21 21.93±1.24 234.52±9.71 185.12±10.23 说明:数值为平均值±标准差。同列数值后不同小写字母表示2种样地类型凋落物蓄积量存在显著差异(P<0.05)
下载: 导出CSV
表 4 根长密度及生物量
Table 4. Root long density and biomass
根系径级/mm 计划烧除样地 未烧除样地 根长密度/
(×10−3cm·cm−3)根系生物量/
(×10−3g·cm−3)占根总生物量百分比/% 根长密度/
(×10−3cm·cm−3)根系生物量/
(×10−3g·cm−3)占根总生物量百分比/% >2.00 9.70±1.10 e 0.28±0.02 e 35.20±4.14 b 29.17±1.86 d 0.76±0.08 b 24.64±2.51 c 0.50~2.00 24.73±2.50 d 0.20±0.03 e 24.82±3.32 c 46.10±4.17 c 0.46±0.04 c 14.77±1.38 d <0.50 59.33±7.41 b 0.33±0.03 d 39.99±1.37 b 162.80±14.62 a 1.89±0.07 a 60.59±2.31 a 说明:数值为平均值±标准差。同列数值后不同小写字母表示不同根系径级存在显著差异(P<0.05)
下载: 导出CSV
表 5 计划烧除前后泥沙流失量及抗冲系数t检验
Table 5. Detection of sediment loss and impact coefficient t test before and after planning
样地类型 因素 不同冲刷历时泥沙流失量/g 抗冲系数/(L·g−1) 1 2 3 4 5 6 7 8 9 10 min 计划烧除样地 均值 1.561 0.236 0.101 0.079 0.058 0.041 0.037 0.031 0.024 0.022 9.217 标准差 0.159 0.064 0.012 0.028 0.014 0.019 0.019 0.018 0.018 0.014 0.908 未烧除样地 均值 0.264 0.060 0.032 0.031 0.031 0.022 0.021 0.017 0.013 0.011 37.496 标准差 0.098 0.006 0.015 0.002 0.012 0.002 0.001 0.002 0.004 0.003 4.739 显著性(双尾)P值 0.007 0.007 0.011 0.126 0.117 0.287 0.291 0.284 0.456 0.264 0.013 说明:当显著性P<0.05时,表示计划烧除前后泥沙流失量及抗冲系数有显著差异
下载: 导出CSV
表 6 土壤抗冲系数与凋落物及根系参数相关性
Table 6. Correlation between soil anti-rinse coefficients and litter and root parameters
参数 凋落物 根系 凋落物蓄积量 自然含水率 最大持水率 最大拦蓄率 有效拦蓄率 根长密度 根系生物量 土壤抗冲系数 0.965** 0.875* 0.952** 0.954** 0.936** 0.976** 0.970** 说明:*表示显著相关 (P<0.05);**表示极显著相关 (P<0.01)
下载: 导出CSV
-
[1] 苏佳佳, 刘志华, 焦珂伟, 等. 气候变化对中国林火干扰空间格局的影响[J]. 生态学杂志, 2021, 40(12): 3810 − 3821. SU Jiajia, LIU Zhihua, JIAO Kewei, et al. Effects of climate change on spatial pattern on forest fire regimes in China [J]. Chinese Journal of Ecology, 2021, 40(12): 3810 − 3821. [2] 刘小菊, 潘存德, 褚江涛, 等. 喀纳斯泰加林林下草本层地上碳密度对林火烈度的火后时间响应[J]. 水土保持研究, 2022, 29(3): 98 − 105. LIU Xiaoju, PAN Cunde, CHU Jiangtao, et al. Response of aboveground carbon density of herbaceous layer to post-fire time of fire severity in Kanas, Xinjiang [J]. Research in Soil and Water Conservation, 2022, 29(3): 98 − 105. [3] 魏书精, 罗碧珍, 孙龙, 等. 黑龙江省温带森林火灾碳排放的计量估算[J]. 生态学报, 2014, 34(11): 3048 − 3063. WEI Shujing, LUO Bizhen, SUN Long, et al. Estimates of carbon emissions caused by forest fires in the temperate climate of Heilongjiang Province, China, from 1953 to 2012 [J]. Acta Ecologica Sinica, 2014, 34(11): 3048 − 3063. [4] 周梅, 杨洪琴, 王广山, 等. 计划烧除对大兴安岭冻土、湿地环境的影响分析[J]. 内蒙古农业大学学报, 2008, 29(2): 73 − 75. ZHOU Mei, YANG Hongqin, WANG Guangshan, et al. The analysis of influence of prescribed burning on permafrost and wetland in Daxinganling Mountian [J]. Journal of Inner Mongolia Agricultural University, 2008, 29(2): 73 − 75. [5] BROCKWAY D G, GATEWOOD R G, PARIS R B. Restoring fire as an ecological process in shortgrass prairie ecosystems: initial effects of prescribed burning during the dormant and growing seasons [J]. Journal of Environmental Management, 2002, 65(2): 135 − 152. [6] 张文文, 王秋华, 龙腾腾, 等. 周期性计划烧除对森林生态系统的影响研究综述[J]. 西南林业大学学报(自然科学), 2021, 41(4): 181 − 188. ZHANG Wenwen, WANG Qiuhua, LONG Tengteng, et al. A review of the effects of periodic prescribed burning on forest ecosystems [J]. Journal of Southwest Forestry University (Natural Sciences) , 2021, 41(4): 181 − 188. [7] 徐伟恒, 黄邵东, 杨磊, 等. 滇东北地区云南松地表可燃物载量及火强度研究[J]. 西部林业科学, 2019, 48(4): 19 − 26. XU Weiheng, HUANG Shaodong, YANG Lei, et al. Surface fuel load and fire intensity of Pinus yunnanensis in northeast Yunnan Province [J]. Journal of West China Forestry Science, 2019, 48(4): 19 − 26. [8] 高仲亮, 舒立福, 王明玉, 等. 云南松烧除火蔓延速率预测模型研究[J]. 安全与环境学报, 2016, 16(4): 82 − 86. GAO Zhongliang, SHU Lifu, WANG Mingyu, et al. Model for predicting the forest fire spreading speed based on the prescriptive combustion of Pinus yunnanensis [J]. Journal of Safety and Environment, 2016, 16(4): 82 − 86. [9] 田荣, 陈奇伯, 黎建强, 等. 计划烧除对云南松林土壤理化性质及可蚀性的影响[J]. 西北林学院学报, 2021, 36(2): 11 − 16. TIAN Rong, CHEN Qibo, LI Jianqiang, et al. Effects of prescribed burning on soil physicochemical properties and erodibility in Pinus yunnanensis forests [J]. Journal of Northwest Forestry University, 2021, 36(2): 11 − 16. [10] 朱利英, 赵春章, 莫旭, 等. 计划烧除攀枝花苏铁林区地面覆盖物对苏铁生长和土壤理化性质的影响[J]. 应用与环境生物学报, 2012, 18(3): 381 − 390. ZHU Liying, ZHAO Chunzhang, MO Xu, et al. Effects of prescribed burning on plant growth and soil properties of a Cycas panzhihuacycasis forest [J]. Chinese Journal of Applied and Environmental Biology, 2012, 18(3): 381 − 390. [11] 龚顺清, 陈奇伯, 黄新会, 等. 多年连续计划烧除对云南松林坡面水土流失的影响[J]. 水土保持研究, 2022, 29(3): 18 − 24. GONG Shunqing, CHEN Qibo, HUANG Xinhui, et al. Prescribed burninng effects on slope runoff and soil loss in a Pinus yunnanensis forest of Yunnan [J]. Research in Soil and Water Conservation, 2022, 29(3): 18 − 24. [12] PACHECO R M, CLARO J. Prescribed burning as a cost-effective way to address climate change and forest management in Mediterranean countries [J]. Annals of Forest Science, 2021, 78(4): 1 − 11. [13] FONTURBEL T, CARRERA N, VEGA J A, et al. The effect of repeated prescribed burning on soil properties: a review[J/OL]. Forests, 2021, 12(6): 767[2022-03-25]. doi: 10.3390/f12060767. [14] TAHMASBIAN I, XU Zhihong, NGUYEN T T N, et al. Short-term carbon and nitrogen dynamics in soil, litterfall and canopy of a suburban native forest subjected to prescribed burning in subtropical Australia [J]. Journal of Soil &Sediments, 2019, 19(12): 3969 − 3981. [15] 王秋华, 单保君, 龚家平, 等. 滇中地区云南松纯林计划烧除研究[J]. 江西农业大学学报, 2018, 40(2): 235 − 240. WANG Qiuhua, SHAN Baojun, GONG Jiaping, et al. A study on prescribed burning in pure forest of Pinus yunnanensis Franch in central Yunnan Province [J]. Acta Agriculturae Universitatis Jiangxiensis, 2018, 40(2): 235 − 240. [16] 王健敏, 李帅锋, 徐凡迪, 等. 计划烧除对思茅松天然林群落结构与物种多样性的影响[J]. 西北林学院学报, 2020, 35(1): 62 − 67. WANG Jianmin, LI Shuaifeng, XU Fandi, et al. Effects of prescribed burning on the community structure and species diversity in Pinu skesiya var. langbiannesis primary forest [J]. Journal of Northwest Forestry University, 2020, 35(1): 62 − 67. [17] 罗斯生, 罗碧珍, 魏书精, 等. 森林火灾对马尾松次生林土壤理化性质的影响[J]. 生态环境学报, 2020, 29(11): 2141 − 2152. LUO Sisheng, LUO Bizhen, WEI Shujing, et al. Effects of forest fires on soil physical and chemical properties in secondary forest of Pinus massoniana [J]. Ecology and Environmental Sciences, 2020, 29(11): 2141 − 2152. [18] 沈颖, 秦涛, 郭银花, 等. 林火对山西太岳山油松林土壤微生物-酶活性的短期影响[J]. 北京林业大学学报, 2022, 44(4): 76 − 85. SHEN Ying, QIN Tao, GUO Yinghua, et al. Short-term effects of forest fire on soil microorganism and enzyme activities of Pinus tabuliformis forest in Taiyue Mountain, Shanxi Province of northern China [J]. Journal of Beijing Forestry University, 2022, 44(4): 76 − 85. [19] 范世才, 方晰, 王振鹏, 等. 植被恢复过程土壤基质改良对湘南紫色土抗蚀性的影响[J]. 水土保持学报, 2021, 25(6): 22 − 30. FAN Shicai, FANG Xi, WANG Zhenpeng, et al. Effect of soil matrix improvement measure on erosion resistance of purple soil during vegetation restoration southern Hunan Province, China [J]. Journal of Soil and Water Conservation, 2021, 25(6): 22 − 30. [20] 杨晓霞, 赵锦梅, 张雪, 等. 祁连山东段山地典型灌丛枯落物及土壤水源涵养功能研究[J]. 干旱区地理, 2022, 45(1): 197 − 207. YANG Xiaoxia, ZHAO Jinmei, ZHANG Xue, et al. Litter and soil water conservation function of typical shrubs in eastern Qilian Mountains [J]. Arid Land Geography, 2022, 45(1): 197 − 207. [21] 孙丽丽, 査轩, 黄少燕, 等. 花岗岩红壤区不同治理模式土壤抗冲性因素试验[J]. 水土保持学报, 2019, 33(5): 34 − 40. SUN Lili, ZHA Xuan, HUANG Shaoyan, et al. Experimental on soil anti-scourbility factors under different management modes in granite red soil region [J]. Journal of Soil and Water Conservation, 2019, 33(5): 34 − 40. [22] 郭明明, 王文龙, 史倩华, 等. 黄土高塬沟壑区退耕地土壤抗冲性及其与影响因素的关系[J]. 农业工程学报, 2016, 32(10): 129 − 136. GUO Mingming, WANG Wenlong, SHI Qianhua, et al. Soil anti-scouribility of abandoned land and its relationship with influencing factors in Loess Plateau Gully region [J]. Transactions of the Chinese Society of Agricultural Engineering, 2016, 32(10): 129 − 136. [23] 刘均阳, 周正朝, 苏雪萌. 植物根系对土壤团聚体形成作用机制研究回顾[J]. 水土保持学报, 2020, 34(3): 267 − 273. LIU Junyang, ZHOU Zhengchao, SU Xuemeng. Review of the mechanism of root system on the formation of soil aggregates [J]. Journal of Soil and Water Conservation, 2020, 34(3): 267 − 273. [24] 孙龙, 窦旭, 胡同欣. 林火对森林生态系统碳氮磷生态化学计量特征影响研究进展[J]. 南京林业大学学报(自然科学版), 2021, 45(2): 1 − 9. SUN Long, DOU Xu, HU Tongxin. Research progress on the effects of forest fire on forest ecosystem C-N-P ecological stoichiometry characteristics [J]. Journal of Nanjing Forestry University (Natural Sciences Edition) , 2021, 45(2): 1 − 9. [25] BASTIAS B A, ZHUANG Zhiqun, BLUMFIELD T, et al. Influence of repeated prescribed burning on the soil fungal community in an eastern Australian wet sclerophyll forest [J]. Soil Biology and Biochemistry, 2006, 38(12): 3492 − 3501. [26] 杨瀛, 胡卸文, 王严, 等. 火后泥石流流域坡面侵蚀物源动储量估算方法探讨[J]. 工程地质学报, 2021, 29(1): 151 − 161. YANG Ying, HU Yuwen, WANG Yan, et al. Preliminary study on methods to calculate dynamic reserves of slope erosioning materials transported by post-fire debris flow [J]. Journal of Engineering Geology, 2021, 29(1): 151 − 161. [27] MATAIX-SOLERA J, DOERR S H. Hydrophobicity and aggregate stability in calcareous topsoils from fire-affected pine forests in southeastern Spain [J]. Geoderma, 2004, 118(1/2): 77 − 88. [28] POLLET J, OMI P N. Effect of thinning and prescribed burning on crown fire severity in ponderosa pine forests [J]. International Journal of Wildland Fire, 2002, 11(1): 1 − 10. [29] 赵春红, 高建恩, 徐震. 牧草调控绵沙土坡面侵蚀机理[J]. 应用生态学报, 2013, 24(1): 113 − 121. ZHAO Chunhong, GAO Jianen, XU Zhen. Mechanisms of grass in slope erosion control in loess sandy soil region of northwest China [J]. Chinese Journal of Applied Ecology, 2013, 24(1): 113 − 121. [30] 王龙, 张连科, 马伊明, 等. 工程扰动对黄土抗蚀性与抗冲性的影响[J]. 水土保持学报, 2021, 35(4): 14 − 20. WANG Long, ZHANG Lianke, MA Yiming, et al. Influence of engineering disturbance on soil anti-erodibility and anti-scourability of loess [J]. Journal of Soil and Water Conservation, 2021, 35(4): 14 − 20. [31] 冯宜明, 李毅, 曹秀文, 等. 甘肃亚高山云杉人工林土壤特性及水源涵养功能对林分密度的响应特征[J]. 自然资源学报, 2018, 33(9): 1529 − 1541. FENG Yiming, LI Yi, CAO Xiuwen, et al. Effects of stand density on soil properties and water conservation function of Picea asperata plantation in the subalpine region of Gansu Province [J]. Journal of Natural Resources, 2018, 33(9): 1529 − 1541. [32] 孙佳美, 余新晓, 李瀚之, 等. 模拟降雨下枯落物调控坡面产流产沙过程及特征研究[J]. 水利学报, 2017, 48(3): 341 − 350. SUN Jiamei, YU Xinxiao, LI Hanzhi, et al. Runoff and sediment yield process and characteristics research on litter slopes in simulated rainfall [J]. Journal of Hydraulic Engineering, 2017, 48(3): 341 − 350. [33] 陈璐, 辛颖, 赵雨森, 等. 大兴安岭火烧迹地土壤抗冲性研究[J]. 森林工程, 2015, 31(3): 21 − 24. CHEN Lu, XIN Ying, ZHAO Yusen, et al. Study on soil anti-scourability of burned area in Greater Xing’ an Mountains [J]. Forest Engineering, 2015, 31(3): 21 − 24. [34] 胡卸文, 侯羿腾, 王严, 等. 火烧迹地土壤根系特征及其对抗剪强度的影响[J]. 水文地质工程地质, 2019, 46(5): 106 − 112. HU Yuwen, HOU Yiteng, WANG Yan, et al. Root characteristics and its influences on shear strength in burned areas [J]. Hydrogeology &Engineering Geology, 2019, 46(5): 106 − 112. [35] 潘天辉, 杜峰, 王月. 陕北黄土区护坡植物根系分布和抗剪增强分析[J]. 水土保持研究, 2020, 27(3): 357 − 363. PAN Tianhui, DU Feng, WANG Yue. Analysis of root distribution and shear strengths of slope protection plants in the loess region of northern Shanxi [J]. Research in Soil and Water Conservation, 2020, 27(3): 357 − 363. [36] 秦倩倩, 刘艳红. 重度火烧干扰下的森林土壤功能[J]. 应用与环境生物学报, 2021, 27(2): 503 − 512. QIN Qianqian, LIU Yanhong. Forest soil function after severe fire disturbance [J]. Chinese Journal of Applied &Environmental Biology, 2021, 27(2): 503 − 512. [37] 王金成, 井明博, 周天林, 等. 黄土高原沟壑区人工沙棘林生长年限对土壤微生物组成的影响[J]. 干旱地区农业研究, 2015, 33(2): 253 − 260. WANG Jincheng, JING Mingbo, ZHOU Tianlin, et al. Effects of different aged artificial seabuckthorn forests on soil microbial-composition in the gully region loess plateau, China [J]. Agricultural Research in the Arid Areas, 2015, 33(2): 253 − 260. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220321
 点击查看大图
点击查看大图
计量
- 文章访问数: 1762
- HTML全文浏览量: 456
- PDF下载量: 65
- 被引次数: 0

