




-
杨柳科Salicaceae柳属Salix植物种类繁多,有450~520种;分布广泛,主要分布在北半球[1],在中国属于原生树种;适应性强,耐湿,抗旱,耐低温,抗风性好,又具有很高的观赏价值。柳蓝叶甲Plagiodera versicolora又名柳圆叶甲,属鞘翅目Coleoptera叶甲科Chrysomelidae,是柳树主要食叶害虫,易在柳树较多或集中的地方大量发生。目前柳蓝叶甲仍以化学防治为主,因此对环境污染较大[2]。如何安全有效地预防和控制柳蓝叶甲的危害成为亟待解决的问题。植物抗虫性是植物与昆虫共同进化过程中形成的遗传性状,具有专一性、累积性、持续性、与环境的协调性、与其他防治措施的兼容性等特点,提高植物抗性是害虫综合防治策略中的一项重要措施[3-5]。在促使植物具有抗虫性的诸多因素中,化学因素起着重要作用[6]。穆丹等[7]研究证明:植物的气味能够有效调节和诱导柳蓝叶甲的行为。MATSUDA等[8]研究发现:绿原酸对某些柳树甲虫具有拮抗作用。IKONEN等[9]研究表明:叶甲拒食的柳叶中有高含量的绿原酸和水杨酸。由于技术的限制,植物次生代谢物基因转录和调控机制仍然没有得到解决。代谢组学技术在理解代谢网络和转录基因功能方面起着关键作用[10-14]。随着质谱、核磁共振等相关技术的迅速发展,代谢组学与其他组学技术的结合成为代谢组学研究的热点[12-14]。本研究选取旱柳S. matsudana、 黄花柳S. caprea、 垂柳S. babylonica以及三蕊柳S. triandra的3个无性系进行抗虫性测定,根据结果将柳树分成抗虫组与非抗虫组,再通过代谢组学技术分析抗虫组与非抗虫组之间的差异次生代谢物,为选育抗虫性强的柳树品种提供依据。
-
选取三蕊柳的3个无性系W10、W4、W21,对照包括旱柳K24,黄花柳HHL,垂柳BD3,共6个材料。
-
在养虫盒底部放入湿润的滤纸,在每个供试材料上摘取完整嫩叶,用浸湿的棉球包裹叶柄以防止叶片失水,把嫩叶放于湿润的滤纸上,接入相同龄期的30只柳蓝叶甲幼虫,每个材料均设3组重复,每组5片叶,7 d后观察幼虫存活数量并计算存活率。
-
植物代谢组学是一种高通量、无偏倚的综合分析[15]。将供试样品根据抗虫性指标测定的结果分为非抗虫组和抗虫组,采集新鲜叶片,液氮速冻用箔纸包好,−80 ℃冰箱保存备用。基于UPLC-Q-TOF-MS和GC-MS的代谢组学分析方法,对不同样品的代谢谱进行分析[16]。OPLS-DA是代谢组学数据分析中常用的分析方法,是PLS-DA的扩展[17],通过去除与分组无关的系统自身差异筛选代谢差异物,使分离达到最大化[18],再通过差异聚类热图分析[19]对样品和指标进行分类。
-
使用Excel 2010对数据进行整理;使用SPSS 20.0对数据进行独立样本t检验以及相关性分析;使用Origin 8.0作图。用OPLS-DA、单维分析(t检验)和差异倍数筛选组间差异代谢物。筛选获得的差异代谢物用
和 https://metlin.scripps.edu/ 数据库进行定性。 -
由图1可见:喂食黄花柳、旱柳、垂柳的幼虫平均存活率分别为90%、98%、93%,其中,喂食旱柳的幼虫存活率最高。喂食三蕊柳3个无性系的幼虫存活率均为0,在接虫后的第4天全部死亡。因此,把柳树分类非抗虫组(旱柳K24、黄花柳HHL和垂柳BD3)以及抗虫组(三蕊柳的3个无性系W10、W4和W21)。
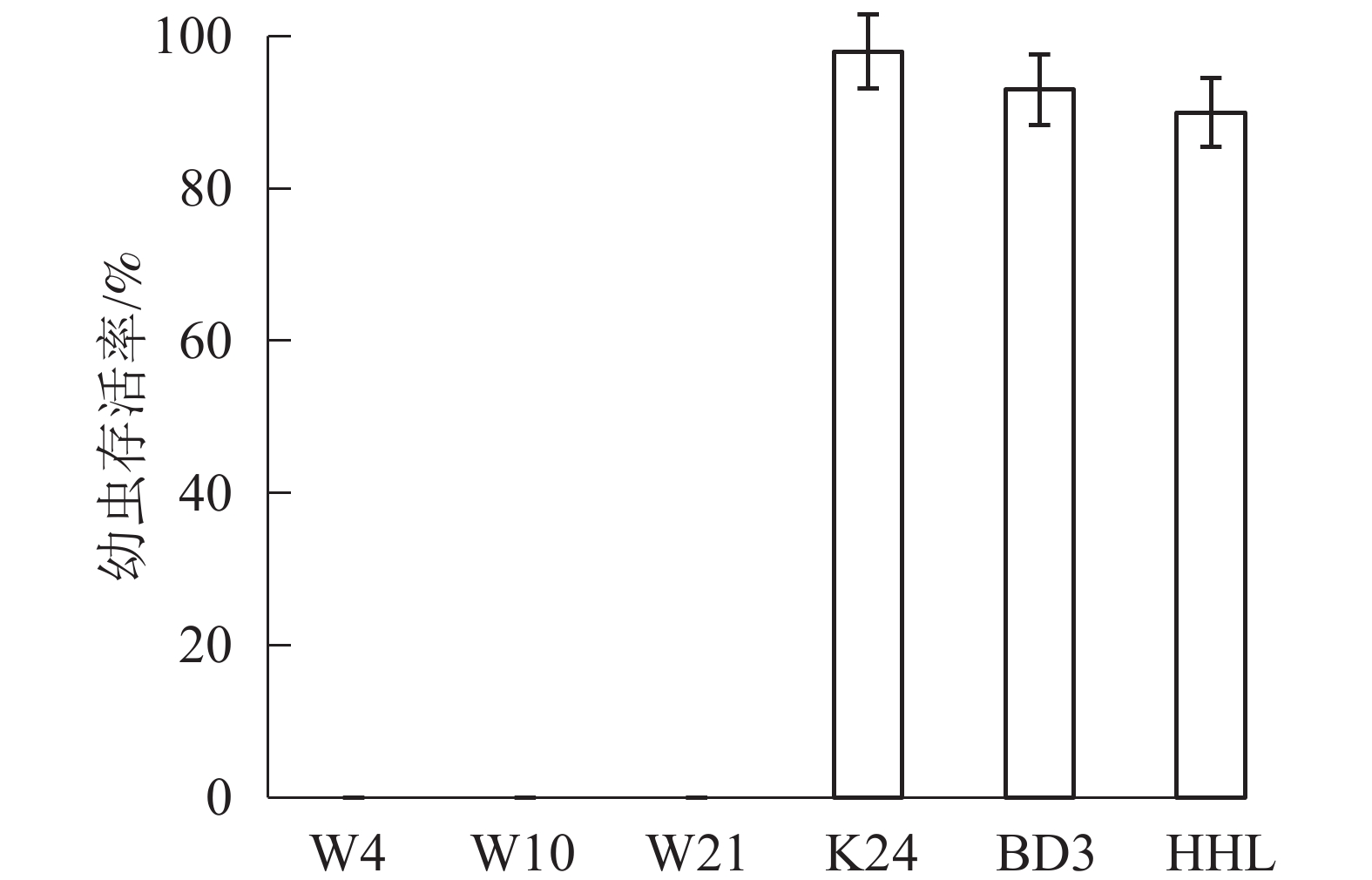
图 1 室内幼虫存活率
Figure 1. Survival rate of indoor larvae
-
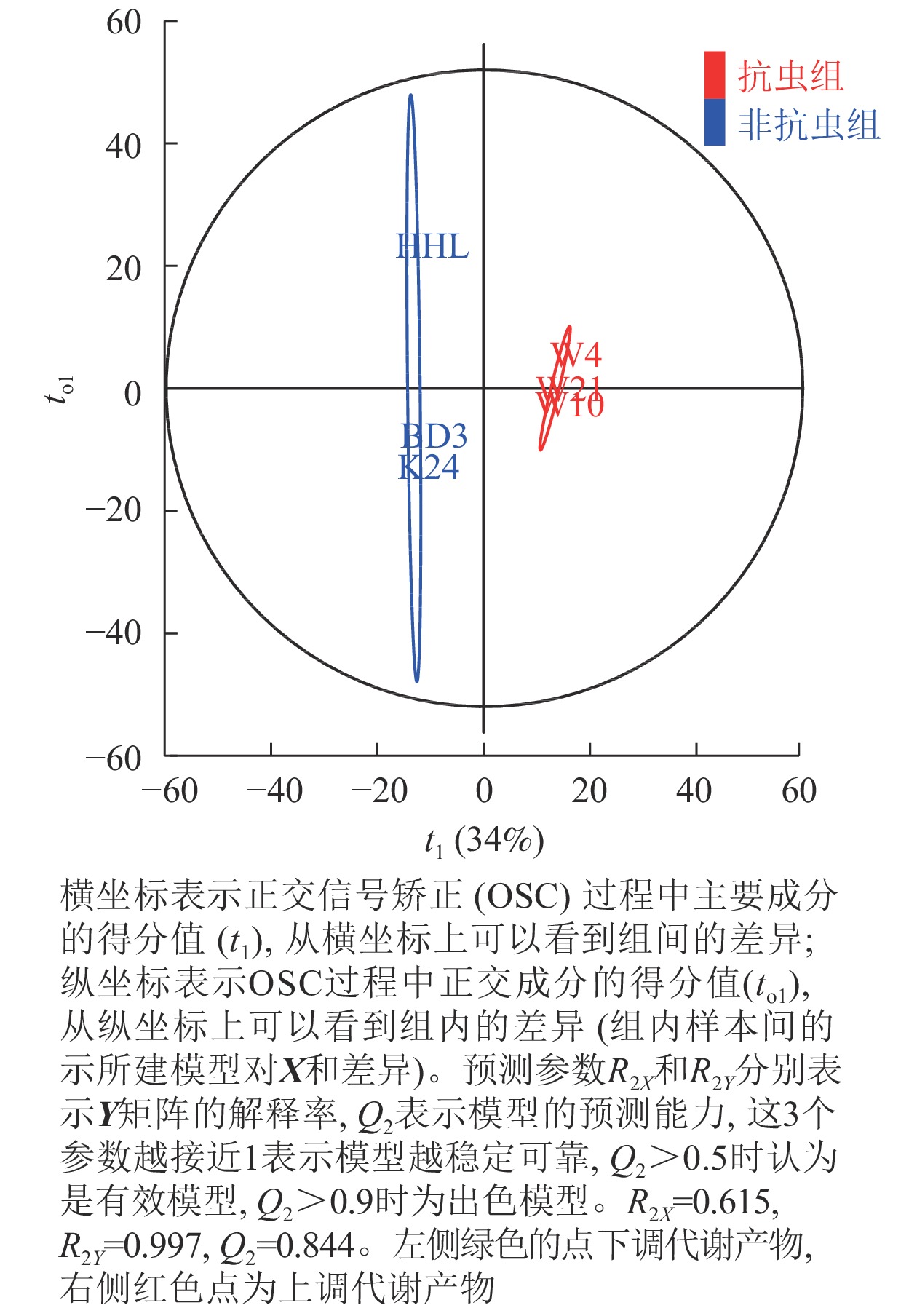
由OPLS-DA得分图(图2)显示:不同抗性组合模型参数中R2X=0.615,说明该模型对自变量X的解释程度为61.5%;R2Y=0.997,表示对分类变量Y的解释程度为99.7%;Q2=0.844,表示该模型对样本变量的预测程度为84.4%,说明OPLS-DA模型稳定性良好且预测能力较强。OPLS-DA的得分图显示非抗虫组与抗虫组的数据点在OPLS1上完全区分,表明两者存在明显化学差异。
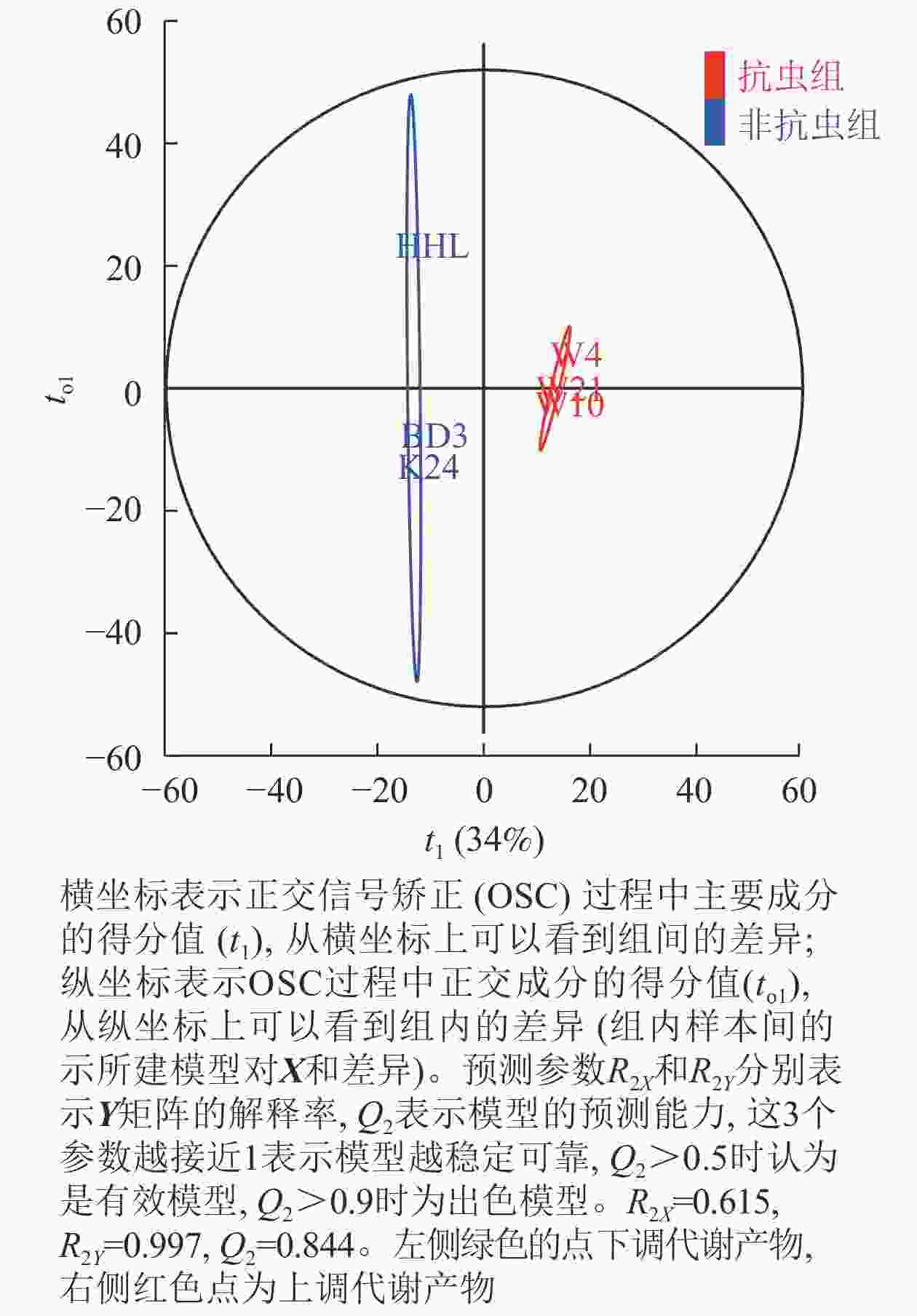
图 2 非抗虫组与抗虫组OPLS-DA得分图
Figure 2. OPLS-DA score chart of non-insect resistant group and insect resistant group
-
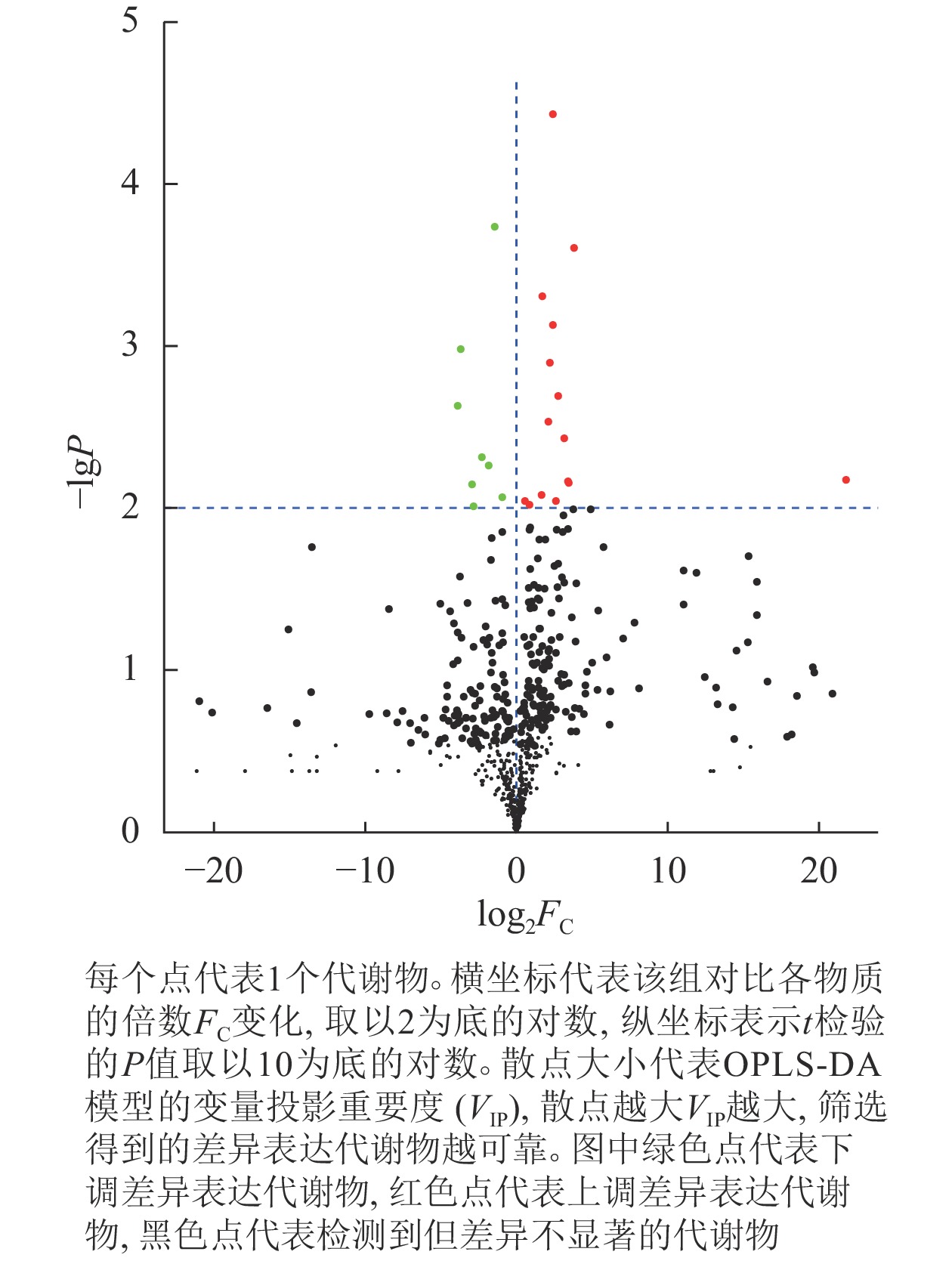
从图3可以看出:在指定阈值P<0.01时,共筛选出23种显著差异代谢物。结果显示:抗虫组与非抗虫组比较,其中上调的差异代谢物数目为15种,下调的代谢物数目为8种。
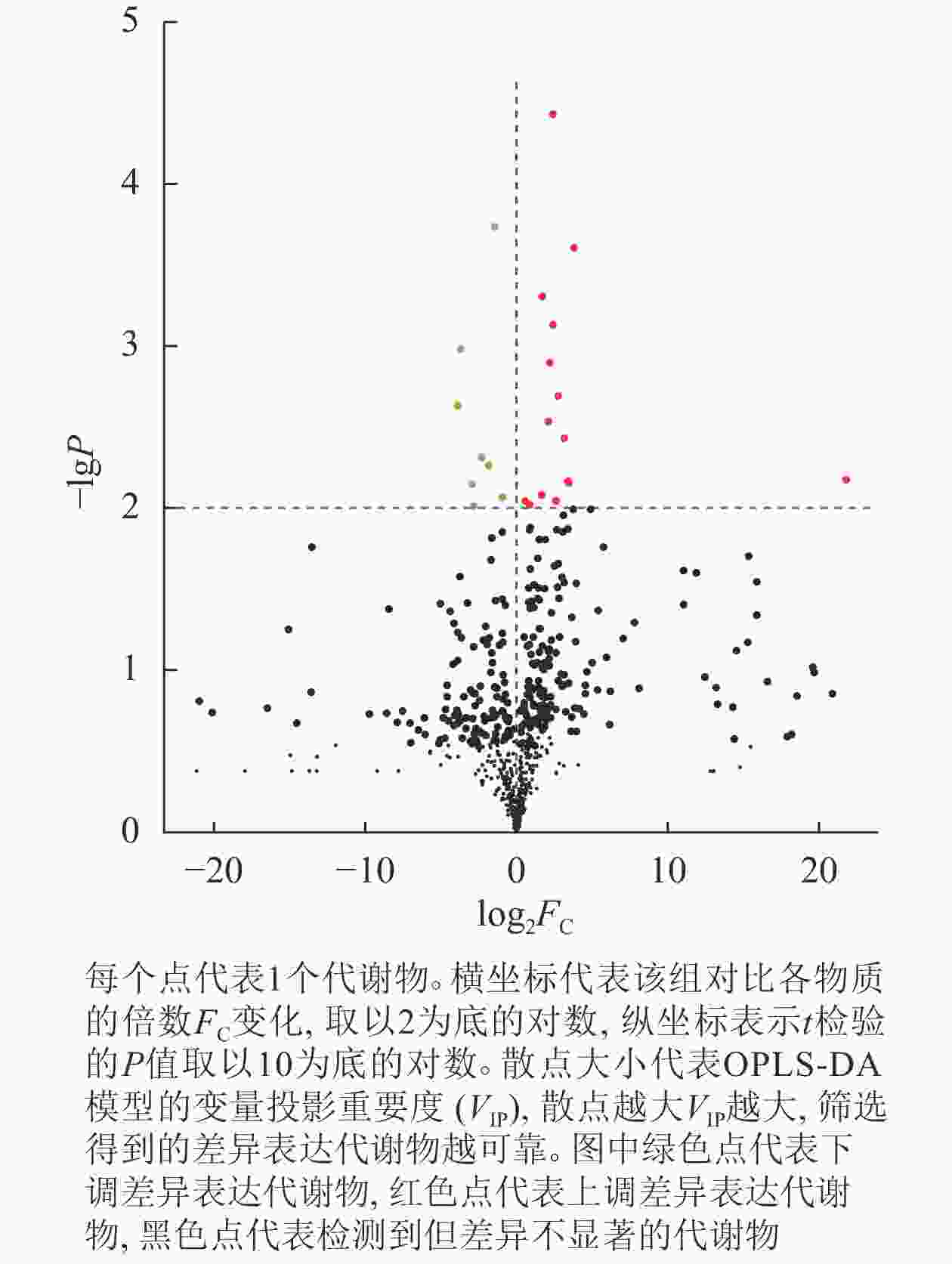
图 3 差异代谢物火山图
Figure 3. Volcano map of differential metabolites
-
不同柳树样本中共检测到651个物质信号,经过标准样品数据库和参考文献比对,对有生物学重复的,采取将差异倍数,t检验的P值和OPLS-DA模型的变量投影重要度(VIP)相结合的方法来筛选差异代谢物,筛选的标准为差异倍数FC>1,t检验P<0.01和VIP>1。由表1可见:经筛选后,非抗虫组与抗虫组共筛选出23种主要的差异代谢产物。其中阿魏酰胍丁胺、抗坏血酸、没食子酸、绿茶儿茶酚、没食子儿茶素、杨梅苷、葡萄酸铜、D-(−)-阿拉伯糖、4-羟基-3-甲氧基肉桂酸乙酯、N-阿魏酰胍丁胺、葡糖酸、2′, 4′, 5, 7-四羟基异黄酮、N-乙酰-DL-色氨酸、异泽兰黄素-3-葡萄糖苷、十六聚甲烷基鞘氨醇等15种产物表现为上调,而槲皮素-3-O-(6″-O-乙酰基)-β-D-吡喃半乳糖苷、橙皮甙、芦丁、聚半乳糖醛酸、2, 5-二羟基苯甲酸、槲皮素O-乙酰己烯苷、3-羟基-5-甲基苯酚-1-氧化基-β-D-葡萄糖、槲皮素等8种产物表现为下调。并且其中异泽兰黄素-3-葡萄糖苷在非抗虫组中并未检测到,但其在抗虫组的含量较高。
表 1 差异代谢物筛选
Table 1. Differential metabolite screening
化合物 CAS编号 非抗虫组
代谢物丰度抗虫组
代谢物丰度FC P VIP 调节变化 阿魏酰胍丁胺 N/A 360 000 1 923 333 5.343 0.001 1.708 上调 抗坏血酸 50-81-7 4 197 336 37 266 667 8.879 0.004 1.647 上调 槲皮素-3-O-(6″-O-乙酰基)-β-D-吡喃半乳糖苷 N/A 59 033 333 16 443 333 0.279 0.005 1.643 下调 没食子酸 149-91-7 74 367 501 333 6.741 0.002 1.697 上调 橙皮甙 520-26-3 20 766 667 1 596 673 0.077 0.001 1.695 下调 绿茶儿茶酚 970-74-1 1 613 333 17 633 333 10.930 0.007 1.708 上调 没食子儿茶素 4233-96-9 10 050 000 32 900 000 3.274 0 1.707 上调 杨梅苷 17912-87-7 2 901 667 39 633 333 13.659 0 1.708 上调 芦丁 153-18-4 48 933 333 6 833 339 0.140 0.010 1.618 下调 葡萄酸铜 815-82-7 122 700 521 000 4.246 0.003 1.661 上调 聚半乳糖醛酸 25990-10-7 58 167 30 233 0.520 0.009 1.608 下调 D-(−)-阿拉伯糖 10323-20-3 674 333 1 220 000 1.809 0.010 1.603 上调 4-羟基-3-甲氧基肉桂酸乙酯 N/A 70 100 221 667 3.162 0.008 1.594 上调 N-阿魏酰胍丁胺 N/A 265 000 1 373 333 5.182 0 1.721 上调 2, 5−二羟基苯甲酸 490-79-9 34 966 667 12 733 333 0.364 0 1.692 下调 槲皮素O-乙酰己烯苷 N/A 53 433 333 10 726 667 0.201 0.005 1.627 下调 葡糖酸 133-42-6 526 333 2 393 333 4.547 0.001 1.689 上调 2′, 4′ , 5, 7-四羟基异黄酮 1156-78-1 2 000 003 12 063 333 6.032 0.009 1.594 上调 3-羟基-5-甲基苯酚-1-氧化基-β-D-葡萄糖 N/A 2 373 333 158 433 0.067 0.002 1.682 下调 槲皮素 20364-84-5 52 666 667 6 833 339 0.130 0.007 1.626 下调 N-乙酰-DL-色氨酸 87-32-1 24 970 264 333 10.586 0.007 1.617 上调 异泽兰黄素 3-葡萄糖苷 N/A 9 31 866 667 3 540 740.740 0.007 1.694 上调 十六聚甲烷基鞘氨醇 N/A 4 260 000 6 223 333 1.461 0.009 1.675 上调 说明:CAS编号是美国化学文摘服务社(chemical abstracts service, CAS)为化学物质编制的登记号;N/A表示该物质并未查询到
CAS编号 -
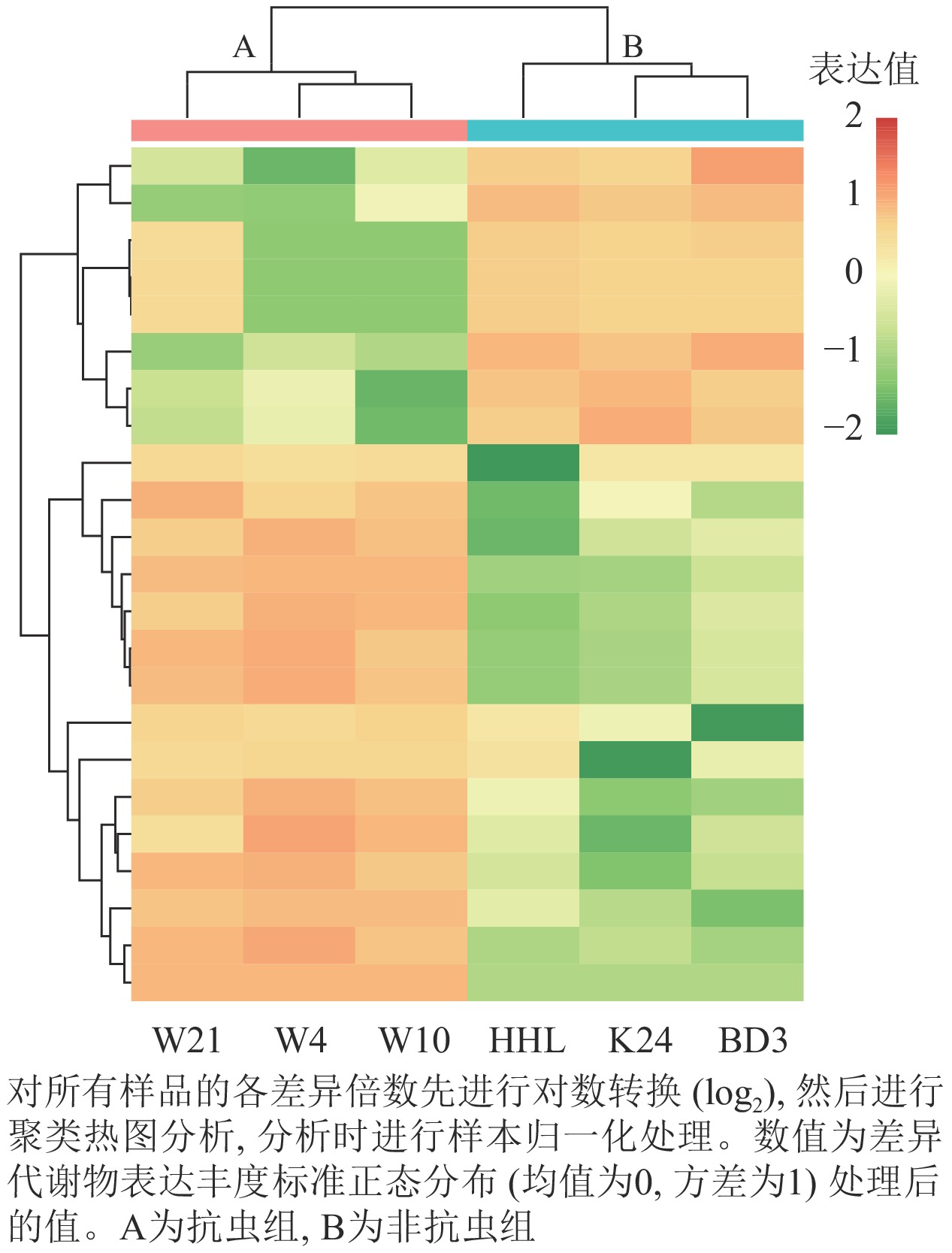
筛选具有显著差异(P<0.05)的代谢物进行聚类热图分析,进一步挖掘特征,并对筛选后的差异代谢物归一化处理。由图4显示:抗虫组样本丰度明显区别于非抗虫组样本,抗虫组样本组内也有差异但不明显,非抗虫组中黄花柳的样本成分分布丰度与垂柳和旱柳存在差异。可以看出:能明显划分成4个区域冷暖色差分异,在非抗虫组丰度高而在抗虫组中丰度低的代谢物,为8种下调代谢产物;在抗虫组中丰度高而在非抗虫组中丰度低的代谢物为15种上调代谢物。
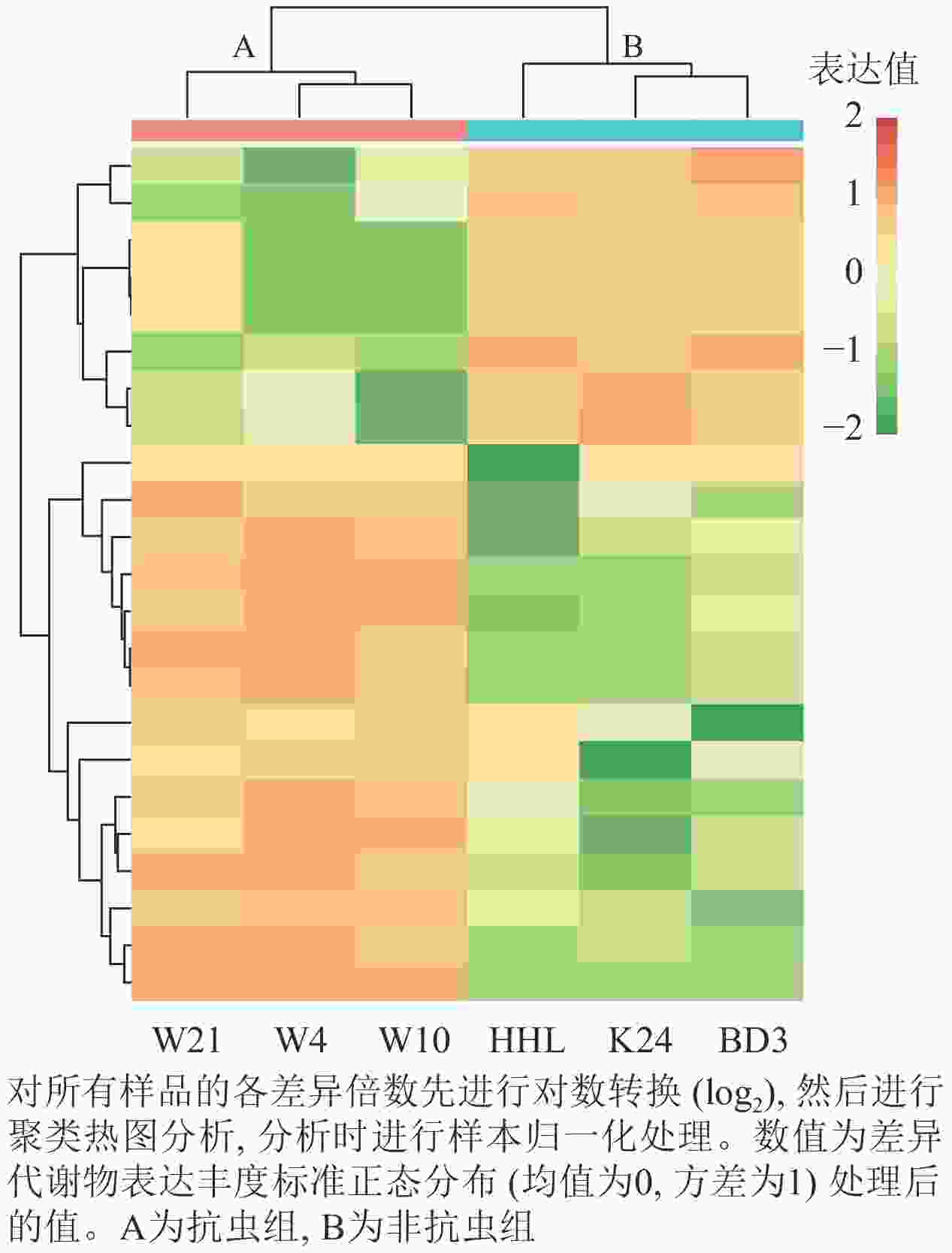
图 4 差异代谢聚类热图
Figure 4. Differential metabolism clustering heat map
-
本研究对4种不同柳树无性系进行了室内饲虫,结果显示三蕊柳对柳蓝叶甲表现出了高抗性;通过应用代谢组学方法对非抗虫组与抗虫组的次生代谢产物进行测定,筛选并最终鉴定出23种主要的差异代谢产物。上调次生代谢物包括阿魏酰胍丁胺、抗坏血酸、没食子酸、绿茶儿茶酚、没食子儿茶素、杨梅苷、葡萄酸铜、D-(−)-阿拉伯糖、4-羟基-3-甲氧基肉桂酸乙酯、N-阿魏酰胍丁胺、葡糖酸、2′, 4′, 5, 7-四羟基异黄酮、N-乙酰-DL-色氨酸、异泽兰黄素-3-葡萄糖苷、十六聚甲烷基鞘氨醇等15种。下调次生代谢物包括槲皮素-3-O-(6″-O-乙酰基)-β-D-吡喃半乳糖苷、橙皮甙、芦丁、聚半乳糖醛酸、2, 5-二羟基苯甲酸、槲皮素O-乙酰己烯苷、3-羟基-5-甲基苯酚-1-氧化基-β-D-葡萄糖、槲皮素等8种。这些差异特定代谢物影响的机制尚待进一步研究。
凌嘉昊[20]通过调查野外虫害等级和室内取食率来对比簸箕柳S. suchowensis和三蕊柳的抗虫性,发现三蕊柳表现出了高抗性,与本研究结果一致;该研究还发现三蕊柳的挥发物会使柳蓝叶甲产生趋避反应,并且鉴定分析显示22种挥发物组分在释放量方面有显著差异。而本研究发现:柳蓝叶甲的幼虫对三蕊柳存在取食行为,在接虫后的第4天全部死亡,表明三蕊柳除挥发物会对柳蓝叶甲产生趋避反应外,其次生代谢产物可能会对柳蓝叶甲幼虫产生毒害作用,或缺乏幼虫生长发育所必需的营养物质而导致其死亡。黄酮类物质是植物中一类重要的次生代谢物质,可以影响昆虫的行为和代谢,使之产生忌避、拒食等行为,破坏昆虫的正常代谢,严重时导致昆虫中毒甚至死亡,是对昆虫有毒的一类次生代谢产物[21]。生物碱是一大类含氮的碱性物质,对昆虫的防御作用主要表现在阻碍昆虫的取食以及引起昆虫中毒2个方面[22]。有研究表明:树木中的酚酸类物质含量越高,对天牛Cerambycidae的抗性就越强,但不同酚酸在不同无性系间的变化规律随其种类的不同而不同[23]。本研究可为培育抗虫性强、性状优良的柳树品种提供理论基础,三蕊柳的具体抗虫机制还有待进一步研究。
Insect resistance and metabolome of four willow species
-
摘要:
目的 研究不同柳树Salix spp. 对柳蓝叶甲Plagiodera versicolora的抗虫性,为选育抗虫性强的柳树品种提供参考。 方法 对旱柳S. matsudana、黄花柳S. caprea、垂柳S. babylonica以及三蕊柳S. triandra的3个无性系进行室内饲虫试验,并对抗虫组与非抗虫组进行差异次生代谢产物的测定与筛选。 结果 各无性系室内饲虫成活率旱柳为98%、垂柳为93%、黄花柳为90%,三蕊柳的3个无性系均为0。非抗虫组与抗虫组有23种主要的差异代谢产物,其中阿魏酰胍丁胺、抗坏血酸、没食子酸、绿茶儿茶酚、没食子儿茶素、杨梅苷、葡萄酸铜、D-(−)-阿拉伯糖、4-羟基-3-甲氧基肉桂酸乙酯、N-阿魏酰胍丁胺、葡糖酸、2′, 4′, 5, 7-四羟基异黄酮、N-乙酰-DL-色氨酸、异泽兰黄素 3-葡萄糖苷、十六聚甲烷基鞘氨醇等15种产物表现为上调,而槲皮素-3-O-(6″-O-乙酰基)-β-D-吡喃半乳糖苷、橙皮甙、芦丁、聚半乳糖醛酸、2, 5-二羟基苯甲酸、槲皮素O-乙酰己烯苷、3-羟基-5-甲基苯酚-1-氧化基-β-D-葡萄糖、槲皮素等8种产物表现为下调。 结论 三蕊柳对柳蓝叶甲的抗虫性最佳;非抗虫组与抗虫组之间的主要次生代谢产物存在显著差异,表明三蕊柳的次生代谢产物可能会对柳蓝叶甲幼虫产生毒害作用,或缺乏幼虫生长发育所必需的营养物质而导致柳蓝叶甲幼虫死亡。图4表1参23 Abstract:Objective This purpose is to explore the insect resistance of different willows (Salix spp.) to Plagiodera versicolora, and to provide reference for the breeding of willow cultivars with strong insect resistance. Method S. caprea, S. matsudana, S. babylonica and 3 clones of S. triandra were tested in laboratory for insect feeding, and the differential secondary metabolites between insect resistant and non insect resistant groups were determined and screened. Result The survival rate of indoor feeding was 98% for S. matsudana, 93% for S. babylonica, 90% for S. caprea and 0 for S. triandra. There were 23 main different metabolites between the non insect resistant group and the insect resistant group, among which 15 were up-regulated, including fer-agmatine, L-ascorbic acid, gallic acid, (−)-epigallocatechin, (+)-gallocatechin, myricitrin, L-(+)-tartaric acid, D-(−)-arabinose, hydroxy-methoxycinnamate, N-feruloyl agmatine, gluconic acid, 2 ′-Hydroxygenistein, N-Acetyl-DL-tryptophan, eupatilin 3-glucoside, and hexadecylsphingosine, and 8 were down-regulated, including quercetin 3-O-(6″-O-malonyl)-galactoside, hesperidin, rutin, D-galactopyranuronate, 2,5-dihydroxy benzoic acid O-hexside, quercetin O-acetylhexoside, 3-hydroxy-5-methylphenol-1-Oxy-β-D-glucose, and bioquercetin. Conclusion S. triandra has the best insect resistance to P. versicolora. There are significant differences in the main secondary metabolites between the non insect resistant group and the insect resistant group, which indicates that the secondary metabolites of S. triandra may be toxic to the larvae of P. versicolora, or lack of nutrients necessary for larvae growth and development, resulting in the death of the larvae. [Ch, 4 fig. 1 tab. 23 ref.] -
Key words:
- willow (Salix spp.) /
- metabolome /
- secondary metabolites /
- Plagiodera versicolora
-
图 2 非抗虫组与抗虫组OPLS-DA得分图
Figure 2 OPLS-DA score chart of non-insect resistant group and insect resistant group
表 1 差异代谢物筛选
Table 1. Differential metabolite screening
化合物 CAS编号 非抗虫组
代谢物丰度抗虫组
代谢物丰度FC P VIP 调节变化 阿魏酰胍丁胺 N/A 360 000 1 923 333 5.343 0.001 1.708 上调 抗坏血酸 50-81-7 4 197 336 37 266 667 8.879 0.004 1.647 上调 槲皮素-3-O-(6″-O-乙酰基)-β-D-吡喃半乳糖苷 N/A 59 033 333 16 443 333 0.279 0.005 1.643 下调 没食子酸 149-91-7 74 367 501 333 6.741 0.002 1.697 上调 橙皮甙 520-26-3 20 766 667 1 596 673 0.077 0.001 1.695 下调 绿茶儿茶酚 970-74-1 1 613 333 17 633 333 10.930 0.007 1.708 上调 没食子儿茶素 4233-96-9 10 050 000 32 900 000 3.274 0 1.707 上调 杨梅苷 17912-87-7 2 901 667 39 633 333 13.659 0 1.708 上调 芦丁 153-18-4 48 933 333 6 833 339 0.140 0.010 1.618 下调 葡萄酸铜 815-82-7 122 700 521 000 4.246 0.003 1.661 上调 聚半乳糖醛酸 25990-10-7 58 167 30 233 0.520 0.009 1.608 下调 D-(−)-阿拉伯糖 10323-20-3 674 333 1 220 000 1.809 0.010 1.603 上调 4-羟基-3-甲氧基肉桂酸乙酯 N/A 70 100 221 667 3.162 0.008 1.594 上调 N-阿魏酰胍丁胺 N/A 265 000 1 373 333 5.182 0 1.721 上调 2, 5−二羟基苯甲酸 490-79-9 34 966 667 12 733 333 0.364 0 1.692 下调 槲皮素O-乙酰己烯苷 N/A 53 433 333 10 726 667 0.201 0.005 1.627 下调 葡糖酸 133-42-6 526 333 2 393 333 4.547 0.001 1.689 上调 2′, 4′ , 5, 7-四羟基异黄酮 1156-78-1 2 000 003 12 063 333 6.032 0.009 1.594 上调 3-羟基-5-甲基苯酚-1-氧化基-β-D-葡萄糖 N/A 2 373 333 158 433 0.067 0.002 1.682 下调 槲皮素 20364-84-5 52 666 667 6 833 339 0.130 0.007 1.626 下调 N-乙酰-DL-色氨酸 87-32-1 24 970 264 333 10.586 0.007 1.617 上调 异泽兰黄素 3-葡萄糖苷 N/A 9 31 866 667 3 540 740.740 0.007 1.694 上调 十六聚甲烷基鞘氨醇 N/A 4 260 000 6 223 333 1.461 0.009 1.675 上调 说明:CAS编号是美国化学文摘服务社(chemical abstracts service, CAS)为化学物质编制的登记号;N/A表示该物质并未查询到
CAS编号 下载: 导出CSV
下载: 导出CSV
-
[1] 施士争, 王红玲, 周洁. 观赏柳树资源多样性及应用展望[J]. 林业工程学报, 2015, 29(2): 1 − 6. SHI Shizheng, WANG Hongling, ZHOU Jie. Ornamental willow resources diversity and application prospect [J]. J For Eng, 2015, 29(2): 1 − 6. [2] 史瑞. 杨树对青杨脊虎天牛抗虫化学机制的研究[D]. 哈尔滨: 东北林业大学, 2008. SHI Rui. Study on Chemical Resistance Mechanism of Poplar Species to Xylotrechus rusticus (L.) [D]. Harbin: Northeast Forestry University, 2008. [3] 许国旺, 杨军. 代谢组学及其研究进展[J]. 色谱, 2003, 21(4): 316 − 320. XU Guowang, YANG Jun. Metabolomics and its research progress [J]. Chin J Chromatogr, 2003, 21(4): 316 − 320. [4] 李薇, 李淑娴. 林木抗虫性研究进展[J]. 西南林业大学学报(自然科学), 2018, 38(5): 180 − 188. LI Wei, LI Shuxian. Advances in insect resistance of forest trees [J]. J Southwest For Univ Na Sci, 2018, 38(5): 180 − 188. [5] 杨军, 宋硕林, CASTRO-PEREZ J, 等. 代谢组学及其应用[J]. 生物工程学报, 2005, 21(1): 1 − 5. YANG Jun, SONG Shuolin, CASTRO-PEREZ J, et al. Metabonomics and its applications [J]. Chin J Biotechnol, 2005, 21(1): 1 − 5. [6] 李会平, 黄大庄, 杨敏生, 等. 林木抗虫机制研究进展[J]. 河北林果研究, 2001, 16(1): 91 − 96. LI Huiping, HUANG Dazhuang, YANG Minsheng, et al. Advance in the study on mechanism of tree resistance to insect [J]. For Ecol Sci, 2001, 16(1): 91 − 96. [7] 穆丹, 桂祥, 汪知重, 等. 柳蓝叶甲对不同植物气味的选择效应[J]. 安徽农学通报, 2016, 22(23): 33 − 35. MU Dan, GUI Xiang, WANG Zhizhong, et al. Selective effect of willow beetle on different plant odors [J]. Anhui Agric Sci Bull, 2016, 22(23): 33 − 35. [8] MATSUDA K, MATSUO H. A flavonoid, luteolin-7-glucoside, as well as salicin and populin, stimulating the feeding of leaf beetles attacking salicaceous plants [J]. Appl Entomol Zool, 1985, 20(3): 305 − 313. [9] IKONEN A, TAHVANAINEN J, ROININEN H. Phenolic secondary compounds as determinants of the host plant preferences of the leaf beetle Agelastica alni [J]. Chemoecology, 2002, 12(3): 125 − 131. [10] 邓彦彦. 3种植物的次生代谢产物及生物功能研究[D]. 沈阳: 沈阳农业大学, 2019. DENG Yanyan. Secondary Metabolites of Three Plants and Their Biological Functions[D]. Shenyang: Shenyang Agricultural University, 2019. [11] 刘青, 李珮, 曾其国, 等. 川栝楼瓤中次生代谢物组成及相对含量分析研究[J]. 中国现代中药, 2020, 22(2): 224 − 230, 236. LIU Qing, LI Pei, ZENG Qiguo, et al. Study on composition and relative content of secondary metabolites in flesh of Sichuan Trichosanthes kirilowii [J]. Modern Chin Med, 2020, 22(2): 224 − 230, 236. [12] 靳鹏博. 遮阴对丹参生长和次生代谢物含量的影响[D]. 杨凌: 西北农林科技大学, 2017. JIN Pengbo. Effect of Shading on Growth and Content of Secondary Metabolites in Saliva miltiorrhiza Bunge[D]. Yangling: Northwest A&F University, 2017. [13] 胥苡. 小麦抗蚜性与次生代谢物的相关性研究[J]. 安徽农业科学, 2015, 43(22): 123 − 124, 127. XU Yi. The correlation between resistance to wheat aphid and secondary metabolites in common wheat [J]. J Anhui Agric Sci, 2015, 43(22): 123 − 124, 127. [14] 李泽坤, 谢倩, 王威, 等. 果树代谢组学研究进展[J]. 西北植物学报, 2014, 34(9): 1909 − 1915. LI Zekun, XIE Qian, WANG Wei, et al. Advance in research of fruit tree metabolomics [J]. Acta Bot Boreali-Occident Sin, 2014, 34(9): 1909 − 1915. [15] 王景顺, 吴秋芳, 路志芳. 植物次生代谢物与林木抗虫性研究进展[J]. 江苏农业科学, 2015, 43(8): 4 − 7. WANG Jingshun, WU Qiufang, LU Zhifang. Research progress on plant secondary metabolites and forest tree pest resistance [J]. Jiangsu Agric Sci, 2015, 43(8): 4 − 7. [16] 赵学丽, 周志军, 郭成博, 等. 柳树单宁含量与光肩星天牛危害的关系[J]. 林业勘查设计, 2011(1): 71 − 73. ZHAO Xueli, ZHOU Zhijun, GUO Chengbo, et al. Relationship between tannin content of willows poplar and damage of Anoplophora glabripennis [J]. For Invest Design, 2011(1): 71 − 73. [17] CONSONNI R, CAGLIANI L R, STOCCHERO M, et al. Triple concentrated tomato paste: discrimination between Italian and Chinese products [J]. J Agric Food Chem, 2009, 57(11): 4506 − 4513. [18] XIA Jianguo, WISHART D S. Web-based inference of biological patterns, functions and pathways from metabolomic data using Metaboanalyst [J]. Nat Protoc, 2011, 6(6): 743 − 760. [19] CHEN Wei, GONG Liang, GUO Zilong, et al. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: application in the study of rice metabolomics [J]. Molec Plant, 2013, 6(6): 1769 − 1780. [20] 凌嘉昊. 簸箕柳和三蕊柳叶片挥发性物质对柳蓝叶甲取食选择性影响及有效化学成分识别[D]. 南京: 南京林业大学, 2020. LING Jiahao. Effect of Volatile Metabolites Emitting from Leaves of Salix suchowensis and Salix triandra on Host Selection for Plagiodera versicolora and Identification of the Functional Chemicals[D]. Nanjing: Nanjing Forestry University, 2020. [21] 张华锋, 陈顺立, 朱建华, 等. 松墨天牛为害对马尾松针叶化学成分的影响[J]. 福建林学院学报, 2004, 24(1): 28 − 31. ZHANG Huafeng, CHEN Shunli, ZHU Jianhua, et al. Effect of Monochamus alternatus on the composition of chemical materials in needles of Pinus massoniana [J]. J For Environ, 2004, 24(1): 28 − 31. [22] 钦俊德. 昆虫与植物的关系[M]. 北京: 科学出版社, 1987. QIN Junde. Relationship of Insects and Plants[M]. Beijing: Science Press, 1987. [23] 李会平, 王志刚, 杨敏生, 等. 杨树单宁与酚类物质种类及含量与光肩星天牛危害之间关系的研究[J]. 河北农业大学学报, 2003, 26(1): 36 − 39. LI Huiping, WANG Zhigang, YANG Minsheng, et al. The relation between tannin and phenol constituents and resistance to Anoplophora glabripennis of various poplar tree species [J]. J Hebei Agric Univ, 2003, 26(1): 36 − 39. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210166
 点击查看大图
点击查看大图

图(4) / 表(1)
计量
- 文章访问数: 2533
- HTML全文浏览量: 596
- PDF下载量: 35
- 被引次数: 0
