




-
稻田土壤是保障国家稻米供给的重要支撑[1]。稻田土壤微生物及相关胞外酶是有机质分解和养分循环的主要驱动力,可灵敏指示土壤质量和健康状况,并对促进水稻Oryza sativa生长具有重要作用[2]。施肥是提高稻田土壤质量、保障水稻产量的主要方式,且与作物养分利用率、农田面源污染和微生物介导的温室气体排放存在密切联系[3-4]。因此,研究施用不同类型肥料对土壤养分、微生物数量、酶活性的影响及其驱动的生物化学过程,对了解土壤质量变化及保障国家粮食安全和改善农业环境具有重要的意义。大量研究证明:尽管化肥能够有效提高水稻产量,但长期过量施用化肥也会导致土壤酸化、作物养分利用率低,造成较为严重的环境污染和肥料浪费[5]。周晓阳等[6]研究发现:化肥的过量施用是导致水旱轮作下稻田土壤酸化和面源污染的重要原因。ZENG等[7]研究发现:长期过量施用氮肥不仅引起土壤结构和质量下降,而且进一步恶化了土壤微环境,降低了微生物多样性,抑制了微生物活性和功能,进而阻碍了微生物介导的养分循环过程。因此,有必要寻求有效合理的施肥方法,减少面源污染,提高土壤肥力和微生物活性与功能,促进稻田土壤健康发展。
增施有机肥与有机肥替代部分化肥是减少化肥用量、改善土壤质量、维持养分平衡和降低环境污染的重要途径之一[8]。温延臣等[9]研究发现:有机肥替代部分化肥能协调有机无机养分平衡供应,保证作物产量。在氮磷钾养分投入量相当的情况下,有机肥替代化肥能够提升土壤微生物量碳氮、水溶性碳氮的含量及其比例,提高水稻产量[10]。李小萌等[11]研究发现:施用有机肥可以改善土壤结构,增加土壤孔隙度,促进土壤微生物的生长和繁殖。炭基肥是利用物理或化学方法将生物质炭与氮、磷、钾等养分通过一定比例混合而成的新型肥料,融合了生物质炭与肥料各自的优点,因具备吸附能力强、养分释放缓慢、肥效持久等特点而受到广泛关注[12]。李春阳[13]研究发现:适量施用炭基肥能够提升稻田土壤速效养分含量,增加土壤微生物数量。炭基肥与有机肥替代部分化肥对水稻产量的促进作用可能与有机物料分解促进养分缓慢释放、提高微生物丰度和酶活性有密切关系。常栋等[14]研究表明:炭基肥能直接为微生物生长提供所需的养分,同时其丰富的孔隙结构可以为微生物的栖息提供良好的环境。然而,潘全良等[15]研究表明:炭基肥对土壤蔗糖酶与土壤过氧化氢酶活性有抑制作用。可见,炭基肥对作物生长和土壤微生物的影响与炭基肥种类、制备原料和组分比例、施用量和方式密切相关,其影响还存在不确定性。目前国内针对炭基肥土壤改良研究集中在作物产量、品质、土壤养分等方面[14-15],其如何影响土壤微生物数量和酶活性尚缺乏系统研究。
基于此,本研究以杭嘉湖平原青紫泥水稻土为供试土壤,利用田间试验,在等量氮、磷、钾养分施用条件下,比较了炭基肥、有机肥替代部分化肥和常规化肥对土壤养分、微生物群落丰度以及土壤酶活性的影响,探讨了土壤酶活性变化的主要影响因素,以期为炭基肥及有机肥替代化肥合理应用和培肥稻田土壤提供科学依据。
-
样地位于浙江省桐乡市屠甸镇(30°32′N,120°24′E)。该区域属于杭嘉湖平原,属亚热带季风气候,年平均气温为16.5 ℃。供试土壤为青紫泥水稻土,土壤基本化学性质如下:pH 6.22、总碳14.50 g·kg−1、全氮1.17 g·kg−1、全磷0.56 g·kg−1、有效磷4.90 mg·kg−1、速效钾119.00 mg·kg−1。
-
采用田间试验,设置4个处理:对照(不施肥,ck)、常规施肥(CF)、炭基肥(BF)与有机肥替代50%化肥(OF)处理。后3者处理合计氮磷钾养分投入一致,其中:氮肥270 kg·hm−2(以氮计),磷肥75 kg·hm−2(以五氧化二磷计),钾肥150 kg·hm−2(以氧化钾计)。以BF处理氮磷钾养分投入为标准,CF和OF投入后养分不足部分用尿素、钙镁磷肥、氯化钾补充,保证各处理之间氮、磷、钾施入量相等。采用随机区组设计,各处理小区面积8 m×4 m,重复3次。小区间用水泥埂隔开,保证各小区不串灌。CF和OF处理分2次施肥,50%作为基肥,50%作为追肥。各处理具体施肥方案如下:CF处理将配方肥按750.0 kg·hm−2、钙镁磷肥125.0 kg·hm−2作为基肥施入,将尿素290.3 kg·hm−2、氯化钾24.2 kg·hm−2作为追肥施入;OF处理将菜籽饼(含全氮50.00 g·kg−1, 全磷1.09 g·kg−1,0.83 g·kg−1)按2700.0 kg·hm−2、钙镁磷肥62.5 kg·hm−2作为基肥施入,将尿素290.3 kg·hm−2、氯化钾196.2 kg·hm−2作为追肥施入。BF处理中炭基肥(18-5-10,含15 %玉米秸秆炭)委托南京勤丰秸秆科技有限公司制备,作为基肥按1500.0 kg·hm−2一次性施入。供试水稻品种为浙优18,2019年6月14日施基肥,6月16日移栽,6月27日追肥,11月4日收获。肥料均匀撒施,各小区田间管理措施均与当地一致。
-
于水稻收获时在每个小区按“S”形采样法,随机采集10个表层(0~20 cm)土壤样品,形成混合样。土壤样品过2 mm筛剔除石砾植物根系及残体,混匀分为3份;一部分放置于4 ℃冰箱储存,用以测定土壤微生物生物量及土壤酶活性,一部分放置于−70 ℃冰箱储存,用于土壤总DNA提取,剩余样品于室温风干后用于土壤基础化学性质测定。
土壤化学性质参考鲁如坤[16]方法测定。土壤pH值采用pH计按土水质量比1.0∶2.5测定;土壤总碳和全氮采用元素分析仪(Vario EL Ⅲ,Elementar,德国)测定;有效磷采取0.5 mol·L碳酸氢钠浸提,钼锑抗比色法测定;速效钾采取醋酸铵浸提,火焰光度计测定。铵态氮(NH4 +-N)和硝态氮(NO3 −-N)采用2 mol·L−1氯化钾浸提,流动分析仪测定。溶解性有机碳(DOC)、溶解性氮(DN)采用去离子水浸提,总有机碳分析仪(日本岛津公司)测定。溶解性有机氮(DON)为溶解性氮与铵态氮和硝态氮含量的差值。土壤微生物生物量碳(MBC)和氮(MBN)采用氯仿熏蒸提取法,总有机碳分析仪(日本岛津公司)测定。
-
称取0.35 g冷冻干燥后的土壤样品,采用土壤DNA提取试剂盒(DNeasy PowerSoil® Kit,QIAGEN,美国),按说明书进行土壤DNA提取。提取后的DNA用超微量紫外分光光度计(NanoDrop One,Thermo,美国)测定其浓度和纯度,保存于−20 ℃。本研究分别用引物对338F和518R[17]、NS1F和FungR[18]、344F和517R[19]对细菌16S rRNA基因、真菌18S rRNA基因和古菌16S rRNA基因丰度进行定量分析。反应体系如下:2×SYBR Premix Ex Taq 10 μL(TaKaRa,日本),10.0 μmol·L−1上游和下游引物各0.5 μL,模板DNA 1.0 μL,无菌双蒸水8.0 μL。质粒及标准曲线的制作参照吴涛等[20]进行。标准曲线及样品测定3次重复,设置空白,反应在荧光定量PCR仪(CFX96TM Real-Time System,Bio-Rad,美国)上进行。
-
利用微孔板荧光法[20]测定α-葡萄糖苷酶(AG)、β-葡萄糖苷酶(BG)、纤维二糖水解酶(CB)、木聚糖苷酶(XYL)、亮氨酸氨基肽酶(LAP)、N-乙酰基-β-D氨基葡萄糖苷酶(NAG)和酸性磷酸酶(PHOS)等7种参与土壤碳氮磷循环相关的酶活性。以4-羟甲基-7-香豆素(MUB)和7-氨基-4-甲基香豆素(MUC)荧光标记的底物作为酶活性反应底物,通过MUB和MUC荧光强度的变化反映酶活性。土壤荧光酶活性测定参考吴涛等[20]的方法,用多功能酶标仪(Synergy™ H1,Biotek,美国)在荧光激发光 365 nm和检测光波长 450 nm 下测定,酶活性单位为 nmol·g−1·h−1。由于不同类型土壤酶活性差异较大,难以综合反映土壤总体酶活性,本研究采用酶的几何平均值表示单个样品总体的土壤酶活性。
-
数据利用SPSS 26.0统计分析,采用单因素方差分析(one-way ANOVA)和邓肯法(Duncan)多重比较检验不同处理间的差异显著性(P<0.05)。相关性分析采用皮尔逊(Pearson)相关分析法进行双尾检验确定显著性。冗余分析(RDA)用于检验土壤环境因子对土壤酶活性变化的显著性,单因素置换多元方差分析(one-way PERMANOVA)(999次置换检验)用于分析施肥处理对土壤酶活性影响的显著性(P<0.05)。
-
由表1可见:与ck相比,在水稻收获期各施肥处理对土壤pH、总碳、全氮、有效磷、速效钾和硝态氮均无影响,但显著影响了土壤铵态氮质量分数(P<0.05);与CF处理相比,BF处理显著提高了土壤总碳质量分数(P<0.05),提高幅度为20.2%;3种施肥处理均提高了土壤铵态氮质量分数,其中CF、BF和OF处理提高幅度分别为84.1%、83.4%和150.7%。
表 1 不同施肥处理对土壤化学性质的影响
Table 1. Effects of different fertilization treatments on soil chemical properties
处理编号 pH 总碳/(g·kg−1) 全氮/(g·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) 硝态氮/(mg·kg−1) 铵态氮/(mg·kg−1) ck 6.24±0.27 a 13.97±1.16 ab 1.93±0.23 a 9.58±0.36 a 139.59±1.06 a 1.18±1.08 a 5.25±2.08 c CF 6.20±0.26 a 12.97±0.15 b 2.23±0.15 a 10.77±1.19 a 157.28±15.69 a 2.86±0.85 a 9.67±0.96 b BF 6.11±0.10 a 15.57±1.18 a 2.27±0.42 a 11.94±2.43 a 133.90±28.88 a 2.09±0.82 a 9.63±2.03 b OF 6.23±0.15 a 14.27±1.24 ab 2.30±0.20 a 11.28±0.43 a 128.97±28.15 a 2.37±1.16 a 13.17±1.60 a 说明:数据为平均值±标准差。同列不同小写字母表示各处理间差异显著(P<0.05) -
由表2所见:BF和OF处理对易氧化态碳(ROC)无影响,但显著提高了溶解性有机碳质量分数(P<0.05),且OF处理提高幅度高于BF处理;仅OF处理显著提高了溶解性有机氮(DON)和微生物生物量碳(MBC)质量分数(P<0.05),提高幅度分别达136.6%和164.0%。由表3可见:与ck相比,仅OF处理显著提高了细菌和真菌丰度及真菌/细菌比(P<0.05),提高幅度为34.9%,98.3%和50.0%;CF和BF处理对细菌和古菌丰度无影响,但CF处理显著降低了真菌丰度(P<0.05)。
表 2 不同施肥处理对土壤溶解性有机碳(DOC)、溶解性有机氮(DON)、微生物生物量碳(MBC)和微生物生物量氮(MBN)质量分数的影响
Table 2. Effects of different fertilization treatments on soil dissolved organic C and N, microbial biomass C and N contents
处理编号 质量分数/(mg·kg−1) MBC/MBN 溶解性有机碳 溶解性有机氮 易氧化态碳 微生物生物量碳 微生物生物量氮 ck 354.49±64.31 b 17.51±4.42 b 3.97±0.02 a 90.36±26.55 b 17.06±3.93 a 5.68±2.62 a CF 414.7±27.81 ab 17.51±2.88 b 3.75±0.26 a 123.63±42.63 b 21.27±5.48 a 6.25±2.84 a BF 484.19±50.53 a 15.02±3.34 b 4.36±0.5 a 117.64±40.05 b 23.47±6.62 a 5.43±2.92 a OF 520.11±65.57 a 41.09±11.84 a 4.14±0.38 a 238.29±102.01 a 26.79±8.02 a 9.09±3.79 a 说明:数据为平均值±标准差。同列不同字母表示各处理间差异显著(P<0.05) 表 3 不同施肥处理对土壤细菌、真菌、古菌基因丰度的影响
Table 3. Effects of different fertilization treatments on the gene abundances of bacteria, fungi and archaeal
处理编号 细菌丰度/(×109拷贝·g−1) 真菌丰度/(×108拷贝·g−1) 古菌丰度/(×107拷贝·g−1) 真菌/细菌/(×10−2) ck 4.62±0.74 b 0.85±0.36 b 1.25±0.26 a 1.78±0.47 b CF 3.61±0.72 b 0.27±0.03 c 1.26±0.26 a 0.75±0.11 ab BF 4.01±0.98 b 0.67±0.33 bc 1.35±0.43 a 1.89±1.43 ab OF 6.24±0.32 a 1.68±0.33 a 1.89±0.42 a 2.68±0.39 a 说明:数据为平均值±标准差。同列不同字母表示各处理间差异显著(P<0.05) -
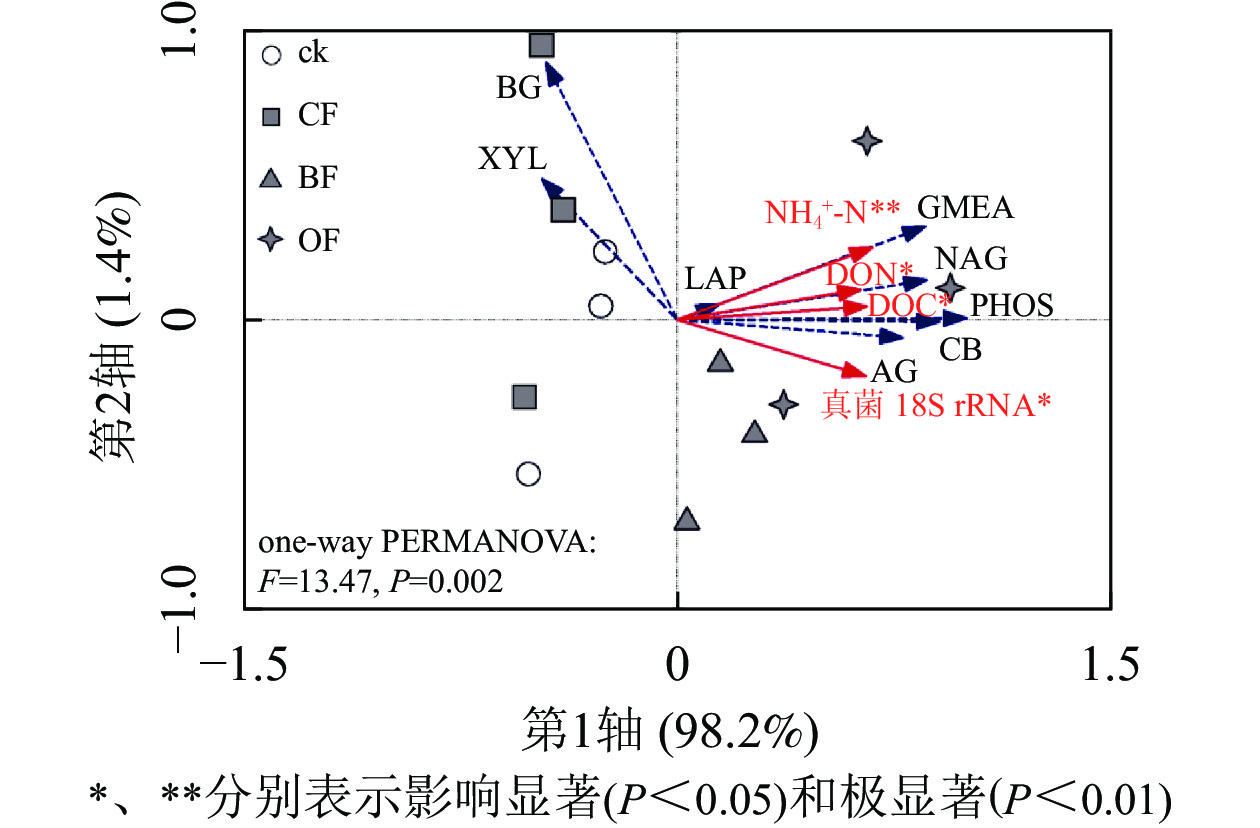
由图1可知:与ck相比,BF和OF处理均提高了土壤α-葡萄糖苷酶(AG)、纤维二糖苷酶(CB)、β-N-乙酰氨基葡萄糖酶(NAG)和酸性磷酸酶(PHOS)的活性,其中OF处理下4种酶活性分别较ck增加了136.7%、100.0%、177.2%和199.3%,BF处理下4种酶活性分别比ck增加111.6%、76.7%、109.4%和97.3%,而CF处理对其无显著影响。相比ck,3种施肥处理对β-葡萄糖苷酶(BG)、木聚糖苷酶(XYL)、亮氨酸氨基肽酶(LAP)活性均无影响。相比CF,BF处理具有更低的BG和XYL活性。OF处理下酶活性几何平均值(GMEA)显著高于ck和CF处理(P<0.05)。单因素置换多元方差分析表明(图2):施肥处理显著改变了土壤酶活性(P<0.05)。冗余分析表明:BF和OF处理下土壤酶活性与ck和CF在第1轴上具有明显分异。铵态氮(NH4 +-N)、溶解性有机碳(DOC)、溶解性有机氮(DON)及真菌18S rRNA丰度与纤维二糖苷酶(CB)、β-N-乙酰氨基葡萄糖酶(NAG)、酸性磷酸酶(PHOS)和酶活性几何平均值(GMEA)正相关,是显著影响土壤酶活性的环境因子;而其他性质对土壤酶活性变化影响较小。
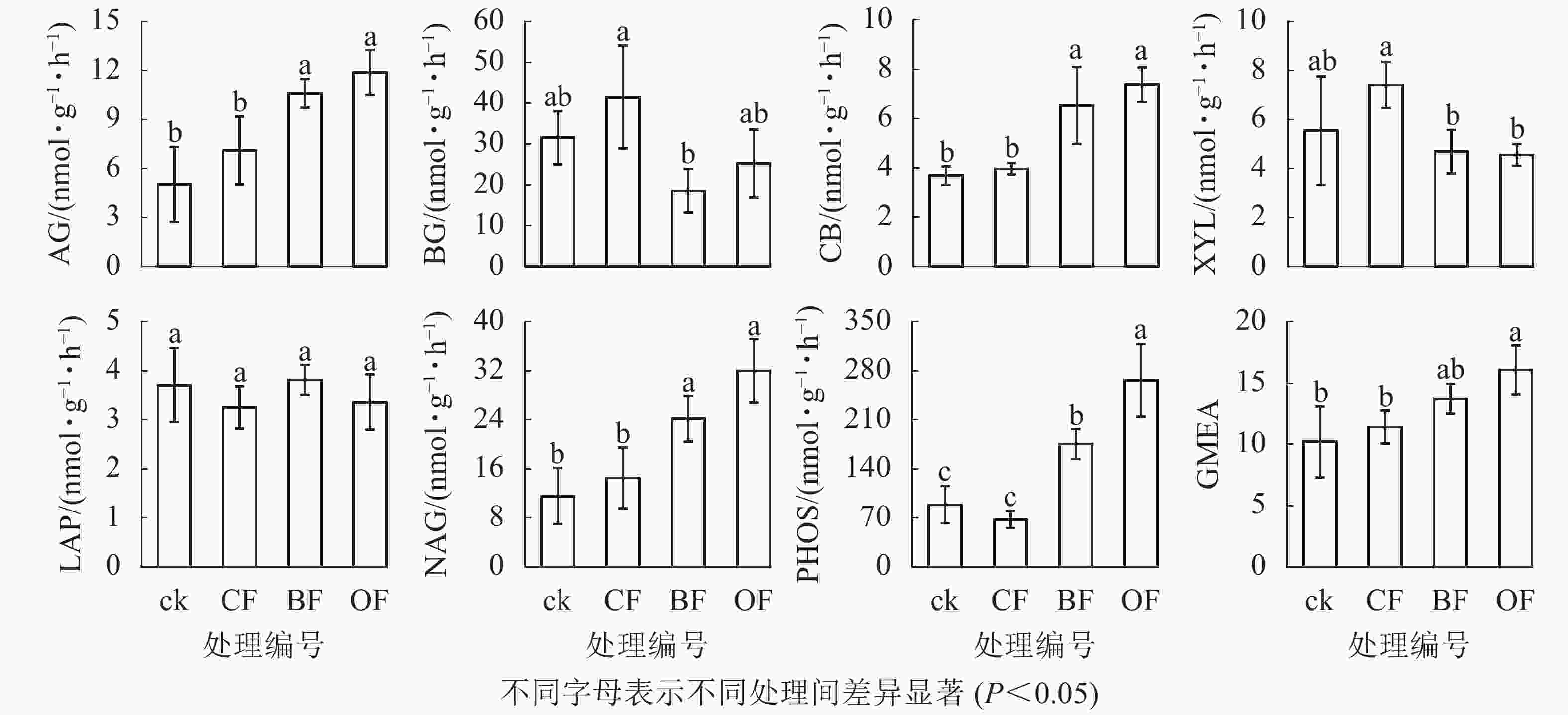
图 1 不同施肥处理对土壤酶活性的影响
Figure 1. Effects of different fertilization treatments on soil enzyme activities
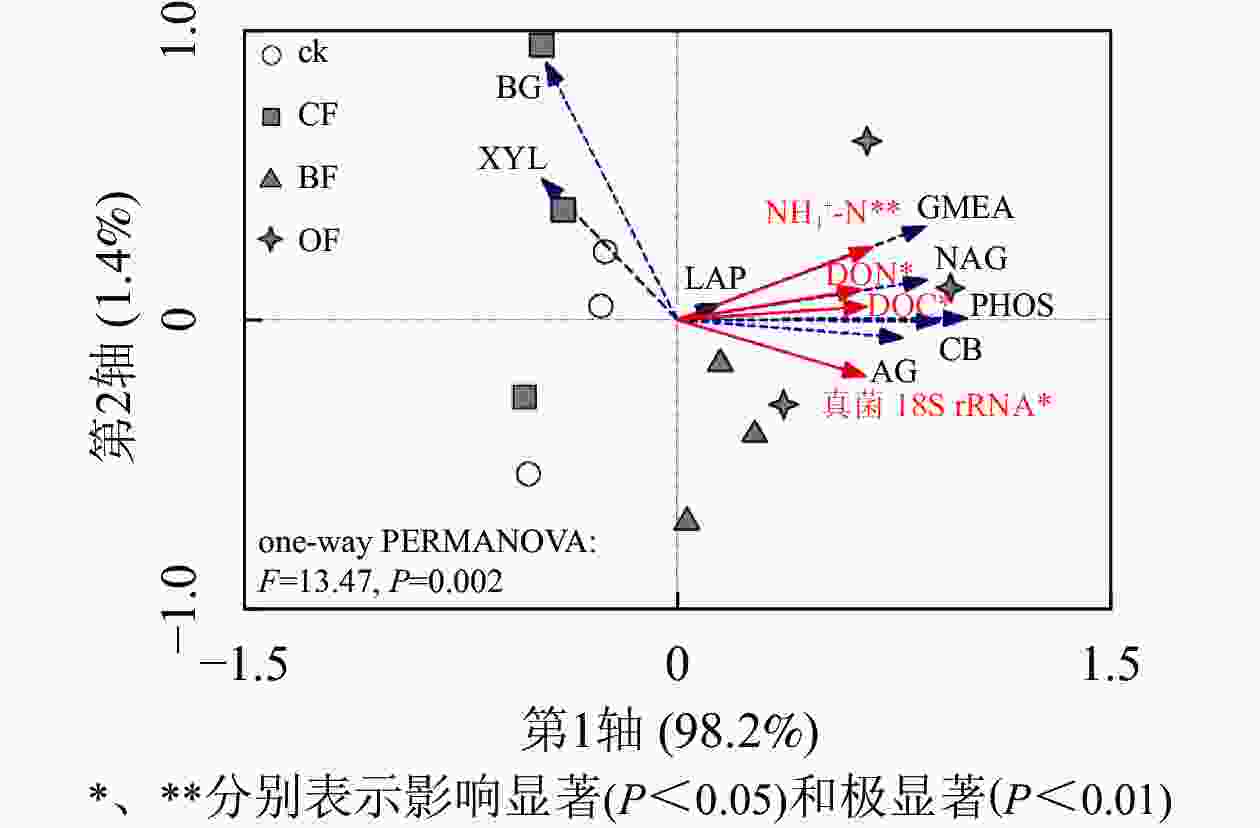
图 2 土壤化学性质和微生物量变化约束下土壤酶活性的冗余分析
Figure 2. Redundancy analysis of soil enzyme activities constrained by changes in soil chemical properties and microbial abundance
-
有机肥单独施用或配施一定量化肥可以显著提高稻田土壤pH,增加土壤有机质和矿质养分含量,从而提高稻田土壤肥力[21]。然而,炭基肥施用对土壤pH及土壤有机质和养分含量的影响结果并非一致[13-14]。李春阳[13]研究发现:适量施用炭基肥能够提高稻田土壤pH以及速效养分含量。杨天昱[22]研究表明:施用炭基肥使辽宁典型稻田土壤有机碳、碱解氮、速效钾含量分别提高了37.55%~41.48%、12.45%~13.55%、12.35%~12.90%。李昌娟等[23]研究表明:施用1.48~3.70 t·hm−2炭基肥5个月后,酸化茶园土壤pH以及铵态氮、硝态氮、速效磷等养分含量显著提高,且随炭基肥施用量的增加而增大。相反,常栋等[14]研究表明:不同用量炭基肥使土壤矿质氮含量降低了30%~49%。本研究中,常规化肥、炭基肥与有机肥替代部分化肥处理均未显著影响土壤pH、总碳、全氮、有效磷和速效钾质量分数,结果与多数报道不一致。这可能与不同肥料的施用量、施肥时间和受试土壤性质等有关。本研究采用菜籽饼替代50%化肥用量,施用量相对偏低,且因菜籽饼相对易分解,可能对水稻收获期土壤有机质、全氮等总养分质量分数影响不大。本研究中炭基肥用量以及其中生物质炭的比例相比李昌娟等[23]的研究结果较低,且只施用1次,可能也无法引起土壤有机碳、氮磷钾养分总量发生显著变化。溶解性有机质作为微生物养分重要来源,在土壤养分供应方面起着重要作用,相比土壤总有机碳和全氮可以更加灵敏反映土壤质量的内在变化[15]。本研究发现:施用有机肥和炭基肥显著提高了土壤水溶性有机碳和铵态氮质量分数,与石丽红等[24]的研究结果较为一致。这可能是因为有机肥和炭基肥具有速效和缓效养分,能够促进植物快速生长,导致更多的有机残余物经分解后以可溶性组分进入土壤。包建平等[25]研究表明:秸秆等有机物料施用可显著提高红壤活性碳组分含量,这是因为秸秆等有机物料分解释放的淀粉、半纤维素和多糖等物质是土壤溶解性有机质和矿质氮的主要来源。
-
本研究发现:仅有机肥替代部分化肥处理显著提高了稻田土壤微生物生物量碳、细菌和真菌的基因丰度,而炭基肥和化肥施用对其影响较小。已有研究表明:长期施用化肥可显著降低土壤微生物生物量,抑制土壤微生物和酶活性,而有机肥替代化肥可以改善土壤环境,增加土壤溶解性有机碳和氮源,从而促进微生物生长和繁殖,加快土壤养分周转速率[26-27]。本研究中菜籽饼营养丰富,碳氮比合适,在水稻收获期仍具有较高的水溶性有机碳和铵态氮质量分数,可能刺激了土壤微生物生长。尽管生物质炭在改善土壤结构、持水性、增加土壤养分含量、提高微生物丰度等方面已被广泛认可[28-29],但炭基肥对土壤结构和微生物丰度的影响相对微弱[30]。陈坤等[30]通过田间定位试验研究表明:猪厩肥以及生物炭均能显著提高土壤微生物磷脂脂肪酸含量和细菌丰度,但炭基肥对其影响较小。相反,陈懿等[31]研究发现:炭基肥增加了植烟土壤细菌、放线菌和真菌数量。炭基肥对土壤微生物的影响可能与炭基肥类型(如有机物料含量)、养分释放状况和水热协调能力等差异有关。本研究表明:炭基肥对细菌和真菌丰度并未产生显著促进作用,可能是单次炭基肥施用难以给微生物提供较多的底物来改善土壤环境。
本研究中,相比对照和化肥,炭基肥及有机肥替代化肥处理显著提高了土壤α-葡萄糖苷酶(AG)、纤维二糖水解酶(CB)、N-乙酰基-β-D氨基葡萄糖苷酶(NAG)和酸性磷酸酶(PHOS)活性,且有机肥提高幅度更大,而对其他酶活性无影响。这一结果表明炭基肥和有机肥部分替代化肥均有利于促进土壤碳、氮、磷转化相关酶活性,可潜在促进土壤有机质分解。有机肥施用对土壤碳、氮、磷转化相关水解酶活性的促进作用已有广泛报道,这与有机肥含有较高的易分解有机质,有利于促进微生物生长,进而促进微生物胞外酶的产生和酶促反应活性有关[26, 32]。包建平等[25]研究表明:秸秆等有机物料增加了土壤可溶性有机碳源和氮源,促进了β-葡萄糖苷酶(BG)和CB及微生物对葡萄糖、天冬氨酸和丁香酸的利用速率。炭基肥施用对土壤酶活性的影响报道相对较少,且结果存在不一致性。如潘全良等[15]发现:炭基肥对土壤蔗糖酶与土壤过氧化氢酶活性有抑制作用,而对土壤脲酶活性无明显影响;而陈懿等[31]研究显示:施用炭基肥可以提高土壤脲酶和过氧化氢酶活性,其中过氧化氢酶活性增幅达10.6%。以上这些迥异的报道可能与炭基肥中的生物质炭材料、本身养分释放特征以及土壤的养分有效性均有一定关系。本研究中施用炭基肥提高了AG、CB、NAG和PHOS活性,可能与炭基肥施用提高了土壤可溶性有机碳和铵态氮有关。冗余分析证实:铵态氮、溶解性有机碳和氮及真菌丰度与CB、NAG、PHOS正相关,是显著影响土壤酶活性的环境因子。也有相关研究表明:炭基肥含有的生物质炭可能通过吸附酶促反应的底物,促进酶促反应活性,抑制或阻碍酶促反应结合位点,进而抑制酶活性[33]。酶生态化学计量学理论认为:微生物氮、磷养分限制条件可进一步促进NAG和PHOS等胞外酶活性[29]。因此,在相同养分投入下,有机肥和炭基肥促进植物养分吸收也可能加剧了微生物的养分限制,从而促进了氮磷获取酶活性。与化肥的快速养分供给特点不同,有机肥和炭基肥一方面通过自身所含养分的缓慢分解与释放提高了土壤可溶性有机质,另一方面通过提高土壤碳、氮、磷转化酶活性促进养分周转,从而加速稻田土壤养分循环。然而,有机肥和炭基肥对土壤酶活性的影响方式可能存在差异:有机肥主要通过自身的大量可降解性碳氮组分提高微生物量,从而提高相关水解酶活性;炭基肥本身易分解组分含量低,并不能显著提高土壤微生物量,但可通过自身孔隙结构和养分缓释性能调控土壤酶活性。
-
在等量氮、磷、钾养分投入下,炭基肥与有机肥替代部分化肥较常规施肥处理对土壤总碳、全氮、有效磷和速效钾质量分数无显著影响,但均提高了土壤α-葡萄糖苷酶、纤维二糖苷酶、β-N-乙酰氨基葡萄糖苷酶和酸性磷酸酶活性,且后者提高幅度更大。有机肥替代部分化肥较常规施肥显著提高了土壤微生物生物量碳质量分数、细菌和真菌丰度,而炭基肥对其无影响。因此,有机肥替代部分化肥和炭基肥处理均可提高土壤碳、氮、磷转化酶活性,促进稻田养分周转。
Effects of biochar-based fertilizer and organic fertilizer substituting chemical fertilizer partially on soil microbial abundances and enzyme activities
-
摘要:
目的 探究炭基肥施用和有机肥替代部分化肥对土壤养分含量、微生物丰度和酶活性的影响,分析土壤酶活性变化的主要驱动因子,为提高稻田土壤质量和新型肥料应用提供科学依据。 方法 在杭嘉湖平原典型稻田进行田间小区试验,供试土壤为青紫泥稻田土。设置4个处理:不施肥对照(ck)、常规施肥(CF)、炭基肥(BF)及有机肥替代50%化肥(OF),3种施肥处理的氮、磷、钾投入量一致。处理开始于2019年6月,于2019年11月水稻Oryza sativa收获后采集0~20 cm土壤样品,测定土壤碳氮组分质量分数、细菌、真菌和古菌丰度及土壤碳氮磷循环相关酶活性。 结果 与CF处理相比,BF和OF处理对土壤pH、总碳、易氧化态碳、全氮、有效磷、速效钾、硝态氮质量分数无显著影响,而OF处理显著提高了土壤铵态氮和溶解性有机氮质量分数。与ck相比,OF处理显著提高了土壤微生物生物量碳质量分数(164%)、细菌16S rRNA (35%)和真菌18S rRNA基因拷贝数(98%)及真菌/细菌比(50%),而BF处理对上述指标无影响。3种施肥处理对β-葡萄糖苷酶、木聚糖苷酶、亮氨酸氨基肽酶活性均无影响,而BF和OF处理显著提高了土壤α-葡萄糖苷酶(111%和136%)、纤维二糖苷酶(77%和100%)、β-N-乙酰氨基葡萄糖苷酶(109%和177%)和酸性磷酸酶(97%和199%)活性,OF处理平均提高幅度高于BF处理。冗余分析表明:土壤铵态氮、溶解性有机碳和氮、真菌丰度是影响土壤酶活性变化的主要驱动因子。 结论 相比常规施肥,有机肥替代部分化肥和炭基肥施用提高了土壤碳氮磷转化相关酶活性,且有机肥替代部分化肥进一步提高了微生物丰度,对促进稻田土壤养分周转具有重要意义。图2表3参33 Abstract:Objective The objective is to provide a scientific basis for improving rice paddy soil quality and for the application of new fertilizer, by investigating the effects of biochar-based fertilizer and organic fertilizer substituting chemical fertilizer partially on soil nutrients contents, microbial abundances and enzyme activities, and investigated the driving factors for soil enzyme activities. Method A field experiment was conducted in a typical rice paddy located in Hangjiahu Plain, which soil is Qingzini paddy soil. Four treatments, namely no fertilizer control (ck), conventional fertilizer (CF), biochar-based fertilizer (BF) and organic fertilizer substitution of 50% chemical fertilizer (OF), were laid out with the three fertilization treatments had consistent input of N, P and K amount. The field trail was initiated on June 2019 and soil sampling were collectted on November, 2019. Topsoils (0−20 cm) were sampled to investigate changes in soil carbon and nitrogen fractions, bacterial, fungal and archaeal abundances and enzyme activities involved in C, N and P cycling. Result Compared with CF, BF and OF treatments had no effects on soil pH, total C and N, available P and K and nitrate contents, but OF significantly increased soil ammonia and dissolved organic C contents. Compared with ck, OF treatment increased the content of soil microbial biomass carbon (MBC) by 164%, bacterial 16S rRNA gene abundance by 35% and fungal 18S rRNA gene abundances by 98% and fungi/bacteria ratio by 50%, while BF and CF had no effects on them. The three fertilization treatments had no effects on the activities of β-glucosidase, β-xylosidase and Leucine aminopeptidase, whereas BF and OF treatments significantly increased the activities of α-glucosidase (AG) by 111%和136%, β-cellobiosidase (BG) by 77%和100%, β-N-acetylglucosaminidase (NAG) by 109%和177% and acid phosphatase (PHOS) by 97%和199%, respectively. Redundant analysis indicated that changes in soil enzyme activities were strongly dependent on the contents of soil ammonia, dissolved organic C and N, and fungal abundances. Conclusion The application of organic fertilizer and biochar-based fertilizer significantly increased soil enzyme activities involved in C, N and P cycling, with the OF treatment further increased soil microbial abundance, which was beneficial for soil nutrient cycling. [Ch, 2 fig. 3 tab. 33 ref.] -
Key words:
- organic fertilizer /
- biochar-based fertilizer /
- soil microbes /
- soil enzymes
-
图 1 不同施肥处理对土壤酶活性的影响
Figure 1 Effects of different fertilization treatments on soil enzyme activities
图 2 土壤化学性质和微生物量变化约束下土壤酶活性的冗余分析
Figure 2 Redundancy analysis of soil enzyme activities constrained by changes in soil chemical properties and microbial abundance
表 1 不同施肥处理对土壤化学性质的影响
Table 1. Effects of different fertilization treatments on soil chemical properties
处理编号 pH 总碳/(g·kg−1) 全氮/(g·kg−1) 有效磷/(mg·kg−1) 速效钾/(mg·kg−1) 硝态氮/(mg·kg−1) 铵态氮/(mg·kg−1) ck 6.24±0.27 a 13.97±1.16 ab 1.93±0.23 a 9.58±0.36 a 139.59±1.06 a 1.18±1.08 a 5.25±2.08 c CF 6.20±0.26 a 12.97±0.15 b 2.23±0.15 a 10.77±1.19 a 157.28±15.69 a 2.86±0.85 a 9.67±0.96 b BF 6.11±0.10 a 15.57±1.18 a 2.27±0.42 a 11.94±2.43 a 133.90±28.88 a 2.09±0.82 a 9.63±2.03 b OF 6.23±0.15 a 14.27±1.24 ab 2.30±0.20 a 11.28±0.43 a 128.97±28.15 a 2.37±1.16 a 13.17±1.60 a 说明:数据为平均值±标准差。同列不同小写字母表示各处理间差异显著(P<0.05)  下载: 导出CSV
下载: 导出CSV
表 2 不同施肥处理对土壤溶解性有机碳(DOC)、溶解性有机氮(DON)、微生物生物量碳(MBC)和微生物生物量氮(MBN)质量分数的影响
Table 2. Effects of different fertilization treatments on soil dissolved organic C and N, microbial biomass C and N contents
处理编号 质量分数/(mg·kg−1) MBC/MBN 溶解性有机碳 溶解性有机氮 易氧化态碳 微生物生物量碳 微生物生物量氮 ck 354.49±64.31 b 17.51±4.42 b 3.97±0.02 a 90.36±26.55 b 17.06±3.93 a 5.68±2.62 a CF 414.7±27.81 ab 17.51±2.88 b 3.75±0.26 a 123.63±42.63 b 21.27±5.48 a 6.25±2.84 a BF 484.19±50.53 a 15.02±3.34 b 4.36±0.5 a 117.64±40.05 b 23.47±6.62 a 5.43±2.92 a OF 520.11±65.57 a 41.09±11.84 a 4.14±0.38 a 238.29±102.01 a 26.79±8.02 a 9.09±3.79 a 说明:数据为平均值±标准差。同列不同字母表示各处理间差异显著(P<0.05)
下载: 导出CSV
表 3 不同施肥处理对土壤细菌、真菌、古菌基因丰度的影响
Table 3. Effects of different fertilization treatments on the gene abundances of bacteria, fungi and archaeal
处理编号 细菌丰度/(×109拷贝·g−1) 真菌丰度/(×108拷贝·g−1) 古菌丰度/(×107拷贝·g−1) 真菌/细菌/(×10−2) ck 4.62±0.74 b 0.85±0.36 b 1.25±0.26 a 1.78±0.47 b CF 3.61±0.72 b 0.27±0.03 c 1.26±0.26 a 0.75±0.11 ab BF 4.01±0.98 b 0.67±0.33 bc 1.35±0.43 a 1.89±1.43 ab OF 6.24±0.32 a 1.68±0.33 a 1.89±0.42 a 2.68±0.39 a 说明:数据为平均值±标准差。同列不同字母表示各处理间差异显著(P<0.05)
下载: 导出CSV
-
[1] 徐春春, 纪龙, 陈中督, 等. 2020年我国水稻产业形势分析及2021年展望[J]. 中国稻米, 2021, 27(2): 1 − 4. XU Chunchun, JI Long, CHEN Zhongdu, et al. Analysis of China’s rice industry in 2020 and the outlook for 2021 [J]. China Rice, 2021, 27(2): 1 − 4. [2] ZHANG Q C, SHAMSI I H, XU D T, et al. Chemical fertilizer and organic manure inputs in soil exhibit a vice versa pattern of microbial community structure [J]. Appl Soil Ecol, 2012, 57: 1 − 8. [3] LI Zhaolei, ZENG Zhaoqi, SONG Zhaopeng, et al. Vital roles of soil microbes in driving terrestrial nitrogen immobilization [J]. Glob Change Biol, 2021, 27: 1848 − 1858. [4] SCHMIDT J E, KENT A D, BRISSON V L, et al. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling[J/OL]. Microbiome, 2019, 7: 146 [2021-08-10]. doi: 10.1186/s40168-019-0756-9. [5] 徐洋, 杨帆, 张卫峰, 等. 2014—2016年我国种植业化肥施用状况及问题[J]. 植物营养与肥料学报, 2019, 25(1): 11 − 21. XU Yang, YANG Fan, ZHANG Weifeng, et al. Status and problems of chemical fertilizer application in crop plantations of China from 2014 to 2016 [J]. J Plant Nutr Fert, 2019, 25(1): 11 − 21. [6] 周晓阳, 徐明岗, 周世伟, 等. 长期施肥下我国南方典型农田土壤的酸化特征[J]. 植物营养与肥料学报, 2015, 21(6): 1615 − 1621. ZHOU Xiaoyang, XU Minggang, ZHOU Shiwei, et al. Soil acidification characteristics in southern China’ s croplands under long-term fertilization [J]. J Plant Nutr Fert, 2015, 21(6): 1615 − 1621. [7] ZENG Jun, LIU Xuejun, SONG Ling, et al. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition [J]. Soil Biol Biochem, 2016, 92: 41 − 49. [8] 张福锁. 科学认识化肥的作用[J]. 中国农技推广, 2017, 33(1): 16 − 19. ZHANG Fusuo. Scientific understanding of the role of chemical fertilizers [J]. China Agric Technol Ext, 2017, 33(1): 16 − 19. [9] 温延臣, 张曰东, 袁亮, 等. 商品有机肥替代化肥对作物产量和土壤肥力的影响[J]. 中国农业科学, 2018, 51(11): 2136 − 2142. WEN Yanchen, ZHANG Yuedong, YUAN Liang, et al. Crop yield and soil fertility response to commercial organic fertilizer substituting chemical fertilizer [J]. Sci Agric Sin, 2018, 51(11): 2136 − 2142. [10] 李文军, 彭保发, 杨奇勇. 长期施肥对洞庭湖双季稻区水稻土有机碳、氮积累及其活性的影响[J]. 中国农业科学, 2015, 48(3): 488 − 500. LI Wenjun, PENG Baofa, YANG Qiyong. Effects of long-term fertilization on organic carbon and nitrogen accumulation and activity in a paddy soil in double cropping rice area in Dongting Lake of China [J]. Sci Agric Sin, 2015, 48(3): 488 − 500. [11] 李小萌, 陈效民, 曲成闯, 等. 生物有机肥与减量配施化肥对连作黄瓜养分利用率及产量的影响[J]. 水土保持学报, 2020, 34(2): 309 − 317. LI Xiaomeng, CHEN Xiaomin, QU Chengchuang, et al. Effects of bio-organic fertilizer combined with reduced fertilizer on nutrient utilization and yield of continuous cropping cucumber [J]. J Soil Water Conserv, 2020, 34(2): 309 − 317. [12] 赵泽州, 王晓玲, 李鸿博, 等. 生物质炭基肥缓释性能及对土壤改良的研究进展[J]. 植物营养与肥料学报, 2021, 27(5): 886 − 897. ZHAO Zezhou, WANG Xiaoling, LI Hongbo, et al. Slow-release property and soil remediation mechanism of biochar-based fertilizers [J]. J Plant Nutr Fert, 2021, 27(5): 886 − 897. [13] 李春阳. 减氮条件下配施不同用量炭基肥对红壤稻田土壤性质的影响[D]. 南昌: 江西农业大学, 2020. LI Chunyang. Effects of Different Amount of Biochar based Fertilizer on Soil Properties of Red Soil Paddy Field under Nitrogen Reduction Condition[D]. Nanchang: Jiangxi Agricultural University, 2020. [14] 常栋, 马文辉, 张凯, 等. 生物炭基肥对植烟土壤微生物功能多样性的影响[J]. 中国烟草学报, 2018, 24(6): 58 − 66. CHANG Dong, MA Wenhui, ZHANG Kai, et al. Effect of biochar fertilizer on microbial functional diversity in tobacco growing soil [J]. Acta Tab Sin, 2018, 24(6): 58 − 66. [15] 潘全良, 宋涛, 陈坤, 等. 连续6年施用生物炭和炭基肥对棕壤生物活性的影响[J]. 华北农学报, 2016, 31(3): 225 − 232. PAN Quanliang, SONG Tao, CHEN Kun, et al. Influences of 6-year application of biochar and biochar-based compound fertilizer on soil bioactivity on brown soil [J]. Acta Agric Boreali-Sin, 2016, 31(3): 225 − 232. [16] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000. LU Rukun. Methods of Soil Agricultural Chemical Analysis[M]. Beijing: China Agricultural Science and Technology Press, 2000. [17] FIERER N, JACKSON J A, VILGALYS R, et al. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays [J]. Appl Environ Microbiol, 2005, 71(7): 4117 − 4120. [18] MAY L A, SMILEY B, SCHMIDT M G. Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage [J]. Can J Microbiol, 2001, 47(9): 829 − 841. [19] BANO N, RUFFIN S, RANSOM B, et al. Phylogenetic composition of arctic ocean archaeal assemblages and comparison with antarctic assemblages [J]. Appl Environ Microbiol, 2004, 70(2): 781 − 789. [20] 吴涛, 冯歌林, 曾珍, 等. 生物质炭对盆栽黑麦草生长的影响及机理[J]. 土壤学报, 2017, 54(2): 525 − 534. WU Tao, FRNG Gelin, ZENG Zhen, et al. Effect of biochar addition on ryegrass growth in a pot experiment and its mechanism [J]. Acta Pedol Sin, 2017, 54(2): 525 − 534. [21] 吕真真, 吴向东, 侯红乾, 等. 有机–无机肥配施比例对双季稻田土壤质量的影响[J]. 植物营养与肥料学报, 2017, 23(4): 904 − 913. LÜ Zhenzhen, WU Xiangdong, HOU Honggan, et al. Effect of different application ratios of chemical and organic fertilizers on soil quality in double cropping paddy fields [J]. J Plant Nutr Fert, 2017, 23(4): 904 − 913. [22] 杨天昱. 生物炭基肥对水稻产量及稻田土壤肥力特性的影响[D]. 沈阳: 沈阳农业大学, 2020. YANG Tianyu. Effects of Biochar-based Fertilizer on Rice Yield and Paddy Soil Fertility Characteristics [D]. Shenyang Agricultural University, 2020. [23] 李昌娟, 杨文浩, 周碧青, 等. 生物炭基肥对酸化茶园土壤养分及茶叶产质量的影响[J]. 土壤通报, 2021, 52(2): 387 − 397. LI Changjuan, YANG Wenhao, ZHOU Biqing, et al. Effects of biochar based fertilizer on soil nutrients, tea output and quality in an acidified tea field [J]. Chin J Soil Sci, 2021, 52(2): 387 − 397. [24] 石丽红, 李超, 唐海明, 等. 长期不同施肥措施对双季稻田土壤活性有机碳组分和水解酶活性的影响[J]. 应用生态学报, 2021, 32(3): 921 − 930. SHI Lihong, LI Chao, TANG Haiming, et al. Effects of long-term fertilizer management on soil labile organic carbon fractions and hydrolytic enzyme activity under a double-cropping rice system of southern China [J]. Chin J Appl Ecol, 2021, 32(3): 921 − 930. [25] 包建平, 袁根生, 董方圆, 等. 生物质炭与秸秆施用对红壤有机碳组分和微生物活性的影响[J]. 土壤学报, 2020, 57(3): 721 − 729. BAO Jianping, YUAN Gensheng, DONG Fangyuan, et al. Effects of biochar application and straw returning on organic carbon fractionations and microbial activities in a red soil [J]. Acta Pedol Sin, 2020, 57(3): 721 − 729. [26] 徐一兰, 唐海明, 肖小平, 等. 长期施肥对双季稻田土壤微生物学特性的影响[J]. 生态学报, 2016, 36(18): 5847 − 5855. XU Yilan, TANG Haiming, XIAO Xiaoping, et al. Effects of different long-term fertilization regimes on the soil microbiological properties of a paddy field [J]. Acta Ecol Sin, 2016, 36(18): 5847 − 5855. [27] SHANG Lirong, WAN Liqiang, ZHOU Xiaoxin, et al. Effects of organic fertilizer on soil nutrient status, enzyme activity, and bacterial community diversity in Leymus chinensis steppe in Inner Mongolia, China [J/OL]. PLoS One, 2020, 15: e0240559[2021-09-20]. doi: 10.1371/journal.pone.0240559.eCollection 2020. [28] LEHMANN J, RILLIG M C, THIES J, et al. Biochar effects on soil biota: a review [J]. Soil Biol Biochem, 2011, 43(9): 1812 − 1836. [29] GUO Kangying, ZHAO Yingzhi, LIU Yang, et al. Pyrolysis temperature of biochar affects ecoenzymatic stoichiometry and microbial nutrient-use efficiency in a bamboo forest soil [J/OL]. Geoderma, 2020, 363: 114162[2021-09-11]. doi: 10.1016/j.geoderma.2019.114162. [30] 陈坤, 徐晓楠, 彭靖, 等. 生物炭及炭基肥对土壤微生物群落结构的影响[J]. 中国农业科学, 2018, 51(10): 1920 − 1930. CHEN Kun, XU Xiaonan, PENG Jing, et al. Effects of biochar and biochar-based fertilizer on soil microbial community structure [J]. Sci Agric Sin, 2018, 51(10): 1920 − 1930. [31] 陈懿, 吴春, 李彩斌, 等. 炭基肥对植烟黄壤细菌、真菌群落结构和多样性的影响[J]. 微生物学报, 2020, 60(4): 653 − 666. CHEN Yi, WU Chun, LI Caibin, et al. Effect of biochar-based fertilizer on bacterial and fungal community composition, diversity in tobacco-planting yellow soil [J]. Acta Microbiol Sin, 2020, 60(4): 653 − 666. [32] MIAO Fuhong, LI Yuan, CUI Song, et al. Soil extracellular enzyme activities under long-term fertilization management in the croplands of China: a meta-analysis [J]. Nutr Cycling in Agroecosyst, 2019, 114: 125 − 138. [33] BAILEY V L, FANSLER S J, SMITH J L, et al. Reconciling apparent variability in effects of biochar amendment on soil enzyme activities by assay optimization [J]. Soil Biol Biochem, 2011, 43(2): 296 − 301. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210619
 点击查看大图
点击查看大图
计量
- 文章访问数: 1418
- HTML全文浏览量: 484
- PDF下载量: 51
- 被引次数: 0
