




-
白斑综合征病毒(WSSV)是线头病毒科Nimaviridae白斑病毒属Whispovirus的唯一成员[1]。在电子显微镜下可以观察到WSSV的核衣壳呈棒状,核衣壳表面有3层膜交叉覆盖,并形成阴影,核衣壳的一端还有一个独特的尾状结构。WSSV的核衣壳由14个环组成,其中包含1个约300 kb的圆形dsDNA基因组[2],是迄今为止被完全测序的最大动物病毒之一[3]。在养殖条件下,感染WSSV的虾类可在3~10 d内死亡[4]。另外,WSSV还会感染其他甲壳类动物,其中就包括克氏原螯虾Procambarus clarkii[5−6]。WSSV危害大、传播快,已对中国虾类养殖业造成了数十亿美元的经济损失。然而常规的化学和物理方法并不能有效地控制WSSV对虾类的感染。因此,找到并采取合理的预防手段才是控制该病毒病的关键。有关物理和化学消毒方法对WSSV的灭活作用已有较多报道[7−10],已确定了温度、紫外线、pH、臭氧、盐度、干燥和化学消毒剂等因素对WSSV灭活的影响。有研究发现:大部分化学试剂对WSSV灭活后,WSSV的免疫原性会受到损害[7, 9−11],而双乙烯亚胺(BEI)对免疫原性的损害相对较小,因此BEI是对含有DNA或RNA基因组动物病毒的首选灭活剂[12−14]。基于此,本研究从WSSV灭活制剂着手,用BEI灭活WSSV,通过口服和注射途径分别建立克氏原螯虾的主动免疫体系,再对其进行抗WSSV感染的效果研究,分析WSSV灭活制剂对WSSV的防治效果,为防治WSSV提供科学依据。
-
LAI等[15]研究可知:超声破碎WSSV后获得的蛋白质和核酸可以感染克氏原螯虾并导致其死亡,其有效性与完整的WSSV病毒一样。因此,本研究对病毒进行超声破碎,之后进行BEI灭活,观察BEI灭活制剂是否具有与超声破碎制剂相同的抗WSSV效果。本研究分为口服和注射免疫,免疫7 d后再进行WSSV攻毒感染。具体分组如表1所示。克氏原螯虾购于安徽宣城,养殖于充气水箱中,水温为25 ℃,每日投喂克氏原螯虾专用饲料。
表 1 WSSV灭活制剂对克氏原螯虾抗WSSV的实验分组
Table 1. Experimental groups of anti-WSSV effects of WSSV inactivated preparation on P. clarkii
组别 实验内容 组别 实验内容 1 口服WSSV对照 6 注射BEI灭活WSSV破碎液12 h,免疫7 d后攻毒 2 注射WSSV对照 7 口服BEI灭活WSSV破碎液24 h,免疫7 d后攻毒 3 注射WSSV超声破碎液对照 8 口服BEI灭活WSSV破碎液1 h,免疫7 d后攻毒 4 注射BEI灭活WSSV破碎液24 h,免疫7 d后攻毒 9 口服BEI灭活WSSV破碎液12 h,免疫7 d后攻毒 5 注射BEI灭活WSSV破碎液1 h,免疫7 d后攻毒 10 磷酸盐缓冲溶液(PBS)阴性对照 -
取经过WSSV感染的克氏原螯虾的尾部肌肉,参考ZHU等[16]的方法提纯WSSV。使用高速分散器将WSSV提纯液快速分散呈匀浆,用超高速冷冻离心机在6 000 g下离心10 min,分离上清液后再次离心,重复3次。离心完成后,肌肉组织已基本去除,WSSV病毒存在于上清液中。将上清液通过300目细胞筛过滤分散后,用离心机在6000 g下离心30 min。再将分离出的上清液通过450 nm的滤膜后,用离心机在30 000 g下离心30 min,最后得到WSSV病毒粒子的白色沉淀。将提取到的病毒粒子用10 mL 磷酸盐缓冲溶液(PBS)溶解后,使用FS 200 T超声波均质机(上海盛溪)破碎溶解的病毒颗粒。破碎完成后,在30 000 g条件下对超声破碎的病毒粒子离心30 min,离心得到的上清液便是WSSV病毒粒子的破碎液,使用100 nm滤膜进行过滤,即为WSSV超声破碎液。
-
将0.12 mol·L−1BEI溶液按1∶60的体积比加入到病毒液中,使BEI的最终分子浓度为2 mmol·L−1。在37 ℃下恒温灭活3个不同时间段(1、12、24 h)的病毒液,期间每2 h搅拌1次。经过规定时间后,剩余BEI以50∶1的体积比例加入质量分数2%的硫代硫酸钠使灭活中止,制成WSSV灭活制剂。每100 mL WSSV灭活制剂中含45 mg WSSV粒子,WSSV灭活制剂中WSSV的质量浓度为0.45 g·L−1。
-
克氏原螯虾养殖于(25±1) ℃控温玻璃缸中,随机分成10组,每组20只,每组设3个重复,投喂配合饲料暂养7 d。将WSSV超声破碎液与饲料混合均匀,混合5 min后用蜂胶液包被,风干10 min后投喂受试克氏原螯虾。将WSSV提纯液与克氏原螯虾饲料混合均匀,混合5 min后用蜂胶液包被,风干10 min后投喂受试克氏原螯虾。7 d后所有实验组进行攻毒研究,期间保证每组克氏原螯虾摄食等量的饲料,同时记录每日死亡率。
-
从表2可见:经BEI处理WSSV 24 h后,注射和口服免疫克氏原螯虾的死亡率为0,说明没有感染性;而BEI灭活WSSV破碎处理液1 h的注射死亡率为100.0%,口服死亡率为10.0%,BEI处理12 h的注射死亡率为33.3%,口服死亡率为0,说明口服免疫更安全,注射免疫危险性比较高,但是BEI处理时间过短也会使病毒没有完全灭活,因此BEI灭活24 h较为安全,后续实验均采用BEI灭活WSSV破碎液24 h的制剂。
表 2 BEI灭活WSSV感染克氏原螯虾的死亡率
Table 2. Mortality of P. clarkii infected with BEI-inactivated WSSV
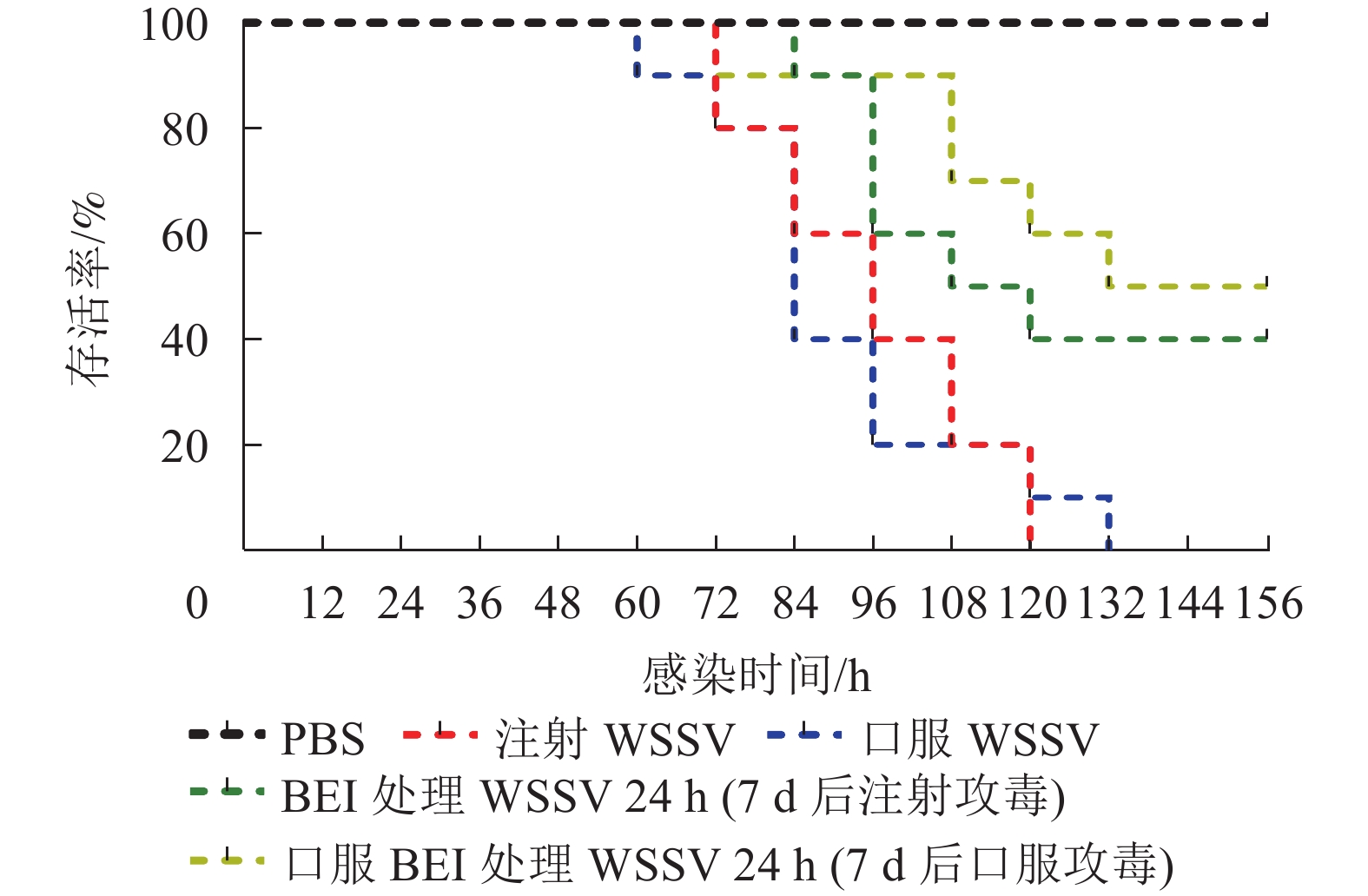
组别 灭活时间/h 死亡率/% 注射BEI灭活WSSV破碎液 1 100.0 12 33.3 24 0 口服BEI灭活WSSV破碎液 1 10.0 12 0 24 0 从表3可见:WSSV无论是通过口服感染还是注射感染,死亡率均为100%。用BEI灭活WSSV破碎液1和12 h分别采用注射或口服免疫克氏原螯虾,虽然免疫过程中死亡了一些,但将活着的克氏原螯虾7 d后攻毒,口服的存活率为30%~40%,注射的效果较差,最后的存活率为20%(图1)。以上结果说明灭活时间延长后,不但灭活更完全,而且灭活制剂的保护效果更好,因此在后续的研究中采用了24 h的灭活时间。用BEI灭活WSSV破碎液24 h制成灭活制剂,采用注射或口服免疫克氏原螯虾7 d后分别注射、口服攻毒,结果发现WSSV灭活制剂的口服免疫效果明显,比起注射WSSV组和口服WSSV组的100%死亡率,注射免疫的克氏原螯虾在攻毒后存活率为40%,口服免疫的存活率为50%(图2);经灭活制剂免疫7 d后,再去感染WSSV,极大地降低了死亡率,免疫效果比较明显。
表 3 BEI灭活WSSV制剂免疫7 d后攻毒克氏原螯虾的死亡率
Table 3. Mortality rate of P. clarkii after 7 days of BEI-inactivated WSSV (ruptured ultrasonically) immunization
组别 灭活时间/h 死亡率/% PBS 0 0 注射WSSV 0 100 口服WSSV 0 100 口服BEI灭活WSSV破碎液 1 70 口服BEI灭活WSSV破碎液 12 60 注射BEI灭活WSSV破碎液 12 80 注射BEI灭活WSSV(7 d后注射攻毒) 24 60 口服BEI灭活WSSV(7 d后口服攻毒) 24 50 
图 1 BEI灭活WSSV破碎液免疫7 d后攻毒克氏原螯虾的存活率
Figure 1. Survival rate of P. clarkii after 7 days of BEI-inactivated WSSV (ruptured ultrasonically) immunization

图 2 BEI灭活WSSV制剂免疫7 d后攻毒克氏原螯虾的存活率
Figure 2. Survival rate of P. clarkii after 7 days of BEI-inactivated WSSV immunization
-
已有研究表明:WSSV囊膜蛋白可以诱导甲壳动物对WSSV产生抵抗力,从而大幅提高WSSV攻毒后的成活率;多糖(如葡聚糖、壳聚糖、甲壳素)也被认为能提高甲壳动物的免疫力,并被广泛用于提高甲壳动物抗细菌性和病毒性疾病的研究[6−8]。由BEI灭活的WSSV灭活制剂,可以保护WSSV囊膜蛋白的完整性[16],使其免疫原性最大限度不受破坏。
本研究表明:用BEI灭活的WSSV灭活制剂免疫接种克氏原螯虾7 d后,可在一定程度上保护克氏原螯虾免受WSSV的侵害。克氏原螯虾对WSSV非常敏感,通过口服和注射途径的克氏原螯虾死亡率都超过90%[17]。在本次研究中,随着灭活时间的延长,注射和口服2 mmol·L−1 BEI灭活WSSV制剂的克氏原螯虾存活率越来越高,说明只要灭活时间足够,使用BEI灭活的WSSV灭活制剂可以成功地保护克氏原螯虾免受WSSV的侵害。NAMIKOSHI等[11]使用甲醛灭活的WSSV也获得了类似的结果,受免疫保护的对虾Penaeus japonicus在WSSV攻毒实验中存活率显著升高。同样,紫外线灭活的WSSV对日本对虾Marsupenaeus japonicus也有一定的免疫保护效果[18]。这就引发了甲壳动物中是否存在免疫记忆的猜想[19],但要证实其中的免疫机制还有待更深入的研究。
综上所述,BEI灭活的WSSV免疫克氏原螯虾的效果十分明显,并且口服免疫的保护效果优于注射免疫。与BEI的灭活纯病毒液24 h相比,BEI灭活的WSSV破碎液减少了灭活时间,但最终也是口服免疫的效果较好。用BEI灭活的WSSV灭活制剂饲喂克氏原螯虾后,可以有效地降低克氏原螯虾感染病毒的死亡率,说明用BEI灭活的WSSV灭活制剂对克氏原螯虾抗WSSV具有保护性效果,可成为抗白斑综合征的一种免疫策略。
-
感谢浙江杭州康德权科技有限公司的经费支持。
Anti-WSSV effect of inactivated preparation of white spot syndrome virus (WSSV) on Procambarus clarkii
-
摘要:
目的 研究经双乙烯亚胺(BEI)灭活的白斑综合征病毒(WSSV)灭活制剂保护克氏原螯虾Procambarus clarkii抗WSSV感染的效果,以期为白斑综合征的防治提供有效的免疫方法。 方法 应用BEI对WSSV和WSSV超声破碎液进行灭活,通过口服和注射分别对克氏原螯虾进行免疫,再对其进行抗WSSV感染的效果研究。 结果 WSSV在BEI处理24 h可以被完全灭活,通过口服途径用灭活制剂免疫克氏原螯虾7 d后攻毒,克氏原螯虾死亡率显著下降,且口服免疫的效果要好于注射免疫。 结论 经BEI灭活24 h的WSSV灭活制剂对克氏原螯虾是安全的,口服免疫后可以显著降低克氏原螯虾感染白斑综合征病毒的死亡率。图2表3参19 -
关键词:
- 白斑综合征病毒 /
- 双乙烯亚胺(BEI) /
- 灭活 /
- 克氏原螯虾
Abstract:Objective This study is to explore the protective effect of white spot syndrome virus (WSSV) preparation inactivated by binary ethyleneimine (BEI) on Procambarus clarkii against WSSV infection, so as to provide an effective immune method for the prevention of WSSV. Method Starting with the preparation of WSSV inactivation preparations, BEI was applied to inactivate WSSV, immunize P. clarkii by oral and injection methods, and then conduct anti-WSSV infection test. Result WSSV could be completely inactivated under BEI treatment for 24 hours. After 7 days of oral immunization with inactivated WSSV, the mortality rate of P. clarkii was significantly decreased. The effect of oral immunization was better than that of injection immunization. Conclusion The inactivated WSSV preparation inactivated by BEI for 24 hours is safe for P. clarkii and can significantly reduce its mortality infected with WSSV. [Ch, 2 fig. 3 tab. 19 ref.] -
图 1 BEI灭活WSSV破碎液免疫7 d后攻毒克氏原螯虾的存活率
Figure 1 Survival rate of P. clarkii after 7 days of BEI-inactivated WSSV (ruptured ultrasonically) immunization
图 2 BEI灭活WSSV制剂免疫7 d后攻毒克氏原螯虾的存活率
Figure 2 Survival rate of P. clarkii after 7 days of BEI-inactivated WSSV immunization
表 1 WSSV灭活制剂对克氏原螯虾抗WSSV的实验分组
Table 1. Experimental groups of anti-WSSV effects of WSSV inactivated preparation on P. clarkii
组别 实验内容 组别 实验内容 1 口服WSSV对照 6 注射BEI灭活WSSV破碎液12 h,免疫7 d后攻毒 2 注射WSSV对照 7 口服BEI灭活WSSV破碎液24 h,免疫7 d后攻毒 3 注射WSSV超声破碎液对照 8 口服BEI灭活WSSV破碎液1 h,免疫7 d后攻毒 4 注射BEI灭活WSSV破碎液24 h,免疫7 d后攻毒 9 口服BEI灭活WSSV破碎液12 h,免疫7 d后攻毒 5 注射BEI灭活WSSV破碎液1 h,免疫7 d后攻毒 10 磷酸盐缓冲溶液(PBS)阴性对照  下载: 导出CSV
下载: 导出CSV
表 2 BEI灭活WSSV感染克氏原螯虾的死亡率
Table 2. Mortality of P. clarkii infected with BEI-inactivated WSSV
组别 灭活时间/h 死亡率/% 注射BEI灭活WSSV破碎液 1 100.0 12 33.3 24 0 口服BEI灭活WSSV破碎液 1 10.0 12 0 24 0
下载: 导出CSV
表 3 BEI灭活WSSV制剂免疫7 d后攻毒克氏原螯虾的死亡率
Table 3. Mortality rate of P. clarkii after 7 days of BEI-inactivated WSSV (ruptured ultrasonically) immunization
组别 灭活时间/h 死亡率/% PBS 0 0 注射WSSV 0 100 口服WSSV 0 100 口服BEI灭活WSSV破碎液 1 70 口服BEI灭活WSSV破碎液 12 60 注射BEI灭活WSSV破碎液 12 80 注射BEI灭活WSSV(7 d后注射攻毒) 24 60 口服BEI灭活WSSV(7 d后口服攻毒) 24 50
下载: 导出CSV
-
[1] FAUQUET C M, MAYO M A, MANILOFF J, et al. Virus Taxonomy: Eighth Report of the International Committee on Taxonomy of Viruses [M]. San Diego: Elsevier Academic Press, 2005. [2] ESCOBEDO-BONILLA C M, ALDAY-SANZ V, WILLE M, et al. A review on the morphology, molecular characterization, morphogenesis and pathogenesis of white spot syndrome virus [J]. Journal of Fish Diseases, 2008, 31(1): 1 − 18. [3] van HULTEN M C, WITTEVELDT J, PETERS S, et al. The white spot syndrome virus DNA genome sequence [J]. Virology, 2001, 286: 7 − 22. [4] LIGHTNER D V. A Handbook of Pathology and Diagnostic Procedures for Diseases of Penaeid Shrimp [M]. Baton Rouge: World Aquaculture Society, 1996. [5] HOSSAIN S, CHAKRABORTY A, JOSEPH B, et al. Detection of new hosts for white spot syndrome virus of shrimp using nested polymerase chain reaction [J]. Aquaculture, 2001, 198: 1 − 11. [6] LO Chufang, HO Chinghui, PENG Shaoen, et al. White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods [J]. Diseases of Aquatic Organisms, 1996, 27: 215 − 225. [7] CHANG Pohshing, CHEN Lijing, WANG Yuchi. The effect of ultraviolet irradiation, heat, pH, ozone, salinity and chemical disinfectants on the infectivity of white spot syndrome baculovirus [J]. Aquaculture, 1998, 166(1/2): 1 − 17. [8] WANG Yuchi, LO Chufang, CHANG Pohshing, et al. Experimental infection of white spot baculovirus in some cultured and wild decapods in Taiwan [J]. Aquaculture, 1998, 164: 221 − 231. [9] BALASUBRAMANIAN G, SUDHAKARAN R, MUSTHAQ S S, et al. Studies on the inactivation of white spot syndrome virus (WSSV) of shrimp by physical and chemical treatment, and seaweed extracts tested in marine and freshwater animal models [J]. Journal of Fish Diseases, 2006, 29(9): 569 − 572. [10] OSEKO N, CHUAH T T, MAENO Y, et al. Examination for viral inactivation of WSSV (white spot syndrome virus) isolated in Malaysia using black tiger prawn (Penaeus monodon) [J]. Japan Agricultural Research Quarterly, 2006, 40(1): 93 − 97. [11] NAMIKOSHI A, WU Jinlu, YAMASHITA T, et al. Vaccination trials with Penaeus japonicus to induce resistance to white spot syndrome virus [J]. Aquaculture, 2004, 229: 25 − 35. [12] BAHNEMANN H G. Binary ethylenimine as an inactivant for foot and mouth disease virus and its application for vaccine production [J]. Archives of Virology, 1975, 47(1): 47 − 56. [13] BAHNEMANN H G. Inactivation of viruses in serum with binary ethyleneimine [J]. Journal of Clinical Microbiology, 1976, 3(2): 209 − 210. [14] AARTHIA D, RAOA K A, ROBINSONB R, et al. Validation of binary ethyleneimine (BEI) used as an inactivant for foot and mouth disease tissue culture vaccine [J]. Biologicals, 2004, 32(3): 153 − 156. [15] LAI Yongyong, ZHU Fei, XU Yinglei. WSSV proteins and DNA genome released by ultrasonic rupture can infect crayfish as effectively as intact virions [J/OL]. Journal of Virological Methods, 2020, 283: 113917[2022-07-01]. doi: 10.1016/j.jviromet.2020.113917. [16] ZHU Fei, DU Huahua, MIAO Zhiguo, et al. Protection of Procambarus clarkii against white spot syndrome virus using inactivated WSSV [J]. Fish Shellfish Immunology, 2009, 26(5): 685 − 690. [17] JHA R K, XU Zirong, PANDEY A. Protection of Procambarus clarkii against white spot syndrome virus using vaccine expressed in Pichia pastoris [J]. Fisheries Science, 2006, 7: 1011 − 1019. [18] ZANG Shaoqing, LÜ Lixia, LIU Chenfei, et al. Metabolomic investigation of ultraviolet ray-inactivated white spot syndrome virus-induced trained immunity in Marsupenaeus japonicus [J/OL]. Frontiers in Immunology, 2022, 13: 885782[2022-07-01]. doi: 10.3389/fimmu.2022.885782. [19] CHANG Yuhsuan, KUMAR R, NG T H, et al. What vaccination studies tell us about immunological memory within the innate immune system of cultured shrimp and crayfish [J]. Developmental and Comparative Immunology, 2018, 80(1): 53 − 66. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220459
 点击查看大图
点击查看大图

计量
- 文章访问数: 1735
- HTML全文浏览量: 801
- PDF下载量: 38
- 被引次数: 0
