





-
牦牛Bos grunniens是中国青藏高原及其毗邻高山、亚高山高寒地区的特有珍稀牛种之一[1]。作为牦牛肉加工过程中产生的主要副产物,牦牛骨钙质丰富,是目前市面上优质的小分子骨胶原肽来源[2]。多肽螯合钙是第3代具有活性结构的生物钙补钙制剂,稳定性好,抗干扰能力强,吸收效果好,生物利用度高。甘林火等[3]发现:L-亮氨酸螯合钙的生物利用度比葡萄糖酸钙要高,具有缓释、药效时间长、不需要消耗胃酸等特点。植物来源的精油可以作为调味剂和天然防腐剂,肉桂精油具有清除自由基、抗油脂氧化的作用和较强的抗菌能力[4],是对抗常见食源性病原体的最有效精油之一[5],主要成分为肉桂醛,此外还含有丁子香酚、芳樟醇和α-pine烯等[6]。HUANG等[7]发现:肉桂精油抗菌活性高,能有效抑制微生物生长,延长草鱼Ctenopharyngodon idellus鱼片保质期。OJAGH等[8]发现富含肉桂油的壳聚糖涂层延长了虹鳟鱼Oncorhynchus mykiss冷藏期间的保质期。本研究利用牦牛骨资源,制备包埋肉桂精油的牦牛骨蛋白肽-钙螯合物,探讨其对水产品中的优势腐败菌维氏气单胞菌Aeromonas veronii的缓释抑菌作用,为水产品的防腐保鲜提供新思路。
-
木瓜蛋白酶购自安琪酵母股份有限公司;维氏气单胞菌由中国农业大学食品科学与营养工程学院水产品加工实验室提供;牦牛骨购自国肽生物科技(北京)有限公司;肉桂精油购自江西恒城天然香料油有限公司。肉桂精油的添加使用遵照GB 2760−2014《食品安全国家标准 食品添加剂使用标准》的相关规定[9]。
-
参考魏洁琼等[10]方法并稍作修改。破碎后的牦牛骨粒经高压灭菌锅蒸煮3.0 h;冷却后除去油脂,过滤得到骨渣和滤液。骨渣∶盐酸(1.0 mmol·L−1)以1∶50(m∶V)酸溶15 h[11],蒸发浓缩,EDTA滴定后用去离子水稀释得到2.0 mmol·L−1的可溶性牦牛骨钙,备用。滤液浓缩,冷冻干燥,得到牦牛骨蛋白;去离子水溶解,调节至60 ℃,pH 7.0[12-13],木瓜蛋白酶(4 000×16.67 nkat·g−1)酶解2.0 h[14],95 ℃水浴灭酶20 min,冷却后离心,取上清液浓缩后冷冻干燥,即为牦牛骨蛋白肽。
-
参照HONG等[15]方法并稍作修改。牦牛骨胶原蛋白肽用去离子水溶解成10.0 g·L−1蛋白肽溶液,离心去除沉淀。在使用均质仪(德国IKA)均质的同时向牦牛骨蛋白肽溶液中逐滴加入牦牛骨钙溶液,得到牦牛骨蛋白肽-钙螯合物。量取10 mL牦牛骨蛋白肽溶液,分别加入0(对照)、10、15、20、25、30 μL肉桂精油,在均质的同时逐滴加入1 mL 0(对照)、0.5、1.0、2.0 mmol·L−1牦牛骨钙溶液,得到包埋肉桂精油的牦牛骨蛋白肽-钙螯合物[16]。
-
参照XIE等[17]方法,通过分子量排阻的高效液相色谱法(SE-HPLC),使用液相色谱仪(日本岛津)测定牦牛骨蛋白肽分子量[18]。仪器参数:TSK凝胶G2000SWXL柱(7.8 mm×300 .0 mm),进样量25.0 μL,流速0.5 mL·min−1,紫外检测器波长214 nm。以三氟乙酸-水-乙腈(体积比为0.1∶54.9∶45.0)为流动相。标准物为细胞色素C(12.362 kDa),抑肽酶(6.5114 kDa),杆菌肽(1.423 kDa),Gly-Gly-Tyr-Arg(四肽,0.451 kDa)和Gly-Gly-Gly(三肽,0.189 kDa)。将混合标准溶液进样,以标准物分子量(M)的对数值为纵坐标,保留时间(t)为横坐标,建立保留时间和分子量对数之间的标准曲线。
-
使用Q-Exactive高分辨质谱仪(美国赛默飞)对样品进行液相色谱串联质谱(LC-MS/MS)全扫描,质荷比为100~1 990 Da[19]。使用Peaks studio 8.0软件结合Mascot MS/MS搜索SwissProt蛋白质数据库,鉴定牦牛骨多肽序列[20]。
-
根据AKBARI等[16]和ZHANG等[21]方法稍作修改,使用可见分光光度计每隔10 min测定1次牦牛骨蛋白肽原始溶液、牦牛骨蛋白肽-钙螯合物和去离子水乳化的牦牛骨蛋白肽(对照)的吸光度(600 nm),计算相对浊度,研究蛋白肽钙螯合物的稳定性。计算公式为:相对浊度=(Ax−A0)/A0×100%。其中:Ax为不同条件下制得的牦牛骨蛋白肽-钙螯合物(或对照)的吸光度,A0为牦牛骨蛋白肽原始溶液的吸光度。
每隔2 h测定1次包埋肉桂精油的牦牛骨蛋白肽-钙螯合物的吸光度(600 nm)[22],以牦牛骨蛋白肽-钙螯合物为对照,研究肉桂精油对螯合物稳定性的影响。
使用ZS90纳米粒径电位分析仪(英国,马尔文)在25 ℃下通过动态光散射(DLS)测定螯合物的平均粒径和多分散指数(PDI)[21]。
-
采用感官评价法和抑菌活性测定缓释效果。取包埋肉桂精油的牦牛骨蛋白肽-钙螯合物和去离子水稀释的等比例肉桂精油样品各1份,室温下静置24 h后,参考朱红梅等[23]进行感官评价。感官指标评分表如表1所示。参照周子雄等[24]的方法并稍作修改测定抑菌活性。活化过夜的维氏气单胞菌菌液[25-26]调整至106 CFU·mL−1,取4份25 mL受试菌菌悬液,分别加入100 μL已灭菌的包埋肉桂精油的牦牛骨蛋白肽-钙螯合物、牦牛骨蛋白肽-钙螯合物、肉桂精油和去离子水(对照),震荡混匀。每隔2 h测定1次菌悬液吸光度(600 nm),检测螯合钙的抑菌效果。抑菌率=(Ac−At)/Ac×100%。其中:Ac为对照组的吸光度,At为不同处理条件下的吸光度。
表 1 感官评价评分标准
Table 1. Standard of sensory evaluation
评分 状态 色泽 香气 异味 协调性 7~9 分散均一,无分层、结块、沉淀 一致,无杂质、霉变 肉桂味浓郁、纯正 气味良好,
无不良风味气味协调 4~6 分散略不均一,无分层、结块、沉淀 不均,有杂质,无霉变 肉桂味较浓郁 稍有异味 气味较协调 1~3 分散不均一,有明显分层、结块、沉淀,
有油滴出现不均,有大量杂质,有霉变 肉桂味不突出 有明显异味 气味不协调 -
所有实验重复3次,数据用平均值±标准差表示。采用RStudio 1.3.1073软件对测定结果进行显著性分析,差异显著水平为0.05。
-
标准曲线方程为:lgM=7.013 8+0.226 9t,计算得到不同分子量区间的蛋白肽占总蛋白的比例。由表2可知:酶解后的牦牛骨蛋白肽肽段的分子量均小于3.0 kDa,其中分子量小于1.0 kDa的牦牛骨蛋白肽占全部肽段的95.867%,说明牦牛骨蛋白肽多为小分子肽。
表 2 牦牛骨蛋白肽的分子量
Table 2. Molecular weight of yak bone peptides
t/min 分子量/kDa 占比/% <15.579 ≥3.0 0 15.579~<16.363 2.0~<3.0 0.040 16.363~<17.688 1.0~<2.0 4.094 17.688~<18.508 0.5~<1.0 34.211 ≥18.508 <0.5 61.656 -
LC-MS/MS测序发现:牦牛骨蛋白肽主要成分为胶原Ⅰ蛋白α1链和α2链,分别含肽段336和285条,肽段覆盖率为55%。共测得945个肽段序列信息,相对分子量为306~1 032 Da,组成肽段的氨基酸为7~37个。对其中分子量小于1.0 kDa且可信度较高的33条蛋白肽段(表3)进一步的分析,牦牛骨蛋白肽可用于制备具备缓释杀菌功能的钙螯合物。
表 3 牦牛骨蛋白肽部分多肽序列鉴定结果
Table 3. Identification of partial peptides sequences of yak bone peptides
主要肽段 可信度 分子量/Da 质核比 功能 GPVGPVG 40.23 581.317 3 582.324 1 SVPGPMG 32.33 643.299 9 644.306 6 VPGPMGP 31.42 653.320 7 654.326 5 APGPVGPAG 34.80 721.375 9 722.382 3 AP(+15.99)GPVGPAG 31.27 737.370 8 738.377 4 羟基化 FP(+15.99)GADGVA 34.14 748.339 1 749.345 7 羟基化 VP(+15.99)GPMGPS 33.76 756.347 6 757.356 1 羟基化 SDGSVGPVG 36.97 773.355 5 774.362 2 SP(+15.99)GN(+0.98)IGPAG 33.70 785.355 5 786.362 5 羟基化 GAAGPTGPIG 30.08 796.407 9 797.415 6 TPGPQ(+0.98)GIAG 30.34 797.391 9 798.399 9 脱酰胺 LPQPPQE 31.34 807.412 7 808.419 7 IQ(+0.98)GPP(+15.99)GPAG 30.19 809.391 9 810.398 9 脱酰胺,羟基化 AP(+15.99)GPQ(+0.98)GFQ 31.21 817.360 6 818.366 5 羟基化,脱酰胺 GPP(+15.99)GPVGPAG 33.96 820.407 9 821.415 4 羟基化 GP(+15.99)AGPIGPVG 34.90 836.439 1 837.445 6 羟基化 GPVGPTGPVG 37.77 836.439 2 837.445 6 TP(+15.99)GPQGLLG 31.36 854.449 8 855.456 9 羟基化 GASGPP(+15.99)GFVG 31.32 860.402 8 861.412 9 羟基化 GPQ(+0.98)GPVGPVG 31.77 864.434 1 865.440 8 脱酰胺 GIQGPP(+15.99)GPAG 30.73 865.429 3 866.436 9 羟基化 AGPSGPAGPTG 36.13 867.408 6 868.415 5 GPP(+15.99)GLQ(+0.98)GVQ 33.31 868.429 0 869.436 0 羟基化,脱酰胺 GPP(+15.99)GPMGPPG 42.39 878.395 6 879.402 2 羟基化 TGPIGPP(+15.99)GPA 41.22 878.449 8 879.457 6 羟基化 GPP(+15.99)GPIGNVG 33.95 879.445 0 880.452 9 羟基化 GPP(+15.99)GPIGN(+0.98)VG 32.42 880.429 0 881.436 0 羟基化,脱酰胺 GPP(+15.99)GFP(+15.99)GAVG 34.44 886.418 5 887.424 9 羟基化 VGPAGPN(+0.98)GFA 40.33 886.418 5 887.425 0 脱酰胺 AGPP(+15.99)GPTGPAG 31.53 893.424 3 894.431 5 羟基化 AAGPP(+15.99)GPTGPA 34.87 907.439 9 908.446 5 羟基化 VGPAGPN(+0.98)GFAG 38.51 943.439 9 944.445 9 脱酰胺 APGAPGPVGPAG 30.84 946.487 2 947.493 7 -
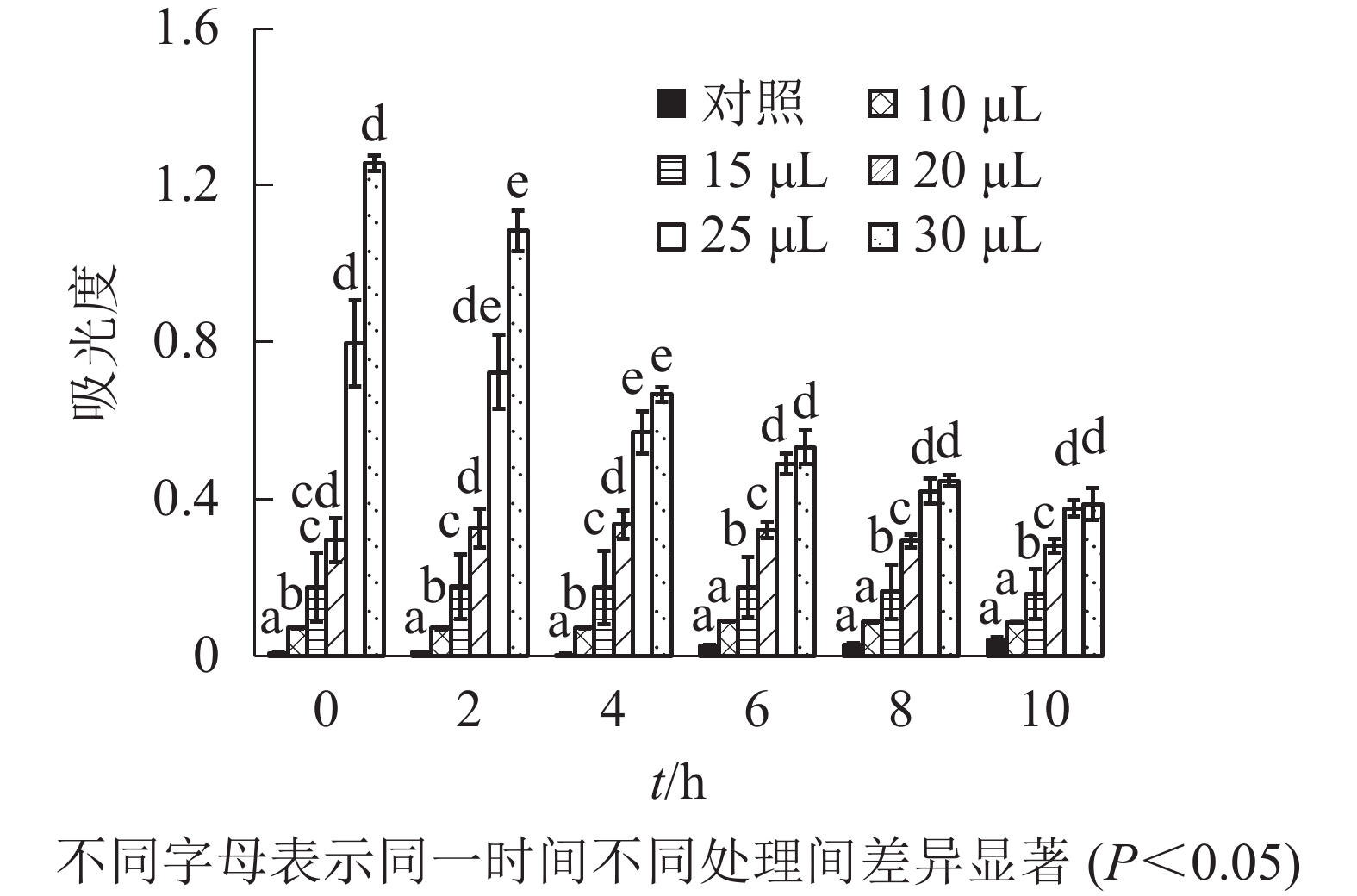
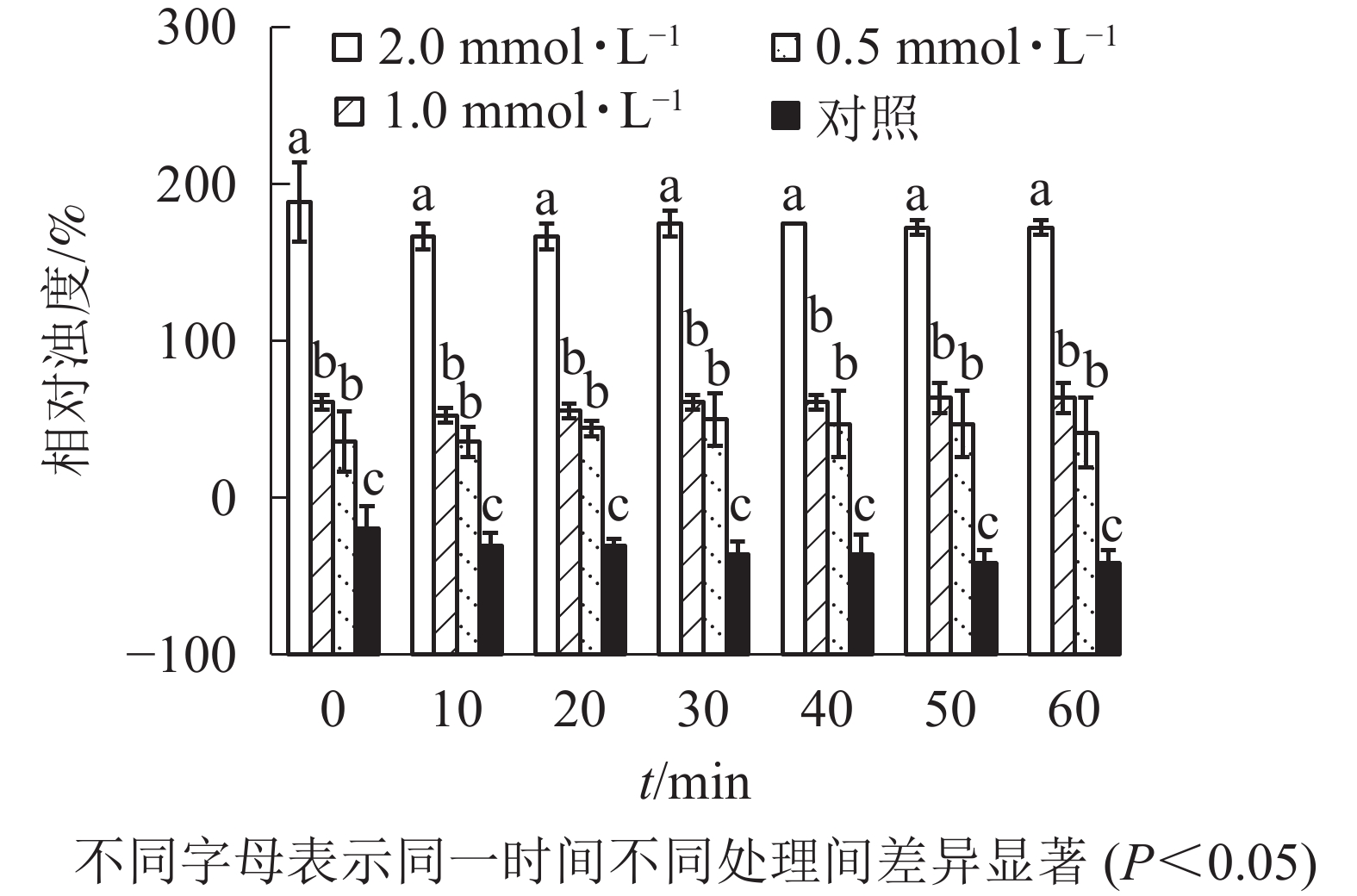
由图1可知:除加入去离子水并乳化的牦牛骨蛋白肽(对照)外,不同浓度牦牛骨钙溶液制成的牦牛骨蛋白肽-钙螯合物的相对浊度存在显著差异(P<0.05)。随牦牛骨钙溶液浓度增加,蛋白肽-钙螯合物相对浊度增加,但不同沉降时间下相对浊度变化不大,说明鳌合效果良好,螯合物稳定,可以用于混合乳液的制备,后续研究选用2.0 mmol·L−1牦牛骨钙溶液制备鳌合物。由图2可知:除牦牛骨蛋白肽-钙螯合物(对照)外,添加不同体积肉桂精油制成的包埋肉桂精油的牦牛骨蛋白肽-钙螯合物的吸光度存在显著差异(P<0.05)。当添加的肉桂精油大于20 μL时,螯合物吸光度随时间急剧下降,稳定性较差;添加的肉桂精油小于20 μL时,螯合物吸光度变化不大,说明稳定性较好。因此后续研究以10 mL牦牛骨蛋白肽中加入20 μL肉桂精油为螯合时肉桂精油的适宜添加量。
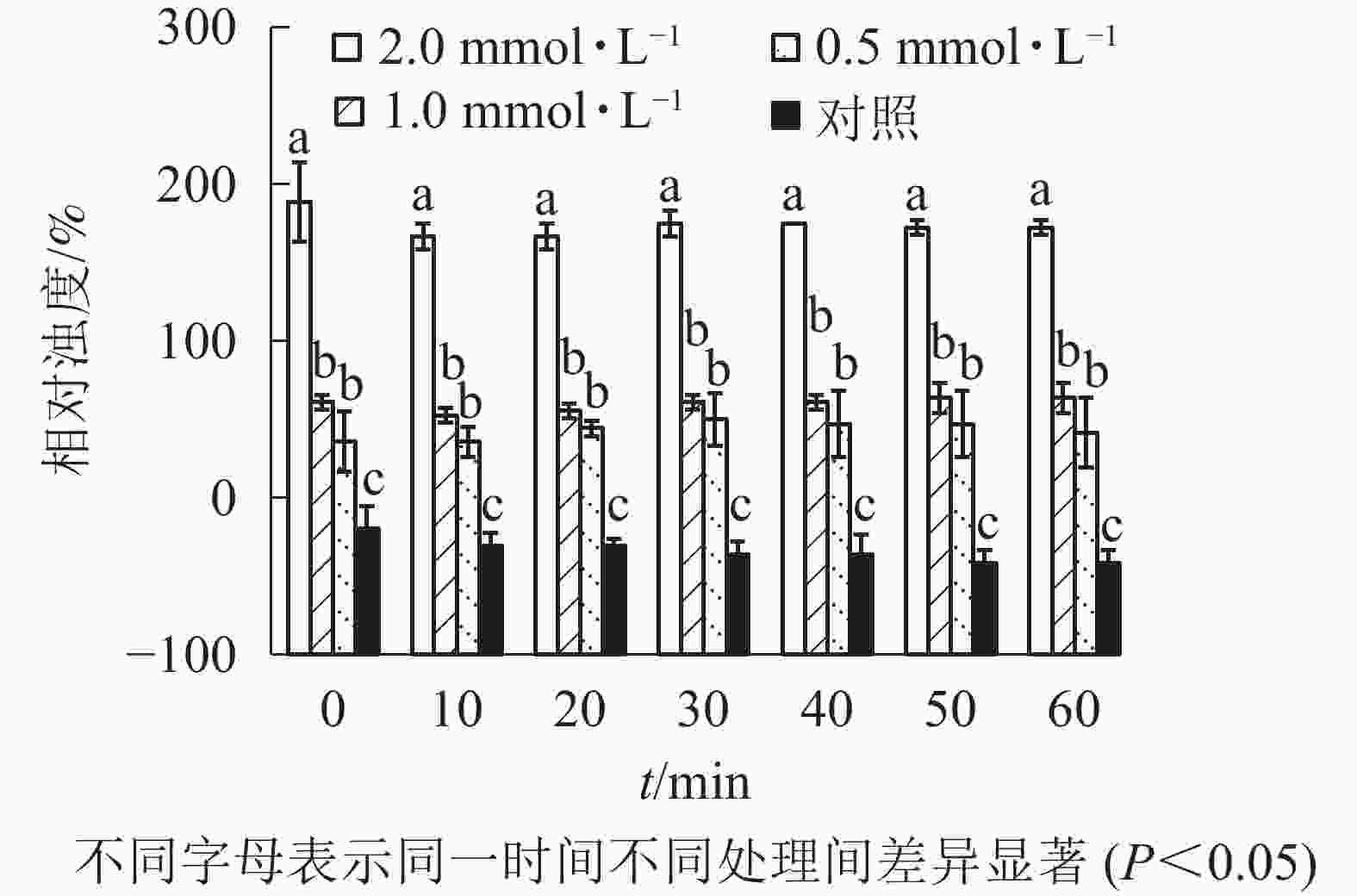
图 1 牦牛骨钙溶液浓度对牦牛骨蛋白肽-钙螯合物稳定性的影响
Figure 1. Stability of yak bone peptides-calcium chelate prepared by different yak bone calcium solution
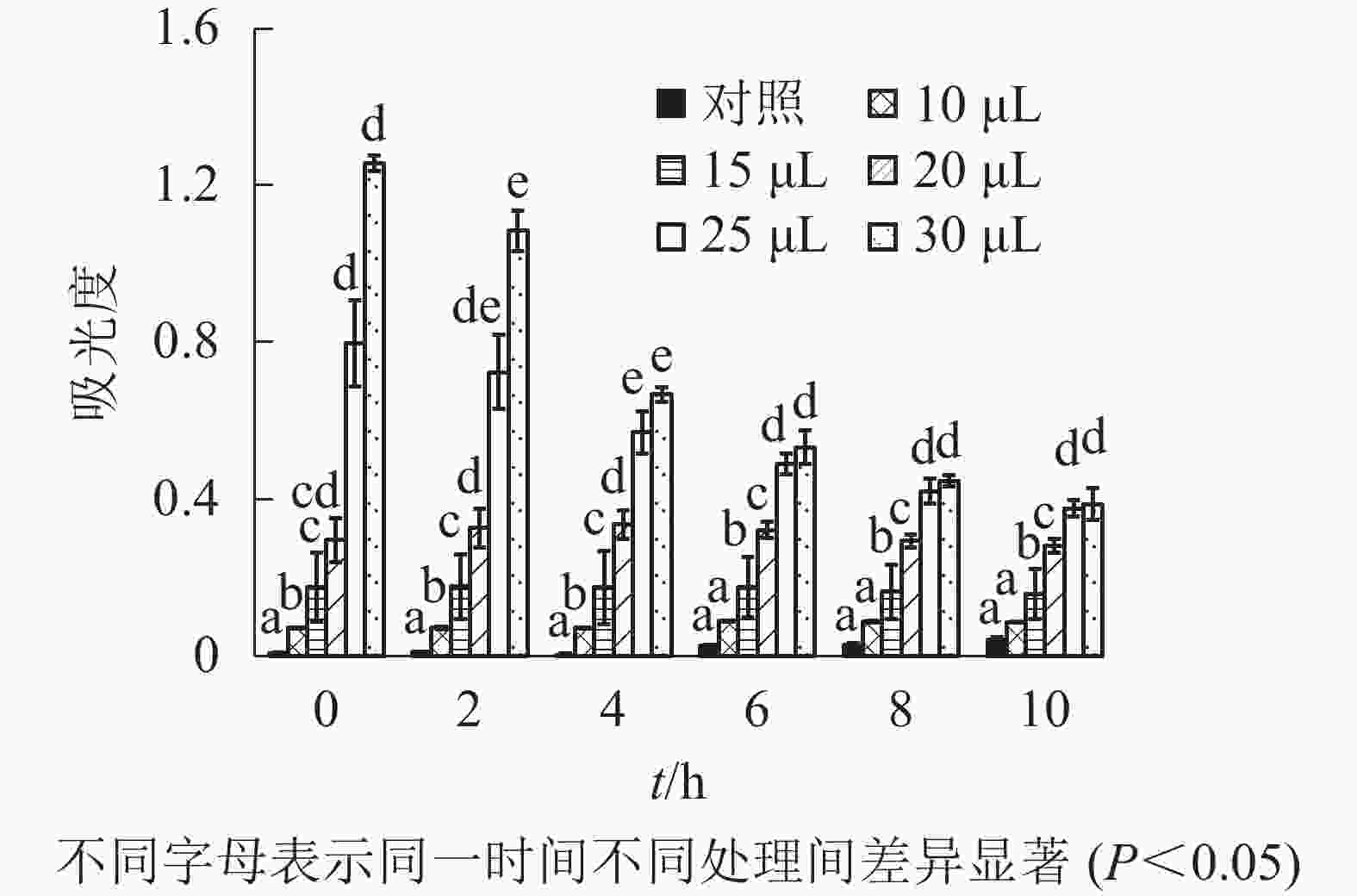
图 2 不同肉桂精油添加量对包埋肉桂精油的牦牛骨蛋白肽-钙螯合物稳定性的影响
Figure 2. Stability of yak bone peptides-calcium chelate prepared by different ratio of cinnamon essential oil
-
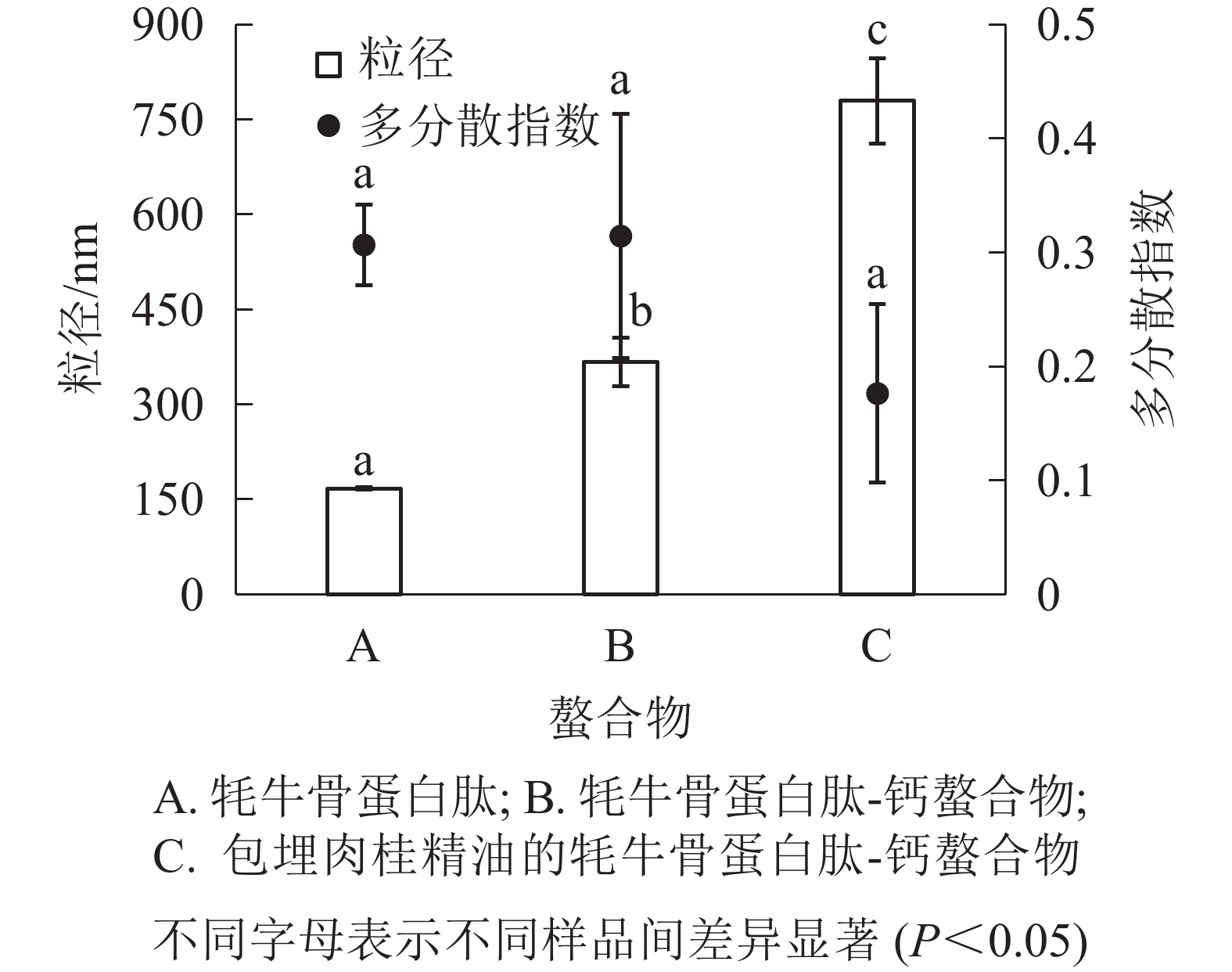
由图3可知:牦牛骨蛋白肽的粒径为167.8 nm,牦牛骨蛋白肽-钙螯合物的粒径为368.2 nm,包埋肉桂精油的牦牛骨蛋白肽-钙螯合物的粒径为780.2 nm,三者差异显著(P<0.05);牦牛骨蛋白肽、牦牛骨蛋白肽-钙螯合物及包埋肉桂精油的牦牛骨蛋白肽-钙螯合物的多分散指数没有显著性差别(P>0.05),且包埋肉桂精油的螯合物多分散指数小于0.2,螯合物分散性较好。
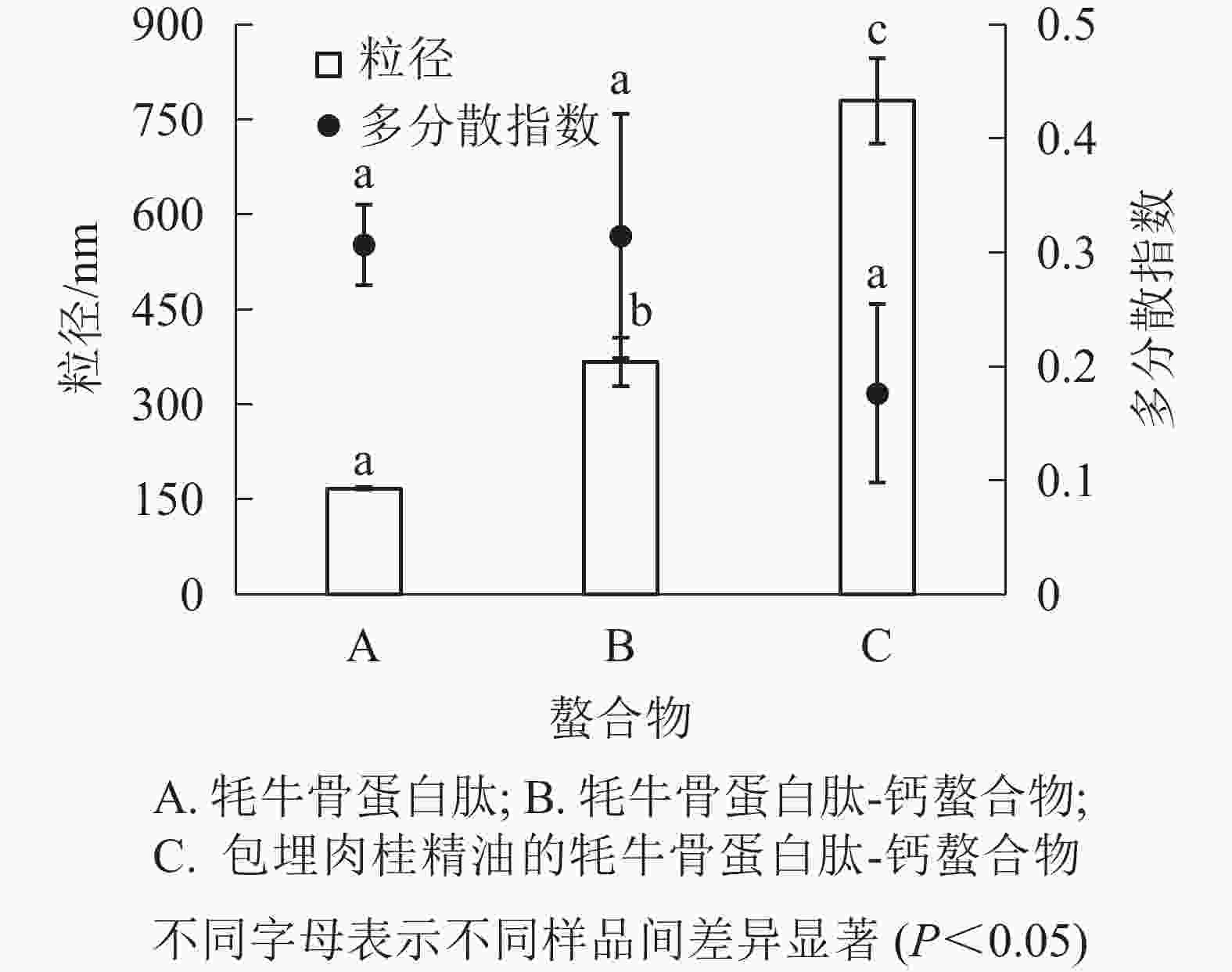
图 3 螯合物的粒径和多分散指数
Figure 3. Particle size and polydispersity coefficient of emulsion of cinnamon essential oil and yak bone peptides-calcium chelate
-
由表4可知:包埋肉桂精油的牦牛骨蛋白肽-钙螯合物的总体评分为7.22分,即该螯合物表观状态良好,分散均一,无分层,色泽一致,无杂质,肉桂香浓郁、风味纯正,气味较协调,无不良气味,其中“香气”项的分值显著高于肉桂精油稀释液(P<0.05)。
表 4 感官评价结果
Table 4. Artificial sensory evaluation results
感官特性 包埋肉桂精油的牦牛骨
蛋白肽-钙螯合物/分肉桂精油稀释液/分 感官特性 包埋肉桂精油的牦牛骨
蛋白肽-钙螯合物/分肉桂精油稀释液/分 状态 7.60±0.70 b 8.65±0.47 a 异味 7.10±0.99 b 8.00±0.67 a 色泽 8.15±0.34 a 8.25±0.59 a 协调性 6.10±0.88 b 8.15±0.34 a 香气 7.15±1.00 a 4.35±0.82 b 平均 7.22 7.48 说明:不同字母表示样品间差异显著(P<0.05) -
由图4可知:在2 h后,对照和牦牛骨蛋白肽-钙螯合物的吸光度明显上升,维氏气单胞菌大量繁殖;相比之下,牦牛骨蛋白肽-钙螯合物的吸光度值增速更快,12 h抑菌率为−13.70%,推测是牦牛骨蛋白肽为菌的繁殖提供营养。肉桂精油稀释液和包埋肉桂精油的牦牛骨蛋白肽-钙螯合物都表现出了良好的抑菌效果,12 h抑菌率分别为70.24%和77.33%。螯合物对维氏气单胞菌的6 h抑菌率为61.41%,且6 h后抑菌率始终高于肉桂精油稀释液,说明螯合物抑菌效果较为稳定。
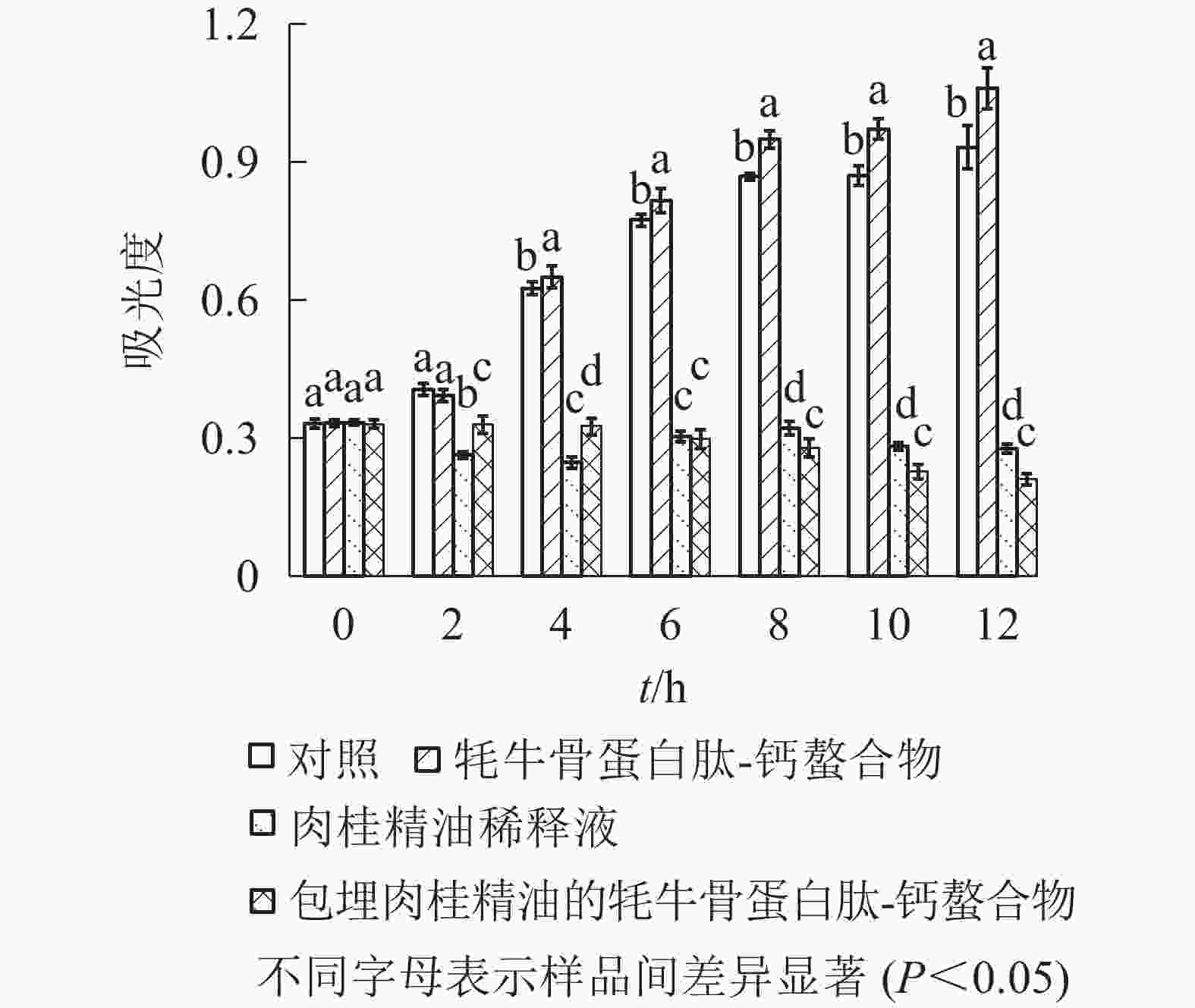
图 4 肉桂精油和螯合物对维氏气单胞菌的抑菌效果
Figure 4. Bacteriostatic effect of cinnamon essential oil and yak bone peptides-calcium chelate on Aeromonas veronii
-
人体从外界摄取的蛋白类营养物质被分解后多以小分子肽的形式被吸收利用,肽的螯合能力受分子量影响;相比之下,小分子肽内部活性基团和螯合位点暴露程度较大,与钙离子的接触机率也较大[27]。本研究中得到的多数牦牛骨蛋白肽分子量小于1.0 kDa,属于小分子肽,因此适用于钙螯合物的制备。
本研究发现:大量牦牛骨蛋白肽在pH小于7时带负电荷,有利于蛋白肽侧链与钙离子间的静电吸引,当负电荷被钙离子中和时,中性蛋白肽受到疏水作用影响,形成蛋白肽-钙螯合物。同时,牦牛骨蛋白肽中还存在大量的脯氨酸(P)和甘氨酸(G);HAINES-BUTTERICK等[28]发现:脯氨酸和甘氨酸通常依次出现在β-折叠的边缘链或β-发夹中,这种蛋白质超二级结构有助于形成球形结构[29],可被应用于药物的释放[30]。因此,牦牛骨蛋白肽可用于制备具缓释杀菌功能的钙螯合物。
浊度是评价多肽螯合钙螯合程度的重要指标之一[21],与螯合物粒径正相关;当悬浮液浊度达到原始分散体的50%且随着钙离子浓度增加不再形成沉淀物时,被认为螯合钙形成良好[19]。AKBARI等[16]发现:随着钙离子浓度增加,螯合物的粒径增加。ZHANG等[21]发现:高浓度钙离子可以平衡肽的负电荷,形成沉淀或附聚物(肽-钙螯合物)。本研究中,随着牦牛骨钙溶液浓度增加,相对浊度显著升高,且未生成沉淀物,与AKBARI等[16]结论一致;因此,使用2.0 mmol·L−1牦牛骨钙溶液可形成螯合钙,且稳定性良好。
多分散指数(PDI)可用来表征纳米乳液粒子的分布情况,PDI越小表示乳液粒子分布越均匀[31]。本研究中,通过高速剪切法向牦牛骨蛋白肽溶液中加入牦牛骨钙溶液,粒径明显增加,PDI较小,说明有新的物质形成,即生成了肽-钙螯合物,且分散性较好;继续加入肉桂精油,乳液粒径出现显著增加,推断是形成了包埋肉桂精油的牦牛骨蛋白肽-钙螯合物。
人工感官评定是最基础的食品质量评价方法[32]。本研究制备的包埋肉桂精油的牦牛骨蛋白肽-钙螯合物在各项评价中表现良好,且“香气”项分值显著高于肉桂精油稀释液,与肉桂精油稀释液相比,静置24 h后肉桂香更浓郁,认为螯合物有一定的缓释作用。
微生物是食物腐败的主要原因,抑制微生物生长是延长保质期和提高水产品质量的有效方法,气单胞菌是水产品中的优势腐败菌[33]。本研究中,包埋肉桂精油的牦牛骨蛋白肽-钙螯合物对气单胞菌表现出明显的抑菌效果,与HUANG等[7]结果相符。随时间增长,螯合物抑菌效果逐渐优于肉桂精油稀释液,说明螯合物具有缓释效果,延长了抑菌时间,有助于延长水产品的货架期。
-
本研究利用牦牛骨资源制备包埋肉桂精油的牦牛骨蛋白肽-钙螯合物;通过分析螯合物相对浊度,确定制备条件;通过测定牦牛骨蛋白肽分子量,分析牦牛骨蛋白肽序列,确认牦牛骨蛋白肽适用于螯合物的制备,并推断螯合物形成机制为牦牛骨蛋白肽的负电荷与钙离子间产生静电吸引作用,受疏水作用影响形成球形肽-钙螯合物;通过测定粒径、多分散指数和浊度,表征螯合物特性,确认螯合物的形成;通过测定抑菌率和感官评价实验,验证包埋肉桂精油的牦牛骨蛋白肽-钙螯合物抑菌效果良好,且具有缓释作用,为新型抑菌剂的开发提供思路。
Preparation and antibacterial effect of yak bone protein peptide-calcium chelate embedded with cinnamon essential oil
-
摘要:
目的 为充分利用牦牛Bos grunniens骨资源,改善其利用率低下,浪费严重的现状;研究包埋肉桂精油的牦牛骨蛋白肽-钙螯合物的形成机制及抑菌效果。 方法 牦牛骨经高温蒸煮,分离出蛋白质和骨渣。酶解蛋白质得到牦牛骨胶原蛋白肽,酸溶骨渣得到可溶性牦牛骨钙。通过肉桂精油与牦牛骨胶原蛋白肽混合均质形成肉桂精油-牦牛骨蛋白肽乳液,乳液与牦牛骨钙通过高速剪切法获得包埋肉桂精油的牦牛骨蛋白肽-钙螯合物。测定螯合钙浊度、粒径、多分散指数和抑菌率,并进行感官评价。 结果 牦牛骨蛋白肽多为小分子肽(<1.0 kDa),适合钙螯合物的制备。pH<7时多数牦牛骨蛋白肽带负电荷,受疏水作用影响,形成牦牛骨蛋白肽-钙螯合物。牦牛骨蛋白肽由大量的脯氨酸(P)和甘氨酸(G)组成,容易构成多肽二级结构β-转角,利于物质释放。包埋肉桂精油的牦牛骨蛋白肽-钙螯合物稳定性良好,平均粒径为780.2 nm;分散性良好,多分散指数小于0.2。牦牛骨蛋白肽、肉桂精油和螯合物对维氏气单胞菌Aeromonas veronii的12 h抑菌率分别为−13.70%、70.24%和77.33%,螯合物抑菌效果良好;6 h后螯合物抑菌率超过肉桂精油。感官评价结果发现:螯合物和肉桂精油的香气评分分别为7.15和4.35分,螯合物肉桂味浓郁程度显著高于肉桂精油(P<0.05),螯合物有一定缓释效果。 结论 牦牛骨蛋白肽和牦牛骨钙适用于研发复合氨基酸矿物质螯合钙产品,包埋肉桂精油的牦牛骨蛋白肽-钙螯合物可用于新型抑菌剂的开发。图4表4参33 Abstract:Objective This study aims to explore the formation mechanism and antibacterial effect of yak bone protein peptide-calcium chelate embedded with cinnamon essential oil, so as to make full use of the bone resource and improve its utilization rate and avoid serious waste. Method The yak bone was autoclaved at high temperature to separate protein and bone residue. Yak bone collagen peptide was obtained from enzymatic hydrolysis of protein, and soluble yak bone calcium was obtained from acid-dissolved bone residue. The cinnamon essential oil and the yak bone collagen peptide were mixed and homogenized to form a cinnamon essential oil-yak bone peptide emulsion, and the calcium chelate of yak bone protein peptide encapsulated in cinnamon essential oil was obtained by high speed shear method. The turbidity, particle size, polydispersity index, antibacterial activity of the chelated calcium were determined and organoleptic evaluation was conducted. Result The yak bone peptides were mostly small molecule peptides (less than 1.0 kDa), and were suitable for the preparation of chelated calcium. When pH value was less than 7, many yak bone peptides were negatively charged and were affected by hydrophobicity to form yak bone protein peptide-calcium chelates. The yak bone peptide was composed of a large number of proline (P) and glycine (G), which could easily form the β-turn, one of the polypeptide secondary structures, and facilitate the release of substances. The yak bone protein peptide-calcium chelate embedded with cinnamon essential oil had good stability, with an average particle size of 780.2 nm. The dispersibility was good and the polydispersity index was less than 0.2. The 12 h antibacterial rates of yak bone protein peptide, cinnamon essential oil and chelate against Aeromonas veronii were −13.70%, 70.24% and 77.33%, respectively. The inhibition rate of chelate to Aeromonas was higher than that of cinnamon essential oil after 6 hours. The organoleptic evaluation revealed that the aroma scores of chelate and cinnamon essential oil were 7.15 and 4.35 respectively, and the cinnamon flavor of chelate was significantly stronger than that of cinnamon essential oil (P<0.05), indicating that the chelate had a certain sustained-release effect. Conclusion Yak bone protein peptide and yak bone calcium are suitable for the development of complex amino acid mineral chelated calcium products, and the yak bone protein peptide-calcium chelate encapsulated with cinnamon essential oil can be used for the development of new antibacterial agents. [Ch, 4 fig. 4 tab. 33 ref.] -
Key words:
- yak bone /
- protein peptide /
- cinnamon essential oil /
- chelate /
- bacteriostasis in vitro
-
图 1 牦牛骨钙溶液浓度对牦牛骨蛋白肽-钙螯合物稳定性的影响
Figure 1 Stability of yak bone peptides-calcium chelate prepared by different yak bone calcium solution
图 2 不同肉桂精油添加量对包埋肉桂精油的牦牛骨蛋白肽-钙螯合物稳定性的影响
Figure 2 Stability of yak bone peptides-calcium chelate prepared by different ratio of cinnamon essential oil
图 3 螯合物的粒径和多分散指数
Figure 3 Particle size and polydispersity coefficient of emulsion of cinnamon essential oil and yak bone peptides-calcium chelate
图 4 肉桂精油和螯合物对维氏气单胞菌的抑菌效果
Figure 4 Bacteriostatic effect of cinnamon essential oil and yak bone peptides-calcium chelate on Aeromonas veronii
表 1 感官评价评分标准
Table 1. Standard of sensory evaluation
评分 状态 色泽 香气 异味 协调性 7~9 分散均一,无分层、结块、沉淀 一致,无杂质、霉变 肉桂味浓郁、纯正 气味良好,
无不良风味气味协调 4~6 分散略不均一,无分层、结块、沉淀 不均,有杂质,无霉变 肉桂味较浓郁 稍有异味 气味较协调 1~3 分散不均一,有明显分层、结块、沉淀,
有油滴出现不均,有大量杂质,有霉变 肉桂味不突出 有明显异味 气味不协调  下载: 导出CSV
下载: 导出CSV
表 2 牦牛骨蛋白肽的分子量
Table 2. Molecular weight of yak bone peptides
t/min 分子量/kDa 占比/% <15.579 ≥3.0 0 15.579~<16.363 2.0~<3.0 0.040 16.363~<17.688 1.0~<2.0 4.094 17.688~<18.508 0.5~<1.0 34.211 ≥18.508 <0.5 61.656
下载: 导出CSV
表 3 牦牛骨蛋白肽部分多肽序列鉴定结果
Table 3. Identification of partial peptides sequences of yak bone peptides
主要肽段 可信度 分子量/Da 质核比 功能 GPVGPVG 40.23 581.317 3 582.324 1 SVPGPMG 32.33 643.299 9 644.306 6 VPGPMGP 31.42 653.320 7 654.326 5 APGPVGPAG 34.80 721.375 9 722.382 3 AP(+15.99)GPVGPAG 31.27 737.370 8 738.377 4 羟基化 FP(+15.99)GADGVA 34.14 748.339 1 749.345 7 羟基化 VP(+15.99)GPMGPS 33.76 756.347 6 757.356 1 羟基化 SDGSVGPVG 36.97 773.355 5 774.362 2 SP(+15.99)GN(+0.98)IGPAG 33.70 785.355 5 786.362 5 羟基化 GAAGPTGPIG 30.08 796.407 9 797.415 6 TPGPQ(+0.98)GIAG 30.34 797.391 9 798.399 9 脱酰胺 LPQPPQE 31.34 807.412 7 808.419 7 IQ(+0.98)GPP(+15.99)GPAG 30.19 809.391 9 810.398 9 脱酰胺,羟基化 AP(+15.99)GPQ(+0.98)GFQ 31.21 817.360 6 818.366 5 羟基化,脱酰胺 GPP(+15.99)GPVGPAG 33.96 820.407 9 821.415 4 羟基化 GP(+15.99)AGPIGPVG 34.90 836.439 1 837.445 6 羟基化 GPVGPTGPVG 37.77 836.439 2 837.445 6 TP(+15.99)GPQGLLG 31.36 854.449 8 855.456 9 羟基化 GASGPP(+15.99)GFVG 31.32 860.402 8 861.412 9 羟基化 GPQ(+0.98)GPVGPVG 31.77 864.434 1 865.440 8 脱酰胺 GIQGPP(+15.99)GPAG 30.73 865.429 3 866.436 9 羟基化 AGPSGPAGPTG 36.13 867.408 6 868.415 5 GPP(+15.99)GLQ(+0.98)GVQ 33.31 868.429 0 869.436 0 羟基化,脱酰胺 GPP(+15.99)GPMGPPG 42.39 878.395 6 879.402 2 羟基化 TGPIGPP(+15.99)GPA 41.22 878.449 8 879.457 6 羟基化 GPP(+15.99)GPIGNVG 33.95 879.445 0 880.452 9 羟基化 GPP(+15.99)GPIGN(+0.98)VG 32.42 880.429 0 881.436 0 羟基化,脱酰胺 GPP(+15.99)GFP(+15.99)GAVG 34.44 886.418 5 887.424 9 羟基化 VGPAGPN(+0.98)GFA 40.33 886.418 5 887.425 0 脱酰胺 AGPP(+15.99)GPTGPAG 31.53 893.424 3 894.431 5 羟基化 AAGPP(+15.99)GPTGPA 34.87 907.439 9 908.446 5 羟基化 VGPAGPN(+0.98)GFAG 38.51 943.439 9 944.445 9 脱酰胺 APGAPGPVGPAG 30.84 946.487 2 947.493 7
下载: 导出CSV
表 4 感官评价结果
Table 4. Artificial sensory evaluation results
感官特性 包埋肉桂精油的牦牛骨
蛋白肽-钙螯合物/分肉桂精油稀释液/分 感官特性 包埋肉桂精油的牦牛骨
蛋白肽-钙螯合物/分肉桂精油稀释液/分 状态 7.60±0.70 b 8.65±0.47 a 异味 7.10±0.99 b 8.00±0.67 a 色泽 8.15±0.34 a 8.25±0.59 a 协调性 6.10±0.88 b 8.15±0.34 a 香气 7.15±1.00 a 4.35±0.82 b 平均 7.22 7.48 说明:不同字母表示样品间差异显著(P<0.05)
下载: 导出CSV
-
[1] 罗惦, 柴林荣, 常生华, 等. 我国青藏高原地区牦牛草地放牧系统管理及优化[J]. 草业科学, 2017, 34(4): 881 − 891. LUO Dian, CHAI Linrong, CHANG Shenghua, et al. Yak grazing management systems and optimization on the Qinghai-Tibet Plateau [J]. Pratacultural Sci, 2017, 34(4): 881 − 891. [2] 魏秋霞, 杨勇, 罗明, 等. 原辅材料对牦牛骨浓缩汤品质影响研究[J]. 中国调味品, 2008(7): 43 − 46. WEI Qiuxia, YANG Yong, LUO Ming, et al. Raw materials were studied on the quality influence of concentrated soup of yak bone [J]. China Condiment, 2008(7): 43 − 46. [3] 甘林火, 翁连进, 刘青, 等. 制备L-亮氨酸螯合钙的新工艺及其生物利用度的研究[J]. 食品与发酵工业, 2008, 34(3): 53 − 57. GAN Linhuo, WENG Lianjin, LIU Qing, et al. The new technology of preparation and the study of bio-availability of the complex of L-leucine calcium [J]. Food Fermentation Ind, 2008, 34(3): 53 − 57. [4] 吴雪辉, 黄永芳, 高强, 等. 肉桂精油的抗氧化作用研究[J]. 食品科技, 2007(4): 85 − 88. WU Xuehui, HUANG Yongfang, GAO Qiang, et al. Study on the antioxidant activities of cinnamon essence oil [J]. Food Sci Technol, 2007(4): 85 − 88. [5] HILBIG J, MA Qiumin, DAVIDSON P M, et al. Physical and antimicrobial properties of cinnamon bark oil co-nanoemulsified by lauric arginate and Tween 80 [J]. Int J Food Microbiol, 2016, 233: 52 − 59. [6] KIM Y G, LEE J H, KIM S I, et al. Cinnamon bark oil and its components inhibit biofilm formation and toxin production [J]. Int J Food Microbiol, 2015, 195: 30 − 39. [7] HUANG Zhan, LIU Xiaochang, JIA Shiliang, et al. The effect of essential oils on microbial composition and quality of grass carp (Ctenopharyngodon idellus) fillets during chilled storage [J]. Int J Food Microbiol, 2018, 266: 52 − 59. [8] OJAGH S M, REZAEI M, RAZAVI S H, et al. Effect of chitosan coatings enriched with cinnamon oil on the quality of refrigerated rainbow trout [J]. Food Chem, 2010, 120(1): 193 − 198. [9] 中华人民共和国国家卫生和计划生育委员会. 食品安全国家标准 食品添加剂使用标准: GB 2760−2014[S]. 北京: 中国标准出版社, 2015. [10] 魏洁琼, 余群力, 韩玲, 等. 牛骨胶原蛋白肽制备工艺优化及抗氧化活性分析[J]. 甘肃农业大学学报, 2020, 55(5): 203 − 211. WEI Jieqiong, YU Qunli, HAN Ling, et al. Optimization of preparation process and antioxidant activity analysis of bovine bone collagen peptide [J]. J Gansu Agric Univ, 2020, 55(5): 203 − 211. [11] 钱峰, 李超. 猪骨渣中可溶性钙的提取及蛋白肽螯合钙的制备[J]. 中国食品添加剂, 2014(8): 68 − 73. QIAN Feng, LI Chao. Study on extraction of soluble calcium from pig bone residue and preparation of chelated peptide calcium compound [J]. China Food Additives, 2014(8): 68 − 73. [12] 瞿瑗, 李诚, 程乐涛. 牦牛骨免疫活性肽的酶解制备研究[J]. 食品工业科技, 2016, 37(3): 271 − 274. QU Yuan, LI Cheng, CHENG Letao. Study on yak bone immune active peptide preparation by enzymatic hydrolysis [J]. Sci Technol Food Ind, 2016, 37(3): 271 − 274. [13] 刘静, 李诚, 胡迤萧, 等. 酶解法制备牦牛骨胶原多肽及其加工性能评价[J]. 食品与生物技术学报, 2018, 37(5): 547 − 554. LIU Jing, LI Cheng, HU Yixiao, et al. Preparation by hydrolysis method and functional properties of yak bone collagen polypeptides [J]. J Food Sci Biotechnol, 2018, 37(5): 547 − 554. [14] 王晓军, 吴婷, 贾伟, 等. 酸法和酶法提取牦牛骨胶原蛋白的特性分析[J]. 食品科学, 2018, 39(12): 101 − 106. WANG Xiaojun, WU Ting, JIA Wei, et al. Comparative analysis of collagen extracted from yak bones using acidic and enzymatic treatment [J]. Food Sci, 2018, 39(12): 101 − 106. [15] HONG Hui, CHAPLOT S, CHALAMAIAH M, et al. Removing cross-linked telopeptides enhances the production of low-molecular-weight collagen peptides from spent hens [J]. J Agric Food Chem, 2017, 65(34): 7491 − 7499. [16] AKBARI A, WU Jianping. Cruciferin nanoparticles: preparation, characterization and their potential application in delivery of bioactive compounds [J]. Food Hydrocolloids, 2016, 54: 107 − 118. [17] XIE Ningning, LIU Shanshan, WANG Chan, et al. Stability of casein antioxidant peptide fractions during in vitro digestion/Caco-2 cell model: characteristics of the resistant peptides [J]. Eur Food Res Technol, 2014, 239(4): 577 − 586. [18] 忻余, 徐康森. 高效液相色谱法测定尿多酸肽注射液中小分子肽的分子量[J]. 药物分析杂志, 2001, 21(3): 191 − 193. XIN Yu, XU Kangsen. HPLC determination of peptide’s molecular weight in uropolypeptide injection [J]. Chin J Pharm Anal, 2001, 21(3): 191 − 193. [19] HONG Hui, AKBARI A, WU Jianping. Small amphipathic peptides are responsible for the assembly of cruciferin nanoparticles [J]. Sci Rep, 2017, 7(1): 7819. doi: 10.1038/s41598-017-07908-z. [20] 张涛, 宋世震, 刘睿. 基于nano LC-MS/MS法的黄明胶胶原蛋白鉴定研究[J]. 南京中医药大学学报, 2020, 36(1): 105 − 109. ZHANG Tao, SONG Shizhen, LIU Rui. Investigation on identifying collagen from Colla Corii Bovis using nano LC-MS/MS method [J]. J Nanjing Univ Tradit Chin Med, 2020, 36(1): 105 − 109. [21] ZHANG Jing, LIANG Li, TIAN Z, et al. Preparation and in vitro evaluation of calcium-induced soy protein isolate nanoparticles and their formation mechanism study [J]. Food Chem, 2012, 133(2): 390 − 399. [22] LUNA-VALDEZ J G, BALANDRÁN-QUINTANA R R, AZAMAR-BARRIOS J A, et al. Structural and physicochemical characterization of nanoparticles synthesized from an aqueous extract of wheat bran by a cold-set gelation/desolvation approach [J]. Food Hydrocolloids, 2017, 62: 165 − 173. [23] 朱红梅, 田建文, 张彦军, 等. 菠萝蜜种子淀粉制备的香草兰精油微胶囊的风味品质分析[J]. 食品工业科技, 2017, 38(21): 253 − 258. ZHU Hongmei, TIAN Jianwen, ZHANG Yanjun, et al. Flavor and quality evaluation of vanilla essential oil microcapsules made from jackfruit seed starch [J]. Sci Technol Food Ind, 2017, 38(21): 253 − 258. [24] 周子雄, 黄庆华, 朱爽, 等. 酶标仪快速测定抗菌物质抑菌活性方法的建立[J]. 微生物前沿, 2014, 3(2): 29 − 35. ZHOU Zixiong, HUANG Qinghua, ZHU Shuang, et al. Establishment of rapid determination method for antibacterial activity by microplate reader [J]. Adv Microbiol, 2014, 3(2): 29 − 35. [25] 刘富康, 方士元, 曲映红. 大青叶粗黄酮提取及其抑菌性研究[J]. 山东化工, 2018, 47(21): 34 − 35. LIU Fukang, FANG Shiyuan, QU Yinghong. Study on extraction and antibacterial activity of crude flavonoids from folium Isatidis [J]. Shandong Chem Ind, 2018, 47(21): 34 − 35. [26] 王步江, 刘金福, 樊秀花, 等. 肉桂精油抑菌活性研究[J]. 食品与机械, 2011, 27(6): 166 − 167. WANG Bujiang, LIU Jinfu, FAN Xiuhua, et al. Antimicrobial activity of cinnamon essential oil [J]. Food Mach, 2011, 27(6): 166 − 167. [27] 刘海军, 乐超银, 邵伟, 等. 生物活性肽研究进展[J]. 中国酿造, 2010(5): 5 − 8. LIU Haijun, YUE Chaoyin, SHAO Wei, et al. Research progress of bioactive peptides [J]. China Brewing, 2010(5): 5 − 8. [28] HAINES-BUTTERICK L, FAJAGOAL K, BRANCO M, et al. Controlling hydrogelation kinetics by peptide design for three-dimensional encapsulation and injectable delivery of cells [J]. Proc Natl Acad Sci USA, 2007, 104(19): 7791 − 7796. [29] FISCHER S, LAMPING M, GOLD M, et al. Synthesis of a biological active β-hairpin peptide by addition of two structural motifs [J]. Bioorg Med Chem, 2017, 25(2): 603 − 608. [30] STOTZ C E, TOPP E M. Applications of model beta-hairpin peptides [J]. J Pharm Sci, 2004, 93(12): 2881 − 2891. [31] 黄剑钊, 黎攀, 许锦伟, 等. 美藤果油纳米乳液的制备及稳定性研究[J]. 食品与发酵工业, 2020, 46(11): 216 − 222. HUANG Jianzhao, LI Pan, XU Jinwei, et al. Study on the preparation and stability of sacha inchi oil nanoemulsion [J]. Food Ferment Ind, 2020, 46(11): 216 − 222. [32] 芮汉明, 郭凯. 食品香气的综合评价技术[J]. 食品工业科技, 2008, 29(7): 277 − 280. RUI Hanming, GUO Kai. Technology of integrated evaluation of food aroma [J]. Sci Technol Food Ind, 2008, 29(7): 277 − 280. [33] 励建荣, 杨兵, 李婷婷. 水产品优势腐败菌及其群体感应系统研究进展[J]. 食品科学, 2015, 36(19): 255 − 259. LI Jianrong, YANG Bing, LI Tingting. Advances in quorum sensing of dominant spoilage bacteria from aquatic products [J]. Food Sci, 2015, 36(19): 255 − 259. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200503
 点击查看大图
点击查看大图
计量
- 文章访问数: 2075
- HTML全文浏览量: 496
- PDF下载量: 35
- 被引次数: 0
