





-
耕地破碎化是影响粮食生产效率的重要因素之一。在同等要素投入的条件下,耕地破碎化程度越高粮食生产效率越低。耕地破碎化是影响中国农业生产效率和耕地质量的重要因素[1-3],研究其时空分布以及变化趋势对于地区农业发展具有重要的实践意义,同时也有利于生态可持续发展[4]。耕地破碎化宏观上表现为耕地的连片度和连通性下降,即单一、均质和集中连片的耕地变为复杂、异质和分割成块的斑块状耕地,具体则包括耕地地块的面积、距离以及分布的景观变化。随着经济不断发展,城市化程度不断提高,一方面耕地抛荒情况普遍存在,另一方面由于道路等基础设施建设的需要,耕地破碎化程度也不断上升。国内外对耕地破碎化的驱动因素、测度方法以及耕地破碎化治理政策等各方面展开了研究。目前,耕地破碎化的研究已形成了较为成熟的指标体系,主要以核密度[5]和景观指数测算耕地的破碎化[6-7],耕地破碎化的指标主要采用遥感数据计算景观指数判断耕地斑块的数量和面积变化[8-9],在此基础上使用熵权法或变异系数法计算一个地区的耕地破碎化综合指数[10]。而景观指数的计算主要以斑块数量、斑块面积和斑块形状为特征指标[11]。在耕地破碎化驱动因素挖掘方面,基础设施的建设、地形地貌的限制以及人多地少的国情使得耕地破碎化愈发严重[5, 9-10]。但已有研究大多聚焦于城市化背景下耕地破碎化的程度,而在城市化背景下,政府也在不断干预耕地保护,如国家耕地保护政策的创新和试点,政府的干预对耕地的破碎化是否有影响?是否会延缓城市化背景下耕地破碎化的趋势?这都有待探讨。长期以来,耕地破碎化是制约浙江省嘉兴市现代农业发展和乡村空间资源高效配置的重要因素,2012年以来嘉兴市大规模开展了农村土地综合整治。研究其耕地破碎化的程度,并对比在大规模推进农村土地综合整治前后的耕地破碎化程度差异,有助于观察在城乡融合发展背景下,耕地破碎化程度在政府干预下的变化。这不仅在理论上有利于完善农村土地综合整治模式和机制,在实践中也有利于其他城市化发展较快的平原地区分析政府干预对耕地破碎化的影响。
-
浙江省嘉兴市位于长江三角洲杭嘉湖平原,如表1所示:嘉兴市下辖2个市辖区(南湖区、秀洲区)、3个县级市(海宁市、平湖市、桐乡市)、2个县(嘉善县、海盐县)。2019年嘉兴市国内生产总值(GDP)为5 370.32亿元,常住人口为480.00 万人,国土面积为4 230 km2。嘉兴市耕地资源基础条件优良,耕地质量等级全省最高,但人均耕地面积较少,耕地面积不断减少,破碎化问题突出。2018年嘉兴市耕地面积为204 897 hm2,相较上年减少了458 hm2,人均耕地面积为427 m2·人−1,低于国际粮农组织公布的人均耕地面积标准533 m2·人−1。另一方面嘉兴市耕地保有量任务重,后备资源接近零,永久基本农田保护率全市平均超过85%,其中平湖市和嘉善县均超过90%。经济增长引致的用地需求与耕地保护矛盾突出。2012年以来,嘉兴市全面实施农村土地综合整治工程发展战略,大力推进耕地连片建设,全市每年复垦导致耕地碎片化的零星建设用地面积约700 hm2。增加耕地的同时,零星分布的耕地图斑明显减少,耕地破碎度情况总体有所好转,耕地利用效益和粮食产量稳步提升。
表 1 嘉兴市区域概况
Table 1. General context of Jiaxing City
市(区、县) 2019年土地面积/km2 2019年常住人口/万人 2019年GDP/亿元 2017年耕地面积/hm2 2018年耕地面积/hm2 嘉兴市 4 230 480.00 5 370.32 205 355 204 897 南湖区 439 68.29 765.37 22 379 22 142 秀洲区 548 62.90 676.35 29 089 28 961 嘉善县 507 59.20 626.81 26 618 26 571 海盐县 590 45.10 539.65 26 019 25 945 海宁市 863 87.83 1 026.57 32 981 33 117 平湖市 557 70.35 765.77 29 321 29 236 桐乡市 727 86.33 968.17 38 947 38 925 说明:数据来源于《嘉兴市2020年统计年鉴》;GDP为国内生产总值 -
本研究数据包括矢量数据以及统计数据两大类。首先本研究收集了嘉兴市2009−2018年(其中,2015年矢量数据缺失)的耕地矢量数据,为“第2次全国土地调查”及年度变更数据,提取了其中的耕地状况数据,包括耕地面积、耕地块数、耕地周长等信息;其次,土地利用数据以及社会经济数据来源于《中国国土资源年鉴》《中国国土资源统计年鉴》《浙江省统计年鉴》和《嘉兴市统计年鉴》等。
-
已有耕地破碎化的研究大多采用景观指数描述耕地破碎化的程度[1,6,10,12 − 13],一方面是由于耕地作为特殊的景观,其空间布局、分布和形态是反映其破碎化程度的具体指标。另一方面,景观指数所包含的数据信息量大且容易获取。参考已有研究[1,12-13]并结合嘉兴市的实际情况,用以下6个指标对耕地破碎化进行描述(表2)。
表 2 耕地破碎化指标体系
Table 2. Index system of farmland fragmentation
指标名称 公式 说明 单位耕地面积(UFA) AUF=AT/QT AUF为单块耕地面积,AT为区域内的耕地总面积,QT为区域内耕地的 总数量。单位耕地面积越大说明耕地的破碎化程度越低,区域内 的耕地越趋向规模经营 耕地密度(FD) DF=QT/AT DF为耕地密度,AT为区域内的耕地总面积,QT为区域内耕地的总数 量。耕地密度是指单位面积的耕地上拥有的耕地数量,耕地密度越大 说明耕地破碎化程度越高,耕地分割越严重 边界密度指数(ED) DE=PT/AT DE为边界密度指数,PT为区域内耕地斑块总周长,AT为区域内耕地总 面积。边界密度指数以单位面积内的耕地地块周长为表征,边界 密度越大说明区域内耕地被划分为更多地块,区域的耕地破碎化程 度越高 区域内地块形状指数(TSI) ITS=${\displaystyle\sum\limits_{i = 1}^{ {Q_{\rm{T}}} } {\left( {\dfrac{ {0.25{P_i} } }{ {\sqrt { {A_i} } } } } \right)} ({A_i}/{A_{\rm{T}}})}$ ITS为区域内地块形状指数,QT是区域内耕地总数量,Pi是区域内i地块周 长,Ai是第i地块面积,AT是耕地总面积。区域内地块形状指数 根据地块周长小的地块形状更规则的原理得到,形状规则的地块 更有利于规模经营。区域内地块形状指数越大,破碎化程度越高 地块数量破碎化指数(FIQ) IFQ=(NT−1)ATmin/AT IFQ为地块数量破碎化指数,NT是区域内耕地斑块总数,ATmin是区域 内最小的耕地斑块的面积,AT是区域内耕地斑块总面积。该指数的数 值为0~1,最小地块的面积越小、区域内耕地地块的数量越少,耕地 的破碎化程度越低。相反则耕地的破碎化程度越高 地块形状破碎化指数(FIS) IFS=1−1/IPS
IPS=${\displaystyle\sum\limits_{i = 1}^{ {Q_{\rm{T} } } } {\left( {\dfrac{ {0.25{P_i} } }{ {\sqrt { {A_i} } } } } \right)} /{Q_{\rm{T}}} }$IFS为地块形状破碎化指数,QT是区域内耕地总数量,Pi是区域内i地 块周长,Ai是第i地块面积,IPS是区域内地块形状指数的平均值。 区域内耕地地块周长越长,形状越不规则,地块的形状破碎化指数 越高 -
为进一步客观描述2009−2018年(2015年除外)嘉兴市的耕地破碎化情况,对耕地破碎化的各指标赋权后综合计算嘉兴市每年的耕地破碎化综合指数。熵值法可以根据各指标的离散程度,客观判断各指标对总指标的贡献度,减小人工赋权的主观性。指标的信息量越小,离散程度越大,熵值越大,则对总指标的评价影响越大。差异性越大的指标对总指标的评价越有影响。本研究拟采用熵值法测算6个指标对破碎化评价的重要性,并据此计算6个指标的权重。最后根据6个指标的权重,加权计算嘉兴市每年的耕地破碎化综合指数。可通过SPSSAU的功能来实现,具体步骤如下。
①在进行熵值法前,为规避6个指标的单位不同以及正负向不同的问题,先对6个指标进行标准化处理。在以上6个指标中,单块耕地面积越小,耕地破碎化程度越高,而耕地密度、边界密度指数、区域内地块形状指数、地块数量破碎化指数和地块形状破碎化指数均为数值越大,耕地破碎化程度越高。因此,对单位耕地面积进行逆向化处理,Pij=(Ximax−Xij)/(Ximax−Ximin);而其他5个指标进行正向化处理,Pij=(Xij−Ximin)/(Ximax−Ximin)。其中,Xij是j县(市、区)i指标的值,Ximin是指标i的最小值,Ximax是指标i的最大值,Pij是j县(市、区)i指标标准化后的值。②测算每个指标的熵值:
${e_i} = - (1/{\rm{ln}}m)\displaystyle\sum\limits_{j = 1}^m {{P_{ij}}} {\rm{ln}}{P_{ij}}$ 。其中,ei是指标i的熵值,m是县的个数,Pij是j县(市、区)指标i标准化后的结果。③计算指标的权重:${W_i} = (1 - {e_i})\Big/\displaystyle\sum\limits_{i = 1}^n {(1 - {e_i})}$ 。其中,$ {e}_{i} $ 是i指标的熵值,n是指标的个数,Wi是指标i的权重。④综合测算各县(市、区)的耕地破碎化综合指数:${F_{i\!j}} = \displaystyle\sum\limits_{i = 1}^n {{P_{i\!j}}} {W_i}$ 。其中,Fij为耕地破碎化综合指数,Pij是j县(市、区)指标i标准化后的结果,Wi是指标i的权重。 -
为从整体上分析嘉兴市耕地破碎化的情况,运用平均断点法将嘉兴市的耕地破碎化综合指数分为3级,一级为(0.60,0.67],二级为(0.67,0.74],三级为(0.74,0.81]。由图1可见:整体上各县(市、区)的耕地破碎度有明显的波动,2009−2012年呈现上升趋势,2013−2018年逐渐下降。分别就3个时间节点的各县(市、区)耕地破碎化的分级情况来看,2009年海盐县和海宁市为一级,其他县为二级;2013年海盐县、海宁市和嘉善县为二级,其他均为三级;直至2018年海宁市和嘉善县为一级,其他县为二级。具体地,位于嘉兴市中心位置的南湖区,耕地破碎化综合指数自2009年的0.679上升至2013年0.778后逐渐下降至2018年的0.699,2009−2013年的年均增长率约3.4%,2013−2018年年均增长率约−2.6%。从耕地的数量和面积来看,南湖区的耕地面积由2009年的24 925 hm2降低至2013年的24 404 hm2,而耕地数量则增加了3 762块;同时,南湖区的GDP由2009年257.56亿元增长至2013年的429.94亿元。究其根源,南湖区的耕地破碎化的加大是城市快速发展背后的土地需求,推动了耕地的面积减小和碎片化[14]。秀洲区2009−2013年耕地破碎化一直呈现增长的趋势,2014年后耕地破碎化减缓。嘉善县、海宁市和桐乡市的耕地破碎化综合指数从2009−2012年呈现波动式上涨后,2013−2018年波动下降。海盐县和平湖市耕地破碎化综合指数在2013年达到最大值后逐渐减小。
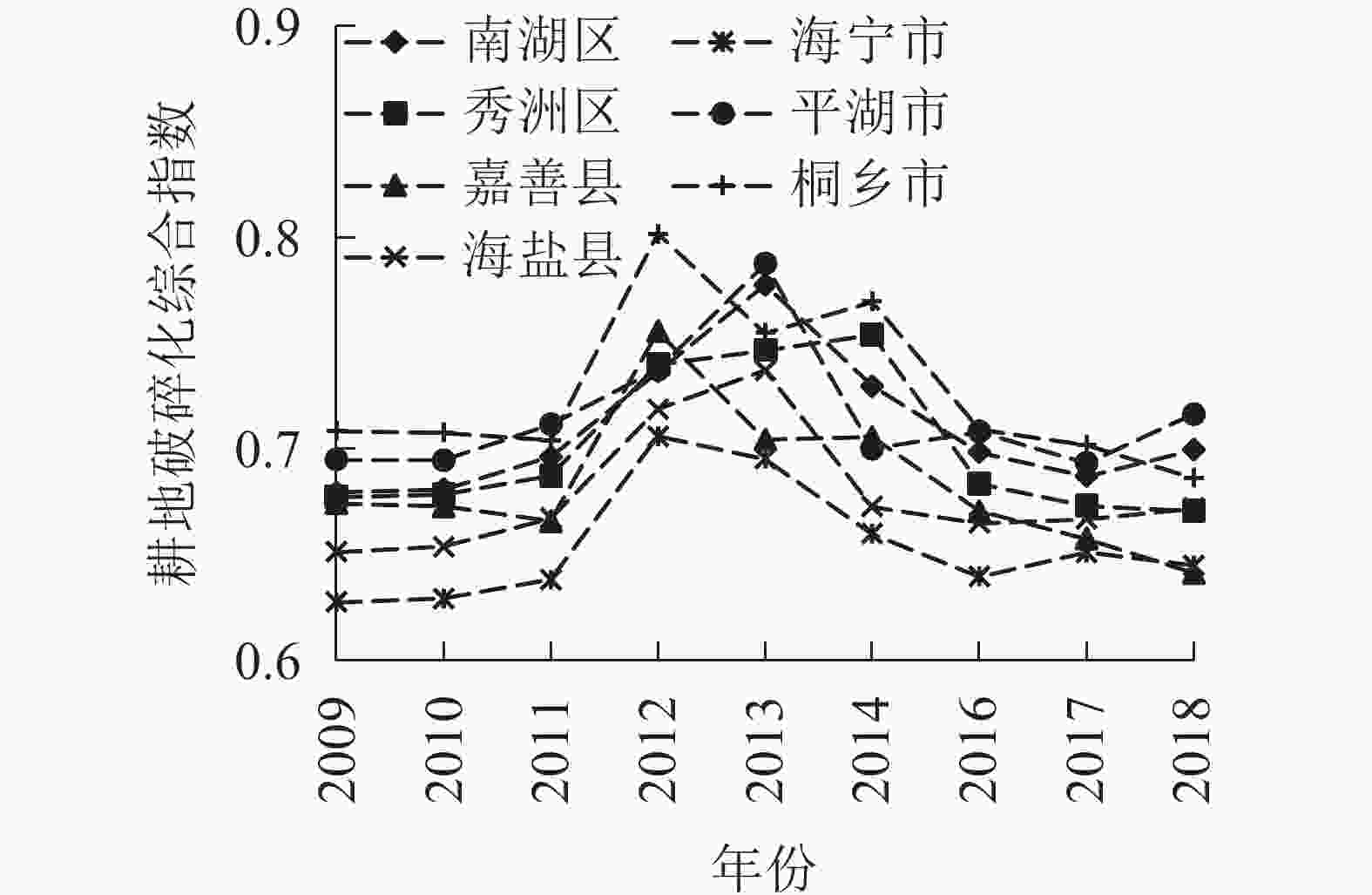
图 1 嘉兴市各县(市、区)耕地破碎化综合指数
Figure 1. Farmland fragmentation comprehensive index of different counties in Jiaxing
-
从嘉兴市各县(市、区)耕地破碎化的空间差异(图2)可以看出:嘉兴市全域耕地破碎化程度呈先上升后下降的趋势。具体来看,2009年,嘉兴市耕地破碎化只有一级区和二级区,因此总体上耕地破碎化程度较轻;相对严重的二级区主要集中在嘉兴市北部,南部的海宁市和海盐县在2009年耕地破碎化程度较低,为耕地破碎化一级区。2013年,嘉兴市中部地区(包括桐乡市、秀洲区、南湖区和平湖市)耕地破碎化程度急剧上升,由二级区变为了三级区;同时南部的海宁市和海盐县也有所恶化,耕地破碎化程度由一级变为了二级。到2018年,中部4个地区(桐乡市、秀洲区、南湖区和平湖市)的耕地破碎化程度有所减缓,变为了二级区,同时北部的嘉善县和南部的海宁市,耕地破碎化程度也有所下降。因此,嘉兴市耕地破碎化情况整体上呈现由中部向外递减的趋势。
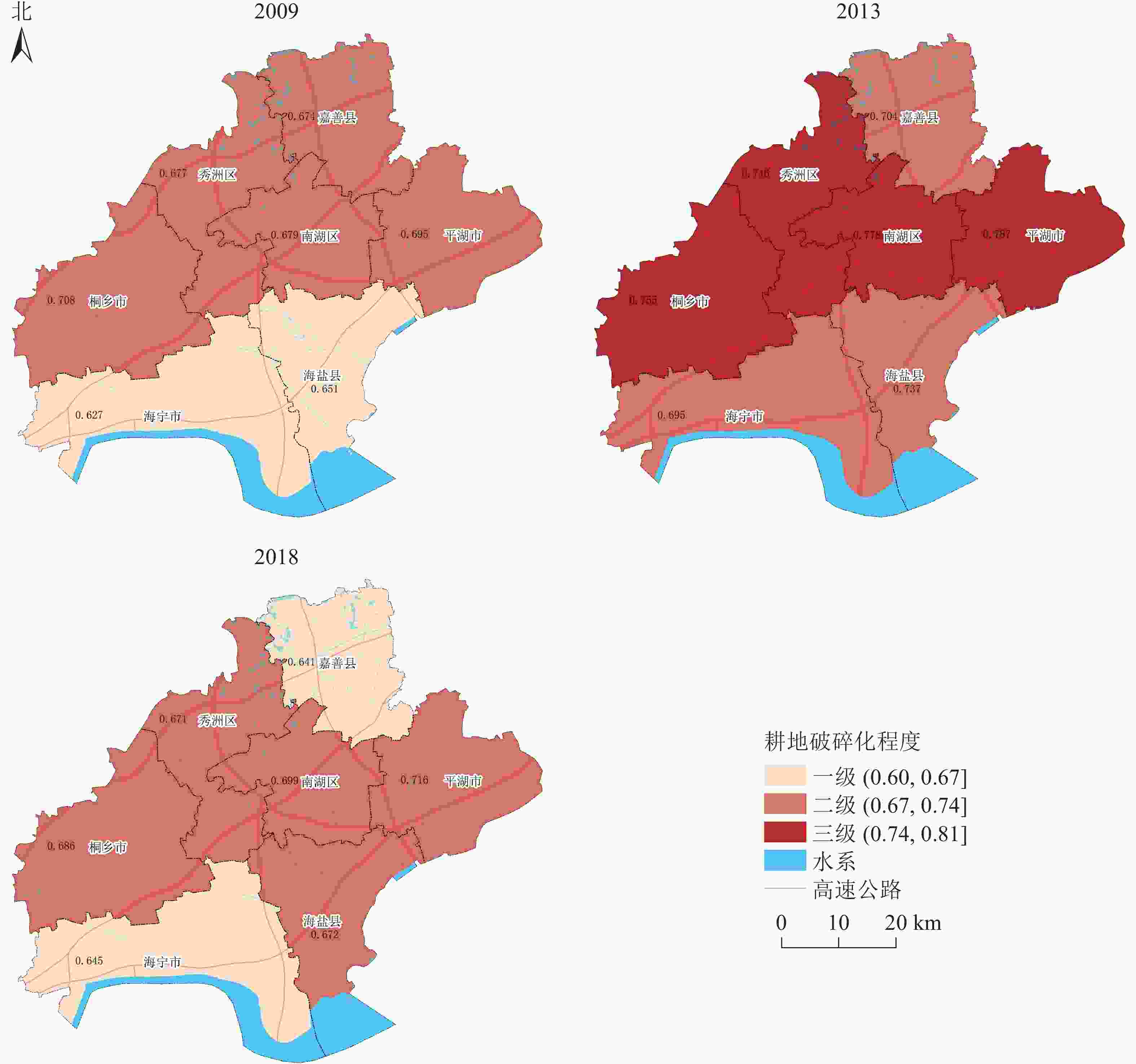
图 2 嘉兴市耕地破碎化时空演变示意图
Figure 2. Jiaxing’s farmland fragmentation and spatio-temporal change
-
2009年,嘉兴市中部的南湖区、桐乡市和平湖市耕地破碎化程度较高,2009−2018年耕地破碎化综合指数平均值分别为0.709、0.727、0.716;秀洲区次之,平均值为0.701;嘉善县、海盐县和海宁市相对较好,平均值分别为0.683、0.678、0.654。2009−2013年,耕地破碎化程度总体加大,各县(市、区)的耕地破碎情况差异也逐渐增加,2009年各县(市、区)耕地破碎化综合指数的变异系数为0.040,且逐渐增加,至2014年为0.057。说明随着耕地破碎化程度的加剧,各县(市、区)的耕地破碎化情况差异逐渐显现。主要表现为南湖区、桐乡市和平湖市耕地破碎化情况更为严重,而相对较好的嘉善县、海盐县和海宁市,耕地进一步破碎的进程较缓。不难推测,由于产业发展的路径依赖,原本就是嘉兴市二、三产业重点发展地区的南湖区、桐乡市和平湖市随着经济的发展耕地破碎化情况不断恶化。
2012年,嘉兴市开始大规模推进农村土地综合整治,以此为平台和抓手,破解制约嘉兴城乡统筹发展的障碍,提出“耕地占补平衡有良方,土地节约集约有增量,农民安居乐业有保障,城乡统筹发展有希望”的发展目标,推动了嘉兴市经济社会可持续发展,为全省乃至全国树立了耕地保护的样板。大规模推进农村土地综合整治与耕地破碎化程度的减轻有着密切的联系。2013−2018年,全市耕地破碎化程度整体减缓,耕地破碎化情况严重的南湖区、桐乡市和平湖市耕地破碎化得到了明显改善,耕地破碎化的空间差异也逐渐减小,各县(市、区)耕地破碎化综合指数的变异系数降低至0.041。
从时间上可以看出,嘉兴市各县(市、区)耕地破碎化程度达到顶峰的时间在2009−2013年,此后,各县(市、区)的耕地破碎化情况均有所好转。而从空间上来看,2009−2013年耕地破碎化出现从中心向周边扩展的趋势,临近主城区南湖区的周边县(市、区)耕地破碎化较严重,相对较远的嘉善县和海宁市耕地破碎化加剧的程度较轻。2013年后,耕地破碎化情况整体上出现好转,这与大规模推进农村土地综合整治密不可分。可见,大规模推进农村土地综合整治是2013年后耕地破碎化程度整体降低的重要影响因素。
-
本研究根据嘉兴市耕地矢量数据,运用熵值法计算嘉兴市各县(市、区)耕地破碎化综合指数后得出:①嘉兴市各县(市、区)地耕地破碎化在2009−2013年整体上呈显著上升趋势,2013−2018年嘉兴市耕地破碎化程度有所减缓,呈现下降趋势。②空间上,嘉兴市耕地破碎化呈现从中部向南北逐渐减少的趋势,2009−2013年耕地破碎化严重的县(市、区)耕地破碎化综合指数增长更快,各县(市、区)的耕地破碎化空间差异加大;2013年后耕地破碎化较严重的地区,耕地破碎化程度明显降低,耕地破碎化空间差异减小。③分析嘉兴市耕地破碎化变化的驱动因素可以发现,2009−2013年经济发展带来的建设用地占用耕地以及农民无序建房等行为是促使耕地破碎化加重的主要原因[15],而在2013年嘉兴市大规模推进农村土地综合整治后,耕地破碎化程度有效减小。嘉兴市的耕地破碎化演变对于全国的耕地保护也具有一定借鉴意义,一方面各地应警惕城市扩张、经济发展带来的耕地破碎等影响耕地质量的因素[16-17],积极推进农村土地综合整治,创新完善整治模式,注重耕地的连片性建设,以有效保护耕地质量,促进农业现代化发展[18],提高耕地的连片度和耕种效率;另一方面,应创新完善耕地保护考核机制[19],禁止任意占用导致的耕地破碎化[20],将耕地的破碎化治理及耕地连片建设纳入耕地保护考核目标内容,以制度促进耕地破碎化的降低[21]。
Spatio-temporal variation of farmland fragmentation in water net plain area
-
摘要:
目的 耕地破碎化是影响中国农业生产效率和规模化经营的重要因素,研究其时空分布以及变化趋势对于地区的农业发展、提高耕地保护水平具有重要的实践意义。 方法 以浙江省嘉兴市为研究对象,计算了2009−2018年嘉兴市7个县级单位的耕地破碎化综合指数,从时间和空间两方面分析各县(市、区)耕地破碎化变化趋势和时空演变特征,并就其变化的驱动因素进行了理论分析。 结果 空间上,嘉兴市耕地破碎化呈现由中部向南北逐渐递减的趋势。在时间上,2009年各县(市、区)耕地破碎度综合指数的变异系数为0.040,变异系数逐渐增加至2014年的0.057。2012年嘉兴市开始大规模推进农村土地综合整治,改变了各县(市、区)耕地破碎化空间差异,2018年各县(市、区)耕地破碎化综合指数的变异系数下降至0.041,同时也减小了各县(市、区)耕地破碎化的空间差异。 结论 需进一步推进农村土地综合整治,创新完善整治模式,并将耕地的破碎化治理及耕地连片建设纳入耕地保护考核目标内容,以制度降低耕地破碎化程度。图2表2参21 Abstract:Objective To promote regional agricultural development, this study is aimed to conduct an investigation of the temporal and spatial distribution and change trend of farmland fragmentation, an important factor that affects the agricultural production efficiency and large-scale management in China. Method With Jiaxing City of Zhejiang Province selected as the research area where the comprehensive index of farmland fragmentation of 7 county-level units in Jiaxing City from 2009 to 2018 was calculated, this paper makes an analysis of the change trend and spatio-temporal evolution characteristics of farmland fragmentation in different counties as well as a theoretical analysis of the driving factors. Result Spatially, the fragmentation of farmland in Jiaxing City has deomonstrated a gradual decrease from the middle to the North and South. In terms of time, the coefficient of variation of the comprehensive index of farmland fragmentation was 0.040 in 2009 and gradually increased to 0.057 in 2014. In 2012, Jiaxing promoted the comprehensive rural land consolidation on a large scale, which changed the trend increasing farmland fragmentation and reduced the spatial difference of farmland fragmentaion in each county, the coefficient of variation of the comprehensive index of farmland fragmentation decreased to 0.041 by 2018. Conclusion It is necessary to further promote the comprehensive consolidation of rural land, innovate and improve the renovation mode and include the control of fragmentation and the promotion of connectivity of farmland as part of farmland protection assessment so as to reduce the fragmentation of farmland. [Ch, 2 fig. 2 tab. 21 ref.] -
图 1 嘉兴市各县(市、区)耕地破碎化综合指数
Figure 1 Farmland fragmentation comprehensive index of different counties in Jiaxing
图 2 嘉兴市耕地破碎化时空演变示意图
Figure 2 Jiaxing’s farmland fragmentation and spatio-temporal change
表 1 嘉兴市区域概况
Table 1. General context of Jiaxing City
市(区、县) 2019年土地面积/km2 2019年常住人口/万人 2019年GDP/亿元 2017年耕地面积/hm2 2018年耕地面积/hm2 嘉兴市 4 230 480.00 5 370.32 205 355 204 897 南湖区 439 68.29 765.37 22 379 22 142 秀洲区 548 62.90 676.35 29 089 28 961 嘉善县 507 59.20 626.81 26 618 26 571 海盐县 590 45.10 539.65 26 019 25 945 海宁市 863 87.83 1 026.57 32 981 33 117 平湖市 557 70.35 765.77 29 321 29 236 桐乡市 727 86.33 968.17 38 947 38 925 说明:数据来源于《嘉兴市2020年统计年鉴》;GDP为国内生产总值  下载: 导出CSV
下载: 导出CSV
表 2 耕地破碎化指标体系
Table 2. Index system of farmland fragmentation
指标名称 公式 说明 单位耕地面积(UFA) AUF=AT/QT AUF为单块耕地面积,AT为区域内的耕地总面积,QT为区域内耕地的 总数量。单位耕地面积越大说明耕地的破碎化程度越低,区域内 的耕地越趋向规模经营 耕地密度(FD) DF=QT/AT DF为耕地密度,AT为区域内的耕地总面积,QT为区域内耕地的总数 量。耕地密度是指单位面积的耕地上拥有的耕地数量,耕地密度越大 说明耕地破碎化程度越高,耕地分割越严重 边界密度指数(ED) DE=PT/AT DE为边界密度指数,PT为区域内耕地斑块总周长,AT为区域内耕地总 面积。边界密度指数以单位面积内的耕地地块周长为表征,边界 密度越大说明区域内耕地被划分为更多地块,区域的耕地破碎化程 度越高 区域内地块形状指数(TSI) ITS=${\displaystyle\sum\limits_{i = 1}^{ {Q_{\rm{T}}} } {\left( {\dfrac{ {0.25{P_i} } }{ {\sqrt { {A_i} } } } } \right)} ({A_i}/{A_{\rm{T}}})}$ ITS为区域内地块形状指数,QT是区域内耕地总数量,Pi是区域内i地块周 长,Ai是第i地块面积,AT是耕地总面积。区域内地块形状指数 根据地块周长小的地块形状更规则的原理得到,形状规则的地块 更有利于规模经营。区域内地块形状指数越大,破碎化程度越高 地块数量破碎化指数(FIQ) IFQ=(NT−1)ATmin/AT IFQ为地块数量破碎化指数,NT是区域内耕地斑块总数,ATmin是区域 内最小的耕地斑块的面积,AT是区域内耕地斑块总面积。该指数的数 值为0~1,最小地块的面积越小、区域内耕地地块的数量越少,耕地 的破碎化程度越低。相反则耕地的破碎化程度越高 地块形状破碎化指数(FIS) IFS=1−1/IPS
IPS=${\displaystyle\sum\limits_{i = 1}^{ {Q_{\rm{T} } } } {\left( {\dfrac{ {0.25{P_i} } }{ {\sqrt { {A_i} } } } } \right)} /{Q_{\rm{T}}} }$IFS为地块形状破碎化指数,QT是区域内耕地总数量,Pi是区域内i地 块周长,Ai是第i地块面积,IPS是区域内地块形状指数的平均值。 区域内耕地地块周长越长,形状越不规则,地块的形状破碎化指数 越高
下载: 导出CSV
-
[1] 李鑫, 欧名豪, 马贤磊. 基于景观指数的细碎化对耕地利用效率影响研究:以扬州市里下河区域为例[J]. 自然资源学报, 2011, 26(10): 1758 − 1767. LI Xin, OU Minghao, MA Xianlei. Analysis on impact of fragmentation based on landscape index to cultivated land use efficiency: a case on Lixiahe District in Yangzhou City [J]. J Nat Resour, 2011, 26(10): 1758 − 1767. [2] TURNER B L, LAMBIN E F, REENBERG A. The emergence of land change science for global environmental change and sustainability [J]. Proc Nat Acad Sci, 2008, 104(52): 20666 − 20671. [3] SU Shiliang, HU Yi’na, LUO Fanghan, et al. Farmland fragmentation due to anthropogenic activity in rapidly developing region [J]. Agric Syst, 2014, 131: 87 − 93. [4] SHRESTHA M K, YORK A M, BOONE C G, et al. Land fragmentation due to rapid urbanization in the phoenix metropolitan area: analyzing the spatiotemporal patterns and drivers [J]. Appl Geogr, 2012, 32(2): 522 − 531. [5] 李鹏山, 吕雅慧, 张超, 等. 基于核密度估计的京津冀地区耕地破碎化分析[J]. 农业机械学报, 2016, 47(5): 281 − 287. LI Pengshan, LÜ Yahui, ZHANG Chao, et al. Analysis of cultivated land fragmentation in Beijing-Tianjin-Hebei region based on kernel density estimation [J]. Trans Chin Soc Agric Mach, 2016, 47(5): 281 − 287. [6] IRWIN E, BOCKSTAEL N E. The evolution of urban sprawl: evidence of spatial heterogeneity and increasing land fragmentation [J]. Proc Nat Acad Sci, 2007, 104(52): 20672 − 20677. [7] DEMETRIOU D, SEE L, STILLWELL J. A parcel shape index for use in land consolidation planning [J]. Trans GIS, 2013, 17(6): 861 − 882. [8] KOSROW D. Density Estimation for Statistics and Data Analysis[M]. London: Chapman and Hall, 1986. [9] 邓楚雄, 刘睿娜, 聂小东, 等. 土地利用变化影响下湘西自治州景观格局的时空演变[J]. 四川师范大学学报(自然科学版), 2020, 43(3): 383 − 392. DENG Chuxiong, LIU Ruina, NIE Xiaodong, et al. The temporal and spatial evolution of landscape pattern under the effects of land use change in Xiangxi Autonomous Prefecture [J]. J Sichuan Norm Univ Nat Sci, 2020, 43(3): 383 − 392. [10] 崔永峰, 赵培强. 连城国家级自然保护区景观格局动态变化分析[J]. 中国水土保持, 2020(5): 44 − 47. CUI Yongfeng, ZHAO Peiqiang. Dynamic changes of landscape pattern in Liancheng National Nature Reserve [J]. Soil Water Conserv China, 2020(5): 44 − 47. [11] 张显源, 蔡忠亮, 李桂娥, 等. 耕地景观破碎化成因及对农村收入影响分析[J]. 测绘科学, 2020, 45(4): 134 − 141. ZHANG Xianyuan, CAI Zhongliang, LI Gui’e, et al. Assessment of determinants of cultivated land fragmentation and its impacts on rural income [J]. Sci Surv Mapp, 2020, 45(4): 134 − 141. [12] 由畅, 周永斌, 于丽芬. 景观破碎化数量分析方法概述[J]. 中国农学通报, 2006, 22(5): 146 − 151. YOU Chang, ZHOU Yongbin, YU Lifen. An introduction of quantitative methods in landscape pattern fragmentation [J]. Chin Agric Sci Bull, 2006, 22(5): 146 − 151. [13] 陈帷胜, 冯秀丽, 马仁锋, 等. 耕地破碎度评价方法与实证研究:以浙江省宁波市为例[J]. 中国土地科学, 2016, 30(5): 80 − 87. CHEN Weisheng, FENG Xiuli, MA Renfeng, et al. Method of cultivated land fragmentation evaluation and empirical research: a case of Ningbo City in Zhejiang Province [J]. China Land Sci, 2016, 30(5): 80 − 87. [14] 刘彦随, 乔陆印. 中国新型城镇化背景下耕地保护制度与政策创新[J]. 经济地理, 2014, 34(4): 1 − 6. LIU Yansui, QIAO Luyin. Innovating system and policy of arable land conservation under the new-type urbanization in China [J]. Econ Geogr, 2014, 34(4): 1 − 6. [15] 朱莉芬, 黄季焜. 城镇化对耕地影响的研究[J]. 经济研究, 2007, 2(1): 137 − 145. ZHU Lifen, HUANG Jikun. Urbanization and cultivated land changes in China [J]. Econ Res, 2007, 2(1): 137 − 145. [16] 祝明霞. 九江市耕地变化及驱动力分析[J]. 贵州农业科学, 2013, 41(3): 153 − 155. ZHU Mingxia. Anlysis on cultivated land change and driving forces in Jiujiang City [J]. Guizhou Agric Sci, 2013, 41(3): 153 − 155. [17] 郭莎莎, 胡守庚, 瞿诗进. 长江中游地区多尺度耕地景观格局演变特征[J]. 长江流域资源与环境, 2018, 27(7): 1637 − 1646. GUO Shasha, HU Shougeng, QU Shijin. Multi-scale analysis of farmland landscape dynamic pattern in the middle reaches of the Yangtze river [J]. Resour Environ Yangtze Basin, 2018, 27(7): 1637 − 1646. [18] 邵晓梅, 杨勤业, 张洪业. 山东省耕地变化趋势及驱动力研究[J]. 地理研究, 2001, 20(3): 298 − 308. SHAO Xiaomei, YANG Qinye, ZHANG Hongye. A study on trend and driving forces of cultivated land use change in Shandong Province [J]. Geogr Res, 2001, 20(3): 298 − 308. [19] 刘旭华, 王劲峰, 刘明亮, 等. 中国耕地变化驱动力分区研究[J]. 中国科学D辑 地球科学, 2005, 35(11): 1087 − 1095. LIU Xuhua, WANG Jinfeng, LIU Mingliang, et al. Spatial heterogeneity of the driving forces of cropland changes in China [J]. Sci China Ser. D Earth Sci, 2005, 35(11): 1087 − 1095. [20] 史芳宁, 刘世梁, 安毅, 等. 城市化背景下景观破碎化及连接度动态变化研究:以昆明市为例[J]. 生态学报, 2020, 40(10): 3303 − 3314. SHI Fangning, LIU Shiliang, AN Yi, et al. Changes of landscape fragmentation and connectivity with urbanization: a case study of Kunming City [J]. Acta Ecol Sin, 2020, 40(10): 3303 − 3314. [21] 吴厚纯, 王成军, 费喜敏. 浙江省耕地数量变化的因素分解分析[J]. 浙江农林大学学报, 2015, 32(6): 933 − 939. WU Houchun, WANG Chengjun, FEI Ximin. Factor decomposition study of farmland change in Zhejiang Province [J]. J Zhejiang A&F Univ, 2015, 32(6): 933 − 939. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20200682
 点击查看大图
点击查看大图

计量
- 文章访问数: 2612
- HTML全文浏览量: 839
- PDF下载量: 65
- 被引次数: 0




