




-
景观格局是大小、形状、属性不同的景观单元在空间上的分布与组合[1]。一定区域内各种自然环境条件和社会经济因素共同作用形成了景观格局[2-3]。景观格局变化研究可以揭示区域景观演化的方向和驱动机制,预测景观变化趋势,进而提高区域景观资源的可持续管理[4]。国内外学者围绕景观格局变化开展了大量的研究工作,研究范围涉及流域[4]、湿地[5-7]、森林[8]与城市[9-12]等,研究内容包括景观格局时空变化[1-2]、驱动机制[5-6, 8]、生态系统服务价值[13-15]、生态环境效应[3-4, 16]、生态安全分析[10, 17]等。江苏省作为经济社会发展发达的省份之一,是景观格局变化的热点研究区域[18-19]。里下河平原位于江苏省中部,是典型的平原河网区,区内众多湖泊水系密布,是江苏极具水乡特色和生态价值的区域,是“南水北调”工程的重要水源地,也是淮河流域里下河地区主要的调蓄性湖泊群,其河湖健康在防洪、调蓄水量、引水排水、水体净化及保护生物多样性等方面具有重要的生态作用[20-21]。近年来里下河平原的景观格局变化显著,区域生物多样性和生态安全遭受严重威胁。当前学者围绕里下河平原的研究主要集中在水质、水系、养殖方面[20, 22-23],有学者探讨里下河平原景观格局的演变特征[6, 9, 24-25],但是都以某个城市为研究对象,基于长时间序列系统研究里下河平原景观格局时空演化的研究还鲜见报道,也未见景观格局变化方向、分布范围及热点方面的分析。基于此,本研究以里下河平原为研究区,选择1980、1990、2000、2010与2018年共5期景观类型数据,综合运用遥感和GIS技术,通过景观生态学和空间热点识别方法,分析里下河平原近40 a来的景观格局时空变化特征及热点区域,全面反映里下河平原景观演变的规律和原因。
-
里下河平原地处江苏省中部,北至苏北灌溉总渠,南至通扬运河,西起里运河(里河,京杭大运河苏中段)、东至串场河(下河),故称里下河平原,面积约11 556.87 km2,跨南通、泰州、扬州、淮安及盐城5个地级市。里下河平原为亚热带季风气候,区内多年平均降水量约1 000 mm,海拔为1.0~4.5 m,呈现中间低、四周高的特征,是著名的洼地[21];全区水网密布,湖荡相连,淡水沼泽湿地分布较为集中。通榆河、卤丁河、兴盐界河、射阳河、泰东河及新通扬运河等纵横交错。里下河平原是江苏省重要的粮食生产和水产养殖区域,对江苏省实现高质量发展具有重要意义。区内现有总人口约1 300万,耕地面积约8 500 km2。
-
采用的1980、1990、2000、2010和2018年共5期土地利用类型矢量数据来源于中国科学院资源环境科学数据中心(
http://www.resdc.cn/ )。在参考GB/T 21010—2007《土地利用现状分类》的基础上,结合研究区景观性质与特点,将景观分为7个一级类型:耕地、林地、草地、湖荡湿地、建设用地、未利用地和河流;其中,湖荡湿地进一步分为湖泊、水库坑塘、滩地与沼泽地。里下河平原5期景观类型图如图1所示。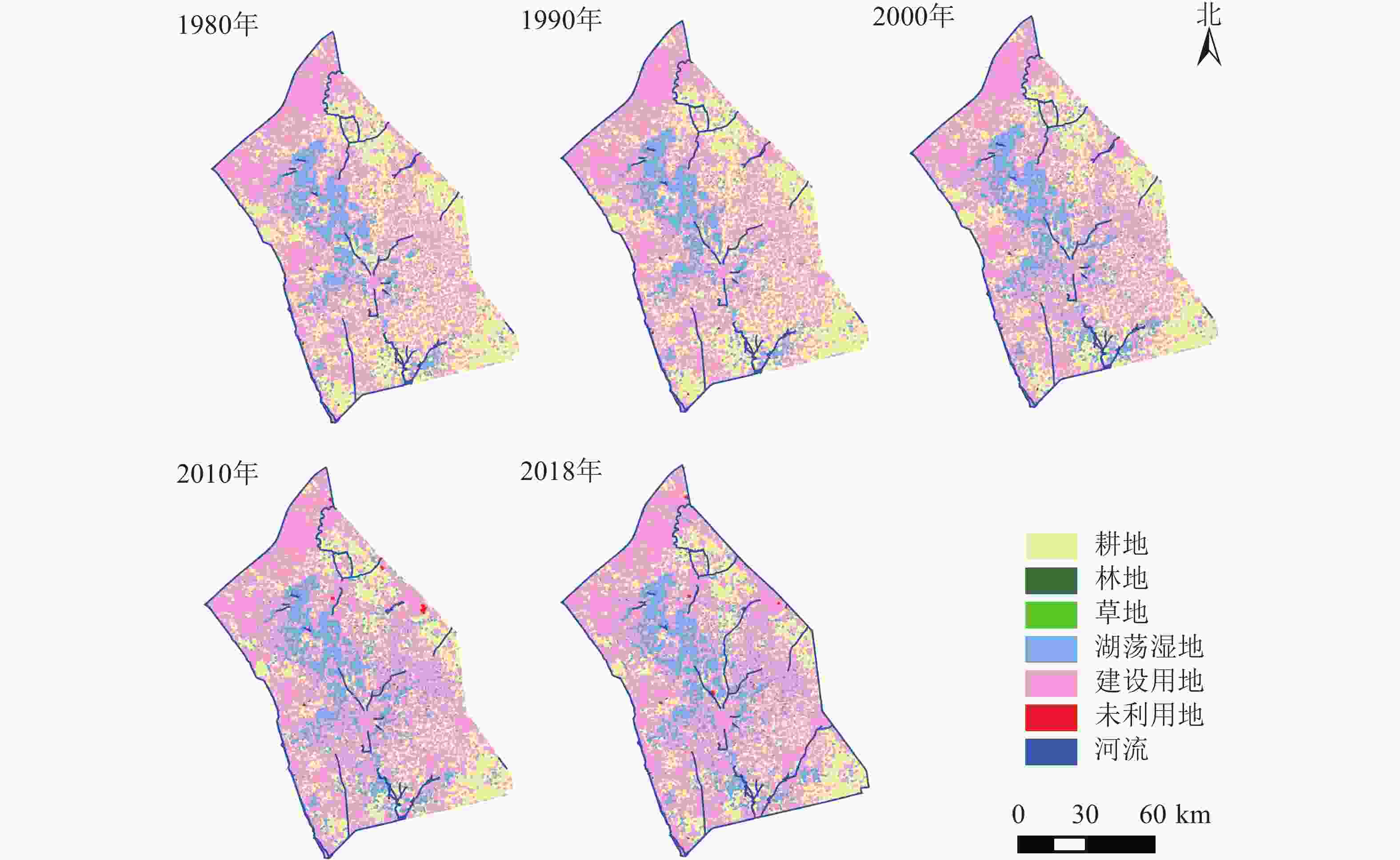
图 1 研究区1980—2018年景观类型示意图
Figure 1. Landscape type of the study area from 1980 to 2018
-
采用景观格局指数[3-4]、景观转移矩阵分析里下河平原近40 a来的景观格局变化规律。根据研究需要,在景观水平上选取7种景观格局指数,包括平均斑块面积(AREA_MN)、平均形状指数(SHAPE_MN)、平均分形维数(FRAC_MN)、景观形状指数(LSI)、聚集度(AI)、香农多样性指数(SHDI)以及香农均匀度指数(SHEI)。研究所用的软件为Fragstats 4.2。基于ArcGIS 10.2软件,采用景观转移矩阵分析研究区各景观类型的面积转移关系,详细的计算方法参考文献[5, 18]。
-
景观综合动态度用来表征研究区景观类型的变化速度[25]。采用景观综合动态度来揭示里下河平原不同研究时段景观类型的动态变化。为探究里下河平原不同时期景观动态变化的空间差异,对研究区进行网格化处理。参考相关文献和考虑研究区实际情况,采用5 km×5 km格网,总计524个格网,分别计算不同时期每个格网的景观综合动态度。格网景观综合动态度的计算公式如下:
$$L_{{\rm{C}}x}{\rm{ = }}\frac{{\displaystyle \sum\limits_{i = 1}^n {\Delta s_{x{{(i - j)}}}} }}{{2\displaystyle \sum\limits_{i = 1}^n s_{{x{i}}} }} \times \frac{1}{T} \times 100\% {\text{。}}$$ 其中:LCx为网格
$x$ 内的景观综合动态度,sxi为格网x上的某景观类型在T时段初期的面积,∆sx(i−j)为格网x上的某景观类型x在T时段内转化为j景观类型面积的绝对值,T为研究时段。运用ArcGIS 10.2中的自然断点法划分等级。 -
标准差椭圆可以在一定程度上反映研究要素的总体轮廓和主导分布方向[26-27]。标准差椭圆工具由3个基本要素组成,长半轴代表数据演化的主导方向,短半轴表示数据分布的范围,具体的计算过程见文献[28]。采用标准差椭圆分析里下河平原1980—2018年不同时期景观类型变化的方向和速度。
-
热点分析可以得到高值或低值要素在空间上的集聚状况,具体的计算公式可参考文献[29-30]。采用Getis-Ord Gi*热点分析工具对里下河平原1980—2018年的景观综合动态度进行冷热点统计。
-
里下河平原优势景观类型为耕地、湖荡湿地与建设用地,近40 a来在景观结构组成上变化明显(表1)。从1980到2018年,耕地面积持续减少了1 000.67 km2,减少幅度达10.45%,耕地在景观结构中的比例也从82.84%下降到74.18%。近40 a,湖荡湿地和河流面积表现出增加的趋势,其中,湖荡湿地面积从902.29 km2增加到1191.81 km2,在景观结构中的比例从7.81%提高到10.31%。这与研究区水产养殖产业的大力发展有关;河流面积从92.88 km2增加到120.44 km2。建设用地面积从1980年的972.55 km2增加到2018年的1 652.38 km2。草地和林地面积相对较小,两者的面积和比例呈波动变化,总体上林地面积有所增加,草地面积呈下降的趋势。研究区从2010年开始出现了未利用地景观,面积为5.73 km2,至2018年,未利用地面积下降到3.15 km2。
表 1 里下河平原1980—2018年景观结构
Table 1. Landscape structure of Lixiahe Plain from 1980 to 2018
景观类型 1980年 1990年 2000年 2010年 2018年 面积/km2 比例/% 面积/km2 比例/% 面积/km2 比例/% 面积/km2 比例/% 面积/km2 比例/% 耕地 9 573.81 82.84 9 544.21 82.58 9 345.89 80.87 8 737.68 75.61 8 573.14 74.18 林地 9.00 0.08 8.72 0.08 8.72 0.08 12.54 0.11 11.91 0.10 草地 6.34 0.05 6.40 0.06 6.10 0.05 4.00 0.03 4.04 0.03 湖荡湿地 902.29 7.81 903.81 7.82 1 025.14 8.87 1 172.42 10.14 1 191.81 10.31 建设用地 972.55 8.42 999.58 8.65 1 076.88 9.32 1 531.51 13.25 1 652.38 14.30 未利用地 0 0 0 0 0 0 5.73 0.05 3.15 0.03 河流 92.88 0.80 94.15 0.81 94.15 0.81 92.99 0.80 120.44 1.04 -
由表2可以看出:里下河平原的景观变化不仅体现在各景观类型的数量变化上,还体现在各景观类型的相互转化。近40 a各景观类型之间发生了显著的相互转化。按绝对量计算,转出面积最大的是耕地(1 126.50 km2),其次是建设用地(84.02 km2),各景观类型转出面积从大到小依次为耕地、建设用地、湖荡湿地、河流、草地、林地、未利用地。从转入看,转入面积最大的是建设用地(763.86 km2),建设用地扩张主要来自耕地和湖荡湿地。其次是湖荡湿地(354.14 km2),耕地转变为湖荡湿地是其面积增加的主要来源,湖荡湿地转入多于转出。转入面积从大到小依次为建设用地、湖荡湿地、耕地、河流、林地、未利用地、草地。
表 2 里下河平原1980—2018年景观转移矩阵
Table 2. Transfer matrix of landscape types in Lixiahe Plain from 1980 to 2018
景观类型 转移面积/km2 耕地 林地 草地 湖荡湿地 建设用地 未利用地 河流 1980年总面积 耕地 8 447.31 2.92 0.35 342.19 747.45 1.78 31.81 9573.81 林地 0.22 7.68 0.38 0.71 9.00 草地 0.19 0.21 2.81 1.61 0.36 1.17 6.34 湖荡湿地 49.97 0.45 0.71 837.68 10.88 1.30 1.30 902.29 建设用地 71.70 0.62 0.07 8.19 888.52 0.07 3.37 972.55 未利用地 − − − − − − − 0 河流 3.75 0.04 0.09 1.77 4.46 0 82.79 92.88 2018年总面积/km2 8573.14 11.91 4.04 1191.81 1652.38 3.15 120.44 11556.87 说明:−表示当年没有某种土地利用类型,空值表示地类之间无相互转化。行为1980年面积,列为2018年面积 由表3可知:近40 a来研究区景观格局变化明显,从1980—2018年,AREA_MN从457.824 9 hm2下降到387.378 5 hm2,减少幅度达15.39%。LSI由21.040 7上升到28.110 5,AI从81.616 5减少到75.048 8。AREA_MN与AI减小、LSI不断增加的变化趋势,表明研究区近40 a来景观破碎化程度不断加剧,斑块形状趋向复杂化。SHDI和SHEI分别从0.605 4和0.337 9增加到0.791 4和0.406 7,景观多样性和异质性增加,景观复杂程度也在增加。研究区耕地景观持续减少、建设用地不断扩张、养殖水面等景观类型快速增加,引起景观类型的多样性和复杂程度增加。
表 3 1980—2018年景观尺度上的景观格局指数
Table 3. Landscape indices at landscape level from 1980 to 2018
年份 AREA_MN/hm2 SHAPE_MN FRAC_MN LSI AI SHDI SHEI 1980 457.824 9 1.106 7 1.014 0 21.040 7 81.616 5 0.605 4 0.337 9 1990 450.155 8 1.106 6 1.014 0 21.389 5 81.292 7 0.612 1 0.341 6 2000 410.060 3 1.109 9 1.014 5 23.630 2 79.201 1 0.651 7 0.363 7 2010 389.467 5 1.137 2 1.017 0 27.545 3 75.572 1 0.760 4 0.390 7 2018 387.378 5 1.139 8 1.017 2 28.110 5 75.048 8 0.791 4 0.406 7 说明:AREA_MN为平均斑块面积,SHAPE_MN为平均形状指数,FRAC_MN为平均分形维数,LSI为景观形状指数,AI为聚集
度,SHDI为香农多样性指数,SHEI为香农均匀度指数研究区景观FRAC_MN总体上持续增大,表明区域内的景观类型受人类活动的影响增强,景观斑块的形状向不规则化发展。从1980—2018年,SHAPE_MN从1.106 7增加到1.139 8,表明景观形状不断趋向复杂化。SHAPE_MN与FRAC_MN表现出一致增加的演变趋势,表明研究区景观类型受到了日趋增强的人类活动干扰。
-
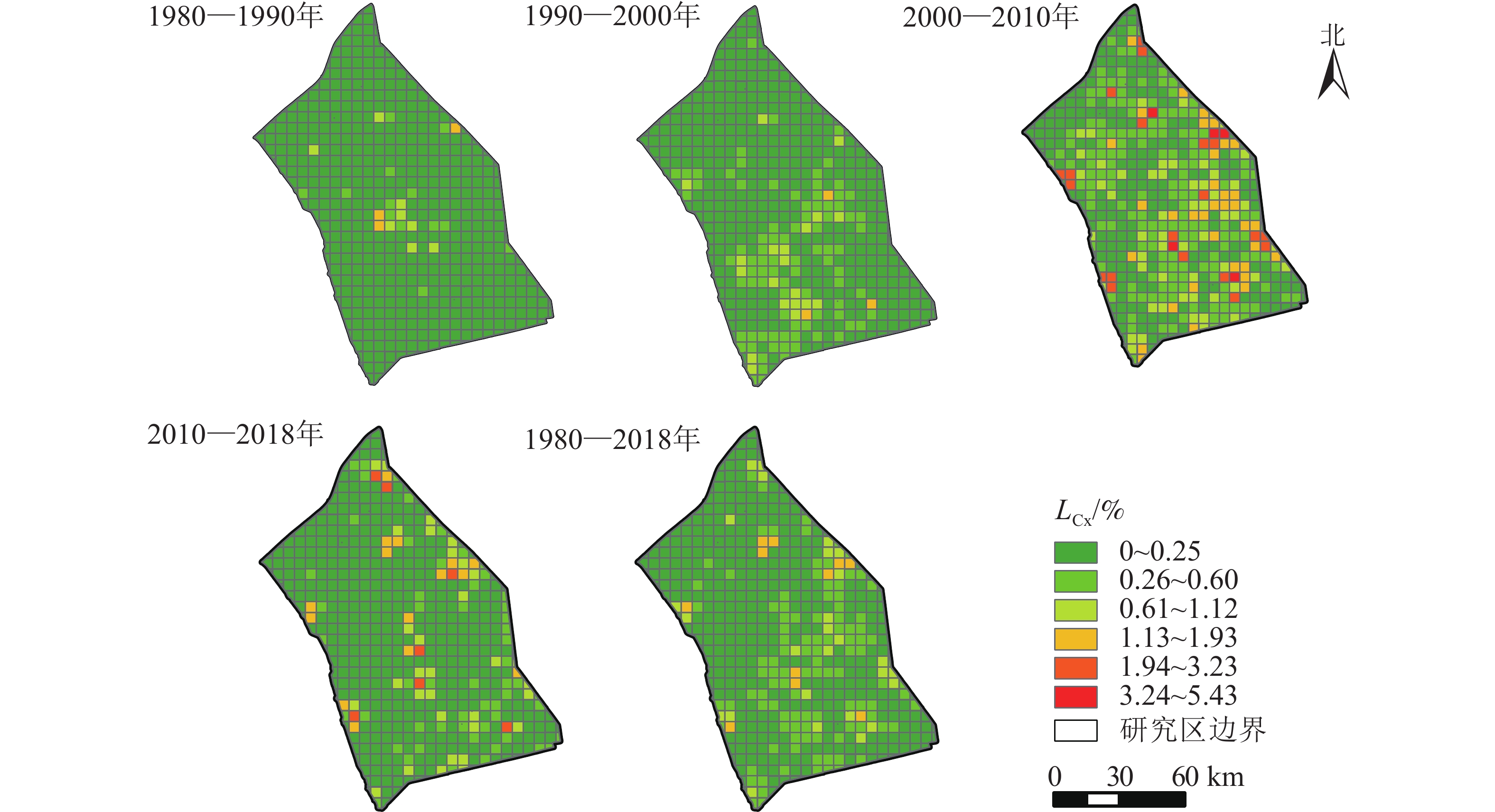
由图2可知:研究区在1980—2018年间,LCx>0.61%的网格面积占研究区7.06%,LCx>1.13%的只占2.10%。4期景观综合动态变化具有明显的时空差异。1980—1990年间,LCx>0.61%的网格面积占比仅2.10%,LCx>1.13%的网格面积只占0.57%,主要分布在里下河中部的兴化市内,其景观动态变化相对较大。1990—2000年,研究区的景观动态发生了显著变化,由中部地区向东北和西南方向扩张,高邮市与江都区内的景观变化速度加快。LCx>0.61%的网格面积增加到4.39%,LCx>1.13%的网格面积占0.57%。2000—2010年是研究区景观变化最为剧烈的时期,大多数区域的景观都发生了不同程度的变化。具有高动态度的网格明显增加,LCx>1.13%的网格面积占研究区的11.45%,LCx>1.94%的网格面积占4.77%。2010—2018年的LCx有明显的回落,景观综合动态变化有所放缓,LCx>1.13%的网格面积减少到4.01%。里下河平原近40 a的景观综合动态度经历了“缓慢变化—快速变化—变化减缓”的发展趋势。
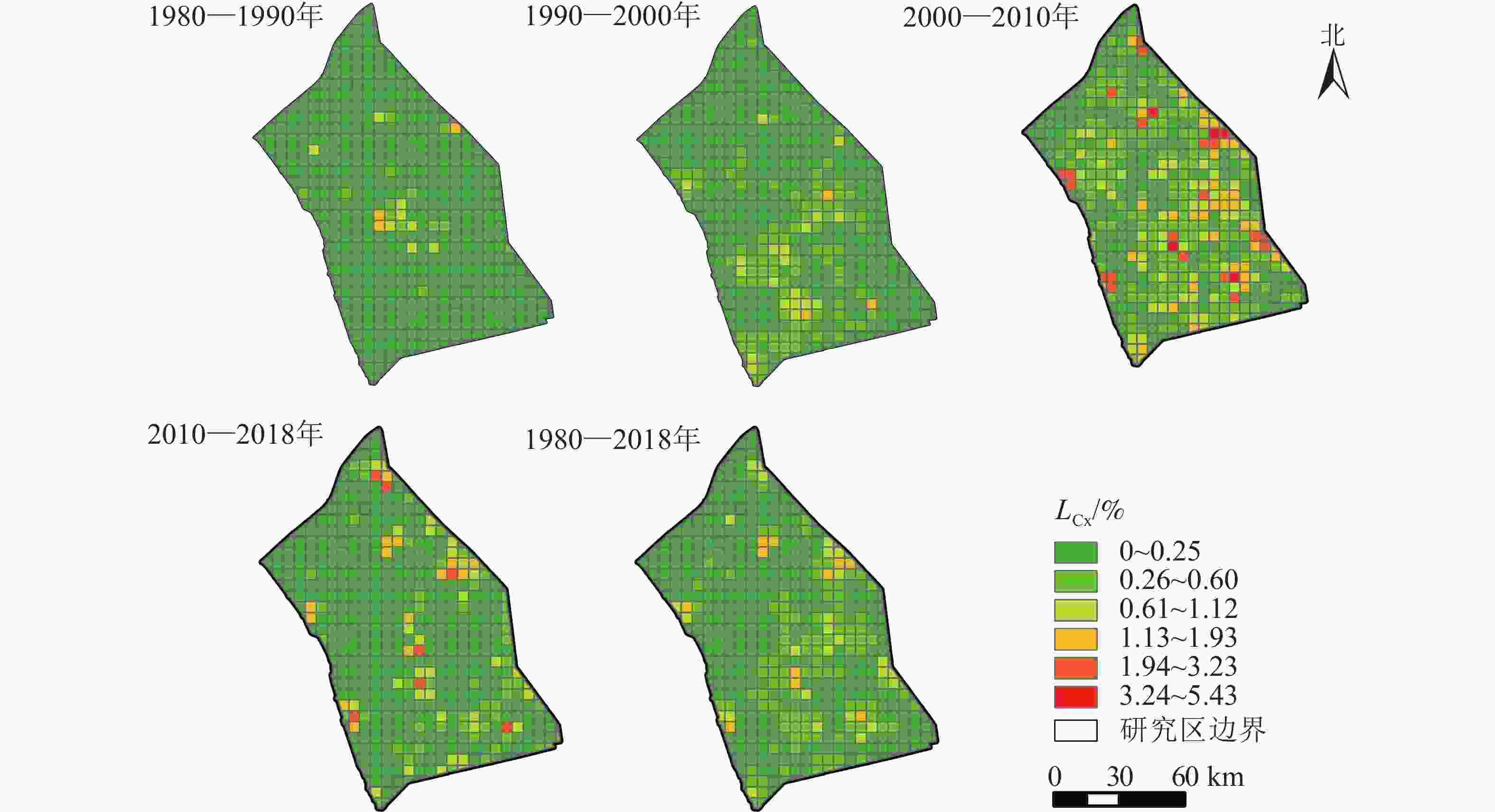
图 2 里下河平原景观综合动态度空间分布示意图
Figure 2. Spatial distribution of landscape integrated dynamic degree in Lixiahe Plain
-
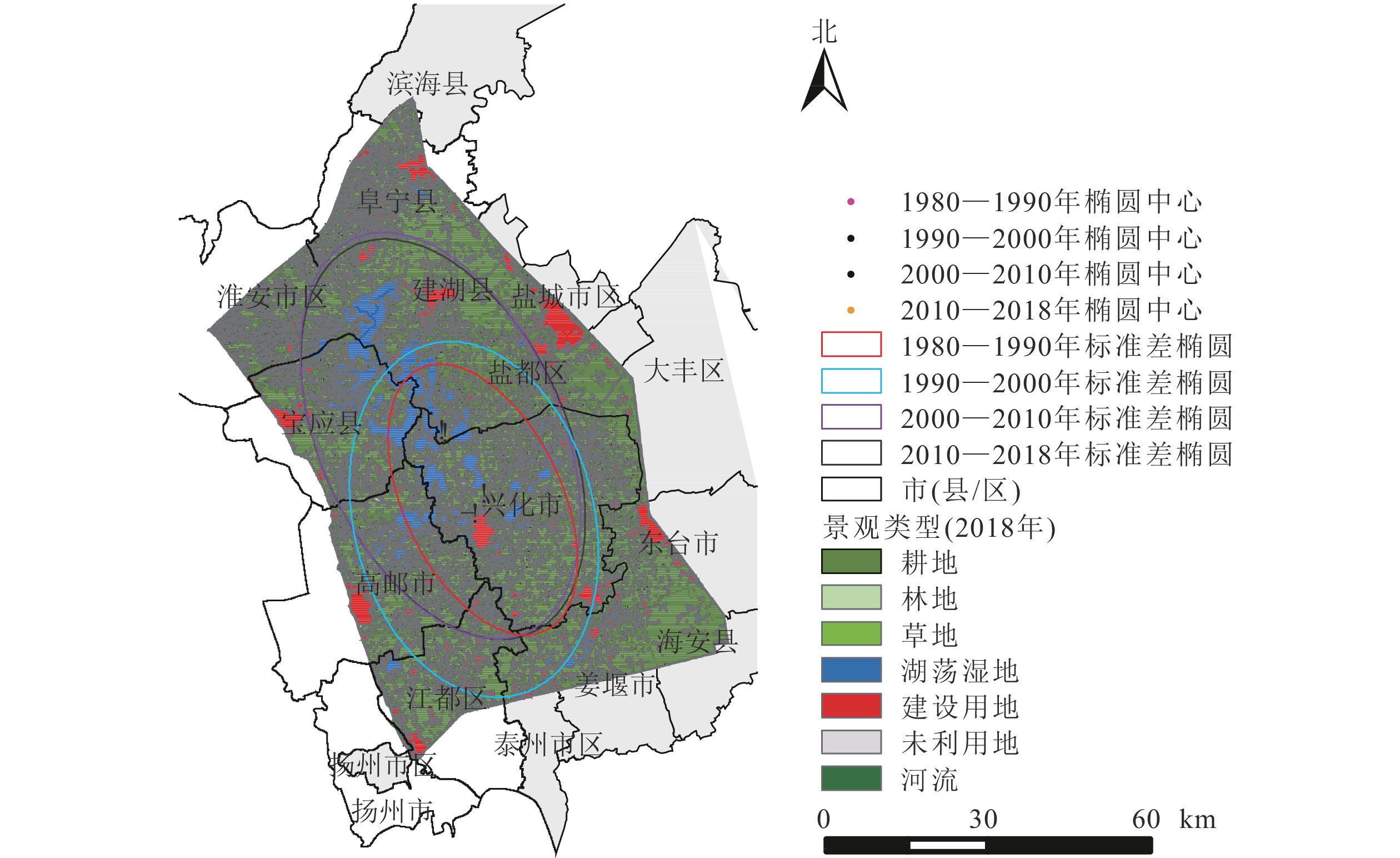
里下河平原景观变化的方向性显著,均以“东南—西北”方向为主轴发生不同角度的倾斜(表4、图3)。其中,1980—1990年与1990—2000年景观变化的中心位于兴化市。自2000年开始,景观类型变化标准差椭圆中心移动至盐都区,有较为明显的向西北方向移动的趋势,主轴的倾斜程度逐渐增大。
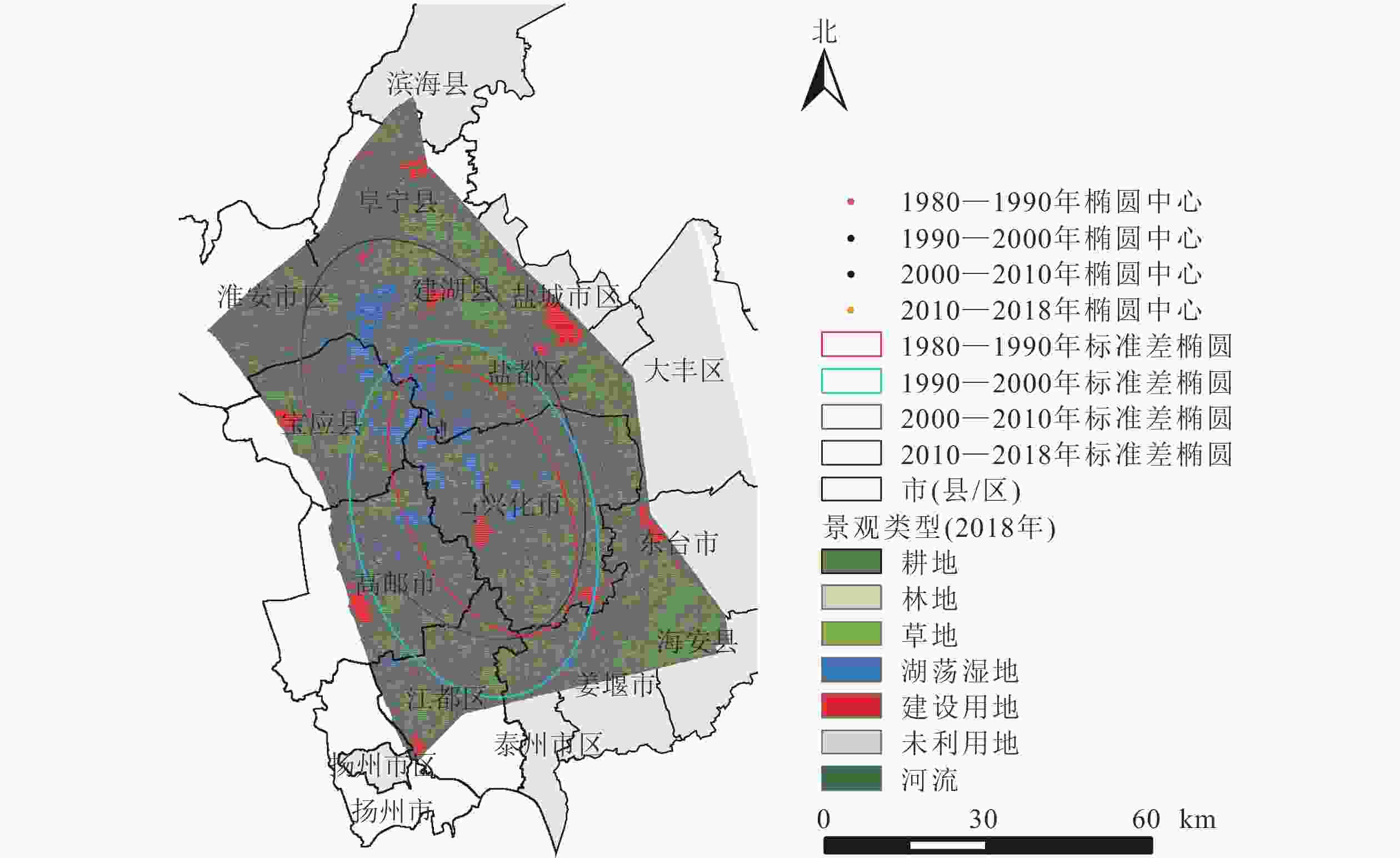
图 3 研究区不同时期景观变化标准差椭圆示意图
Figure 3. Standard deviation ellipse of landscape change
表 4 各研究区间标准差椭圆参数
Table 4. Standard deviation ellipse parameters in each period
起止年份 椭圆长/km 椭圆面积/km2 中心点纬度(N) 中心点经度(E) 标准距离长轴长度/km 标准距离短轴长度/km 旋转角度/(°) 1980—1990 189.24 2487.46 32°59′20″ 119°50′35″ 38.21 20.72 154.53 1990—2000 251.29 4725.24 32°56′46″ 119°48′36″ 47.56 31.63 166.19 2000—2010 283.28 5662.46 33°09′04″ 119°45′04″ 56.58 31.86 156.66 2010—2018 282.17 5661.47 33°08′31″ 119°45′25″ 56.03 32.16 155.35 研究期间景观类型标准差椭圆的大小变化明显,椭圆面积在1980—1990、1990—2000、2000—2010及2010—2018年4个时期内分别为2 487.46、4 725.24、5 662.46及5 661.47 km2,表明里下河平原景观变化在近40 a内呈现“先加剧后放缓”的特征。总体来说,4个时期的标准差椭圆变化幅度较大,范围基本覆盖了里下河平原经济社会发展的主要区域。表明研究区的景观类型从局部区域的剧烈变化扩展为更大区域的变化。
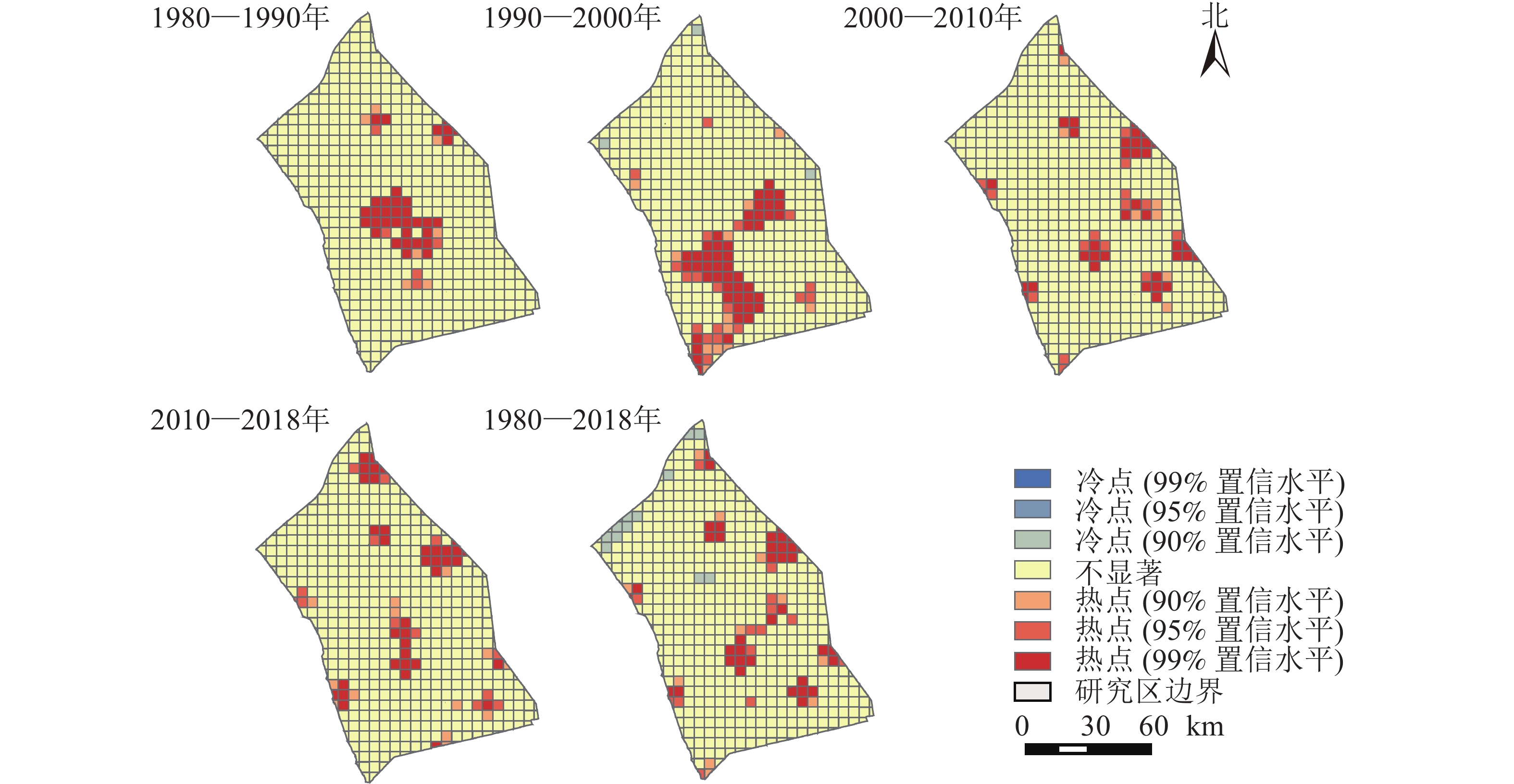
以格网为研究单元,采用ArcGIS空间统计中的Getis-Ord Gi*热点分析工具,选取置信度大于90%的具有显著统计特征的冷热点,探究研究区近40 a来景观变化的冷热点区域(图4)。整体而言,研究区西北方向的景观类型发生变化的概率较小。研究区中部、东北与西南部分地区是景观类型变化的热点区域,以兴化市、盐都区、高邮市及江都区的景观变化最为显著。4个时期的热点形成了明显的区域差异。具体表现为1980—1990年间的中部热点、1990—2000年的中部与西南热点到2020—2010、2010—2018年间的东北方向上的开发热点。除1990—2000年存在冷点区域外,其他3个时期无冷点区域。里下河平原历来是以耕地利用为主的农业开发地区,随着产业结构调整、人口增长和城镇化,里下河平原已经发展成为人口密集、产业集中和交通基础设施相对完善的地区。
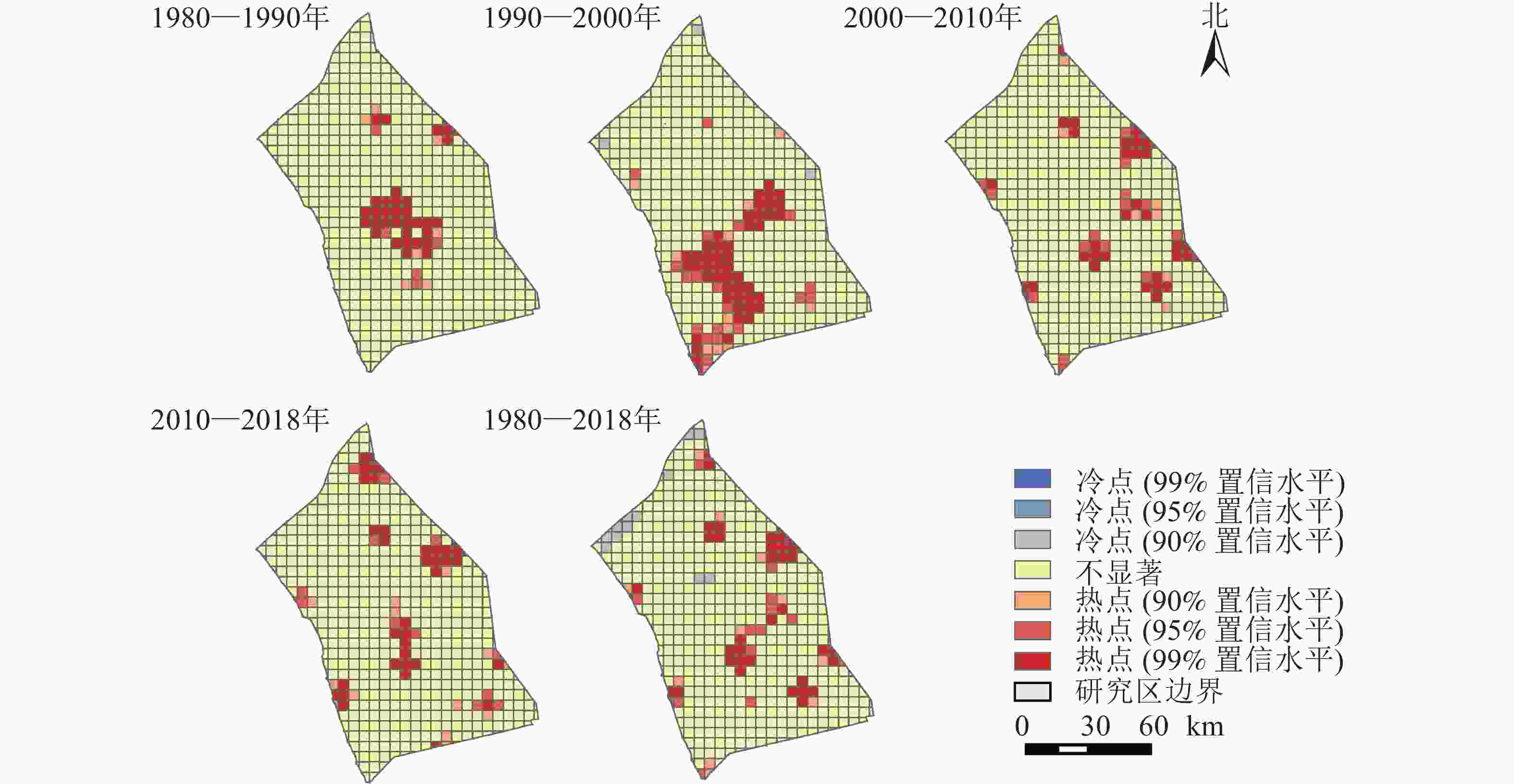
图 4 近40 a里下河平原景观动态变化热点示意图
Figure 4. Hot spots of landscape dynamic changes in Lixiahe Plain in the past 40 years
-
本研究系统探究了里下河平原近40 a来景观格局的时空演化规律和热点区域。里下河平原在1980—2018年景观格局变化明显,景观破碎化程度不断加剧,景观多样性、异质性与复杂程度增加。这与相关学者的研究成果基本一致,徐连芳等[6]研究表明:2002—2013年扬州市湿地景观整体呈破碎化趋势,景观多样性增加,湖泊面积下降,水库坑塘面积上升。SHI等[18]研究表明:江苏省在1990—2010年建设用地持续增加,耕地不断减少,2000—2010年土地利用动态度明显高于1990—2000年。马帅等[19]研究表明:2000—2015年江淮生态经济区土地利用与景观格局变化显著。整体上景观异质性程度提高,类型趋于多样化,人为因素对景观格局的影响不断扩大。蔡汉等[31]研究发现:受到快速城市化影响,扬州市耕地景观生态安全不断降低,社会经济影响力明显高于自然影响。
景观格局指数的变化表明:人口增加、城市化发展、产业结构调整、生态保护政策等人为因素是里下河平原景观格局变化的主要驱动力,深刻影响着里下河平原景观格局的变化。里下河平原是典型的平原河网地区,20世纪80年代该地区产业结构发生了调整,开展了大规模的围湖养殖运动,使里下河中部兴化市、西南部的高邮市和江都区景观格局发生了剧烈变化,部分天然湖泊、滩地被开发成养殖水面。近40 a来,水库坑塘的面积增加了418.81 km2,湖泊、滩地等自然湿地的面积减少了132.78 km2。2000—2010年是里下河平原景观格局变化最剧烈的时期,主要是由于人口增长、城镇化进程加快和养殖产业的发展,加剧了土地需求的多样化,大量耕地向建设用地和养殖水面转化,部分天然湖泊也转变成养殖鱼塘。江苏省城镇化率由2000年的41.5%上升至2010年的60.6%[32]。这一阶段耕地面积减少幅度达6.51%,城镇建设用地面积从1 076.88增加到1 531.51 km2,扩张幅度达42.22%。2010—2018年景观格局变化相对放缓,部分地区的养殖水面逐步恢复为天然湖泊,同时耕地保护政策使耕地面积减少幅度较前一个时期有所放缓。这得益于系列生态保护政策的实施。2008年,江苏省颁布实施《里下河腹部地区湖泊湖荡保护规划》,2010年里下河湖泊湖荡管理与保护联席会议制度建立,里下河湖泊湖荡网格化管理自2017年开始实施。2015年《江苏省湿地保护总体规划(2015—2030年)》正式施行。2017年《泰州市里下河生态经济示范区发展规划》正式实施,启动兴化市西北部湖荡湿地、大纵湖、姜堰区天然湖泊退圩还湖等生态修复工程。兴化市和盐都区也联合开展大纵湖的生态修复和退渔还湖,政策因素对里下河平原景观格局变化的影响日益增强。
本研究还结合标准差椭圆和热点分析探究了研究区景观格局变化的方向、空间分布范围及热点,但文中未做进一步的深入探究,如不同驱动因素对景观格局变化的影响力、贡献度及协同影响,不同类型的人类活动对景观格局演变及生态安全等方面的研究。里下河平原是重要的生态保护区域,城镇化进程显著加快,人类活动对区域景观格局的影响日益增强,在协调经济社会发展的同时要合理规划景观格局,要加大对天然湖泊湿地的恢复力度,增强河湖生态系统的连通性,提高区域景观格局的整体连续性。
-
①近40 a来里下河平原景观类型面积变化显著,耕地、草地面积分别减少了1 000.67、2.30 km2,建设用地、湖荡湿地、河流、林地面积分别增加了679.83、289.52、27.56、2.91 km2,耕地、建设用地和湖荡湿地用地面积变化较为剧烈。②近40 a来里下河平原景观类型之间相互转化频繁,尤其以耕地与湖荡湿地、建设用地之间的相互转化最为显著。耕地转出为建设用地和湖荡湿地的比例分别达66.35%和30.31%。耕地景观持续转出,湖荡湿地的净转入速率不断增加。建设用地在研究时段内都有来自耕地、湖荡湿地等景观类型的输入。林地、草地与耕地之间也存在相互转化,少量耕地与建设用地转化为未利用地。除耕地、草地的转出量大于转入量以外,其他景观类型的转入量均大于转出量。③标准差椭圆分析表明:近40 a研究区景观变化的方向性显著,以“东南—西北”为主轴倾斜,景观变化呈现“先加剧后放缓”的特征。兴化市、高邮市、盐都区及江都区是景观的热点变化区域。
Spatial-temporal variations of landscape pattern and hot spot analysis in the Lixiahe Plain of Jiangsu Province from 1980 to 2018
-
摘要:
目的 探讨景观格局时空变化及其冷热点,为区域生态安全与高质量发展提供科学依据。 方法 以江苏里下河平原1980、1990、2000、2010与2018年共5期景观类型数据为基础,采用景观格局指数、景观转移矩阵、景观综合动态度等方法,分析里下河平原近40 a景观格局的时空变化特征,基于热点分析探究景观变化的冷热点区域。 结果 研究区耕地、湖荡湿地和建设用地为优势景观类型,耕地是面积占比最多的景观,但整体上持续减少了1 000.67 km2,湖荡湿地和建设用地面积不断扩张,建设用地扩张面积达679.83 km2,草地和未利用地面积有所减少,林地面积有所增加。近40 a来,研究区各景观类型之间发生了复杂的相互转化,最显著的是耕地与湖荡湿地、建设用地之间的相互转化。研究期间格网景观综合动态度变化幅度为0~5.43%。里下河平原景观变化具有明显的方向性和聚集性,以“东南—西北”方向为主轴发生倾斜,兴化市、高邮市、盐都区及江都区是景观变化的热点区域。 结论 里下河平原景观格局变化显著,景观多样性增加、破碎化加剧,人类活动对景观格局的影响增强。自然湿地恢复政策的实施对研究区景观生态安全产生了积极影响。图4表4参32 Abstract:Objective With an exploration of the temporal and spatial changes of landscape pattern as well as an investigation of its cold and hot spots, this paper is aimed to provide a scientific basis for regional ecological security and high-quality development. Method With landscape type data collected of the Lixiahe Plain, Jiangsu Province over the five periods of 1980, 1990, 2000, 2010 and 2018, an analysis was made of the characteristics of temporal and spatial changes of its landscape pattern in the past 40 years using landscape pattern metrics, transfer matrix and comprehensive dynamic index. Then, an investigation was conducted of the cold and hot spot regions of landscape changes on the basis of hot spot analysis. Result Cultivated land, lake wetland and construction land were the dominant landscape types in the study area with the coverage of cultivated land accounting for the largest proportion, but having decreased by 1 000.67 km2 in the past 40 years whereas the areas of lake wetland and construction land increasingly expanded, with the area of construction land increasing by 679.83 km2 and the area of grassland and unused land showed a slightly decreasing trend, while that of forest increased slightly. The landscape types had undergone complex mutual transformations in the past 40 years with those of cultivated land, lake wetland and construction land being the most remarkable with the comprehensive dynamic index of the grid landscape varying between 0 and 5.43% over the study period. The landscape change of the Lixiahe Plain had obvious directionality and agglomeration: the center of change was tilted along the “southeast-northwest” direction, and the hot spots of landscape change were located at Xinghua City, Gaoyou City, Yandu District and Jiangdu District. Conclusion The landscape pattern of the Lixiahe Plain underwent significant changes due to the increase of landscape diversity and the intensification of fragmentation and the impact of human activities on the landscape pattern. The implementation of the natural wetland restoration policy had a positive impact on the landscape ecological security in the Lixiahe Plain. [Ch, 4 fig. 4 tab. 32 ref.] -
Key words:
- landscape pattern /
- spatial-temporal variations /
- hot spots analysis /
- Lixiahe Plain /
- Jiangsu Province
-
图 2 里下河平原景观综合动态度空间分布示意图
Figure 2 Spatial distribution of landscape integrated dynamic degree in Lixiahe Plain
图 4 近40 a里下河平原景观动态变化热点示意图
Figure 4 Hot spots of landscape dynamic changes in Lixiahe Plain in the past 40 years
表 1 里下河平原1980—2018年景观结构
Table 1. Landscape structure of Lixiahe Plain from 1980 to 2018
景观类型 1980年 1990年 2000年 2010年 2018年 面积/km2 比例/% 面积/km2 比例/% 面积/km2 比例/% 面积/km2 比例/% 面积/km2 比例/% 耕地 9 573.81 82.84 9 544.21 82.58 9 345.89 80.87 8 737.68 75.61 8 573.14 74.18 林地 9.00 0.08 8.72 0.08 8.72 0.08 12.54 0.11 11.91 0.10 草地 6.34 0.05 6.40 0.06 6.10 0.05 4.00 0.03 4.04 0.03 湖荡湿地 902.29 7.81 903.81 7.82 1 025.14 8.87 1 172.42 10.14 1 191.81 10.31 建设用地 972.55 8.42 999.58 8.65 1 076.88 9.32 1 531.51 13.25 1 652.38 14.30 未利用地 0 0 0 0 0 0 5.73 0.05 3.15 0.03 河流 92.88 0.80 94.15 0.81 94.15 0.81 92.99 0.80 120.44 1.04  下载: 导出CSV
下载: 导出CSV
表 2 里下河平原1980—2018年景观转移矩阵
Table 2. Transfer matrix of landscape types in Lixiahe Plain from 1980 to 2018
景观类型 转移面积/km2 耕地 林地 草地 湖荡湿地 建设用地 未利用地 河流 1980年总面积 耕地 8 447.31 2.92 0.35 342.19 747.45 1.78 31.81 9573.81 林地 0.22 7.68 0.38 0.71 9.00 草地 0.19 0.21 2.81 1.61 0.36 1.17 6.34 湖荡湿地 49.97 0.45 0.71 837.68 10.88 1.30 1.30 902.29 建设用地 71.70 0.62 0.07 8.19 888.52 0.07 3.37 972.55 未利用地 − − − − − − − 0 河流 3.75 0.04 0.09 1.77 4.46 0 82.79 92.88 2018年总面积/km2 8573.14 11.91 4.04 1191.81 1652.38 3.15 120.44 11556.87 说明:−表示当年没有某种土地利用类型,空值表示地类之间无相互转化。行为1980年面积,列为2018年面积
下载: 导出CSV
表 3 1980—2018年景观尺度上的景观格局指数
Table 3. Landscape indices at landscape level from 1980 to 2018
年份 AREA_MN/hm2 SHAPE_MN FRAC_MN LSI AI SHDI SHEI 1980 457.824 9 1.106 7 1.014 0 21.040 7 81.616 5 0.605 4 0.337 9 1990 450.155 8 1.106 6 1.014 0 21.389 5 81.292 7 0.612 1 0.341 6 2000 410.060 3 1.109 9 1.014 5 23.630 2 79.201 1 0.651 7 0.363 7 2010 389.467 5 1.137 2 1.017 0 27.545 3 75.572 1 0.760 4 0.390 7 2018 387.378 5 1.139 8 1.017 2 28.110 5 75.048 8 0.791 4 0.406 7 说明:AREA_MN为平均斑块面积,SHAPE_MN为平均形状指数,FRAC_MN为平均分形维数,LSI为景观形状指数,AI为聚集
度,SHDI为香农多样性指数,SHEI为香农均匀度指数
下载: 导出CSV
表 4 各研究区间标准差椭圆参数
Table 4. Standard deviation ellipse parameters in each period
起止年份 椭圆长/km 椭圆面积/km2 中心点纬度(N) 中心点经度(E) 标准距离长轴长度/km 标准距离短轴长度/km 旋转角度/(°) 1980—1990 189.24 2487.46 32°59′20″ 119°50′35″ 38.21 20.72 154.53 1990—2000 251.29 4725.24 32°56′46″ 119°48′36″ 47.56 31.63 166.19 2000—2010 283.28 5662.46 33°09′04″ 119°45′04″ 56.58 31.86 156.66 2010—2018 282.17 5661.47 33°08′31″ 119°45′25″ 56.03 32.16 155.35
下载: 导出CSV
-
[1] 邓楚雄, 刘睿娜, 聂小东, 等. 土地利用变化影响下湘西自治州景观格局的时空演变[J]. 四川师范大学学报(自然科学版), 2020, 43(3): 383 − 392. DENG Chuxiong, LIU Ruina, NIE Xiaodong, et al. The temporal and spatial evolution of landscape pattern under the effects of land use change in Xiangxi Autonomous Prefecture [J]. J Sichuan Norm Univ Nat Sci, 2020, 43(3): 383 − 392. [2] 陈利顶, 孙然好, 刘海莲. 城市景观格局演变的生态环境效应研究进展[J]. 生态学报, 2013, 33(4): 1042 − 1050. CHEN Liding, SUN Ranhao, LIU Hailian. Eco-environmental effects of urban landscape pattern changes: progresses, problems and perspectives [J]. Acta Ecol Sin, 2013, 33(4): 1042 − 1050. [3] 李阳兵, 王世杰, 李瑞玲, 等. 黔东山地丘陵区景观格局演变及其生态效应[J]. 山地学报, 2005, 23(1): 89 − 95. LI Yangbing, WANG Shijie, LI Ruiling, et al. Landscape pattern change of mountain-hill areas and its ecological effects in the east of Guizhou Province [J]. J Mount Sci, 2005, 23(1): 89 − 95. [4] 张宏锋, 欧阳志云, 郑华, 等. 新疆玛纳斯河流域景观格局变化及其生态效应[J]. 应用生态学报, 2009, 20(6): 1408 − 1414. ZHANG Hongfeng, OUYANG Zhiyun, ZHENG Hua, et al. Landscape pattern change and its ecological effect in Manas River Basin of Xinjiang [J]. Chin J Appl Ecol, 2009, 20(6): 1408 − 1414. [5] 吴金华, 房世峰 刘宝军, 等. 乌裕尔河-双阳河流域湿地景观格局演变及其驱动机制[J]. 生态学报, 2020, 40(3): 4279 − 4290. WU Jinhua, FANG Shifeng, LIU Baojun, et al. Landscape pattern evolution of wetland and its driving mechanism in Wuyuer-Shuangyang River Basin [J]. Acta Ecol Sin, 2020, 40(3): 4279 − 4290. [6] 徐连芳, 叶亚平. 扬州市湿地景观格局演变及驱动力研究[J]. 水生态学杂志, 2015, 36(3): 44 − 50. XU Lianfang, YE Yaping. Evolution of the wetland landscape pattern and its driving forces in Yangzhou [J]. J Hydroecol, 2015, 36(3): 44 − 50. [7] 张华兵, 刘红玉, 郝敬锋, 等. 自然和人工管理驱动下盐城海滨湿地景观格局演变特征与空间差异[J]. 生态学报, 2012, 32(1): 101 − 110. ZHANG Huabing, LIU Hongyu, HAO Jingfeng, et al. Spatiotemporal characteristics of landscape change in the coastal wetlands of Yancheng caused by natural processes and human activities [J]. Acta Ecol Sin, 2012, 32(1): 101 − 110. [8] 曹嘉铄, 邓政宇, 胡远东, 等. 神农架林区景观格局时空演变及其驱动力分析[J]. 浙江农林大学学报, 2021, 38(1): 155 − 164. CAO Jiashuo, DENG Zhengyu, HU Yuandong, et al. Spatial and temporal evolution and driving forces of the landscape pattern in Shennongjia Forest District [J]. J Zhejiang A&F Univ, 2021, 38(1): 155 − 164. [9] 吴健生, 罗可雨, 赵宇豪. 深圳市近20年城市景观格局演变及其驱动因素[J]. 地理研究, 2020, 39(8): 1725 − 1738. WU Jiansheng, LUO Keyu, ZHAO Yuhao. The evolution of urban landscape pattern and its driving forces of Shenzhen from 1996 to 2015 [J]. Geogr Res, 2020, 39(8): 1725 − 1738. [10] 朱亚楠, 蒲春玲. 乌鲁木齐市土地利用景观格局变化及生态安全分析[J]. 生态科学, 2020, 39(2): 133 − 144. ZHU Ya’nan, PU Chunling. Analysis on landscape pattern change and ecological security of land use in Urumqi [J]. Ecol Sci, 2020, 39(2): 133 − 144. [11] DENG Jinsong, WANG Ke, YANG Hong, et al. Spatial-temporal dynamics and evolution of land use change and landscape pattern in response to rapid urbanization [J]. Landscape Urban Plann, 2009, 92(3/4): 187 − 198. [12] DADASHPOOR H, AZIZI P, MOGHADASI M. Land use change, urbanization, and change in landscape pattern in a metropolitan area [J]. Sci Total Environ, 2019, 655: 707 − 719. [13] 邵大伟, 吴殿鸣. 景观破碎化对生态系统服务价值的影响作用分析[J]. 长江流域资源与环境, 2020, 29(11): 2436 − 2449. SHAO Dawei, WU Dianming. Analysis on the effect of landscape fragmentation on ecosystem service value: a case study of Suzhou [J]. Resour Environ Yangtze Basin, 2020, 29(11): 2436 − 2449. [14] 贾艳艳, 唐晓岚, 杨阳, 等. 长江沿岸芜湖段景观格局演变及生态系统服务价值分析[J]. 西北林学院学报, 2019, 34(6): 249 − 258. JIA Yanyan, TANG Xiaolan, YANG Yang, et al. Landscape pattern changes and ecological service values in Wuhu Section along the Yangtze River [J]. J Northwest For Univ, 2019, 34(6): 249 − 258. [15] ZHANG Jing, QU Mei, WANG Cheng, et al. Quantifying landscape pattern and ecosystem service value changes: a case study at the county level in the Chinese Loess Plateau [J]. Glob Ecol Conserv, 2020, 23: 1 − 14. [16] ZHU Congmou, ZHANG Xiaoli, ZHOU Mengmeng, et al. Impacts of urbanization and landscape pattern on habitat quality using OLS and GWR models in Hangzhou, China [J/OL]. Ecol Indic, 2020, 117: 106654[2021-02-15]. doi: 10.1016/j.ecolind.2020.106654. [17] MA Libang, BO Jie, LI Xiaoyang, et al. Identifying key landscape pattern indices influencing the ecological security of inland river basin: the middle and lower reaches of Shule River Basin as an example [J]. Sci Total Environ, 2019, 674: 424 − 438. [18] SHI Ge, YE Peng, DING Liang, et al. Spatio-temporal patterns of land use and cover change from 1990 to 2010: a case study of Jiangsu Province, China [J/OL]. Int J Environ Res Public Health, 2019, 16: 907[2021-02-15]. doi: 10.3390/ijerph16060907. [19] 马帅, 程浩, 林晨, 等. 江淮生态经济区土地利用景观格局时空变化[J]. 水土保持研究, 2021, 28(1): 292 − 299. MA Shuai, CHENG Hao, LIN Chen, et al. Spatialtemporal changes of land use and landscape pattern in Jianghuai ecological economic zone [J]. Res Soil Water Conserv, 2021, 28(1): 292 − 299. [20] 吕慧华, 周峰, 李娜, 等. 苏北里下河典型区河网水系演变特征研究[J]. 长江流域资源与环境, 2018, 27(2): 380 − 385. LÜ Huihua, ZHOU Feng, LI Na, et al. Evolution characteristics of the river network in the typical area of Lixiahe Region [J]. Resour Environ Yangtze Basin, 2018, 27(2): 380 − 385. [21] 谭少华, 倪绍祥, 周飞. 苏北里下河地区湿地资源可持续利用的思考[J]. 生态与农村环境学报, 2003, 19(3): 61 − 64. TAN Shaohua, NI Shaoxiang, ZHOU Fei. Sustainable utilization of wetland in Lixiahe region in north Jiangsu Province [J]. Rural Eco-Environ, 2003, 19(3): 61 − 64. [22] 马小雪, 王腊春. 里下河地区主要水环境污染物的空间分布特性研究[J]. 水资源与水工程学报, 2014, 25(6): 1 − 6. MA Xiaoxue, WANG Lachun. An analysis of temporal and spatial variation characteristics of water quality in Lixiahe Abdominal Area [J]. J Water Resour Water Eng, 2014, 25(6): 1 − 6. [23] 谢奎, 程家驿, 毕朱, 等. 江苏省养殖池塘与池塘养殖业地理分布格局研究[J]. 中国农业资源与区划, 2020, 41(3): 178 − 188. XIE Kui, CHENG Jiayi, BI Zhu, et al. Study on geographical distribution pattern of aquaculture pond and pond-culture industry in Jiangsu Province [J]. Chin J Agric Resour Reg Plann, 2020, 41(3): 178 − 188. [24] 李鑫, 欧名豪, 马贤磊. 基于景观指数的细碎化对耕地利用效率影响研究: 以扬州市里下河区域为例[J]. 自然资源学报, 2011, 26(10): 1758 − 1767. LI Xin, OU Minghao, MA Xianlei. Analysis on impact of fragmentation based on landscape index to cultivated land use efficiency: a case on Lixiahe District in Yangzhou City [J]. J Nat Resour, 2011, 26(10): 1758 − 1767. [25] 尚正永, 张小林, 卢晓旭, 等. 城市主要功能用地景观格局的演变特征[J]. 长江流域资源与环境, 2012, 21(12): 1528 − 1535. SHANG Zhengyong, ZHANG Xiaolin, LU Xiaoxu, et al. Evolution characteristics of landscape pattern of urban functional land use [J]. Resour Environ Yangtze Basin, 2012, 21(12): 1528 − 1535. [26] 张丽, 杨国范, 刘吉平. 1986—2012年抚顺市土地利用动态变化及热点分析[J]. 地理科学, 2014, 34(2): 185 − 191. ZHANG Li, YANG Guofan, LIU Jiping. The dynamic changes and hot spots of land use in Fushan City from 1986 to 2012 [J]. Sci Geogr Sin, 2014, 34(2): 185 − 191. [27] 李一琼, 白俊武. 近20 a来苏州土地利用动态变化时空特征分析[J]. 测绘科学, 2018, 43(6): 58 − 64. LI Yiqiong, BAI Junwu. Analysis of temporal and spatial dynamic changes of land use of Suzhou in recent 20 years [J]. Sci Surv Mapp, 2018, 43(6): 58 − 64. [28] 李丹, 梅晓丹, 张玉娟, 等. 绥芬河市土地利用变化的时空格局研究[J]. 测绘科学, 2019, 44(3): 78 − 85. LI Dan, MEI Xiaodan, ZHANG Yujuan, et al. Research of spatiotemporal pattern of land use change in Suifenhe City [J]. Sci Surv Mapp, 2019, 44(3): 78 − 85. [29] 雷金睿, 陈宗铸, 陈小花, 等. 1980—2018年海南岛土地利用与生态系统服务价值时空动态变化[J]. 生态学报, 2020, 40(14): 1 − 14. LEI Jinrui, CHEN Zongzhu, CHEN Xiaohua, et al. Spatio-temporal changes of land use and ecosystem services value in Hainan Island from 1980 to 2018 [J]. Acta Ecol Sin, 2020, 40(14): 1 − 14. [30] CAO Yu, CAO Yu, LI Guoyu, et al. Linking ecosystem services tradeoffs, bundles and hotspot identification with cropland management in the coastal Hangzhou Bay area of China [J/OL]. Land Use Polic, 2020, 97: 104689[2021-02-15]. doi: 10.1016/j.landusepol.2020.104689. [31] 蔡汉, 朱权, 罗云建, 等. 快速城市化地区耕地景观生态安全格局演变特征及其驱动机制[J]. 南京林业大学学报(自然科学版), 2020, 44(5): 181 − 188. CAI Han, ZHU Quan, LUO Yunjian, et al. Evolution characteristics and driving mechanisms of cultivated landscape eco-security patterns in rapid urbanization areas [J]. J Nanjing For Univ Nat Sci Ed, 2020, 44(5): 181 − 188. [32] 陈小卉, 胡剑双. 新中国成立以来江苏城镇化和城乡规划的回顾与展望[J]. 规划师, 2019, 19(35): 25 − 31. CHEN Xiaohui, HU Jianshuang. Review and prospect of urbanization and urban-rural planning in Jiangsu Province since the foundation of the People’s Republic of China [J]. Planners, 2019, 19(35): 25 − 31. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210178
 点击查看大图
点击查看大图

计量
- 文章访问数: 2514
- HTML全文浏览量: 580
- PDF下载量: 68
- 被引次数: 0


