






-
山核桃Carya cathayensis为落叶乔木,是含油率极高的胡桃科Juglandaceae高档干果,其果实为世界四大名优坚果之一[1-2]。因其口味独特、营养价值高而受到人们的广泛喜爱。浙江省杭州市临安区是中国山核桃主产区之一,种植历史悠久[3]。近几十年来,临安山核桃产业迅猛发展,现有种植面积已达5.3万 hm2,已成为当地林农重要的经济来源[4]。山核桃在临安广泛种植,但由于各镇林地有着不同的土壤特征,导致经济效益存在较大差异,即使在同一乡镇,产量大小年份的情况也依然存在[5]。这些问题在一定程度上限制了山核桃产业的发展。国内对于山核桃生长规律、规范施肥、产量提升等方面多有研究。杨慧思等[6]发现:山核桃产地土壤中大量及微量元素的空间分布特征与变异规律有着对应关系;张红桔等[7]揭示了山核桃产区主要养分因子和产量的关系以及空间分布特征;丁立忠等[8]研究结果指出:近10 a临安7 个山核桃主产镇的林地土壤有机碳含量总体呈升高趋势,而土壤碱解氮、有效磷和速效钾含量下降明显,养分现状不容乐观。但是有关水解酶活性与土壤肥力的关系,以及在山核桃产区空间上的分布特点还没有详细的报道和深入研究。水解酶与土壤中营养物质循环、能量转化以及环境质量等密切相关,并参与了土壤环境中重要的生物化学过程[9]。水解酶活性的空间分布特点可以灵敏地反映不同区域土壤中物质循环的速率,这种速率极大程度影响着林地土壤生态系统的功能。同时,水解酶活性能表征土壤养分转化的潜力,度量土壤污染程度和生产力,是人们评价土壤质量和土壤健康的重要经验指标之一[10],因此,对土壤水解酶活性空间分布特点的研究十分重要。
本研究分析山核桃林土壤水解酶活性的空间分布特点,并通过地统计方法揭示土壤养分与水解酶的空间异质性及其影响因子,为林农掌握土壤肥力特性及养分转化潜力,合理经营山核桃林提供理论依据和技术支撑,对山核桃产业可持续发展具有重要的指导意义。
-
研究区位于浙江省杭州市临安区(31°14′N,119°42′E),是中国山核桃的核心产区,属亚热带季风气候,年平均气温为16.0 ℃,极端最高和最低气温分别为41.7和−13.3 ℃。山核桃产区土壤以红壤土类分布最广,多发育于泥岩、页岩、砂岩、凝灰岩、花岗岩、流纹岩以及第四世纪红土[11]。山核桃是该地重要的经济树种,主要分布在海拔50~1100 m的丘陵和山地。林农一般在每年5月上旬和9月上旬将氮肥、复合肥与微肥配合施用。
样地按1 km×1 km网格在全市范围内布设,与临安区森林资源分布图相叠加,有山核桃分布的网格点即为山核桃林样地。根据均匀分布原则,2019年7—8月在山核桃分布较集中的岛石、龙岗、清凉峰、昌化、河桥、湍口、太阳等7个镇,共选取259个样地。在选定的区域内,采集0~20 cm的5个样点土壤样品后,混合,并在样地中心以全球定位系统(GPS)定位,记录经纬度。土壤样品带回实验室,去除石块、植物残体等异物,用木棒碾碎后过2 mm筛。将土壤样品分成2份,一份置于4 ℃冰箱保存,尽快测定土壤酶活性;另一份摊开后在室内自然风干,用于测定土壤养分指标和pH。
-
测定涉及与土壤碳、氮、磷循环的α-葡萄糖苷酶(AG)、β-葡萄糖苷酶(BG)、纤维二糖水解酶(CBH)、木糖苷酶(XYL)、亮氨酸氨基肽酶(LAP)、N-乙酰-β-氨基葡萄糖苷酶(NAG)、酸性磷酸酶(PHOS)等7种土壤水解酶。土壤酶活性测定参照SAIYA-CORK等[12]的荧光微孔板检测技术。具体操作为:称取2 g鲜土于离心管中,加入30 mL提前配置好的pH为5.0的醋酸铵缓冲液,在25 ℃ 180 r·min−1摇床上震荡30 min,再用70 mL醋酸铵缓冲液冲洗至烧杯中,用磁力搅拌器搅拌1 min,用排枪取200 μL土壤悬液于96孔板中,并立即加入50 μL反应底物,放入25 ℃培养箱中避光培养3 h,取出后迅速加入15 μL 0.5 mol·L−1的氢氧化钠结束反应,用多功能酶标仪检测吸光值并计算土壤酶活性。土壤养分指标和pH参照鲁如坤[13]方法测定:土壤pH采用土水比(质量比)为1.0∶2.5的悬浊液,微电极法测定;土壤有机质采用浓硫酸-重铬酸钾外加热法测定;土壤碱解氮采用碱解扩散法测定;土壤有效磷采用盐酸-氟化铵(HCl-NH4F)浸提,钼锑抗比色法测定;土壤速效钾采用醋酸铵浸提,火焰光度计测定。
-
水解酶活性的空间分布特征采用地统计分析方法。半方差变异函数是研究区域化变量模型,其公式为:
$$ y=\frac{1}{2 N} \sum_{i=1}^{N}[Z(x_i)-Z(x_i+h)]^{2} 。 $$ 其中:y表示间隔距离为h点的半方差值,h为空间间隔点的距离,即步长;N为样点观测数值成对的数目;Z(xi+h)和Z(xi)为样点测定值[14]。常用的半方差变异函数模型有球状(Spherical)、高斯(Gaussian)、指数(Exponential)[15]模型。模型中,C0代表块金值,C+C0代表基台值,当块基比C0/(C+C0)<25%时,表明变量具有强烈的空间自相关,即主要受到结构性变异的影响;当C0/(C+C0)为25%~75%时,变量属于中等程度空间自相关;当C0/(C+C0)>75%的,变量空间自相关程度较弱,即主要受随机因素影响[16-17]。
采用全局莫兰指数(Ig)来体现研究区域土壤水解酶的空间自相关水平。使用GeoDa软件对样点构建空间权重矩阵,可以根据局部莫兰指数(IL)绘制局部空间自相关聚类图[18],采用Canoco 5.0对酶活性高、低聚集样点间总体差异采用999次的蒙特卡罗排列检验(Monte Carlo permutation test,999 permutations,full model),然后找出土壤酶活性关系密切的环境因子进行冗余分析(RDA)。
$$ {I_{\rm{g}}} = {\frac{{N\displaystyle \sum\limits_{i = 1}^N {{W_{ij}}({Z_i} - \overline Z )({Z_j} - \overline Z )} }}{{\displaystyle \sum\limits_{i = 1}^N {\displaystyle \sum\limits_{j = 1}^N {{W_{ij}}{{({Z_i} - \overline Z )}^2}} } }}_{}} ; $$ $$ {I_{\rm{L}}} = \frac{{{Z_i} - \overline Z }}{{{\sigma ^2}}}\sum\limits_{i = 1}^N {[{W_{ij}}({Z_i} - \overline Z )]} 。 $$ 其中:
$\overline Z$ 为变量Z的平均值;Zi、Zj分别是变量Z在空间i、j处的数值(i≠j);σ2是变量Z的方差;Wij是Zi与Zj之间的空间权重函数,在选定尺度内时赋予权重值为1,超过尺度时权重值为0。当Ig>0,表示目标样点与邻近样点有空间结构上的相似性,存在空间集聚区;当Ig<0,表示空间区域异常[19]。 -
用SPSS 22.0对土壤水解酶活性与土壤性质数据进行描述性统计分析,计算变异系数,Pearson相关性分析设置显著水平为0.05。在Excel 2007中进行数据的对数转化。采用因子分析中的主成分分析法(PCA)和系统聚类分析,对山核桃林地土壤肥力情况进行评价[20]。通过因子分析确定参评土壤指标主成分特征值和特征向量,选取特征值较高的关键主成分,计算各主成分得分,再利用得分公式求出各样点土壤肥力分值(IIF)[21],采用类平均法对分值进行系统聚类。肥力分值计算采用肥力指数和法。公式为:
$$ I_{\rm{IF}} = \frac{{{x_1}}}{{\displaystyle \sum\limits_{i = 1}^m {{x_i}} }}{F_1} + \frac{{{x_2}}}{{\displaystyle \sum\limits_{i = 1}^m {{x_i}} }}{F_2} + \frac{{{x_3}}}{{\displaystyle \sum\limits_{i = 1}^m {{x_i}} }}{F_3}+ \cdots + \frac{{{x_n}}}{{\displaystyle \sum\limits_{i = 1}^m {{x_i}} }}{F_n} 。 $$ 可简化为:IIF=λ1F1+λ2F2+λ3F3
$+\cdots+ $ λnFn。其中:Fi表示单个主成分得分;λi表示对应主成分解释的总变异。 -
土壤pH平均为5.76,最小为4.50,说明山核桃林地土壤主要为酸性土壤,且酸化较为严重。土壤有机质为5.41~98.08 g·kg−1,平均为37.39 g·kg−1;土壤有效磷为0.52~22.43 mg·g−1;土壤速效钾为22.06~466.07 mg·g−1;土壤碱解氮为28.62~192.53 mg·g−1。研究区域土壤变异系数从大到小依次为有效磷、速效钾、有机质、碱解氮、pH。土壤酶活性变异系数从大到小依次为CBH、XYL、AG、NAG、LAP、BG、PHOS,其中CBH、XYL为高度变异,AG、NAG、LAP、BG、PHOS为中等变异(表1)。根据ZHANG等[22]对变异系数的划分,当变异系数<10%时为弱变异,在10%~90%时为中等变异,>90%时则为高度变异。研究区域有效磷质量分数变异系数>90%,具有明显的变异性,为高度变异。这可能是由于山核桃产区地势起伏较大,在雨水冲刷后有效磷极易流失,且林农施用磷肥量不均匀,土壤利用率较低,常年累积造成。土壤pH变异系数为10.28%,变异较小,与张红桔等[7]研究结果基本一致,说明山核桃林地土壤总体pH比较接近。
表 1 土壤性质描述性统计分析
Table 1. Descriptive statistics of soil properties
项目 有机质/
(g·kg−1)有效磷/
(mg·kg−1)速效钾/
(mg·kg−1)碱解氮/
(mg·kg−1)pH AG/
(mol·g−1·h−1)最小值 5.41 0.52 22.06 28.62 4.50 0.12 最大值 98.08 22.43 466.07 192.53 7.48 1.67 平均值 37.39 4.23 113.77 132.40 5.76 0.35 标准差 15.38 3.90 72.58 43.47 0.59 0.30 变异系数/% 41.15 92.20 63.80 32.83 10.28 88.12 项目 BG/
(mol·g−1·h−1)CBH/
(mol·g−1·h−1)XYL/
(mol·g−1·h−1)LAP/
(mol·g−1·h−1)NAG/
(mol·g−1·h−1)PHOS/
(mol·g−1·h−1)最小值 4.58 0.04 0.17 0.10 0.15 18.31 最大值 192.62 63.06 62.91 32.95 93.60 1042.63 平均值 47.06 8.14 7.44 4.44 17.75 160.43 标准差 31.50 8.09 6.91 3.62 15.27 89.10 变异系数/% 66.95 99.48 92.98 81.54 86.07 55.54 -
土壤有机质和碱解氮与7种水解酶均呈极显著正相关(P<0.01);有效磷与BG 、NAG呈极显著正相关,与CBH呈显著正相关(P<0.05);pH与CBH呈极显著正相关(P<0.01),与BG呈显著正相关(P<0.05),与PHOS、XYL呈极显著负相关(P<0.01);速效钾与7种水解酶均没有表现出明显的相关性(表2)。土壤酶主要来源于土壤微生物、土壤动物和植物根系的分泌,土壤养分质量分数可以直接影响土壤动植物长势、微生物的活性与分布,整合了土壤理化条件的信息[23],因此土壤水解酶活性与土壤养分因子密切相关,水解酶活性也通常被作为土壤质量的生物活性指标[24]。对土壤水解酶活性与土壤肥力指标及pH的相关性分析发现:土壤有机质和碱解氮与7种酶存在极显著正相关(P<0.01)。土壤有机质是评价土壤肥力的重要指标,土壤有机质的形成与分解都与酶的作用有关[25]。水解酶可以吸附在土壤有机质上,以酶-腐殖质复合物的形式从土壤中提取出来,并仍可保留有活性。土壤碱解氮主要集中在土壤表层,其含量受人为施肥的影响较大[26]。山核桃林农为提高产量大量撒施氮肥。氮素供应的增加可以使植物细胞原生质合成加快,细胞数量增多,有了更多的水解酶产出渠道。此外,土壤微生物也会通过分泌多种水解酶固定氮素[27]。PHOS与有效磷质量分数和pH之间存在显著正相关和极显著负相关。由于PHOS会参加土壤磷的矿化作用过程,使土壤有机态磷转化为植物可吸收的无机态磷,有效磷质量分数增加。pH是控制土壤中磷有效性和PHOS活性的关键因子,在山核桃林地土壤酸化的环境下,pH小幅降低可能有利于植物对有效磷的吸收,PHOS参与矿化作用的活性增强[28]。
表 2 土壤水解酶与养分因子及pH相关性分析表
Table 2. Correlation coefficients of soil hydrolase activities and soil nutrient factors and pH
水解酶 有机质 有效磷 速效钾 碱解氮 pH AG 0.355** 0.061 0.060 0.419** 0.102 BG 0.406** 0.172** 0.066 0.354** 0.147* CBH 0.356** 0.158* 0.060 0.275** 0.196** XYL 0.302** 0.088 −0.090 0.278** −0.283** LAP 0.170** 0.042 −0.015 0.230** −0.028 NAG 0.431** 0.267** 0.114 0.357** 0.109 PHOS 0.272** 0.123* 0.007 0.346** −0.286** 说明:*P<0.05, **P<0.01 -
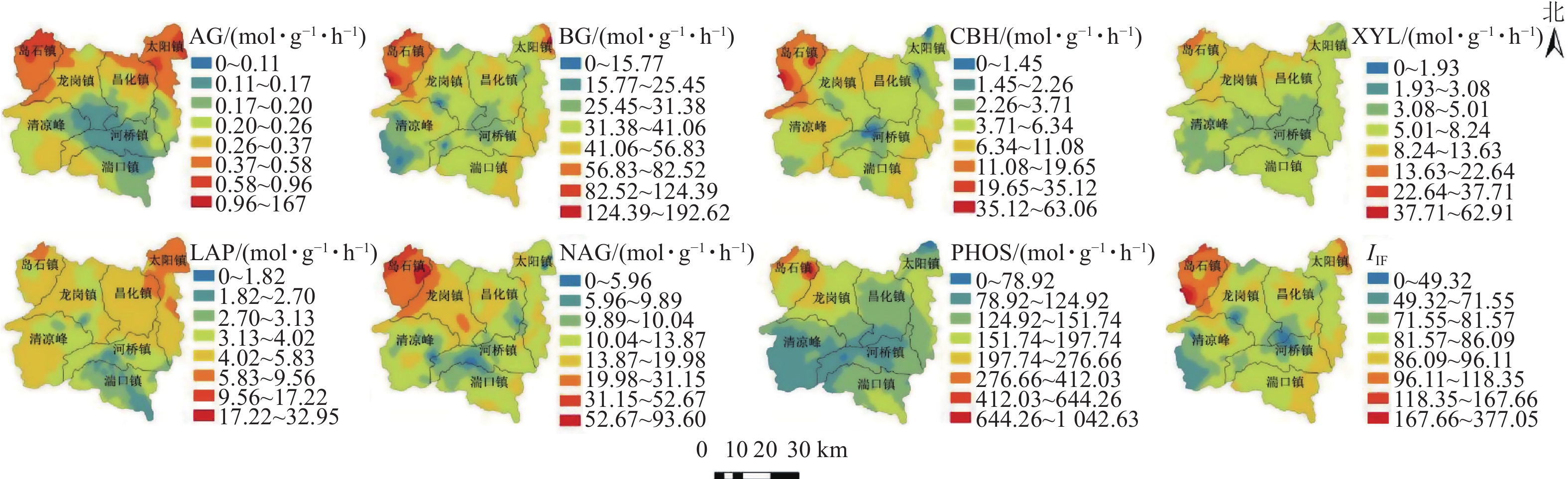
7种土壤水解酶活性在山核桃林地表现出不同的空间异质性(表3)。在GS+ 9.0中进行方差变异函数拟合,分别选用最优模型。不同水解酶拟合模型的差异,表示相应的酶在土壤中的空间变化规律不同。本研究中,AG、BG、CBH、LAP、NAG、XYL活性具有中等空间自相关性,活性均呈现斑块状分布;不同水解酶活性区域分布呈现差异性,块基比分别为55%、42%、56%、49%、66%、47%、78%,说明它们的变异情况主要受到人为因素和结构性因素共同影响。PHOS块基比为78%,具有较弱的空间自相关,其活性空间分布主要受随机因素如施肥方式和耕作强度的影响[29]。本研究中, 虽然各酶活性的空间自相关性存在差异,但变程均大于最小采样距离,因此,本研究所选择的采样距离能够反映土壤水解酶活性在研究区域最小尺度下的空间变异特征。从土壤水解酶活性的空间分布情况来看(图1):活性较高的区域位于临安区西北方向的岛石镇。该镇有中国“山核桃第一镇”的美称,这可能与当地特色生态化经营方式有关,如:林下种植茶Camellia sinensis、黑麦草Lolium perenne、油菜Brassica napus等适生植物,丰富林下土壤生态结构的同时起到涵养水肥、改良酸性土质的作用;林间饲养家禽吃掉害虫、消灭虫卵,产生的鸡粪也可作为有机肥为山核桃树生长提供养分。这些特色经营方式都为植物根系发育、土壤微生物的快速新陈代谢提供了有利的条件,从而提高了相关水解酶参与复杂生化反应的活性。
表 3 土壤水解酶活性半方差函数理论模型及其相关参数
Table 3. Theoretical model of semi-variance function of soil hydrolase activities and its related parameters
水解酶 函数模型 块金值(C0) 基台值(C+C0) 块基比[C0/(C+C0)] 变程 决定系数 AG 球状模型 0.050 0.090 0.55 9.63 0.46 BG 指数模型 136.800 324.400 0.42 8.76 0.65 CBH 指数模型 0.140 0.250 0.56 1.60 0.43 XYL 高斯模型 0.080 0.170 0.47 2.12 0.37 LAP 高斯模型 10.970 21.960 0.49 11.20 0.73 NAG 高斯模型 197.150 294.260 0.66 27.30 0.45 PHOS 球状模型 0.032 0.041 0.78 14.60 0.54 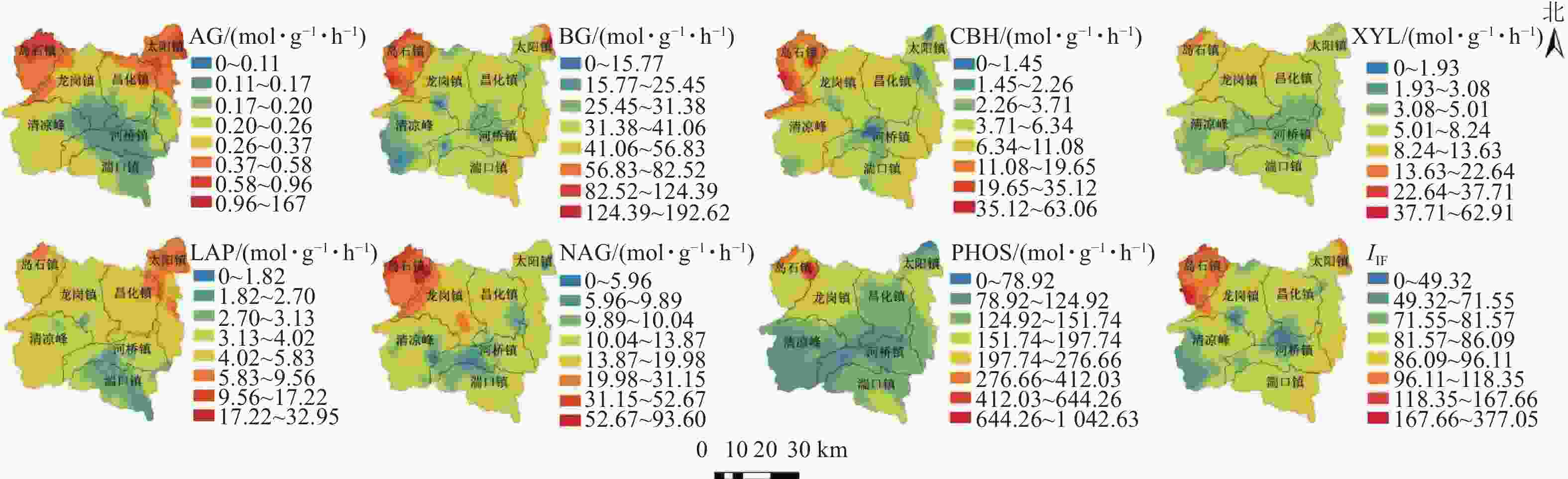
图 1 水解酶活性空间分布示意图
Figure 1. Spatial distributions of soil hydrolase activities
根据全局莫兰指数,AG、BG、CBH、XYL、LAP、NAG、PHOS等7种水解酶活性在空间分布上都存在相关性(Ig>0),且存在高低值聚集区域。在本研究中,7种水解酶活性在高低值聚类上呈现出了相似的特点,岛石镇为水解酶活性高值聚集(high-high)区域,部分水解酶(如AG)在太阳镇北部也有高值聚集的现象;低值聚集(low-low)区域多位于清凉峰以及河桥、龙岗、昌化三镇交界处附近(图2)。在冗余分析结果中,高低值聚集区域样点的水解酶活性在第1轴有明显的分离,第1轴和第2轴分别有82.92%和14.42%的贡献度(图3)。结果显示:碱解氮、有机质、pH与水解酶活性有极显著正相关(P<0.01)。水解酶活性高低值聚类结果和土壤肥力分值高低值聚类情况相似,说明土壤水解酶活性大小和周围可利用的营养物质关系十分密切。由于山核桃林地土壤常年受到不同程度的人为经营干扰,造成这种现象的原因多为当地经营方式的不同,如氮肥、有机肥的投入是增加土壤肥力较为直接的方式,因此各区域出现高低值聚集的现象可以一定程度反映当地肥力水平以及施肥情况。岛石镇高值聚集,一方面可能是由于岛石镇山核桃林氮肥、有机肥常年投入量高于清凉峰等区域,同时岛石镇明确规定当地所有山核桃林地禁用除草剂,防止除草剂的不合理使用破坏产区生态平衡,影响山核桃产量。另一方面,岛石镇相对其他镇海拔较高,大部分产区山高树茂,年降水量充沛,林下、林间生态系统的结构与功能较为完整,因此土壤微生物活动旺盛,作物根系发达,从而成为水解酶活性的高值聚集区。而清凉峰以及河桥、龙岗、昌化交界处的冒尖山、石柱山、云台山、鸡哺山等区域地势极为陡峭,当地山核桃林水土流失现象严重,养分较为贫乏,相对其他区域处于较低的水平,水解酶活性受到影响,产生低值聚集。杭瑞高速经过清凉峰镇与龙岗镇南部,该区域交通便利,人类活动造成了一定程度的干扰。当地存在铅锌矿、钨钼矿以及铜矿等正在开发的金属矿[30]。采矿活动产生的粉尘、废水和尾矿渣可能会对土壤水解酶活性和分布造成一定影响[31]。
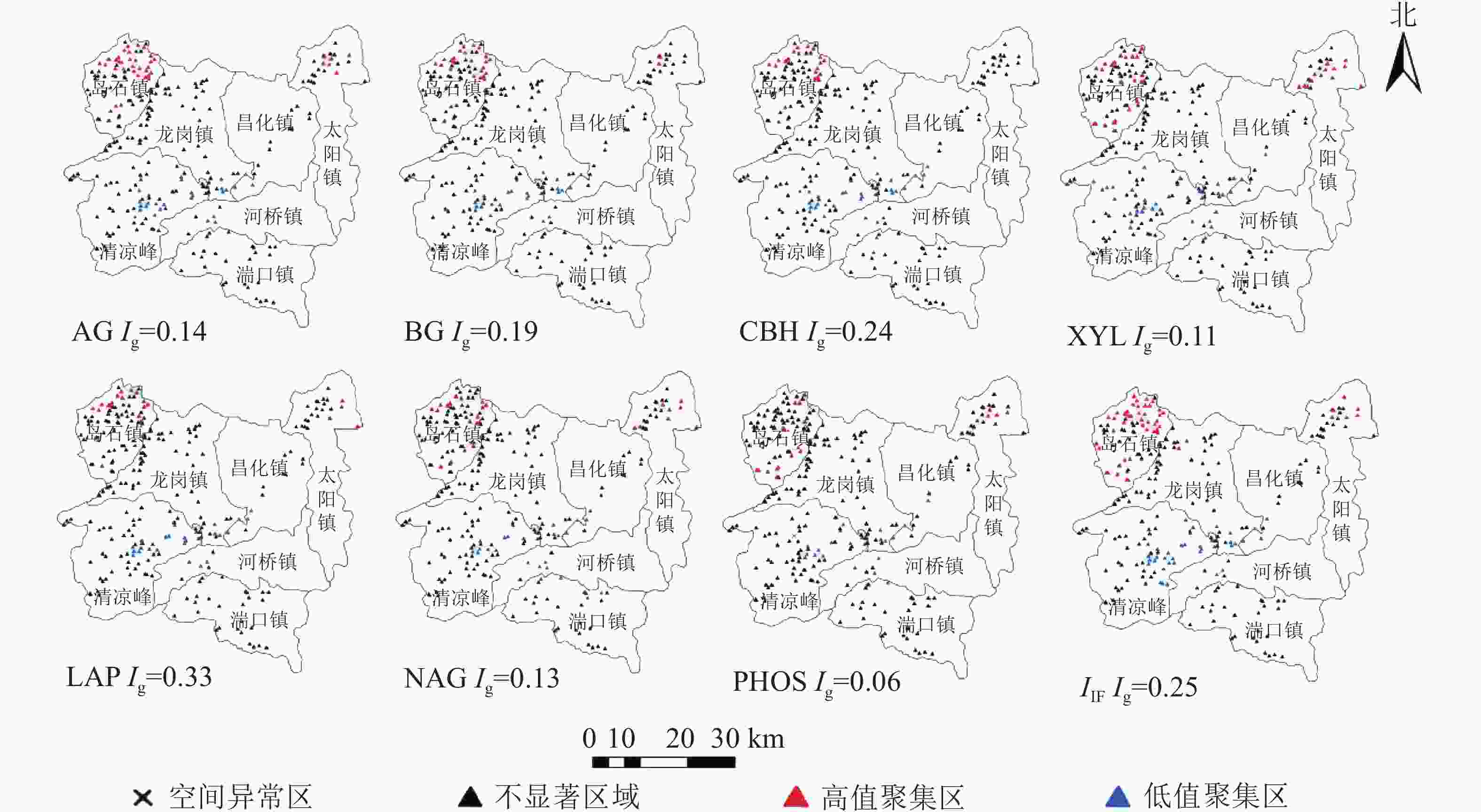
图 2 采样点高低值聚类示意图
Figure 2. High-low-value cluster of sampling points
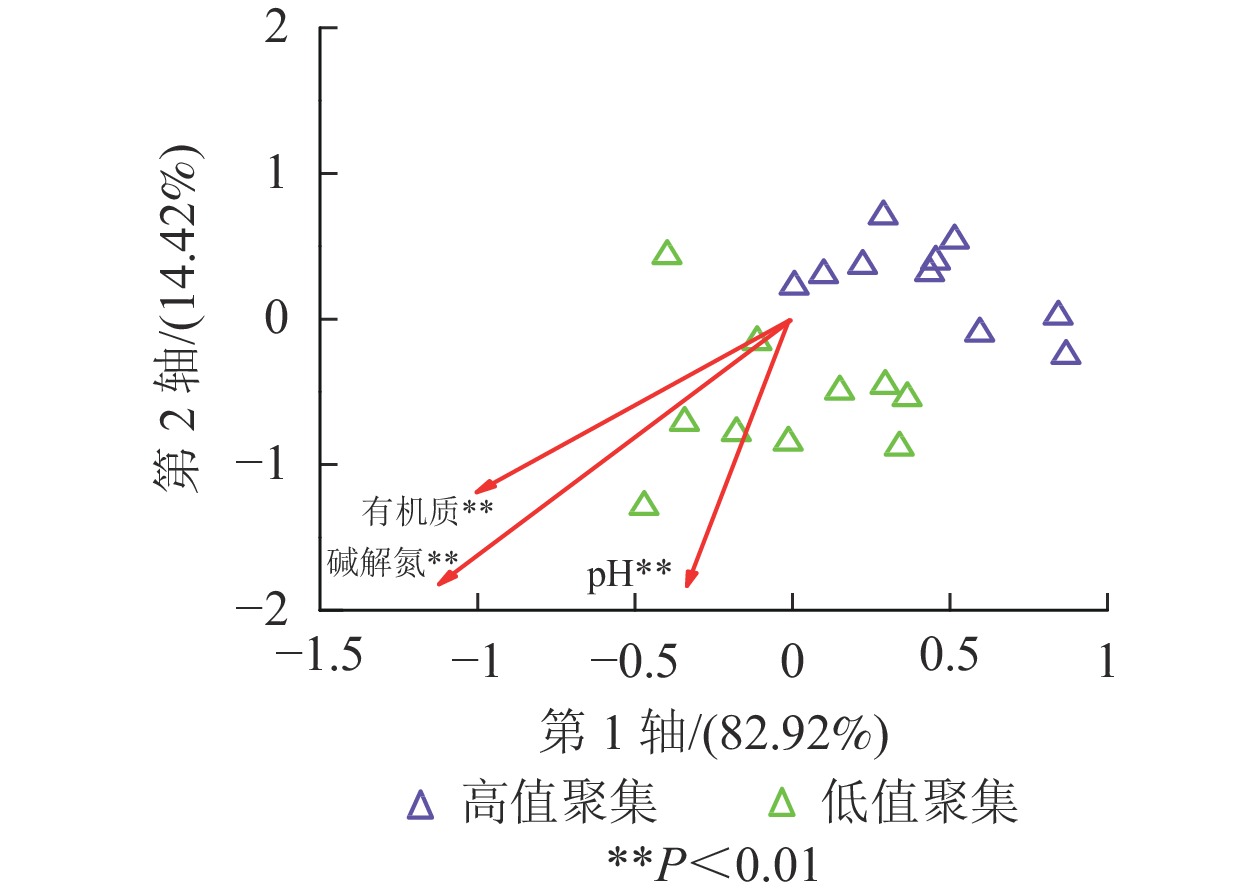
图 3 高低聚类点冗余分析结果
Figure 3. Redundancy analysis of high-high and low-low clustering points
-
据浙江省地方标准,山核桃林地土壤样地中碱解氮、有效磷、速效钾、有机质位于丰富等级的样地分别占64%、56%、23%、45%,绝大部分样点各养分指标等级处于中等以上(表4),能够满足山核桃的生长发育需求,但是由于各区域土壤中养分比例以及酸碱度的不同,综合肥力状况也有所差异。在主成分分析结果中,IIF均值为91.67,变幅为35.29~277.05,变异系数为42.73%。第1主成分解释了48.39%的总变异,第2主成分解释了26.50%的总变异,第3主成分解释了17.12%的总变异(表5)。采用类平均法对土壤肥力分值(IIF)进行系统聚类,将259个样本分为4类,即第Ⅰ类IIF≥146.83,第Ⅱ类87.11≤IIF<146.83,第Ⅲ类59.72≤IIF<87.11,第Ⅳ类IIF<59.72,分别对应土壤肥力高、较高、中、低4个等级。结果显示:IIF变异系数为42.73%,属于中等变异,但其中有174个样本处于中低水平,肥力相对较差。从采样区域土壤肥力分值所占比例来看,山核桃林地有58.7%的样地土壤肥力低于平均水平,有32.7%的样地土壤肥力为Ⅰ和Ⅱ等级,大部分样地土壤肥力处于Ⅲ、Ⅳ等级,说明大部分山核桃林地土壤肥力还有提高的空间。从使用IIF绘制的空间分布图来看,岛石镇山核桃林地土壤肥力在所有山核桃产区中最高,该区域的土壤管理方法值得借鉴。沈一凡等[32]研究了近10 a山核桃林地主要分布区域的土壤养分变化情况,发现林地土壤酸化的现象一直在加重,肥力也有不断下降的趋势。这是由于大多数山核桃林农缺乏相关技术指导和对立地环境的认知,长期施用以氮素为主的化学肥料造成的。而且从20世纪80年代开始,山核桃林地不断扩张,但大多数新兴产区酸化严重,土壤宜肥、宜种性较差。针对这一现象,还需要增施有机肥,并施用一定量的石灰,逐渐改善各地土壤酸化的情况,规范林地生草管理和生态化采收技术,以稳步提升山核桃林地的土壤肥力。
表 4 山核桃土壤肥力指标丰缺等级及各等级占比
Table 4. Level of soil fertility indexs and the proportion of each level
项目 碱解氮 有效磷 速效钾 有机质 质量分数/(mg·kg−1) 占比/% 质量分数/(mg·kg−1) 占比/% 质量分数/(mg·kg−1) 占比/% 质量分数/(g·kg−1) 占比/% 缺乏 <80 6 <5 12 <80 34 <10 7 中等 80~120 30 5~10 32 80~110 43 10~40 48 丰富 >120 64 >10 56 >110 23 >40 45 说明:土壤肥力指标丰缺等级参考浙江省地方标准 DB33/T 2205—2019《山核桃分区施肥技术规范》 表 5 主成分贡献率与各因子得分
Table 5. Principal component contribution rates and each factor score
因子 主成分得分 第1主成分(48.39%) 第2主成分(26.50%) 第3主成分(17.12%) 有机质 0.144 0.268 0.225 有效磷 0.071 0.451 0.116 速效钾 0.027 0.481 0.046 碱解氮 0.061 0.040 0.652 AG 0.160 −0.058 0.039 BG 0.205 0.017 −0.299 CB 0.182 0.064 −0.393 XYL 0.177 −0.240 0.042 LAP 0.122 −0.238 0.345 NAG 0.179 0.111 −0.130 PHOS 0.184 −0.219 0.100 -
研究区山核桃林土壤水解酶活性均具有较好的空间变异结构和空间分布格局,结构性变异占总变异的比例较小。研究区山核桃林土壤受到人为因素的干扰较多,人为施肥与经营强度是影响其空间格局形成的最直接因素。土壤水解酶活性空间分布和养分分布联系密切,在养分质量分数较高的区域有高值聚集的现象,低值聚集区域多位于清凉峰等区域。特色的生态经营方式可以使土壤水解酶活性处于相对较高的水平,从而提高山核桃的宜种性。
研究区山核桃林土壤酸化较为普遍,平均pH为5.76,严重限制了山核桃的生长。岛石镇、太阳镇北部土壤肥力得分较高。从总体来看,大部分区域土壤各肥力指标等级处于中等以上,但有过半土壤综合肥力未达到平均水平。产区内部各镇土壤肥力也有着明显差异,大部分区域土壤肥力还有待提高;土壤水解酶活性变异系数较高,且与有机质、碱解氮、pH、有效磷等肥力因子有较强的相关性。
Spatial distribution characteristics of soil hydrolase activities and soil fertility evaluation of Carya cathayensis forests in Lin’an District
-
摘要:
目的 探索山核桃Carya cathayensis产区土壤水解酶活性空间分布特征和土壤肥力状况。 方法 在浙江省杭州市临安区山核桃主产区选取了259个样地,测定了土壤α-葡萄糖苷酶(AG)、β-葡萄糖苷酶(BG)、纤维二糖水解酶(CBH)、木糖苷酶(XYL)、亮氨酸氨基肽酶(LAP)、N-乙酰-β-氨基葡萄糖苷酶(NAG)、酸性磷酸酶(PHOS)等7种水解酶活性和主要的肥力指标,并运用主成分分析、地统计分析、相关性分析、冗余分析等方法,分析了临安区山核桃林地土壤中7种水解酶活性的空间异质性、影响因素以及土壤肥力状况。 结果 AG、BG、CBH、LAP、NAG、XYL、PHOS活性的块基比C0/(C+C0)分别为55%、42%、56%、49%、66%、47%、78%,全局莫兰指数(Ig)均大于0。土壤中碱解氮、有效磷、速效钾、有机质位于丰富等级的样地数分别占64%、56%、23%、45%,平均pH为5.76。58.7%的样地土壤肥力低于平均水平,仅32.7%的土壤肥力为Ⅰ、Ⅱ等级,大部分土壤肥力处于Ⅲ、Ⅳ等级 。 结论 7种土壤水解酶中,AG、BG、CBH、LAP、NAG、XYL具有中等空间自相关性,它们的变异情况受人为扰动和地形结构因素的共同影响。PHOS具有较弱的空间自相关,其活性空间分布主要受人为干扰的影响。7种水解酶活性均存在空间相关性,高低聚类情况相似。在岛石镇附近出现高值聚集,在清凉峰以及河桥、龙岗、昌化交界处附近出现低值聚集的情况,有机质、pH、碱解氮是影响水解酶活性高低值聚类的关键因素。土壤肥力指标分级和综合肥力得分结果表明:大部分林地土壤养分足以支撑山核桃林正常生长,但综合肥力还有待提高。图3表5参32 Abstract:Objective This study aims to explore the spatial distribution characteristics of soil hydrolase activity and soil fertility in Caya cathayensis forests. Method 259 sample plots were selected from the main C. cathayensis producing areas in Lin’an District of Hangzhou City, Zhejiang Province to determine the main fertility indicators and the activities of 7 hydrolases such as α-glucosidase (AG), β-glucosidase (BG), cellobiosidase (CBH), xylosidase (XYL), leucine amino peptidase (LAP), N-acetyl-glucosaminidase (NAG), and acid phosphatase (PHOS). Principal component analysis, geostatistical analysis, pearson correlation analysis, and redundancy analysis were used to analyze the soil fertility as well as the spatial variation of 7 hydrolase activities and their influencing factors. Result The spacial structure ratios [C0/(C+C0) ] of AG, BG, CBH, LAP, NAG, XYL, and PHOS were 55%, 42%, 56%, 49%, 66%, 47% and 78% respectively, and the global Morans’I (Ig) was greater than 0. Available N, available P, available K, and organic matter in the rich soil accounted for 64%, 56%, 23% and 45%, respectively, and the average pH was 5.76. The soil fertility of 58.7% of the sample plots was below average. Most of the C. cathayensis plots were in level Ⅲ and Ⅳ, while only 32.7% of the plots were in levelⅠand Ⅱ. Conclusion Among the 7 soil hydrolases, AG, BG, CBH, LAP, NAG, and XYL have moderate spatial autocorrelation, and their variation is jointly affected by human interference and topographic structure factors. PHOS has weak spatial autocorrelation, and its spatial distribution is mainly affected by human interference. The activities of 7 hydrolases have spatial correlations and similar high and low clustering. High value aggregation occurs near Daoshi town while low value clustering occurs near Qingliangfeng town and the boundary of Heqiao, Longgang and Changhua towns. Soil pH, organic matter and available N are the key factors affecting the high and low value clustering of hydrolase activity. The results of soil fertility index classification and comprehensive fertility score show that most soil nutrients are sufficient to support the normal growth of C. cathayensis forests, but the comprehensive fertility needs to be improved. [Ch, 3 fig. 5 tab. 32 ref.] -
Key words:
- Carya cathayensis /
- enzyme activity /
- spatial variation /
- high and low clustering /
- soil fertility
-
图 3 高低聚类点冗余分析结果
Figure 3 Redundancy analysis of high-high and low-low clustering points
表 1 土壤性质描述性统计分析
Table 1. Descriptive statistics of soil properties
项目 有机质/
(g·kg−1)有效磷/
(mg·kg−1)速效钾/
(mg·kg−1)碱解氮/
(mg·kg−1)pH AG/
(mol·g−1·h−1)最小值 5.41 0.52 22.06 28.62 4.50 0.12 最大值 98.08 22.43 466.07 192.53 7.48 1.67 平均值 37.39 4.23 113.77 132.40 5.76 0.35 标准差 15.38 3.90 72.58 43.47 0.59 0.30 变异系数/% 41.15 92.20 63.80 32.83 10.28 88.12 项目 BG/
(mol·g−1·h−1)CBH/
(mol·g−1·h−1)XYL/
(mol·g−1·h−1)LAP/
(mol·g−1·h−1)NAG/
(mol·g−1·h−1)PHOS/
(mol·g−1·h−1)最小值 4.58 0.04 0.17 0.10 0.15 18.31 最大值 192.62 63.06 62.91 32.95 93.60 1042.63 平均值 47.06 8.14 7.44 4.44 17.75 160.43 标准差 31.50 8.09 6.91 3.62 15.27 89.10 变异系数/% 66.95 99.48 92.98 81.54 86.07 55.54  下载: 导出CSV
下载: 导出CSV
表 2 土壤水解酶与养分因子及pH相关性分析表
Table 2. Correlation coefficients of soil hydrolase activities and soil nutrient factors and pH
水解酶 有机质 有效磷 速效钾 碱解氮 pH AG 0.355** 0.061 0.060 0.419** 0.102 BG 0.406** 0.172** 0.066 0.354** 0.147* CBH 0.356** 0.158* 0.060 0.275** 0.196** XYL 0.302** 0.088 −0.090 0.278** −0.283** LAP 0.170** 0.042 −0.015 0.230** −0.028 NAG 0.431** 0.267** 0.114 0.357** 0.109 PHOS 0.272** 0.123* 0.007 0.346** −0.286** 说明:*P<0.05, **P<0.01
下载: 导出CSV
表 3 土壤水解酶活性半方差函数理论模型及其相关参数
Table 3. Theoretical model of semi-variance function of soil hydrolase activities and its related parameters
水解酶 函数模型 块金值(C0) 基台值(C+C0) 块基比[C0/(C+C0)] 变程 决定系数 AG 球状模型 0.050 0.090 0.55 9.63 0.46 BG 指数模型 136.800 324.400 0.42 8.76 0.65 CBH 指数模型 0.140 0.250 0.56 1.60 0.43 XYL 高斯模型 0.080 0.170 0.47 2.12 0.37 LAP 高斯模型 10.970 21.960 0.49 11.20 0.73 NAG 高斯模型 197.150 294.260 0.66 27.30 0.45 PHOS 球状模型 0.032 0.041 0.78 14.60 0.54
下载: 导出CSV
表 4 山核桃土壤肥力指标丰缺等级及各等级占比
Table 4. Level of soil fertility indexs and the proportion of each level
项目 碱解氮 有效磷 速效钾 有机质 质量分数/(mg·kg−1) 占比/% 质量分数/(mg·kg−1) 占比/% 质量分数/(mg·kg−1) 占比/% 质量分数/(g·kg−1) 占比/% 缺乏 <80 6 <5 12 <80 34 <10 7 中等 80~120 30 5~10 32 80~110 43 10~40 48 丰富 >120 64 >10 56 >110 23 >40 45 说明:土壤肥力指标丰缺等级参考浙江省地方标准 DB33/T 2205—2019《山核桃分区施肥技术规范》
下载: 导出CSV
表 5 主成分贡献率与各因子得分
Table 5. Principal component contribution rates and each factor score
因子 主成分得分 第1主成分(48.39%) 第2主成分(26.50%) 第3主成分(17.12%) 有机质 0.144 0.268 0.225 有效磷 0.071 0.451 0.116 速效钾 0.027 0.481 0.046 碱解氮 0.061 0.040 0.652 AG 0.160 −0.058 0.039 BG 0.205 0.017 −0.299 CB 0.182 0.064 −0.393 XYL 0.177 −0.240 0.042 LAP 0.122 −0.238 0.345 NAG 0.179 0.111 −0.130 PHOS 0.184 −0.219 0.100
下载: 导出CSV
-
[1] 张璐璐, 贾桂民, 叶建丰, 等. 浙江临安山核桃干腐病发生发展规律[J]. 浙江农林大学学报, 2013, 30(1): 148 − 152. ZHANG Lulu, JIA Guimin, YE Jianfeng, et al. Frequency of Carya cathayensis canker disease in Lin’an City, Zhejiang Province [J]. J Zhejiang A&F Univ, 2013, 30(1): 148 − 152. [2] 吕惠进. 浙江临安山核桃立地环境研究[J]. 森林工程, 2005, 21(1): 1 − 3, 6. LÜ Huijin. The natural stands conditions of Carya Cathayensis Sarg. in Lin’an County of Zhejiang Province [J]. For Eng, 2005, 21(1): 1 − 3, 6. [3] 袁紫倩, 叶正钱, 李皓, 等. 影响山核桃林地土壤生产性能的主要肥力因子及其临界区间[J]. 植物营养与肥料学报, 2020, 26(1): 163 − 171. YUAN Ziqian, YE Zhengqian, LI Hao, et al. Main soil fertility factors and their critical ranges for Chinese walnut(Carya cathayensis Sarg. ) production [J]. J Plant Nutr Fert, 2020, 26(1): 163 − 171. [4] 丁立忠, 潘伟华, 马闪闪, 等. 测土配方施肥对临安山核桃生长和产量的影响[J]. 经济林研究, 2018, 36(4): 33 − 39. DING Lizhong, PAN Weihua, MA Shanshan, et al. Effects of testing soil for formulated fertilization on growth and yield in Carya cathayensis [J]. Nonwood For Res, 2018, 36(4): 33 − 39. [5] 张春苗, 张有珍, 姚芳, 等. 临安山核桃主产区土壤pH 值和有效养分的时空变化[J]. 浙江农林大学学报, 2011, 28(6): 845 − 849. ZHANG Chunmiao, ZHANG Youzhen, YAO Fang, et al. Temporal and spatial variation of soil pH and nutrient availability for Carya cathayensis orchards in Lin’an [J]. J Zhejiang A&F Univ, 2011, 28(6): 845 − 849. [6] 杨惠思, 赵科理, 叶正钱, 等. 山核桃品质对产地土壤养分的空间响应[J]. 植物营养与肥料学报, 2019, 25(10): 1752 − 1762. YANG Huisi, ZHAO Keli, YE Zhengqian, et al. Spatial response of Carya cathayensis quality to soil nutrients [J]. J Plant Nutr Fert, 2019, 25(10): 1752 − 1762. [7] 张红桔, 马闪闪, 赵科理, 等. 山核桃林地土壤肥力状况及其空间分布特征[J]. 浙江农林大学学报, 2018, 35(4): 664 − 673. ZHANG Hongju, MA Shanshan, ZHAO Keli, et al. Soil fertility and its spatial distributionfor Carya cathayensis stands in Lin’an, Zhejiang Province [J]. J Zhejiang A&F Univ, 2018, 35(4): 664 − 673. [8] 丁立忠, 金锦, 张智勇, 等. 临安不同产区镇山核桃林地土壤肥力水平的变化研究[J]. 浙江林业科技, 2020, 40(3): 45 − 50. DING Lizhong, JIN Jin, ZHANG Zhiyong, et al. Changes of soil fertility in Carya cathayensisstands in major production towns of Lin’an City [J]. J Zhejiang For Sci Technol, 2020, 40(3): 45 − 50. [9] GRANDY S A, NEFF J C, WEINTRAU M N. Carbon structure and enzyme activities in alpine and forest ecosystems [J]. Soil Biol Biochem, 2007, 39(11): 2701 − 2711. [10] 莫雪, 陈斐杰, 游冲, 等. 黄河三角洲不同植物群落土壤酶活性特征及影响因子分析[J]. 环境科学, 2021, 41(2): 896 − 904. MO Xue, CHEN Feijie, YOU Chong, et al. Characteristics and factors of soil enzyme activity for different plant communities in Yellow River Delta [J]. Environ Sci, 2021, 41(2): 896 − 904. [11] 祝小祥, 徐祖祥, 徐进, 等. 临安山核桃主产区土壤理化性状变化的研究[J]. 农学学报, 2014, 4(6): 32 − 35, 40. ZHU Xiaoxiang, XU Zuxiang, XU Jin, et al. A study on the change in soil physical and chemical propertiesin the Chinese hickory production area of Lin’an [J]. J Agric, 2014, 4(6): 32 − 35, 40. [12] SAIYA-CORK K R, SINSABAUGH R L, ZAK D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil [J]. Soil Biol Biochem, 2002, 34(9): 1309 − 1315. [13] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2002. LU Rukun. The Analysis Method of Soil Agricultural Chemistry [M]. Beijing: China Agricultural Science and Technology Press, 2000. [14] WEBSTER R, OLIVER M A. Geostatistics for Environmental Scientists[M]. Totnes: John Wiley & Sons, 2007. [15] 史舟, 李艳. 地统计学在土壤学中的应用[M]. 北京: 中国农业出版社, 2006. SHI Zhou, LI Yan. Application of Geostatistics in Soil Science [M]. Beijing: China Agricultural Press, 2006. [16] KATSALIROU E, SHIPING D, NOFNIGER D L, et al. Spatial structure of microbial biomass and activity in prairie soil ecosystems [J]. Eur J Soil Biol, 2010, 46: 181 − 189. [17] ZHAO Keli, FU Weijun, QIU Qiaozhen, et al. Spatial patterns of potentially hazardous metals in paddy soils in a typical electrical waste dismantling area and their pollution characteristics [J]. Geoderma, 2019, 337(3): 453 − 462. [18] 王强, 郑梦蕾, 叶治山, 等. 基于Moran’s I的菜地土壤属性空间分布格局分析[J]. 农业环境科学学报, 2020, 39(10): 2297 − 2306. WANG Qiang, ZHENG Menglei, YE Zhishan, et al. Analysis of spatial distribution pattern of vegetable soil properties based on Moran’s I [J]. J Agro-Environ Sci, 2020, 39(10): 2297 − 2306. [19] 陶吉兴, 傅伟军, 姜培坤, 等. 基于Moran’s I和地统计学的浙江森林土壤有机碳空间分布研究[J]. 南京林业大学学报(自然科学版), 2014, 38(5): 97 − 101. TAO Jixing, FU Weijun, JIANG Peikun, et al. Using Moran’s I and geostatistics to analyze the spatial distribution of organic carbon in forest soil of Zhejiang Province [J]. J Nanjing For UnivNat Sci Ed, 2014, 38(5): 97 − 101. [20] 黄进, 陈金华, 张方敏. 基于主成分分析的安徽省冬小麦气候灾损风险的时空演变[J]. 应用生态学报, 2021, 32(9): 3185 − 3194. HUANG Jin, CHEN Jinhua, ZHANG Fangmin. Spatio-temporal evolution of climate-induced reduction risk for winter wheat in Anhui Province based on principal component analysis. [J]. Chin J Appl Ecol, 2021, 32(9): 3185 − 3194. [21] 赵瑞芬, 程滨, 滑小赞, 等. 基于主成分分析的山西省核桃主产区土壤肥力评价[J]. 山西农业大学学报(自然科学版), 2020, 40(6): 61 − 68. ZHAO Ruifen, CHENG Bin, HUA Xiaozan, et al. Evaluation of soil fertility status in main walnut production areas in Shanxi based on principal component analysis [J]. J Shanxi Agric Univ Nat Sci Ed, 2020, 40(6): 61 − 68. [22] ZHANG Xinyi, SUI Yueyu, ZHANG Xudong, et al. Spatial variability of fertility properties in black soil of Northeast China [J]. Pedosphere, 2007, 17(1): 19 − 29. [23] TRASAR-CEPEDA C, LEIRÓS M C, GIL-SOTRES F. Hydrolytic enzyme activities in agricultural and forest soils: some implications for their use as indicators of soil quality [J]. Soil Biol Biochem, 2008, 40(9): 2146 − 2155. [24] 张贾宇, 佘婷, 鄂晓伟, 等. 杨树人工林幼林阶段林下植被管理对土壤微生物生物量碳、氮及酶活性的影响[J]. 生态学报, 2021, 41(24): 9898 − 9909. ZHANG Jiayu, SHE Ting, E Xiaowei, et al. Effects of understory vegetation management on soil microbial biomass carbon and nitrogen and extracellular enzyme activities in the early stages of poplar plantation [J]. Acta Ecol Sin, 2021, 41(24): 9898 − 9909. [25] 万忠梅, 吴景贵. 土壤酶活性影响因子研究进展[J]. 西北农林科技大学学报(自然科学版), 2005, 33(6): 87 − 92. WAN Zhongmei, WU Jinggui. Study progress on factors affecting soil enzyme activity [J]. J Northwest A&F Univ Nat Sci Ed, 2005, 33(6): 87 − 92. [26] 刘烁, 王秋兵, 史文娇, 等. 喀斯特典型集水区土壤水解酶活性空间异质性及其影响因素[J]. 应用生态学报, 2018, 29(8): 2615 − 2623. LIU Shuo, WANG Qiubing, SHI Wenjiao, et al. Spatial heterogeneity of soil hydrolase activities and their influencing factors in a typical Karst catchment of Guizhou Province, China [J]. Chin J Appl Ecol, 2018, 29(8): 2615 − 2623. [27] 刘霜, 张心昱. 不同植物根际土壤碳氮水解酶活性热点区的空间分布特征[J]. 生态学报, 2020, 40(13): 4462 − 4469. LIU Shuang, ZHANG Xinyu. Spatial distribution of carbon and nitrogen acquiring hydrolase activity hotspots in rhizosphere soils of different plants [J]. Acta Ecol Sin, 2020, 40(13): 4462 − 4469. [28] 白尚斌, 张彦东, 王政权. 落叶松根际 pH 值与供磷水平及土壤磷有效性的关系[J]. 林业科学, 2001, 37(4): 130 − 133. BAI Shangbin, ZHANG Yandong, WANG Zhengquan. The relationship between pH changes and P-availability in rhizosphere of Larix gmelinii [J]. Sci Silv Sin, 2001, 37(4): 130 − 133. [29] 王政权, 王庆成. 森林土壤物理性质的空间异质性研究[J]. 生态学报, 2000, 20(6): 945 − 950. WANG Zhengquan, WANG Qingcheng. The spatial heterogeneity of soil physical properties in forests [J]. Acta Ecol Sin, 2000, 20(6): 945 − 950. [30] 张红桔, 赵科理, 叶正钱, 等. 典型山核桃产区土壤重金属空间异质性及其风险评价[J]. 环境科学, 2018, 39(6): 2893 − 2903. ZHANG Hongju, ZHAO Keli, YE Zhengqian, et al. Spatial variation of heavy metals in soils and its ecological risk evaluation in a typical Carya cathayensis production area [J]. Environ Sci, 2018, 39(6): 2893 − 2903. [31] 蒲生彦, 王宇, 陈文英, 等. 植物根际土壤酶对重金属污染的响应机制研究综述[J]. 生态毒理学报, 2020, 15(4): 11 − 20. PU Shengyan, WANG Yu, CHEN Wenying, et al. Review on the mechanism of plant rhizosphere soil enzyme response to heavy metal pollution [J]. Asian J Ecotoxicol, 2020, 15(4): 11 − 20. [32] 沈一凡, 钱进芳, 郑小平, 等. 山核桃中心产区林地土壤肥力的时空变化特征[J]. 林业科学, 2016, 52(7): 1 − 12. SHEN Yifan, QIAN Jinfang, ZHENG Xiaoping, et al. Spatial-temporal variation of soil fertility in Chinese walnut (Carya cathayensis) plantation [J]. Sci Silv Sin, 2016, 52(7): 1 − 12. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20210417
 点击查看大图
点击查看大图
计量
- 文章访问数: 1881
- HTML全文浏览量: 442
- PDF下载量: 126
- 被引次数: 0






