





-
桑Morus是一种重要的木本植物,具有28条染色体[1],不仅可以作为家蚕的重要食物,还具有一定的经济、药用及生态价值。但是桑葚在成熟的过程中易脱落,使得桑葚变软变黑,影响其产业价值,因此,如何采用有效的手段对桑葚脱落进行调控十分重要。
果实脱落是植物正常发育过程中的一种常见现象,但是,在一定程度上会限制果树的产量。果实脱落主要受环境因素、酶、生理代谢共同调控[2],其中,环境因素主要包括生物胁迫(如病虫害[3])和非生物胁迫(温度胁迫[4]、水分胁迫[5]、光胁迫[6]等);生理代谢则包括一些激素代谢(如脱落酸[7]、乙烯[8]等植物激素)和糖代谢[9];由于纤维素和果胶是植物细胞壁的主要组成成分,因此纤维素酶、果胶酶、多聚半乳糖醛酸酶[10]等与果实脱落相关[11]。
目前,桑葚脱落的分子机制还未见报道。本研究以白桑Morus alba为供试材料,对其进行转录组测序分析,探究桑葚在脱落过程中的生化成分代谢的分子机制,旨在为进一步了解桑葚果实脱落的分子机制提供参考。
-
2021年4月在西南大学家蚕基因组生物学国家重点实验室桑树资源种质基地,选取无病害、健康状况较好的白桑幼果果柄。以落果果柄 (YD) 为实验组,取6个果柄为1份,设3个生物学重复,标记为YD1、YD2、YD3。正常果果柄 (YN) 为对照组,同样取6个果柄为1份,设3个生物学重复,标记为YN1、YN2、YN3。取材后立即在液氮中速冻并储存于−80 ℃冰箱保存。
-
将正常果和脱落果果柄组织清洗后用质量分数为4%的多聚甲醛过夜固定,用包埋剂包埋材料。待包埋剂彻底凝固后进行冷冻切片。切片厚度为5 μm,将切片后的材料吸附于阳离子载玻片上,室温干燥30 min后,用磷酸缓冲盐溶液(PBS)洗涤样品,去除包埋剂后置于显微镜下观察。
-
采用TRIzol法提取落果果柄和正常果果柄的总RNA,用分光光度计检测RNA样品的浓度和纯度,以保证是否可以进行下一步测序分析。
-
样品由华大基因进行测序及分析,利用华大智造测序平台BGISEQ测序,所得的原始数据为raw reads。过滤掉低质量、接头污染以及未知碱基N含量过高的reads,过滤后的数据称为clean reads。将clean reads比对到参考基因组上,再进行后续分析。
-
将测序所得原始数据提交到美国国家生物技术信息中心(NCBI)的SRA数据库中,检索号为: PRJNA811258。
-
使用DEseq2方法检测样品之间的差异表达基因(DEG)[12]。根据基因本体论数据库(GO)和京都基因与基金组百科全书(KEGG)注释结果以及官方分类,将差异基因进行功能分类,同时使用R软件中的phyper函数进行富集分析。详细说明见Wiki网站
。校正后的P≤0.05为显著富集。 -
将桑葚幼果的落果果柄与正常果果柄RNA用PrimesciptTM RT reagent kit with gDNA Eraser (Takara)反转录,在primer premier 5.0软件设计定量引物(表1),内参基因为 Actin,利用NovoStart® SYBR qPCR SuperMix Plus(novoprotein) 试剂盒进行荧光定量PCR。反应程序为:95 ℃ 30 s,95 ℃ 3 s,60 ℃ 30 s,共40个循环。用2−ΔΔCt算法处理数据,利用GraphPad Prism 8.0.2软件作图。
表 1 转录组数据RT-qPCR验证引物序列
Table 1. Primer sequences used in RT-qPCR validation of transcriptome data
基因ID 基因名 正向引物(5′→3′) 反向引物(5′→3′) XM_010096892.1 GH3.6 ACACTAACTACACCAGCCCAAA ACTTAATAGCACGAATGAACCC XM_010102492.2 bHLH78 TATTACCTTCGTCGTCCCCTCCTA ACCACTCTTTTCGTTTCCTTCACC XM_010113436.1 SAUR15 TGAAGAAGCCGAGAAGGAGTA GGTGGTAGGAGAAGGGATAAC XM_024169608.1 PHO1 GCCAATAACGACAGGAAA AACAACCCGTGAACAAAC -
桑果是带果柄脱落,为了研究桑树正常果柄和脱落果柄的差异,对其进行冷冻切片并观察显微结构。从图1可知:正常幼果果柄与枝干相连的区域内细胞规则,形态一致,而脱落幼果果柄与枝干相连处细胞小且致密。

图 1 落果与正常果果柄纵切面显微结构图
Figure 1. Longitudinal section microscopic structures of abscised and normal surviving young fruit peduncles
-
基于BGISEQ对桑葚幼果的落果与正常果果柄离区进行转录组测序,6个样品获得262.92 Mb的原始序列。原始测序数据经一系列质量控制后,每个样本获得超过6.3 Gb的高质量纯净测序数据量,每项碱基质量大于20的碱基数量占总碱基数量的比例 (Q20) 均大于96%,每项碱基质量大于30 的碱基数量占总碱基数量的比例 (Q30) 均大于91%。将每个样本的高质量纯净序列与测序并组装的桑树基因组序列进行比对,比对率均高于59%(表2),表明本研究转录组测序数据质量较高,可用于后续的分析。
表 2 测序数据统计
Table 2. Statistics of sequencing data
样本
名称原始序列
数/Mb纯净序列
数/Mb纯净碱基
数/GbQ20/% Q30/% YD1 43.82 42.48 6.37 96.50 91.31 YD2 43.82 42.74 6.41 96.53 91.34 YD3 43.82 42.58 6.39 96.53 91.39 YN1 43.82 42.61 6.39 96.46 91.19 YN2 43.82 42.58 6.39 96.55 91.38 YN3 43.82 42.65 6.40 96.55 91.39 -
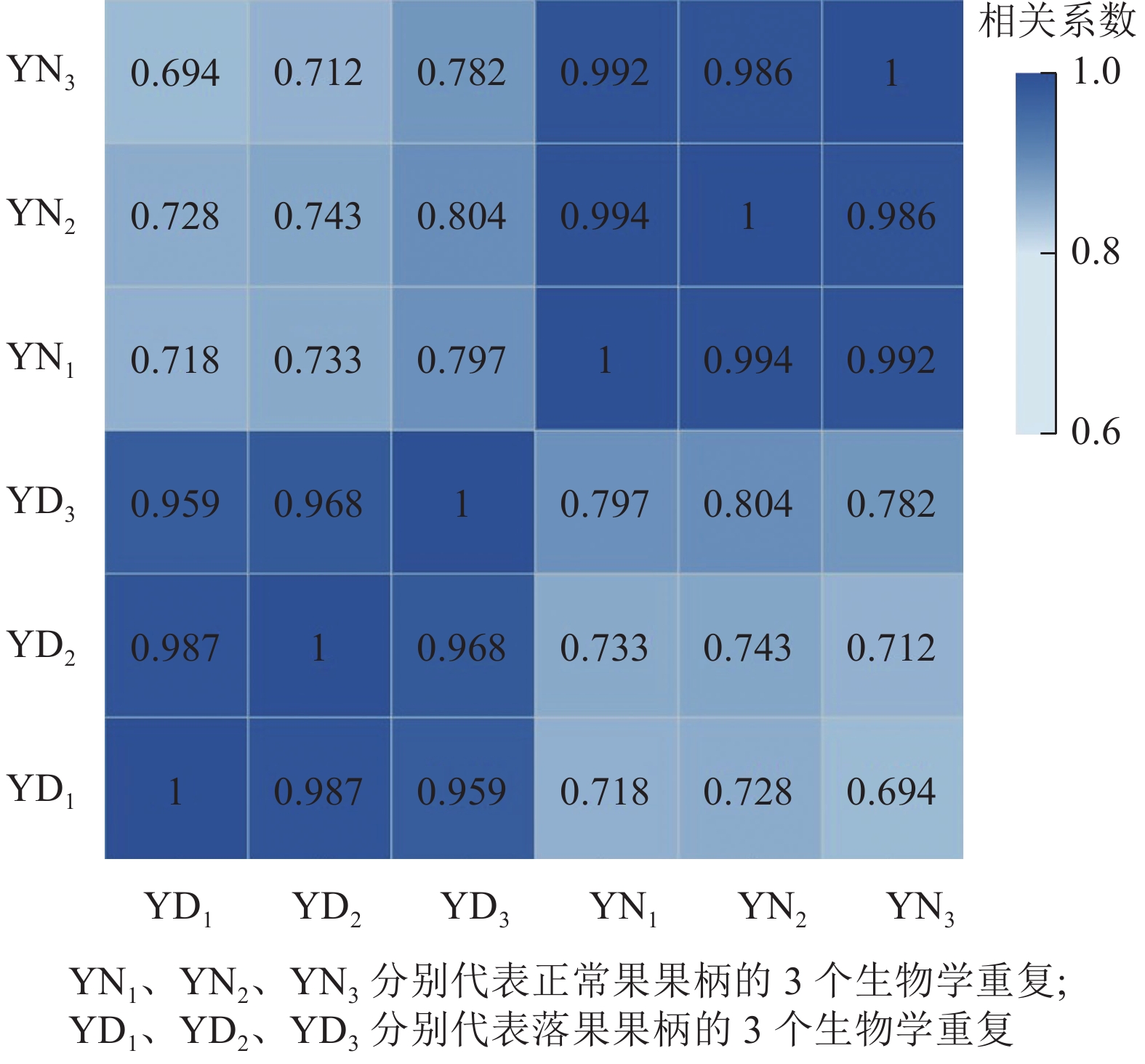
用每2个样品之间的Pearson相关系数以反映样本间基因表达的相关性(图2)。结果发现:Pearson相关系数为0.69~1.00,说明各样本间重复性和相关性较好。
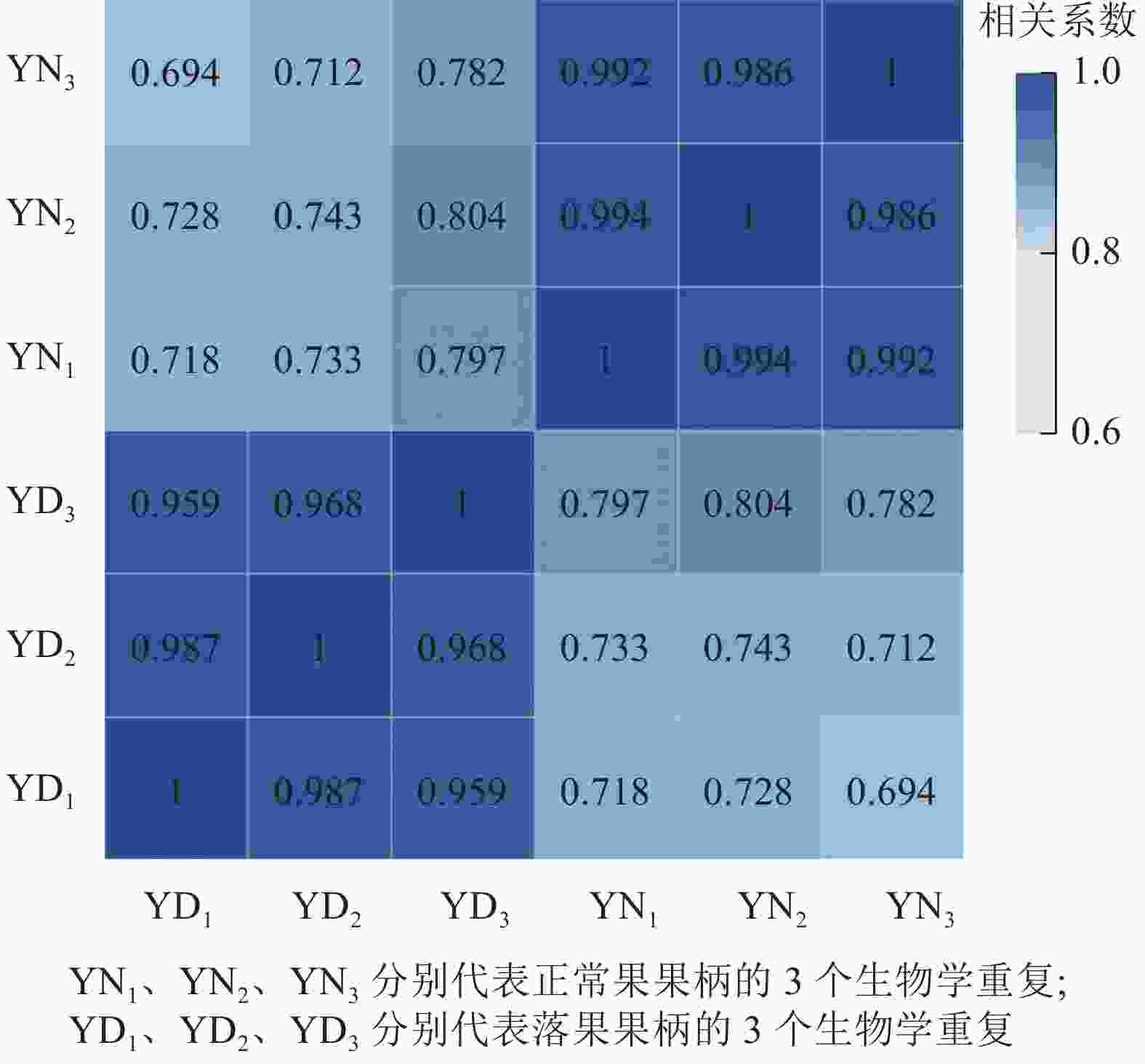
图 2 落果与正常果果柄的相关性热图
Figure 2. Correlation heatmap of abscised and normal surviving young fruit peduncles
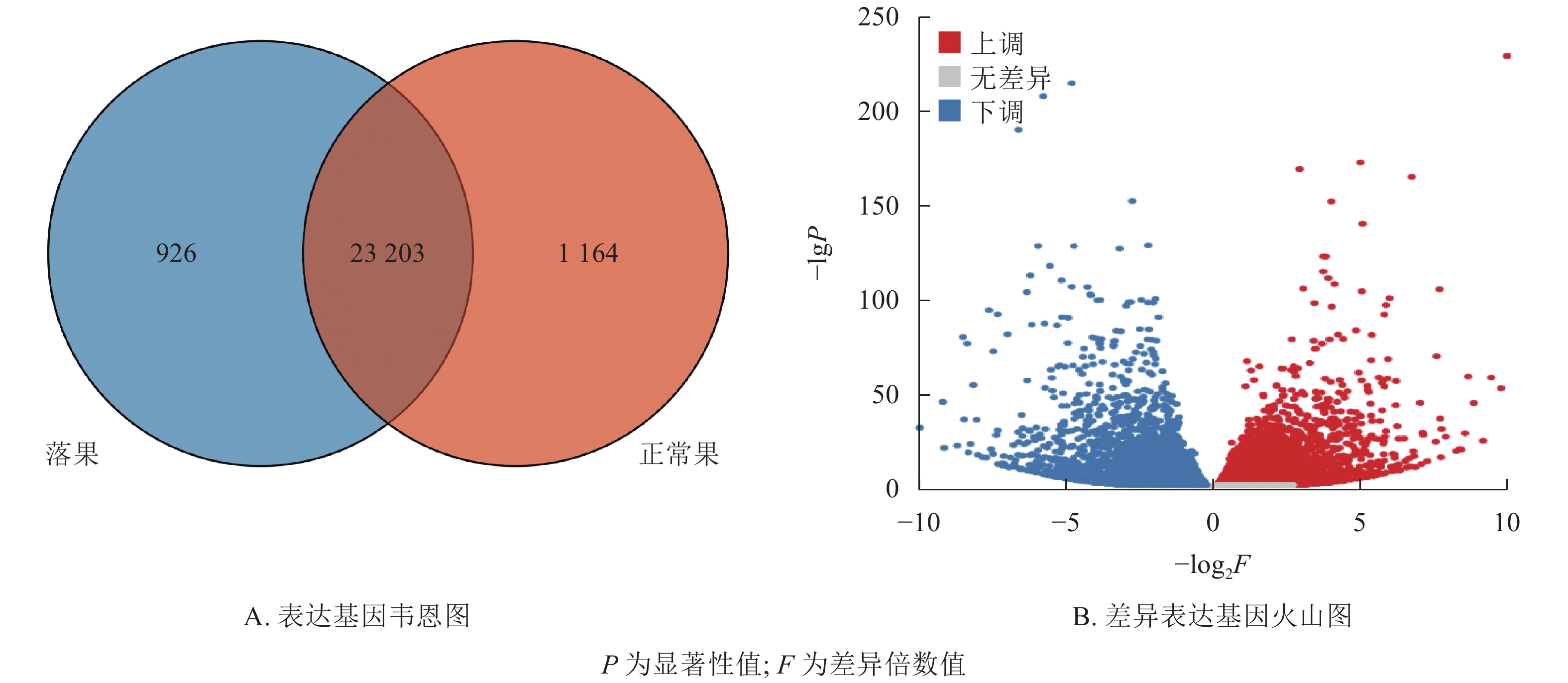
在桑葚幼果果柄组织中,共鉴定到25 293个基因(图3A)。其中,共表达的基因有23 203个,占91.7%。正常果果柄特有基因为1 164个,占4.6%,落果果柄组特有基因为926个,占3.7%。由此可见,落果果柄基因较正常果果柄基因少。根据差异筛选标准,在2组中共筛选到10 481个差异表达基因(图3B),其中,5 239个差异表达基因上调表达,5 242个差异表达基因下调表达。

图 3 落果与正常果果柄的表达基因和差异表达基因
Figure 3. Expressed genes and differentially expressed genes of abscised and normal surviving young fruit peduncles
-
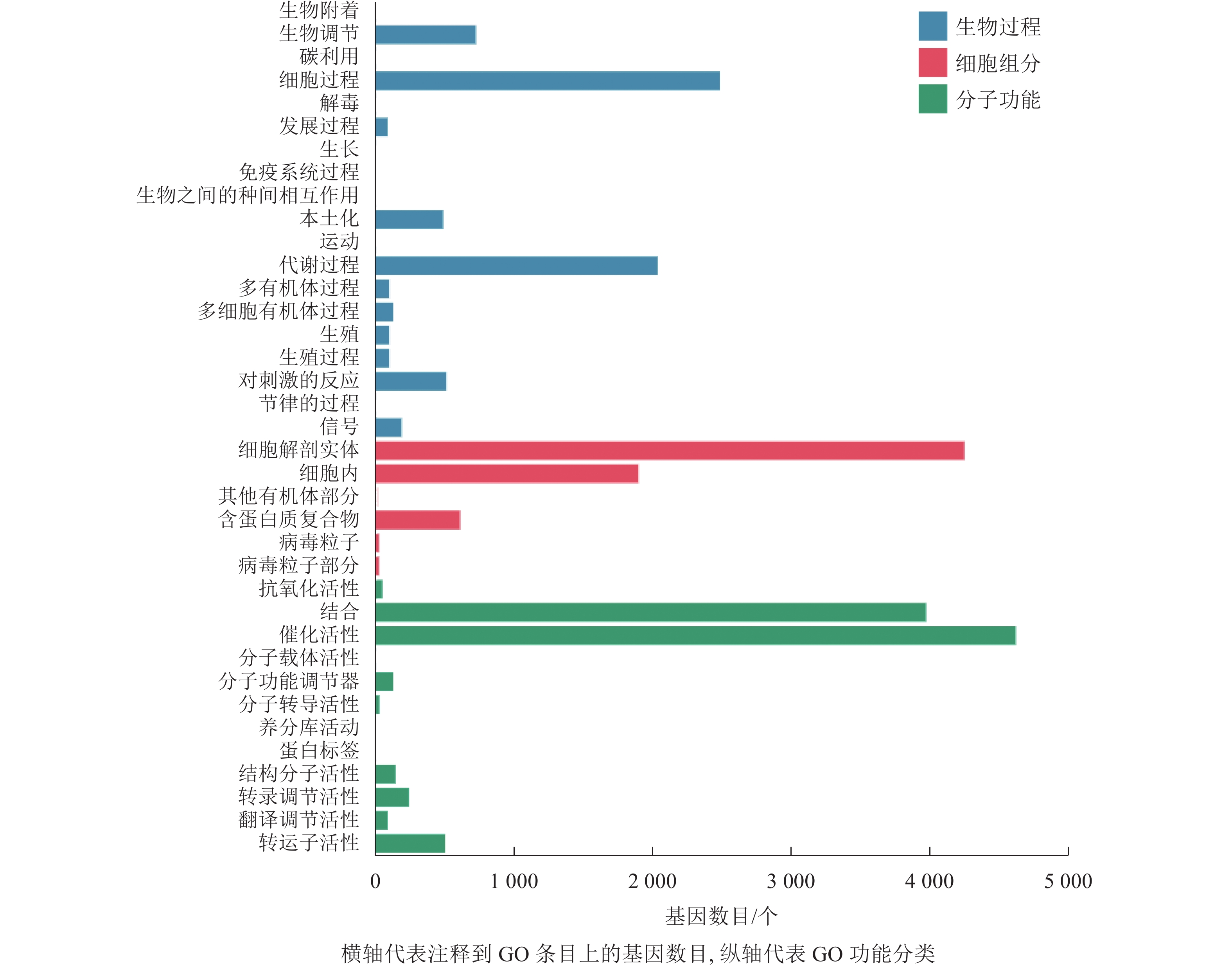
本研究对差异表达基因进行GO功能富集分析,共发现37个显著性GO条目。生物过程共富集到7 064个基因,显著富集到19个条目,涉及生物调节、细胞过程、代谢过程、对刺激的反应等,其中富集到生物过程中最多的差异表达基因为细胞过程和代谢过程基因;细胞组分共富集到6 857个基因,显著富集到6个条目,涉及细胞解剖实体、细胞内、其他有机部分、含蛋白质复合物、病毒粒子等,富集到细胞组分最多的差异表达基因为细胞解剖实体和细胞内基因;分子功能共富集到9 842个基因,显著富集到12个条目,涉及催化活性、转运蛋白活性、转录调节活性、分子功能调节剂等,富集到分子功能最多的差异表达基因为催化活性和结合基因(图4)。
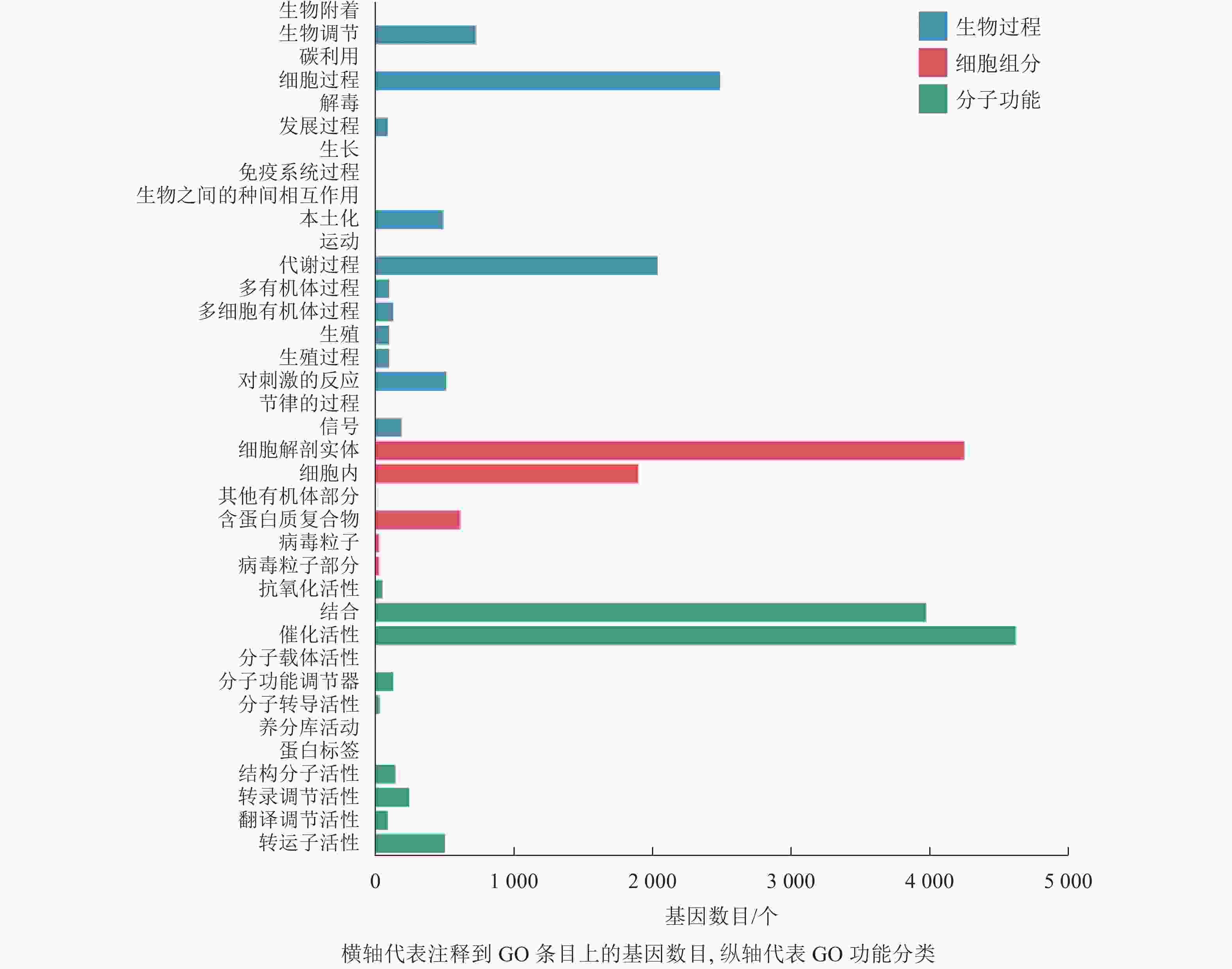
图 4 落果与正常果果柄的差异表达基因GO分类柱状图
Figure 4. Histogram of GO classification of differentially expressed genes of abscised and normal surviving young fruit peduncles
GO富集分析结果显示:大多数差异基因集中在催化活性、膜的组成成分、氧化还原酶活性、膜的内在成分,分别为4 623、2 774、777、1 781个基因(图5)。其中,具催化活性的基因最多,说明在桑葚的落果过程中可能有很多重要的酶参与,从而发生一系列的催化反应。

图 5 落果与正常果果柄的差异表达基因GO富集气泡图
Figure 5. GO enrichment bubble chart of differentially expressed genes of abscised and normal surviving young fruit peduncles
-
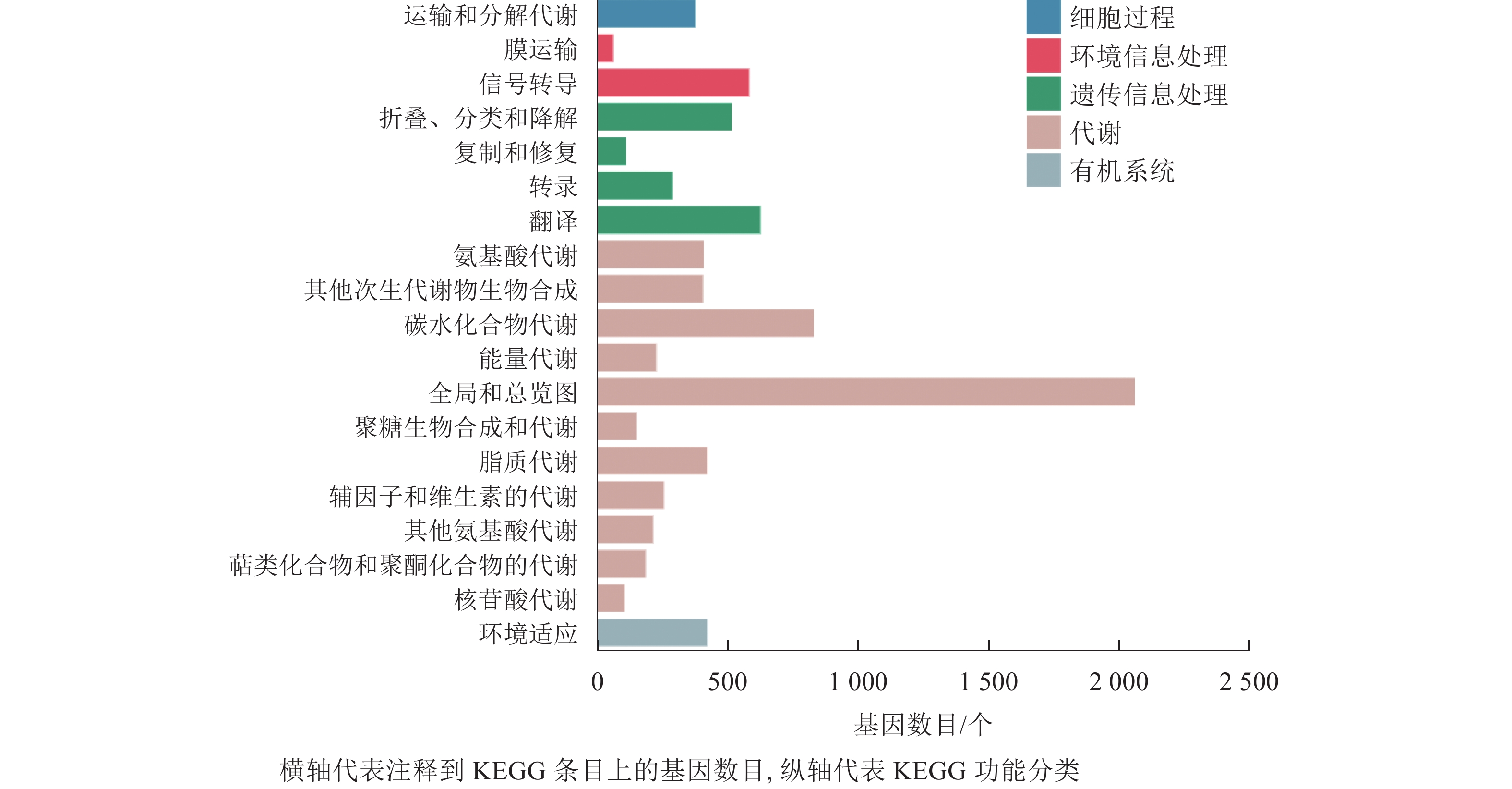
在本研究中,细胞过程富集的基因为380个,富集到1个条目,为运输和分解代谢;环境信息处理共富集到653个基因,富集到2个条目,为膜运输和信号转导;遗传信息处理共富集到1 558个基因,包含4个条目,为折叠分类和降解、复制和修复、转录、翻译;代谢通路富集到5 315个基因,包含11个条目,为氨基酸代谢、其他次生代谢物的生物合成、碳水化合物代谢、能量代谢等;有机系统共富集到427个基因,含1个条目(图6)。分析发现:代谢通路富集的基因最多,说明桑葚在脱落过程中代谢反应尤为明显。
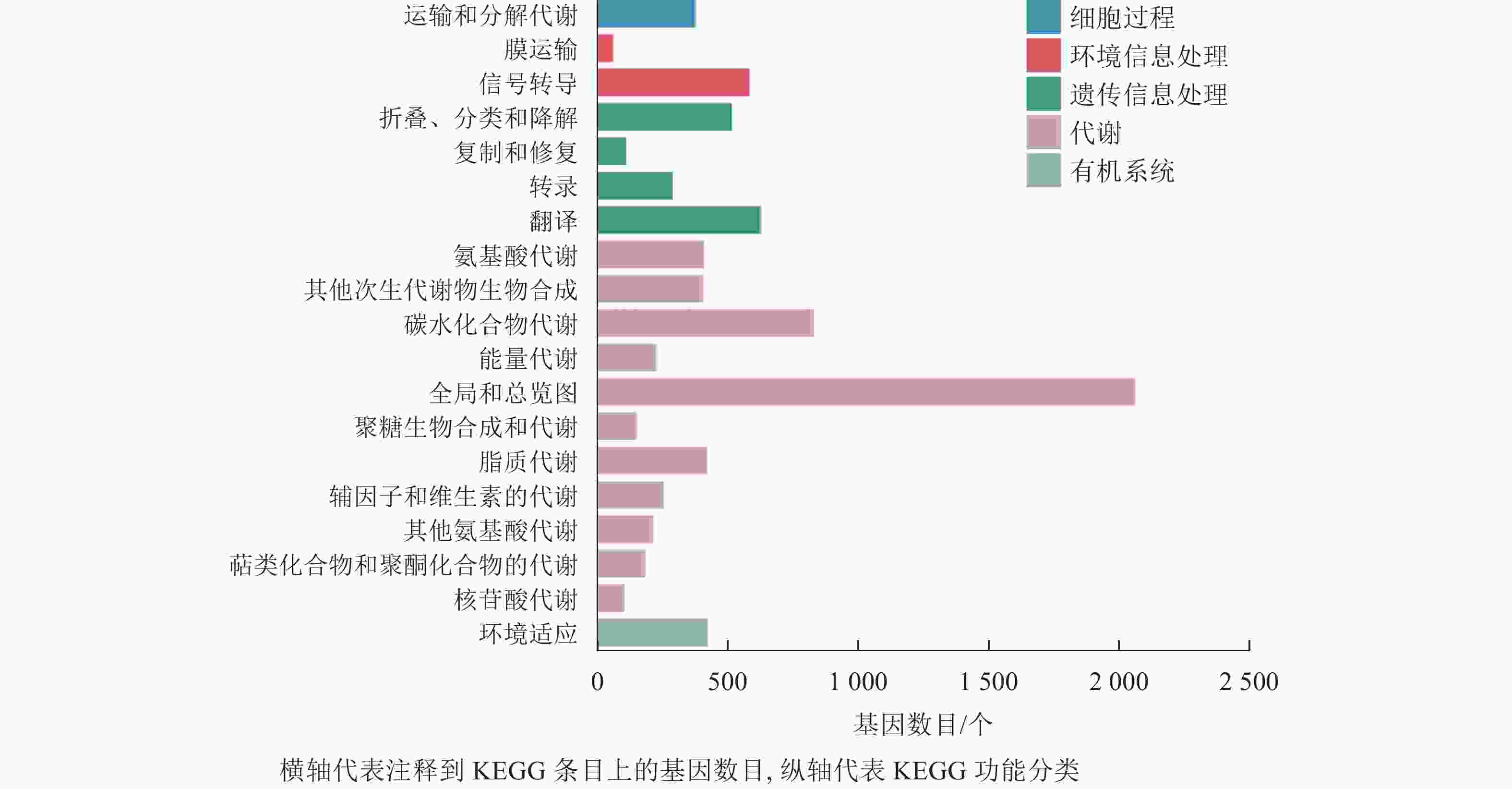
图 6 落果与正常果果柄的差异表达基因KEGG分类柱状图
Figure 6. Histogram of KEGG classification of differentially expressed genes of abscised and normal surviving young fruit peduncles
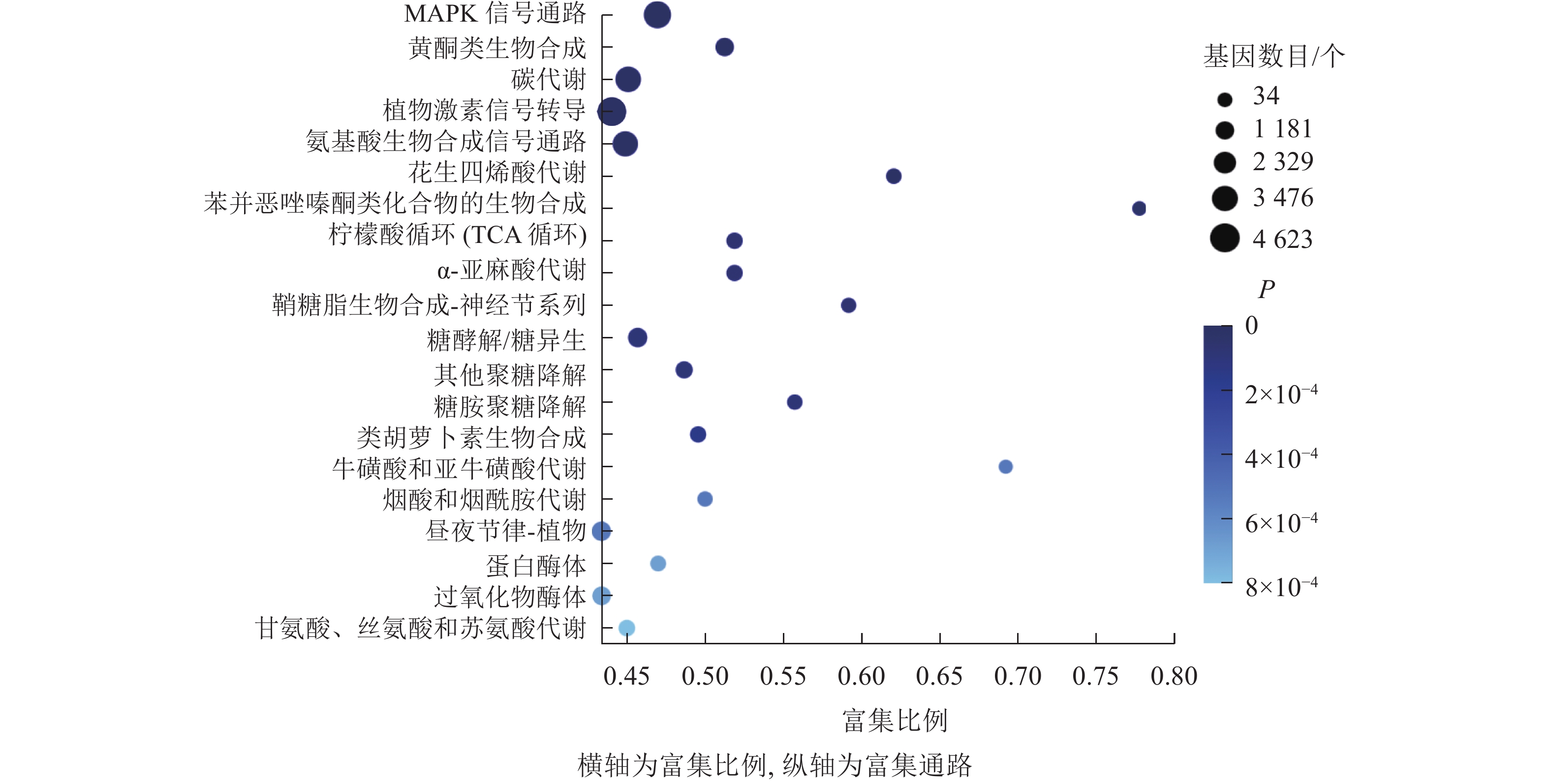
KEGG通路富集分析结果显示:大多数差异表达基因集中在MAPK信号通路、黄酮类生物合成、柠檬酸循环、植物激素信号转导、氨基酸的生物合成通路,分别含293、102、55、318、251个基因(图7)。其中,植物激素信号转导途径的差异表达基因数量最多,说明在桑葚果实脱落过程中植物激素起到了十分重要的作用。
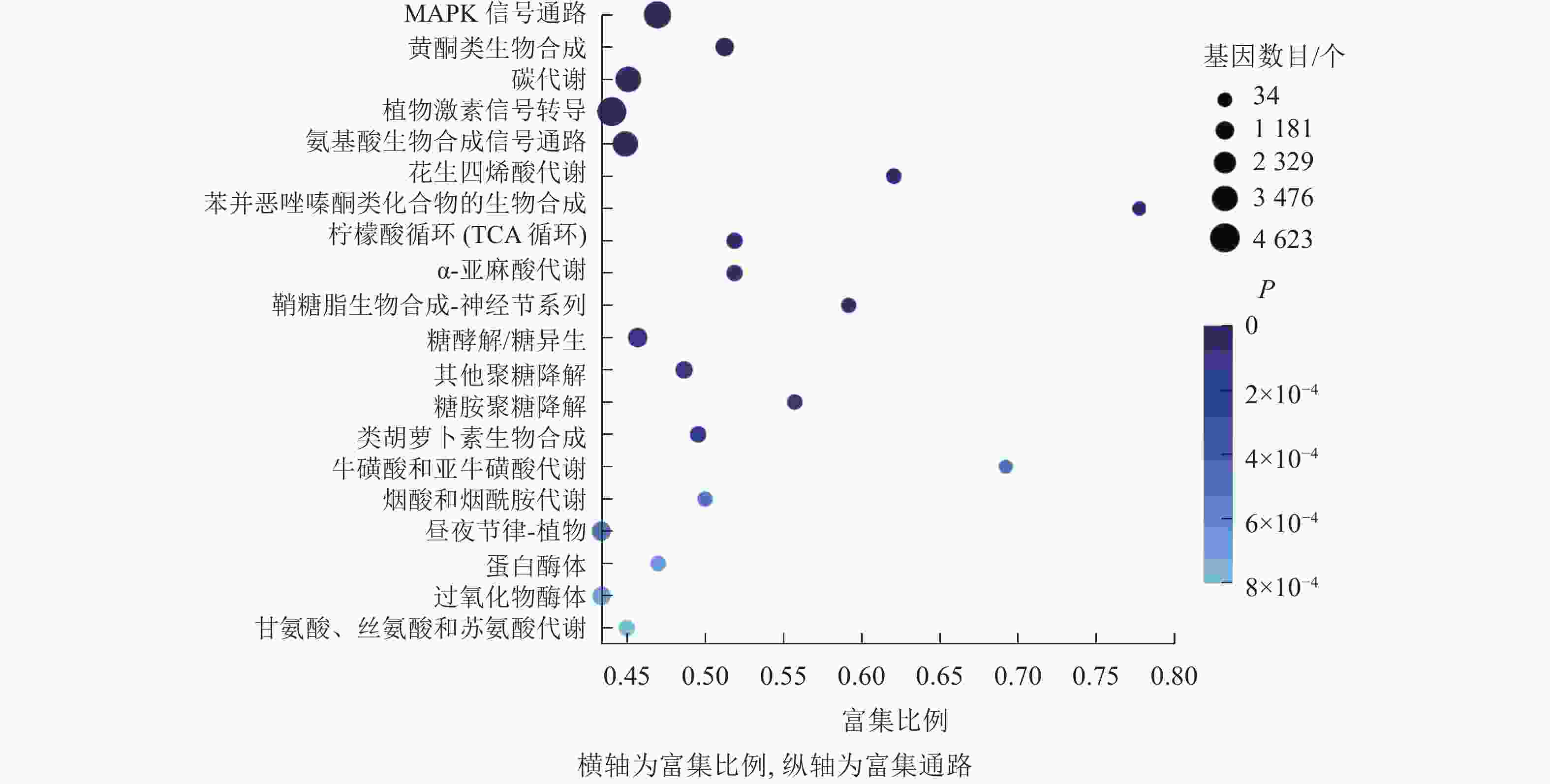
图 7 落果与正常果果柄的差异表达基因的KEGG富集气泡图
Figure 7. KEGG enrichment bubble diagram of differentially expressed genes of abscised and normal surviving young fruit peduncles
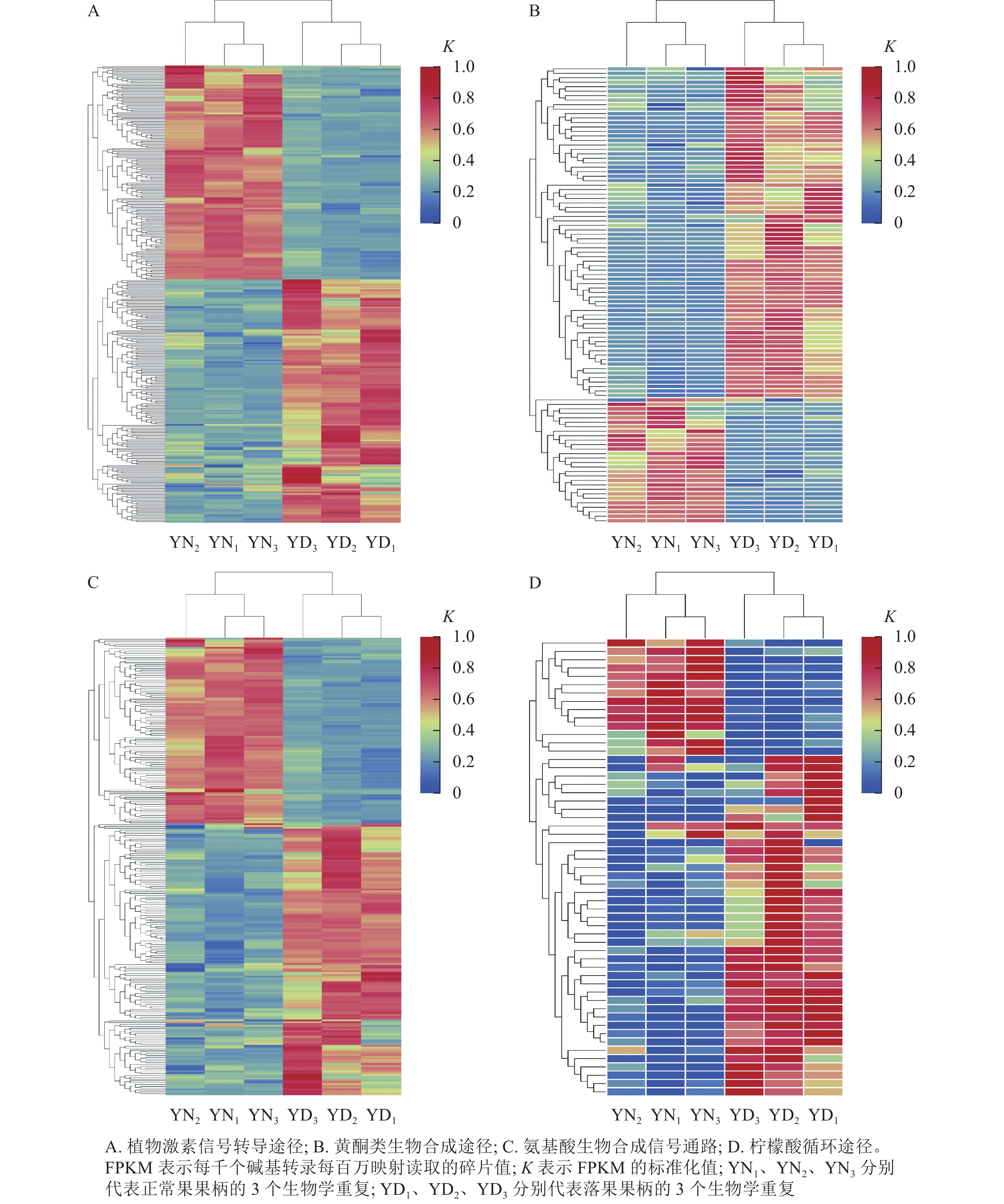
在桑树的代谢通路中,植物激素信号转导途径的差异表达基因数量最多,为318个基因,其中,落果果柄中有51.9% (156个)的差异表达基因的表达量高于正常果果柄(图8A);在黄酮类生物合成途径中,有102个差异基因富集,其中有73.5% (75个)的差异基因在落果果柄中表达量高(图8B);在氨基酸生物合成通路中,共有251个差异表达基因富集,其中有59.8% (150个)的差异表达基因在落果果柄中表达量更高(图8C);在柠檬酸循环中,有55个差异表达基因富集,其中有74.5% (41个)的差异表达基因在落果果柄中表达量更高(图8D)。不难发现,落果果柄有超过一半的差异表达基因表达量高于正常果果柄。说明植物激素、黄酮类等次生代谢物以及柠檬酸等物质在桑葚脱落过程中发挥了重要的作用。
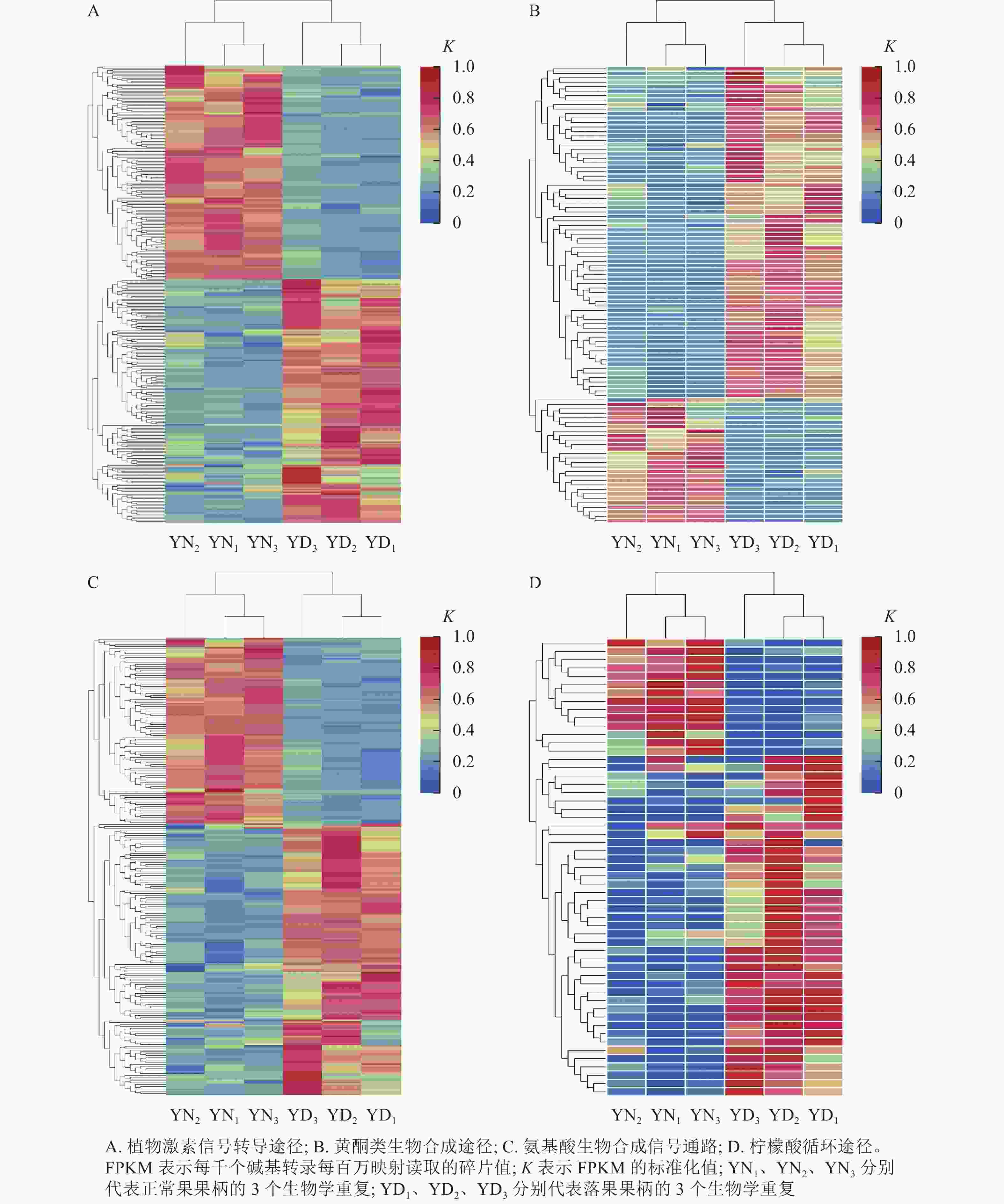
图 8 落果与正常果果柄的代谢途径的差异表达基因热图
Figure 8. Heat map of differentially expressed genes in metabolic pathways of abscised and normal surviving carpopodium
-
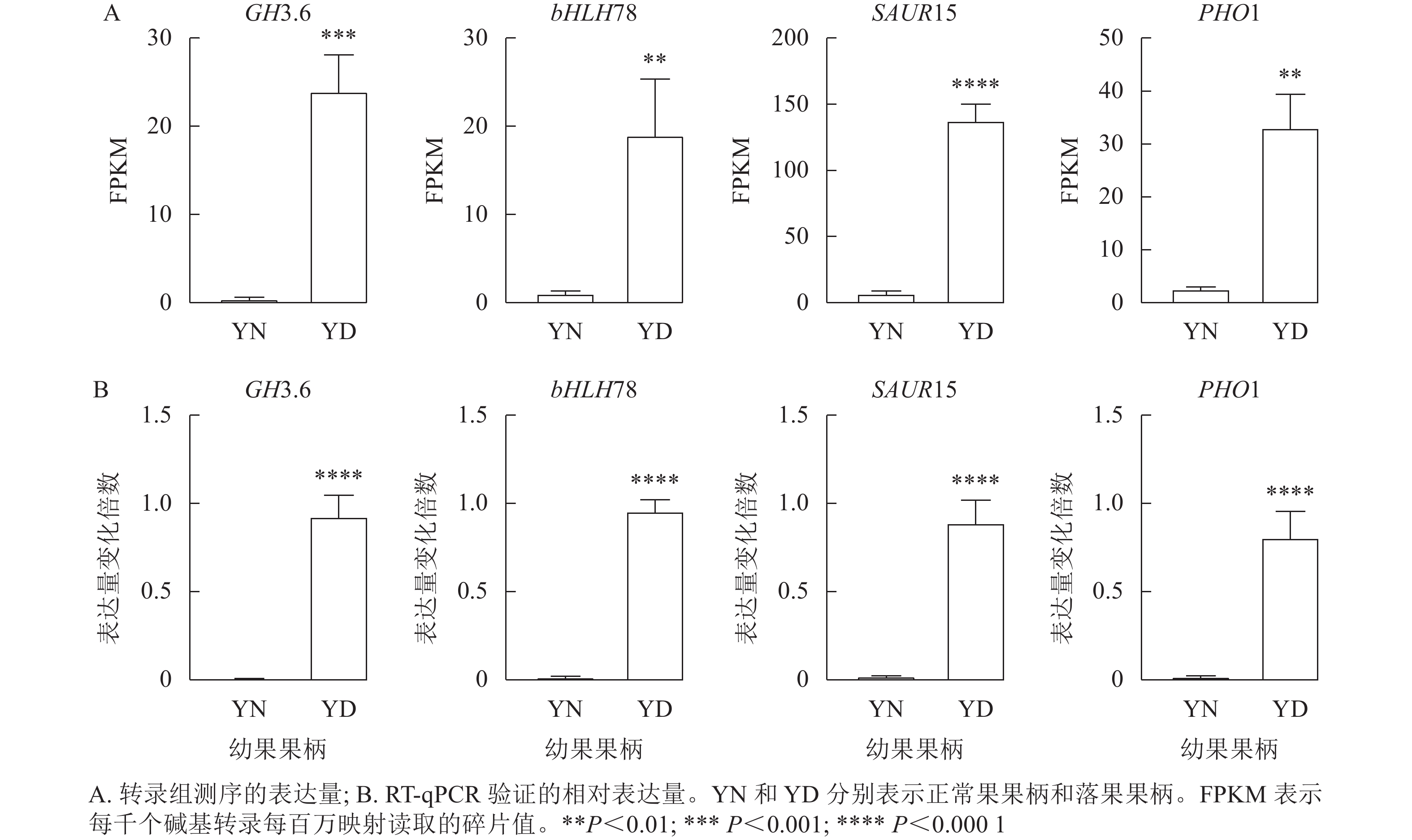
由于植物激素对于落花落果以及保花保果都具有十分重要的作用,于是在植物激素信号转导途径中筛选4个差异十分显著的基因,分别为GH3.6 (XM_010096892.1)、bHLH78 (XM_010102492.2)、SAUR15 (XM_010113436.1)、PHO1 (XM_024169608.1)。将这4个基因的每千个碱基转录每百万映射读取的碎片值(FPKM)绘制直方图,GH3.6在落果组中的表达水平是正常果组的20多倍,bHLH78在落果组中的表达水平约为正常果组20倍,PHO1(磷转运蛋白)在落果组中的表达水平是正常果组的30多倍,SAUR15在落果组中的表达水平是正常果组的100多倍(图9A)。由此推测,这4个基因均参与了桑葚的果实脱落。为了验证转录组数据的准确性,对这4个基因进行RT-qPCR验证,结果发现:4个差异表达基因的荧光定量相对表达量的变化趋势与转录组表达趋势一致,说明该转录组数据可靠(图9B)。
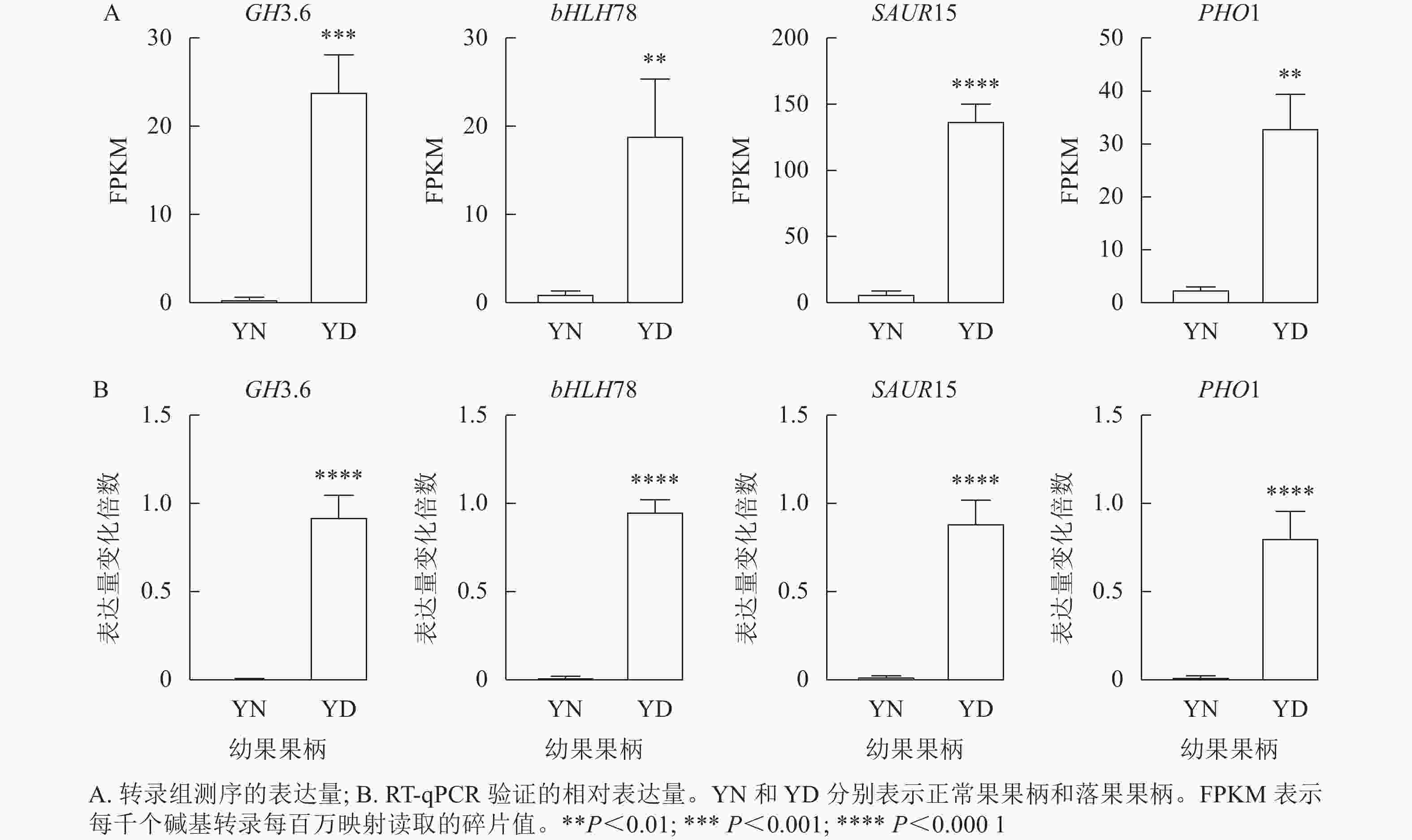
图 9 4个差异表达基因的RT-qPCR验证
Figure 9. RT-qPCR validation of four differentially expressed genes
-
近年来,为了提高果树产量,越来越多果实脱落的相关研究被报道,如番茄Lycopersicon esculentum[13]、荔枝Litchi chinensis[14]、扁桃Amygdalus communis[15]等。桑树作为一种具有重要经济价值的植物,其生理落果引起学者们的关注。2021年,有研究者对不同种类的长果桑Morus macroura的生理落果进行研究,发现高浓度的脱落酸(ABA)和乙烯(ETH)能够促进落果,而高浓度的赤霉素(GA3)和生长素(IAA)抑制落果[16]。但是,关于桑树果实脱落的分子机制还尚不清楚。
植物激素是植物生长发育过程中十分重要的物质,参与植物的落花落果,与落花落果常见的相关植物激素有IAA、GA3、细胞分裂素、ETH、ABA[17]。
IAA主要是促进植物的生长发育,抑制果实的脱落,如果生长素在运输途径中受到阻碍则会导致植物果实的脱落[18−19]。GH3.6是吲哚乙酸酰胺合成酶的基因,能够催化IAA氨基化,使生长素失活[20],这与本研究的结论一致。Small auxin-up RNA (SAUR) 基因是一类生长素早期响应基因,SAUR15能够调控植物的生长发育并参与环境胁迫响应[21]。在本研究中,SAUR15的表达水平在落果组高于正常果组100多倍,说明SAUR15基因可能参与桑葚果实脱落;ABA主要是促进果实脱落[7],这是由于ABA能够增加纤维素酶的活性进而促进果实脱落[22]。有学者通过研究无核荔枝ABA合成关键酶LcNCED与生理落果的关系,验证了ABA对于落果的作用[14];GA主要是通过作用于IAA影响果实脱落[23];ETH主要促进果实的成熟、衰老、脱落,这在多种植物中被报道,如番茄[13]等;细胞分裂素能够促进坐果,延迟果实的脱落[24]。
此外,在对落果的研究中还发现了大量的基因和转录因子的调控,如JOINTLESS[25]、LATERAL SUPPRESSOR(LS)[26]、MACROCALYX[27]主要调控离区的形成,MYB[28]、WRKY[29]、bHLH[30]、bZIP[31]等转录因子能参与植物的器官脱落。bHLH78属于bHLH转录因子,参与植物生长和代谢[32],调节花青素的生物合成[33]。本研究中,bHLH78的表达水平在落果组较高,说明其可能参与桑葚的脱落。此外,还有一些重要的酶及蛋白调控植物的器官脱落,如纤维素酶、果胶酶、多聚半乳糖醛酸酶、扩展蛋白等[34]。在本研究中,磷转运蛋白(PHO1)在落果果柄组的表达水平是正常果柄组的30多倍。此前研究表明:ABA调控依赖于PHO1的表达[35],说明PHO1基因可能参与桑葚的脱落。
-
本研究GO富集分析结果显示:有4 623个基因具有催化活性,说明在果实脱落过程中,有多种重要的酶发挥催化效应;KEGG通路富集分析结果显示:大多数差异表达基因集中在黄酮类生物合成、柠檬酸循环、植物激素信号转导、氨基酸的生物合成等通路中,说明在果实脱落过程中,植物激素、糖类、次生代谢物质等发挥了重要的作用,从而调控果实的脱落。本研究筛选了4个显著的差异表达基因,均在桑葚脱落过程中参与反应,可为今后进一步研究桑树的果实脱落提供参考。
Transcriptome analysis of abscised and normal surviving young fruit of Morus alba
-
摘要:
目的 果实的脱落是一种常见的生理现象,会在一定程度上限制果实的产量。探究桑葚幼果在脱落过程中的代谢及生化反应,可为桑葚的脱落机制研究提供参考。 方法 通过高通量测序对白桑Morus alba幼果在落果和正常果的果柄离区进行转录组测序,筛选差异表达基因,并对其进行功能注释和代谢通路分析,采用实时荧光定量PCR技术对转录组数据进行验证。 结果 经华大基因转录组测序分析共计获得262.92 Mb的原始序列;桑葚幼果果柄在2种状态下共筛选到10 481个差异表达基因,其中,有5 239个差异表达基因上调表达,5 242个差异表达基因下调表达。经基因本体数据库(GO)功能富集分析,共发现37个显著性条目,大多数差异基因具有催化活性、膜的组成成分、氧化还原酶活性、膜的内在成分等功能;京都基因与基金组百科全书(KEGG)富集分析发现:大多数差异表达基因集中在柠檬酸循环、植物激素信号转导途径、黄酮类生物合成等代谢通路上。 结论 桑葚在脱落过程中受植物激素的调控,氨基酸、黄酮类次生代谢物和碳水化合物等物质也能调控果实脱落。图9表2参35 Abstract:Objective This study, with mulberry (Morus) fruits as the subjects, is aimed to conduct an investigation into its fruit abscission, a common physiological process which limits the yield of fruit so as to clarify the underlying abscission mechanism of the metabolism and biochemical reactions. Method High-throughput sequencing was employed to conduct transcriptome analysis of the young fruit of Morus alba in the abscission zone between the abscised and normal surviving mulberry fruits to screen the differentially expressed genes before they were subjected to functional annotation and metabolic pathway analysis, and the verification of transcriptome data using real-time quantitative PCR technology. Result A total of 262.92 Mb of raw reads and 10 481 differentially expressed genes were obtained by Beijing Genomics institution (BGI, Shenzhen, China), of which 5 239 were up-regulated, whereas 5 242 were down-regulated; As was shown by the GO functional enrichment analysis, a total of 37 significant items were found, and most of the differentially expressed genes were featured with catalytic activity, membrane composition, oxidoreductase activity and membrane intrinsic components. It was shown by the KEGG enrichment analysis that most differentially expressed genes were concentrated in metabolic pathways such as citric acid cycle, plant hormone signal transduction pathway, and flavonoid biosynthesis. Conclusion Mulberry is regulated by plant hormones and substances such as amino acids, flavonoid secondary metabolites and carbohydrates during the abscission process. [Ch, 9 fig. 2 tab. 35 ref.] -
Key words:
- Morus alba /
- fruit abscission /
- transcriptome
-
图 1 落果与正常果果柄纵切面显微结构图
Figure 1 Longitudinal section microscopic structures of abscised and normal surviving young fruit peduncles
图 2 落果与正常果果柄的相关性热图
Figure 2 Correlation heatmap of abscised and normal surviving young fruit peduncles
图 3 落果与正常果果柄的表达基因和差异表达基因
Figure 3 Expressed genes and differentially expressed genes of abscised and normal surviving young fruit peduncles
图 4 落果与正常果果柄的差异表达基因GO分类柱状图
Figure 4 Histogram of GO classification of differentially expressed genes of abscised and normal surviving young fruit peduncles
图 5 落果与正常果果柄的差异表达基因GO富集气泡图
Figure 5 GO enrichment bubble chart of differentially expressed genes of abscised and normal surviving young fruit peduncles
图 6 落果与正常果果柄的差异表达基因KEGG分类柱状图
Figure 6 Histogram of KEGG classification of differentially expressed genes of abscised and normal surviving young fruit peduncles
图 7 落果与正常果果柄的差异表达基因的KEGG富集气泡图
Figure 7 KEGG enrichment bubble diagram of differentially expressed genes of abscised and normal surviving young fruit peduncles
图 8 落果与正常果果柄的代谢途径的差异表达基因热图
Figure 8 Heat map of differentially expressed genes in metabolic pathways of abscised and normal surviving carpopodium
图 9 4个差异表达基因的RT-qPCR验证
Figure 9 RT-qPCR validation of four differentially expressed genes
表 1 转录组数据RT-qPCR验证引物序列
Table 1. Primer sequences used in RT-qPCR validation of transcriptome data
基因ID 基因名 正向引物(5′→3′) 反向引物(5′→3′) XM_010096892.1 GH3.6 ACACTAACTACACCAGCCCAAA ACTTAATAGCACGAATGAACCC XM_010102492.2 bHLH78 TATTACCTTCGTCGTCCCCTCCTA ACCACTCTTTTCGTTTCCTTCACC XM_010113436.1 SAUR15 TGAAGAAGCCGAGAAGGAGTA GGTGGTAGGAGAAGGGATAAC XM_024169608.1 PHO1 GCCAATAACGACAGGAAA AACAACCCGTGAACAAAC  下载: 导出CSV
下载: 导出CSV
表 2 测序数据统计
Table 2. Statistics of sequencing data
样本
名称原始序列
数/Mb纯净序列
数/Mb纯净碱基
数/GbQ20/% Q30/% YD1 43.82 42.48 6.37 96.50 91.31 YD2 43.82 42.74 6.41 96.53 91.34 YD3 43.82 42.58 6.39 96.53 91.39 YN1 43.82 42.61 6.39 96.46 91.19 YN2 43.82 42.58 6.39 96.55 91.38 YN3 43.82 42.65 6.40 96.55 91.39
下载: 导出CSV
-
[1] JIAO Feng, LUO Rongsong, DAI Xuelei, et al. Chromosome level reference genome and population genomic analysis provide insights into the evolution and improvement of domesticated mulberry (Morus alba) [J]. Molecular Plant, 2020, 13(7): 1001 − 1012. [2] 齐明芳, 许涛, 郭泳, 等. 园艺植物器官脱落研究进展[J]. 沈阳农业大学学报, 2010, 41(6): 643 − 648. QI Mingfang, XU Tao, GUO Yong, et al. Advance of organ abscission research on horticultural plants [J]. Journal of Shenyang Agricultural University, 2010, 41(6): 643 − 648. [3] CHUNG K R, SHILTS T, ERTÜRK U, et al. Indole derivatives produced by the fungus Colletotrichum acutatum causing lime anthracnose and postbloom fruit drop of citrus [J]. FEMS Microbiology Letters, 2003, 226(1): 23 − 30. [4] COTTEE N S, BANGE M P, WILSON I W, et al. Developing controlled environment screening for high-temperature tolerance in cotton that accurately reflects performance in the field [J]. Functional Plant Biology, 2012, 39(8): 670 − 678. [5] 伏健民, 束怀瑞. 春季干旱对金冠苹果不同部位叶片衰老和脱落的影响[J]. 果树科学, 1993, 10(2): 65 − 68. FU Jianmin, SU Huairui. Effect of water stress in spring on senescence and abscission of leaves on different position in golden delicious apple trees [J]. Journal of Fruit Science, 1993, 10(2): 65 − 68. [6] 吴建阳, 李彩琴, 李建国. 荔枝ACS1基因的分离及其与幼果脱落的关系[J]. 果树学报, 2017, 34(7): 817 − 827. WU Jianyang, LI Caiqin, LI Jianguo. Isolation of ACS1 gene and the relationship between its expression and fruitlet abscission in litchi [J]. Journal of Fruit Science, 2017, 34(7): 817 − 827. [7] OHKUMA K, LYON J L, ADDICOTT F T, et al. Abscisin Ⅱ, an abscission accelerating substance from young cotton fruit [J]. Science, 1963, 142(3599): 1592 − 1593. [8] BURG S P. Ethylene, plant senescence and abscission [J]. Plant Physiology, 1968, 43: 1503 − 1511. [9] YANG Ziqin, ZHONG Xiumei, FAN Yan, et al. Burst of reactive oxygen species in pedicel-mediated fruit abscission after carbohydrate supply was cut off in longan (Dimocarpus longan)[J/OL]. Frontiers in Plant Science, 2015, 6: 360[2022-02-18]. doi: 10.3389/fpls.2015.00360. [10] 寇晓虹, 罗云波. 植物多聚半乳糖醛酸酶功能研究进展[J]. 生物技术通报, 2003(5): 15 − 18. KOU Xiaohong, LUO Yunbo. Research advance in function of plant polygalacturonase [J]. Biotechnology Bulletin, 2003(5): 15 − 18. [11] GOLDENTAL-COHEN S, BURSTEIN C, BITON I, et al. Ethephon induced oxidative stress in the olive leaf abscission zone enables development of a selective abscission compound[J/OL]. BMC Plant Biology, 2017, 17(1): 87[2022-02-20]. doi: 10.1186/s12870-017-1035-1. [12] LOVE M I, HUBER W, ANDERS S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2[J/OL]. Genome Biology, 2014, 15(12): 550[2022-02-20]. doi: 10.1186/s13059-014-0550-8. [13] DEL CAMPILLO E, BENNETT A B. Pedicel breakstrength and cellulase gene expression during tomato flower abscission [J]. Plant Physiology, 1996, 111(3): 813 − 820. [14] 陈哲, 胡福初, 年宇薇, 等. 无核荔枝ABA生物合成关键酶LcNCED基因克隆及其在生理落果阶段中的表达分析[J]. 热带作物学报, 2018, 39(2): 300 − 307. CHEN Zhe, HU Fuchu, NIAN Yuwei, et al. Expression analysis and cloning of NCED gene during physiological fruit drop period of seedless litchi fruit [J]. Chinese Journal of Tropical Crops, 2018, 39(2): 300 − 307. [15] 郭春苗, 杨波, 木巴热克·阿尤普, 等. 扁桃酸性转化酶在生理落果期的特征分析及与落果的关系[J]. 分子植物育种, 2019, 17(14): 4785 − 4790. GUO Chunmiao, YANG Bo, Mubareke Ayoupu, et al. Characteristics of acid invertase (AcAI) and its relationship with fruit drop during the physiological fruit drop of almond [J]. Molecular Plant Breeding, 2019, 17(14): 4785 − 4790. [16] 刘岩, 林天宝, 潘美良, 等. 不同长果桑生理性落果规律的调查[J]. 蚕桑通报, 2021, 52(2): 11 − 15. LIU Yan, LIN Tianbao, PAN Meiliang, et al. Investigation on the physiological abscission rule of long mulberry fruits in different varieties [J]. Bulletin of Sericulture, 2021, 52(2): 11 − 15. [17] SANTNER A, ESTELLE M. Recent advances and emerging trends in plant hormone signalling [J]. Nature, 2009, 459(7250): 1071 − 1078. [18] DRAZETA L, LANG A, CAPPELLINI C, et al. Vessel differentiation in the pedicel of apple and the effects of auxin transport inhibition [J]. Physiologia Plantarum, 2004, 120(1): 162 − 170. [19] KÜHN N, SERRANO A, ABELLO C, et al. Regulation of polar auxin transport in grapevine fruitlets (Vitis vinifera L. ) and the proposed role of auxin homeostasis during fruit abscission[J/OL]. BMC Plant Biology, 2016, 16(1): 234[2022-02-21]. doi: 10.1186/s12870-016-0914-1. [20] STASWICK P E, SERBAN B, ROWE M, et al. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid [J]. The Plant Cell, 2005, 17(2): 616 − 627. [21] 李风光. 桑树生长素早期响应基因Mul-SAUR15的功能研究[D]. 泰安: 山东农业大学, 2020. LI Fengguang. Biological Function of the Auxin Early Response Gene SAUR15 in Mulberry[D]. Tai’an: Shandong Agricultural University, 2020. [22] CHEN Bingxian, MA Jun, XU Zhenjiang, et al. Abscisic acid and ethephon regulation of cellulase in the endosperm cap and radicle during lettuce seed germination [J]. Journal of Integrative Plant Biology, 2016, 58(10): 859 − 869. [23] 樊卫国, 安华明, 刘国琴, 等. 刺梨果实与种子内源激素含量变化及其与果实发育的关系[J]. 中国农业科学, 2004, 37(5): 728 − 733. FAN Weiguo, AN Huaming, LIU Guoqin, et al. Changes of endogenous hormones contents in fruit, seeds and their effects on the fruit development of Rosa roxburghii [J]. Scientia Agricultura Sinica, 2004, 37(5): 728 − 733. [24] 周蕾, 魏琦超, 高峰. 细胞分裂素在果实及种子发育中的作用[J]. 植物生理学通讯, 2006, 42(3): 549 − 553. ZHOU Lei, WEI Qichao, GAO Feng. The effect of cytokinins on fruit and seed development [J]. Plant Physiology Communications, 2006, 42(3): 549 − 553. [25] QI Xiaoxiao, HU Shi, ZHOU Hongsheng, et al. A MADS-box transcription factor of ‘Kuerlexiangli’(Pyrus sinkiangensis Yu) PsJOINTLESS gene functions in floral organ abscission [J]. Gene, 2018, 642: 163 − 171. [26] NAKANO T, FUJISAWA M, SHIMA Y, et al. Expression profiling of tomato pre-abscission pedicels provides insights into abscission zone properties including competence to respond to abscission signals[J/OL]. BMC Plant Biology, 2013, 13: 40[2022-02-25]. doi: 10.1186/1471-2229-13-40. [27] NAKANO T, KIMBARA J, FUJISAWA M, et al. MACROCALYX and JOINTLESS interact in the transcriptional regulation of tomato fruit abscission zone development [J]. Plant Physiology, 2012, 158(1): 439 − 450. [28] LIAO Wenbin, YANG Yiling, LI Yayun, et al. Genome-wide identification of cassava R2R3 MYB family genes related to abscission zone separation after environmental-stress-induced abscission[J/OL]. Scientific Reports, 2016, 6: 32006[2022-02-10]. doi: 10.1038/srep32006. [29] CHEN Ligang, ZHANG Liping, LI Daibo, et al. WRKY8 transcription factor functions in the TMV-cg defense response by mediating both abscisic acid and ethylene signaling in Arabidopsis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(21): 1963 − 1971. [30] WANG Jinyan, HU Zhongze, ZHAO Tongmin, et al. Genome-wide analysis of bHLH transcription factor and involvement in the infection by yellow leaf curl virus in tomato (Solanum lycopersicum)[J/OL]. BMC Genomics, 2015, 16(1): 39[2022-02-22]. doi: 10.1186/s12864-015-1249-2. [31] LOVISETTO A, GUZZO F, TADIELLO A, et al. Characterization of a bZIP gene highly expressed during ripening of the peach fruit [J]. Plant Physiology and Biochemistry, 2013, 70: 462 − 470. [32] SUN Xi, WANG Yu, SUI Na. Transcriptional regulation of bHLH during plant response to stress [J]. Biochemical and Biophysical Research Communications, 2018, 503(2): 397 − 401. [33] ALBERT N W, BUTELLI E, MOSS S M A, et al. Discrete bHLH transcription factors play functionally overlapping roles in pigmentation patterning in flowers of Antirrhinum majus [J]. The New Phytologist, 2021, 231(2): 849 − 863. [34] 文晓鹏, 仇志浪, 洪怡. 果树落果的生理及分子机制研究进展[J]. 山地农业生物学报, 2018, 37(4): 1 − 17, 101. WEN Xiaopeng, QIU Zhilang, HONG Yi. Advances in physiological and molecular mechanisms underlying the fruit abscission of fruit trees [J]. Journal of Mountain Agriculture and Biology, 2018, 37(4): 1 − 17, 101. [35] ZIMMERLI C, RIBOT C, VAVASSEUR A, et al. PHO1 expression in guard cells mediates the stomatal response to abscisic acid in Arabidopsis [J]. The Plant Journal, 2012, 72(2): 199 − 211. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220205
 点击查看大图
点击查看大图

计量
- 文章访问数: 1971
- HTML全文浏览量: 459
- PDF下载量: 98
- 被引次数: 0
