





-
土地资源作为人类生存发展的基本要素和载体,根据功能可划分为生产空间、生活空间和生态空间(“三生空间”)[1]。资源型城市与其他类型城市相比,受到城镇化发展与资源开采的双重压力,生态空间被生活和生产空间挤压的时段更长,“三生空间”冲突更为强烈[2]。并且,由于土地功能不断变更,土地利用敏感度增加,景观格局剧烈变化,面临的生态安全问题也更为复杂与紧迫[3]。因此,科学提升资源型城市生态安全是促进土地可持续发展的关键。资源型城市边缘区作为城市建设中最富变化的区域,不仅涉及城乡之间的过渡,矿区与非矿区之间的矛盾也尤为突出[4],需给予重点关注。生态系统服务价值(ESV)和景观生态风险(ERI)作为评估区域生态环境的重要内容,与维护生态安全密切相关[5]。现阶段,多数学者将生态系统服务价值与景观生态风险作为2个独立的科学问题,分别从格局、过程、功能等方面开展大量研究[6−7]。但是,近期研究发现,生态系统服务价值的时空异质性可以体现景观格局与生态过程的最终结果,是修正景观生态风险的最佳指示指标[8−9]。因此,将生态系统服务价值与景观生态风险评价结合研究,有助于精确分析区域生态环境变化过程[10−11],同时也可开展生态分区等相关研究[12−13],为提升资源型城市边缘区生态安全提供新的切入点。
本研究以典型的资源型城市大庆城市边缘区为例,运用地学信息图谱、时空立方体模型、空间自相关分析等方法研究1980—2020年“三生空间”土地利用、景观风险生态系统服务价值,深入探讨资源型城市人类活动对土地利用变化的影响、“三生空间”耦合协调发展趋势及未来发展路径。
-
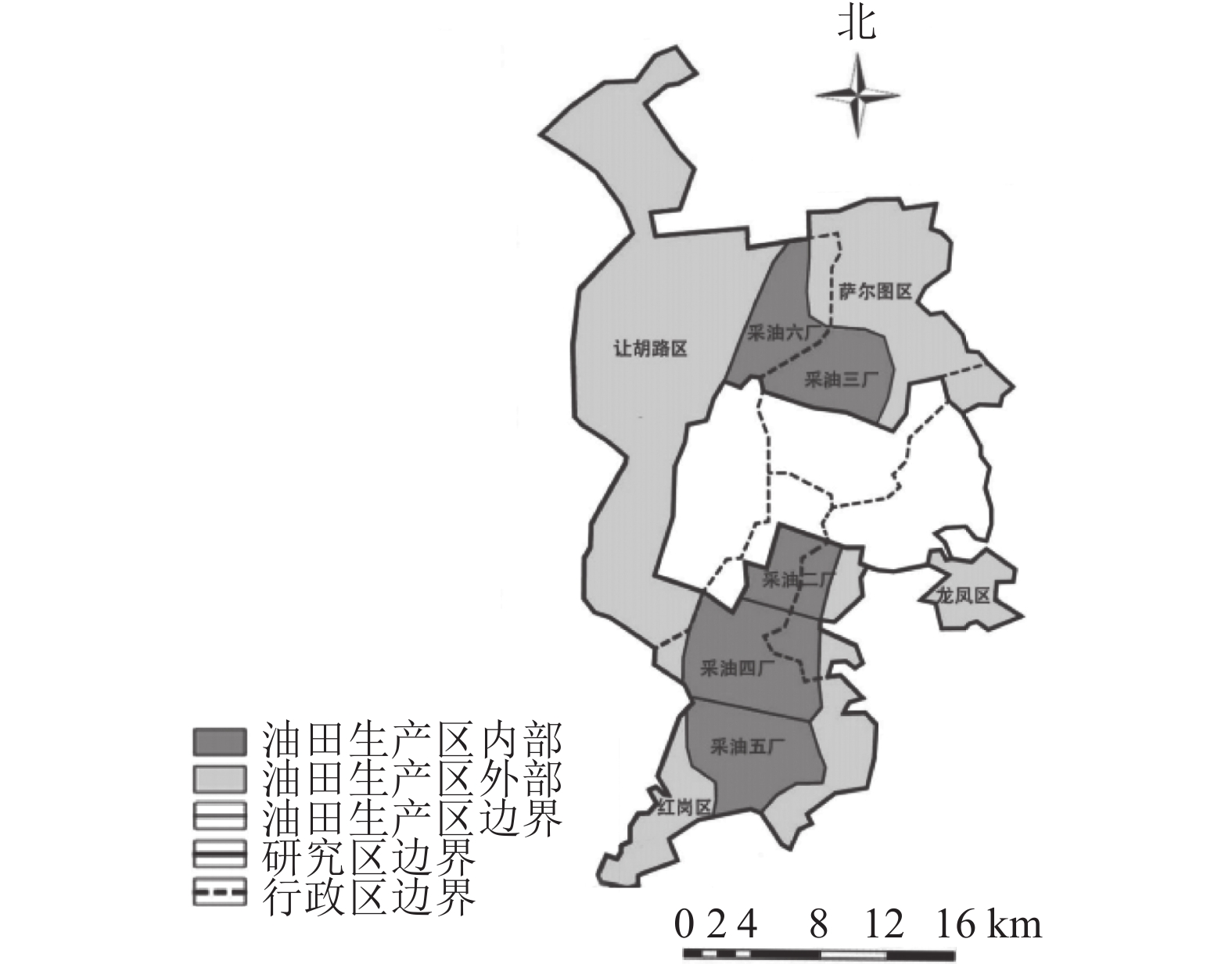
大庆市(45°23′~47°29′N,123°45′~125°47′E)地处黑龙江省西部,松嫩平原中部。东部与绥化市、哈尔滨市相接,南部与吉林省隔江(松花江)相望,西部、北部与齐齐哈尔市接壤。大庆市蕴含丰富的石油资源,大庆油田曾是中国最大的油田。本研究以大庆城市总体规划(2011—2020年)为依据,划定城市边缘区与油田生产区(简称矿区,图1)。
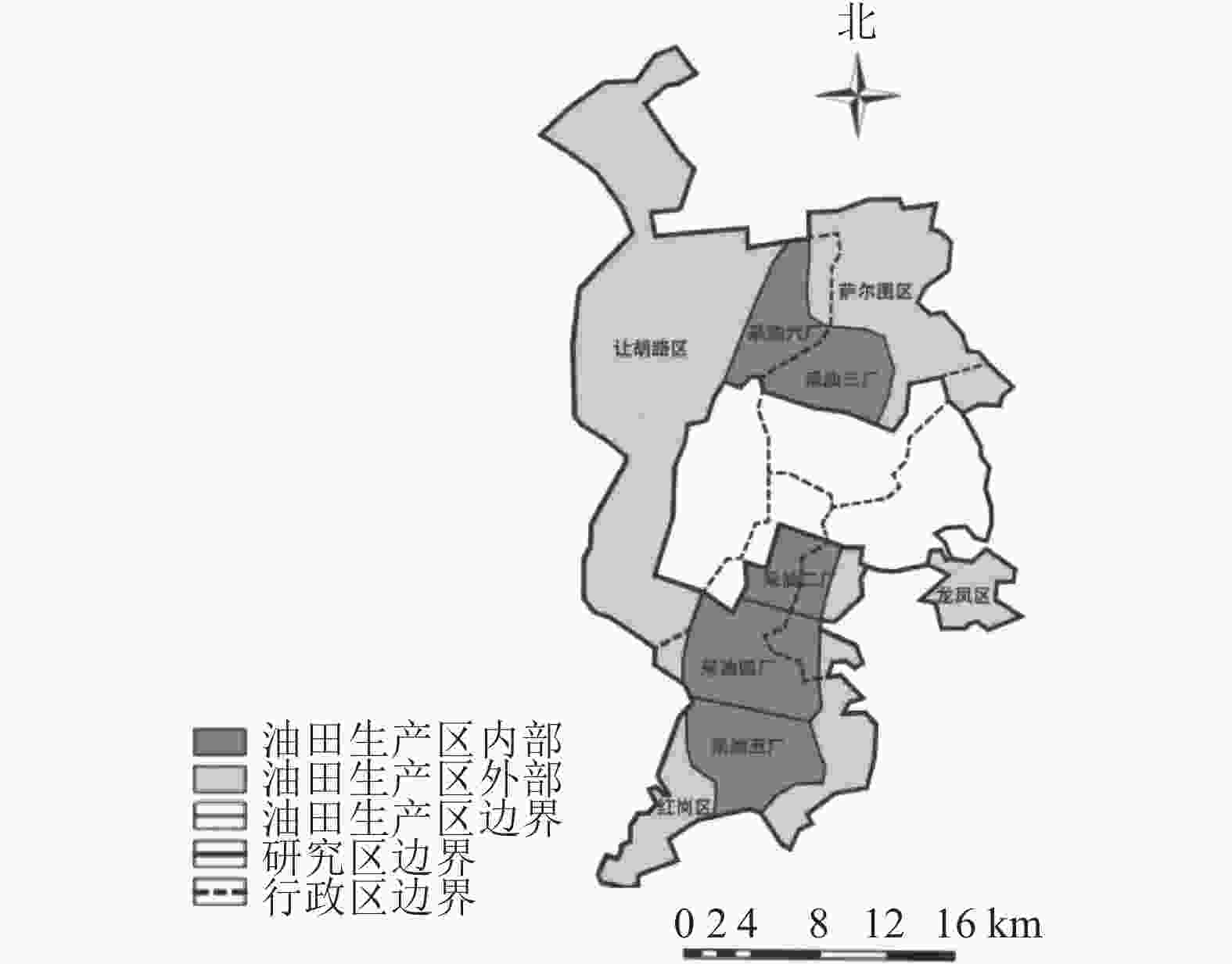
图 1 研究区示意图
Figure 1. Location of the study area
-
1980、1990、2000、2010、2020年大庆城市边缘区土地利用现状数据来源于地理监测云平台,空间分辨率为30 m。行政区划数据源于国家基础地理信息中心。粮食数据来自《中国农村统计年鉴 1981—2021年》和黑龙江省 2020 年国民经济和社会发展统计公报。本研究根据研究区土地利用特点,参考关于“三生空间”的相关文献[14−15],将土地利用分类系统分为3个一级土地利用空间与8个二级土地利用空间(表1)。
表 1 土地利用主导功能分类
Table 1. Land use function classification
一级地类 二级地类 类型 赋分 生产空间 农业生产空间 水田、旱地 6 工业生产空间 工交建设用地 3 生活空间 城市生活空间 城镇用地 1 乡村生活空间 农村居民点 2 生态空间 草地生态空间 高覆盖度草地、中覆盖度草地、低覆盖度草地 4 水域生态空间 河流、湖泊、水库坑塘等 8 林地生态空间 有林地、灌木林、疏林
地、其他林地5 其他生态空间 盐碱地、沙地、裸土地等 7 -
为了将研究数据进行可视化表达,从空间上更直观地分析研究区景观生态风险与生态系统服务价值的变化情况,借鉴已有研究和结合研究区实际情况,利用ArcGIS渔网工具,将研究区划分成1 km×1 km的单元网格(评价单元),共得到2 342个评价单元。
-
以 1980、1990、2000、2010和2020年“三生空间”数据为基本地理单元,利用ArcGIS构建 1980—1990、1990—2000、2000—2010、2010—2020年2位数编码地学信息图谱。计算公式如下:Nc=Na×10+Nb。其中,Na和Nb分别表示研究初期a年和末期b年土地“三生空间”二级类型图谱单元编码;Nc为土地转移新生图谱编码。
-
景观生态风险可以衡量生态系统受到的外界干扰强度和内部抵抗能力[16]。计算公式如下:
$$ {E}_{\mathrm{R}\mathrm{I}k}=\sum\limits_{i = 1}^n \frac{{A}_{ki}}{{A}_{k}}\times {R}_{i}=\sum\limits _{i=1}^{n}\frac{{A}_{ki}}{{A}_{k}}\left({E}_{i}\times {F}_{i}\right)=\sum\limits _{i=1}^{n}\frac{{A}_{ki}}{{A}_{k}}\left\{\left({a_1}{C}_{i}+{b_1}{S}_{i}+{c_1}{D}_{i}\right)\times {F}_{i}\right\} 。 $$ 其中:ERIk为第k个评价单元土地利用景观生态风险;Aki为第k个评价单元土地利用类型i的面积;Ak为第k个评价单元的总面积;Ri为土地利用类型i的景观损失度指数;Ei为土地利用类型i的景观干扰度指数;Ci、Si和 Di分别为景观破碎度指数、景观分离度指数和景观优势度指数;a1、b1、c1 为各景观指数权重,且三者相加为1,根据参考文献[17−18],将3个指数分别赋值为0.502、0.301、0.197;Fi为景观i类型的景观脆弱度指数,根据参考文献[19−21]以及结合研究区土地利用类型的变化情况,对8个土地利用空间空间赋分(表1),并进行归一化处理。
-
生态系统服务功能是人们从生态系统获取的效益,包含直接和间接的、有形和无形的效益[22]。计算公式为:${E}_{\mathrm{S}\mathrm{V}}=\displaystyle \sum\limits _{i=1}^{n}{E}_{\mathrm{S}\mathrm{V}k}=\displaystyle \sum\limits _{i=1}^{n}\left({V}_{\mathrm{C}i}\times {A}_{i}\right)$。其中,ESV为研究区生态系统服务功能总价值(元);ESVk为第k个评价单元生态系统服务功能价值(元);Ai为评价单元内第i类土地利用类型面积(hm2);n为土地利用类型数;VCi为单位面积土地利用类型i的生态系统服务价值(元·hm−2)。其中,生态系统服务价值以谢高地等[23]的中国陆地生态系统服务单位面积价值为基础,辅以文献[24−25]所提供的比例关系确定。在确定农业生活空间生态系统服务价值时,从《大庆统计年鉴》中获取 1980、1990、2000、2010、2020年粮食作物种植面积(hm2)及粮食作物总产值(元)数据,得出大庆市农田提供食物生产服务的单价平均值为1 079.49 元·hm−2。按照自然断点法分类并采用相对指标[26],从景观生态风险与生态系统服务价值上划分为低、较低、中、较高、高共5类区域。
-
以时空立方体模型为基础[27],计算每个立方体条柱的Getis-Ord Gi*统计量,探测某特征在时空尺度的热点或冷点,结合M-K检验法对时空尺度的热点分析结果进行趋势检验,从而识别数据聚集程度的时空趋势[28]。
-
空间自相关分析可以探索某属性值是否具有空间集聚性和异质性[29]。运用GEODA空间分析工具,通过局部空间自相关分析[30]确定未来“三生空间”的管控区域与发展方向。
-
研究区总面积为2342 km2,其中矿区面积为639.52 km2,占研究面积的23.86%。根据1980—2010年土地面积统计可知(表2),研究区生态空间面积较广,各类型面积从大到小依次为草地、其他、水域、林地。生产空间次之,各类型面积从大到小依次为农业、工业。生活空间最少,各类型面积从大到小依次为乡村、城市。根据矿区内外“三生空间”比例分布可知,农业生产空间、草地生态空间矿区内外分布较为均等;城市生活空间、水域生态空间、林地生态空间分布矿区外大于矿区内;乡村生活空间、工业生产空间、其他生态空间分布矿区外小于矿区内。
表 2 1980—2020年研究区“三生空间”面积与分布比例统计
Table 2. Statistics on the area and distribution ratio of the “production-living-ecological space” in the study area in 1980−2020
一级
地类二级
地类1980年 1990年 2000年 2010年 2020年 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 生产
空间农业 673.87 97.79 99.10 668.83 97.48 99.03 766.09 99.58 99.17 732.65 97.57 98.74 726.15 88.44 95.72 工业 8.37 2.21 0.90 9.11 2.52 0.97 9.11 0.42 0.83 11.17 2.43 1.26 45.31 11.31 4.28 生活
空间城市 10.25 0.12 15.19 16.91 0.12 22.38 16.91 0.11 23.09 28.42 8.18 30.75 30.53 8.04 29.73 乡村 80.10 99.88 72.35 90.39 99.88 72.24 93.82 99.89 76.91 94.04 91.82 69.25 102.54 91.96 70.27 生态
空间水域 213.28 7.03 12.28 293.65 14.06 15.72 272.94 12.02 17.09 268.78 11.44 15.75 272.56 3.65 17.20 草地 1229.57 53.76 65.89 925.63 42.60 50.06 833.10 42.15 50.35 855.32 43.49 47.83 804.85 40.35 45.24 林地 19.24 0.31 1.20 94.49 0.82 6.17 11.75 0.98 7.12 97.63 0.92 6.76 121.25 1.05 8.54 其他 460.67 38.90 19.58 600.97 47.11 26.70 607.12 48.42 30.78 606.85 47.51 28.57 592.15 49.77 27.90 -
由表3可知:1980—2020年,生产空间与生态空间之间土地转移最为活跃,面积为418.44 km2,生活与生态空间之间转移次之,面积为88.11 km2,生产与生活之间转移最少,面积为25.56 km2。其中,1980—1990年,主要为矿外生态与生产空间之间相互转移;1990—2000年,主要为矿外生态空间转移为生产空间;2000—2010年,主要为矿区外生产与生态空间之间相互转移,及矿区内生活空间转移为生态空间;2010—2020年,主要为矿外生产与生态空间相互转移,及矿内生态空间转移为生活空间。
表 3 1980—2020年研究区“三生空间”面积转移与分布比例统计
Table 3. Statistics on the area transfer and distribution ratio of the “production-living-ecological space” in the study area in 1980−2020
时段 面积 生产—生活 生产—生态 生活—生产 生活—生态 生态—生产 生态—生活 1980—1990年 矿外面积比例/% 62.54 86.38 8.17 59.35 96.53 48.81 矿内面积比例/% 37.46 13.62 91.83 40.65 3.47 51.19 转移总面积/km2 12.07 62.68 0.66 0.11 79.48 1.18 1990—2000年 矿外面积比例/% 1.62 87.46 33.33 0 90.69 0.74 矿内面积比例/% 98.38 12.54 66.67 0 9.31 99.26 转移总面积/km2 1.61 5.12 0 0 97.38 0.12 2000—2010年 矿外面积比例/% 69.27 65.29 59.70 4.44 87.70 99.37 矿内面积比例/% 30.73 34.71 40.30 95.56 12.30 0.63 转移总面积/km2 5.19 64.75 3.54 36.15 49.55 5.59 2010—2020年 矿外面积比例/% 100 98.59 0 0 34.91 14.87 矿内面积比例/% 0 1.41 0 0 65.09 85.13 转移总面积/km2 2.49 25.54 0 0 33.94 44.96 -
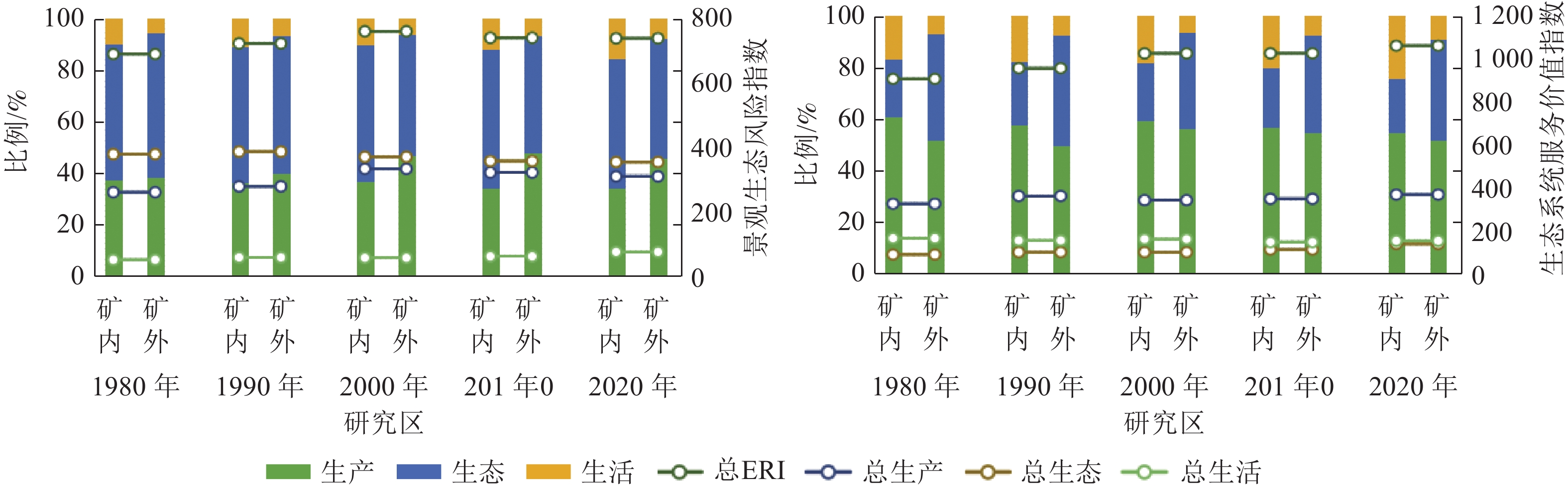
研究区总体景观生态风险与生态系统服务价值指数呈波动上升态势(图2)。研究期初风险与价值指数较低,1980—1990年变化幅度最大,呈现大幅增长趋势。从“三生空间”角度分析可知,景观生态风险指数:生活空间与生产空间呈上升趋势,生态空间呈下降趋势。“三生空间”生态系统服务价值指数均呈上升趋势。

图 2 1980—2020年研究区景观生态风险和生态系统服务价值变化趋势
Figure 2. Changing trend of landscape ecological risk and ecosystem service value in 1980−2020
-
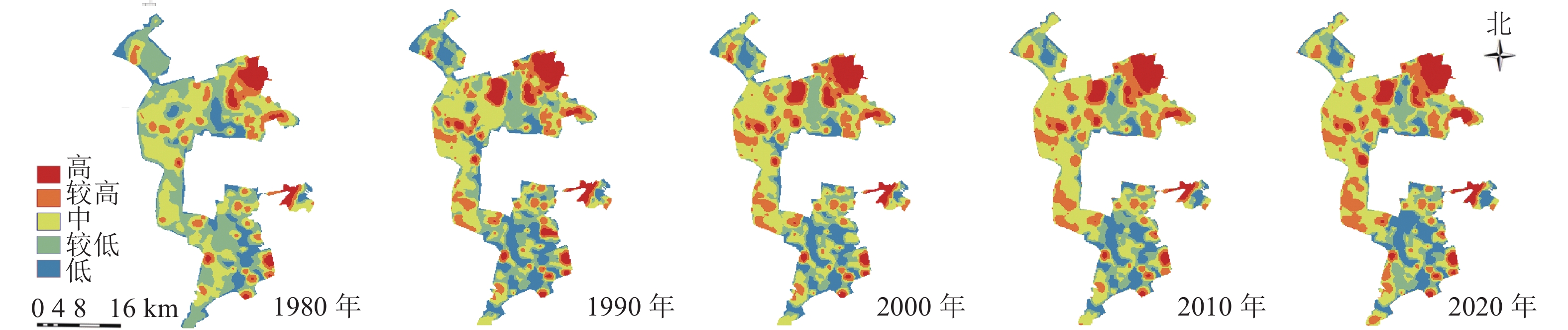
研究区1980—2020年景观生态风险的分布格局图(图3)与生态系统服务价值分布格局图(图4)显示:低风险区主要由生活与生态空间组成,较低风险区主要由生产与生态空间组成;高与较高风险区主要位于矿区内部,其中较高风险区主要由生产与生态空间组成,高风险区主要由生态空间组成。中价值区主要由矿区外生产用地组成,低与较低价值区主要由矿区内部生态空间与生产空间组成,高与较高价值区主要由矿区外部生态空间组成。
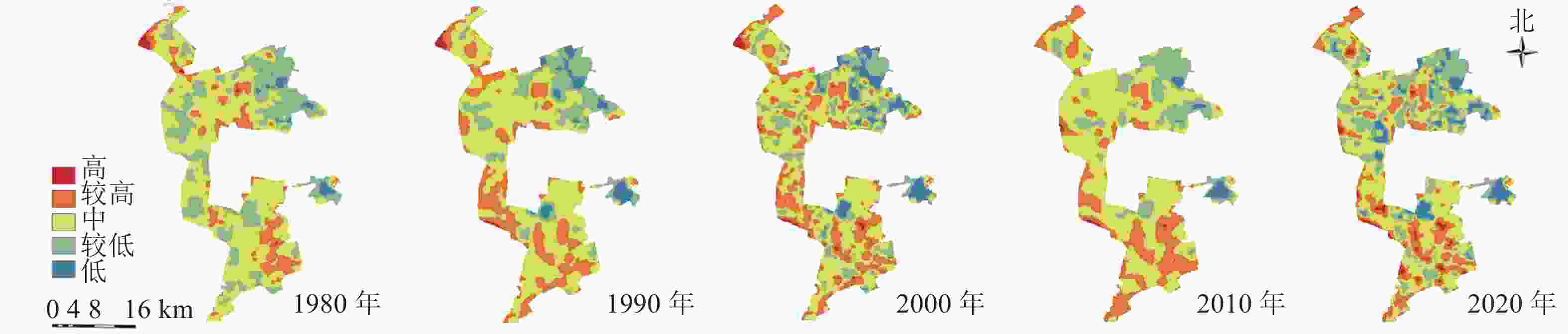
图 3 1980—2020年研究区景观生态风险分布示意图
Figure 3. Landscape ecological risk distribution map in 1980−2020
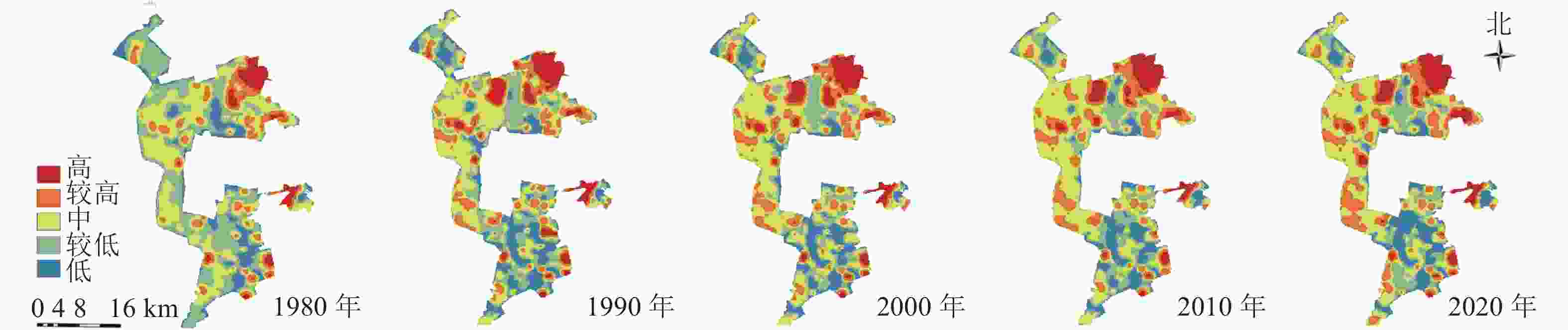
图 4 1980—2020年研究区生态系统服务价值分布示意图
Figure 4. Ecosystem service value distribution map in 1980−2020
-
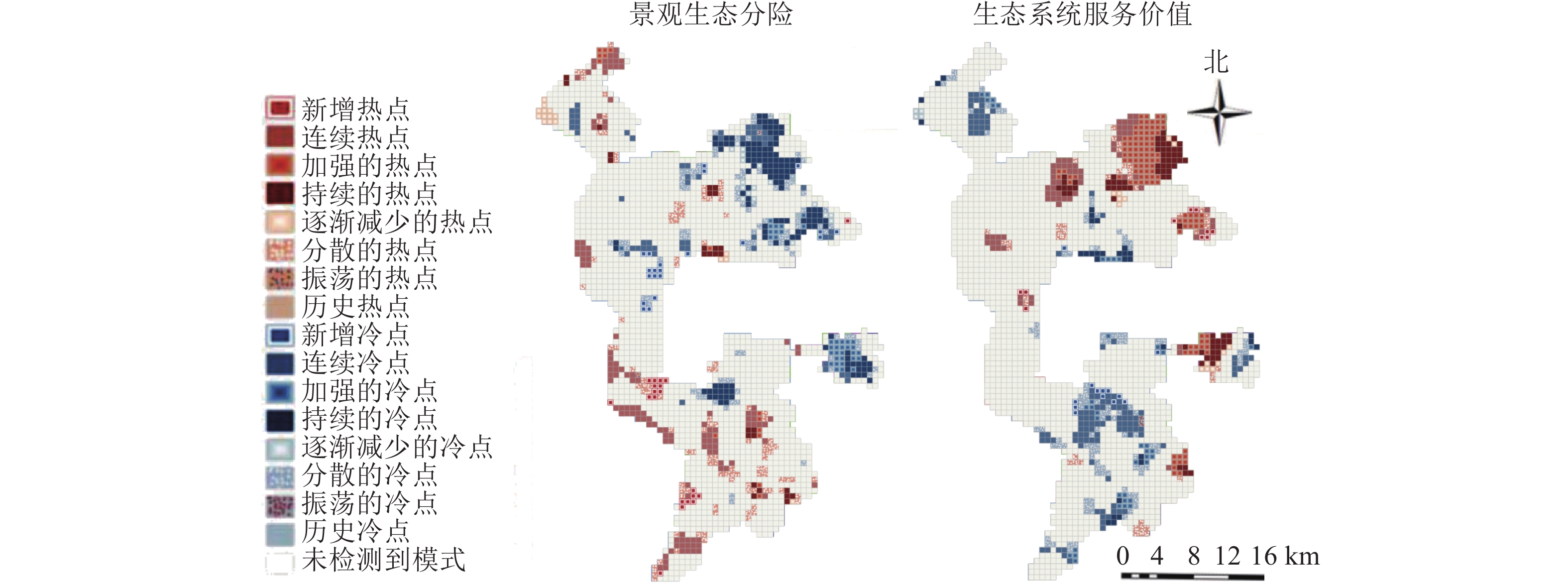
研究区景观生态风险与生态系统服务价值时空热点分析图(图5)显示:冷、热点分别由连续、持续、加强、减弱、新增、分散共6个部分组成。根据研究区景观生态风险时空热点分析可知:持续与连续冷点主要为水域生产空间、城市生活空间与矿区内部工业生产空间组成。新增冷点主要为矿区外部其他生态空间向农村生活空间转移。分散冷点主要为生态用地与生产用地的过渡区,减弱冷点为矿区外生态空间向生产空间转移,加强冷点为矿区外生产空间向生活空间转移。持续与连续热点主要为其他生态空间中的盐碱地组成,新增热点为矿区外生态用地向生产用地转移,分散热点为生态用地与生产用地的过渡区,减弱热点为矿区外草地生态空间内部转移。

图 5 景观生态风险和生态系统服务价值时空热点聚类示意图
Figure 5. Spatial and temporal hotspot analysis of landscape ecological risk and ecosystem service
根据生态系统服务价值时空热点分析可知:持续与连续热点主要为生态空间,由高覆盖度草地与水域组成。新增与加强的热点主要为矿区外部生态空间内部转移,体现为草地生态空间向水域生态空间转移。分散热点主要为生态用地与生产用地的过渡区,减弱热点为矿区外生态空间向生产空间转移。持续与连续冷点主要由其他生态用地中的盐碱地组成,新增冷点为矿区内生态用地向生产用地的转移,分散冷点为生态用地与生产用地的过渡区,加强冷点为矿区内生态空间内部转移体现为草地生态空间向其他生态空间转移。
-
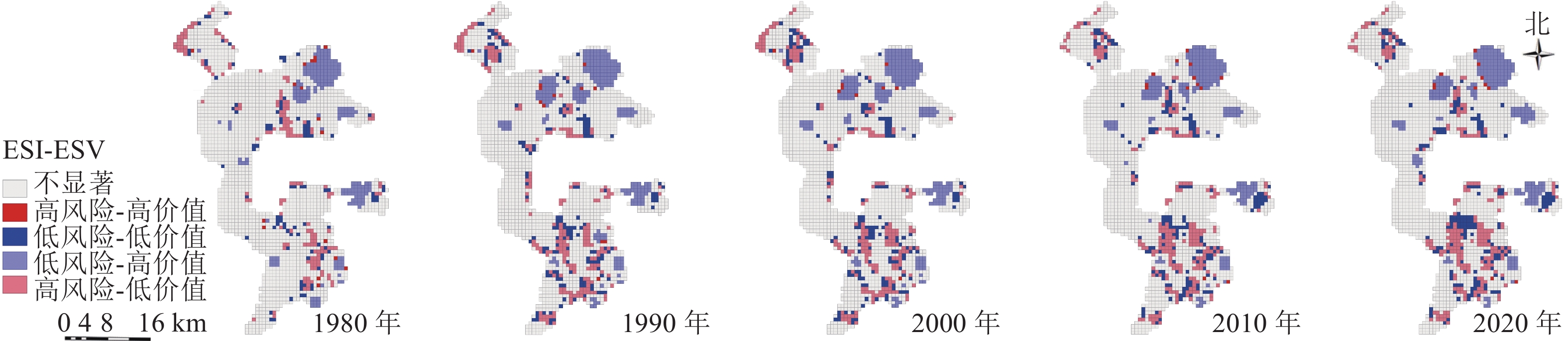
利用局部空间自相关分析,将研究区空间关系分为高风险-高价值区、低风险-低价值区、低风险-高价值区、高风险-低价值区和不显著区共5种(图6),结果显示:低风险-高价值区域主要由矿区外水域生态空间组成,是未来生态保护的重点区域。高风险-低价值区域主要由矿区内其他生态空间(盐碱地)组成,是未来生态修复的重点区域。这2类空间是维护区域生态安全稳定的核心区域,需要重点管控。研究区空间关系主要变化年限为1980—1990年,其原因主要为矿区外草地生态空间向水域生态空间的转移,以及矿区内部草地生态空间向其他生态空间的转移。高风险-高价值区域与低风险-低价值区域主要为“三生空间”过渡区域,其中高风险-高价值区域主要位于矿区外,是水域生态空间与农业生产空间的过渡区域,低风险-低价值区域主要由矿区内部生产与生态过渡区域组成,这2类空间虽然占比较少,变化较为稳定,但是是影响区域未来生态安全的主要风险源。因此,未来应将生产空间作为主要风险源进行科学防范。
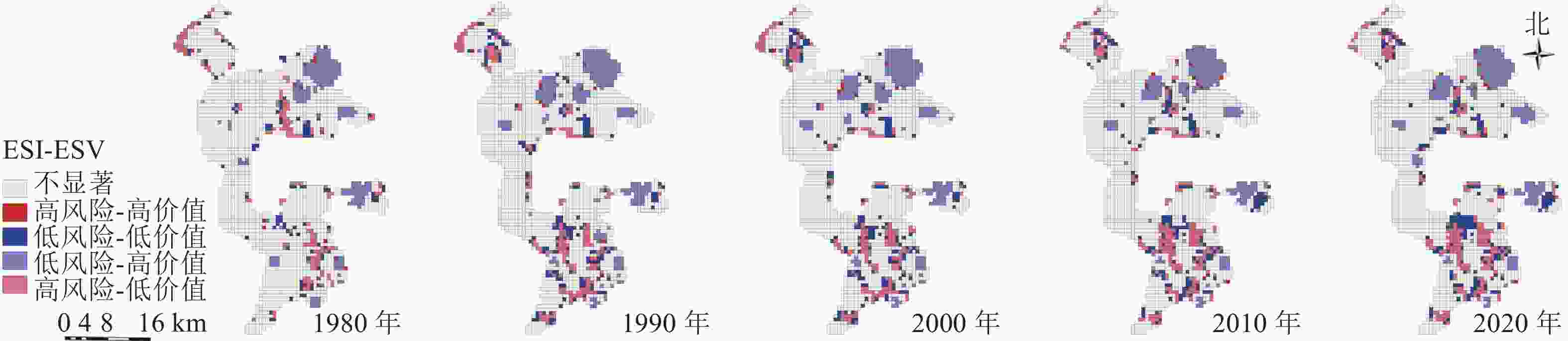
图 6 1980—2020年景观生态风险与生态系统价值关系聚类示意图
Figure 6. Cluster map of the relationship between landscape ecological risk and ecosystem value in 1980−2020
-
资源型城市“三生空间”土地利用变化的影响机制与人类活动关系密切,通过探讨人类扰动因子对“三生空间”土地利用的影响,有助于未来土地利用的决策制定。
研究区生产空间变化主要受到产业产值的影响。生活空间变化主要与人口因素相关,由于城市边缘区经济发展较为落后,与中心城区距离较近,使得农村人口多去城市务工,边缘区农村人口数量流失明显。生态空间变化主要与油田开采相关,油田开采破坏地表植被使得土地裸露,并且开采过程中消耗大量地下水资源,引起地表沉降形成多个湖泊的同时也加重土地盐碱程度。这与其他资源型城市研究结果相似[31]。相比之下,非资源型城市的生产空间变化主要受到经济发展水平的影响,生活空间变化主要与城市规划和管理相关,生态空间变化则主要与环境保护政策相关[32]。
-
景观生态风险与生态系统服务价值的耦合性能够间接反映出区域生态系统中“三生空间”的协调性,探讨生态、生产、生活空间之间的耦合协调关系对于维护资源型城市边缘区生态安全具有重要意义。
资源型城市受到资源不可再生性影响,一般要经历兴起期、成长期、繁荣期、衰退期(再生期) 4个阶段。与非资源型城市相比,其“三生空间”耦合协调性不仅受到城市规划建设的影响[33],也受资源发展周期影响[34]。1980—2020年间,研究区“三生空间”呈由失调向协调转变的趋势,矿区内“三生空间”耦合协调程度低于矿区外,这与其他资源型城市研究结果[35]相似。2000年前,大庆市处于兴起期与成长期,这一时段大庆市进行大规模的城市建设与油田开采活动,使得“三生空间”呈现“高耦合、低协调”的极度失调特征。2000年后,大庆市进入繁荣期与衰退期,这一时期城市建设逐渐稳步,同时石油储备降低、开采活动减弱,“三生空间”呈现协调发展特征。
-
1980—2020 年,研究区景观生态系统服务价值虽然有所改善,但景观生态风险处于上升趋势,说明生态安全仍面临一定压力。根据风险-价值的关系分析结果,结合大庆市国土空间总体规划(2021—2035年),从“三生空间”视域协调土地关系推动构建生态安全格局:①划定城市生态修复区,将矿区内其他生态空间(盐碱地)作为重点修复区域。要特别注重矿区内部生态环境保护,做到石油开采与环境建设兼顾,提高矿区生态环境质量。②保护城市水源地与湿地,将矿区外部水域生态空间作为核心保护区域,对于该区生态保护策略应侧重原有景观的维护,将其作为城市重要的生态斑块进行管理,助力构建“一核一区一带、多廊多点”的城市生态安全格局。③生产空间作为大庆城市边缘区主要的风险源,禁止农业生产空间侵占生态空间,防止工业生产空间无序扩张,建立风险预警机制,强化生态安全格局屏障。
-
本研究基于 1980、1990、2000、2010 和 2020 年的土地利用现状分类图,从“三生空间”的视域,以大庆市边缘区为例,定量研究资源型城市景观生态风险指数及生态系统服务价值时空变化特征,并利用空间回归模型进一步分析两者之间的关系,主要结论如下:①根据“三生空间”土地分布与转移分析可知:研究区生态空间面积较广,生产空间次之,生活空间最少。矿区外土地转移活跃度大于矿区内,其中,生产空间与生态空间之间转移最为活跃;②根据风险与价值的时空特征分析可知,1980—2020年,研究区“三生空间”景观生态风险与生态系统服务价值呈上升趋势。根据时空热点分析可知,景观生态风险与生态系统服务价值的高低分布有明显的区域性特点且较为稳定。其中,矿区内风险高于矿区外,矿区内价值低于矿区外。③根据景观生态风险与生态系统服务价值关联分析可知:风险与价值之间存在显著相关性。其中,低风险高价值区域主要由矿区外水域生态空间组成,高风险低价值区域主要由矿区内其他生态空间组成。
综上所述,未来大庆城市边缘区在生态环境建设过程中应以缩小矿区内外差异为首要任务,降低人为因素(特别是油田开采)对“三生空间”的扰动,加强各区政府与油田管理局之间的协作机制,共同统筹区域“三生空间”建设,重点保护矿区外水域生态空间,修复矿区内盐碱地,并防止生产空间进一步扩张。
Research on land use change and risk-value of “production-living-ecological space” in a resource-based city
-
摘要:
目的 以大庆城市边缘区为研究对象,在生产空间、生活空间和生态空间(“三生空间”)视域下,探究资源型城市土地利用及其风险和价值时空变化特征,为维护区域生态安全提供保障。 方法 基于1980、1990、2000、2010、2020年共5期多源数据,利用地学信息图谱、时空立方体模型、局部空间自相关分析等方法,分析“三生空间”土地分布特征、土地利用转移规律及风险-价值时空异质性、空间关联性,探讨人类活动对“三生空间”变化的影响、“三生空间”耦合协调发展趋势以及土地未来发展路径。 结果 研究区“三生空间”面积从大到小依次为生态空间、生产空间、生活空间。其中,生产空间与生态空间之间的转移最为活跃,1980—2020年面积为418.44 km2。研究区景观生态风险与生态系统服务价值指数呈上升趋势。矿区内呈现高风险、低价值特征。矿区外呈现低风险、高价值特征。 结论 受人口数量、油田开采与产业产值等人为干扰因素的影响,矿区内“三生空间”失调,矿区外“三生空间”协调。大庆城市边缘区在生态环境建设过程中应以缩小矿区内外差异为首要任务,加强各区政府与油田管理局之间的协作机制,使矿区内外协同发展。图6表3参35 Abstract:Objective This study aims to explore the spatio-temporal characteristics of land use and risk-value changes in resource-based cities from the perspective of “production-living-ecological space”, taking Daqing urban fringe as an example, so as to provide guarantee for maintaining regional ecological security. Method Based on the multi-source data of five periods in 1980, 1990, 2000, 2010 and 2020, the distribution and transfer characteristics of land use, spatio-temporal heterogeneity spatial correlation of risk−value are analyzed by using geo-information atlas, spatio-temporal cube model, local spatial auto-correlation analysis and other methods. The factor analysis method and coupling degree coordination model are used to further explore the impact of human activities on the spatial changes of land use, the development trend of “production-living-ecological space” coupling coordination, and the future development path of the city. Result The area of “production-living-ecological space” in the study area: ecological space>production space>living space. Among them, the transfer between production space and ecological space is the most active, with an area of 418.44 km2 during 1980−2020. The landscape ecological risk and ecosystem service value index in the study area showed an upward trend. The oilfield area is characterized by high risk and low value. Outside the oilfield area is characterized by low risk and high value. Conclusion Affected by factors such as population, oilfield exploitation, and industrial output, the “production-living-ecological space” inside the oilfield is imbalanced, while the “production-living-ecological space” outside the oilfield is coordinated. In the process of ecological environment construction in the urban fringe of Daqing, the primary task should be to narrow the differences between the inside and outside of the oilfield area, strengthen the cooperation mechanism between the regional governments and the oilfield administration, so as to achieve the coordinated development between the inside and outside of the oilfield area. [Ch, 6 fig. 3 tab. 35 ref.] -
图 2 1980—2020年研究区景观生态风险和生态系统服务价值变化趋势
Figure 2 Changing trend of landscape ecological risk and ecosystem service value in 1980−2020
图 3 1980—2020年研究区景观生态风险分布示意图
Figure 3 Landscape ecological risk distribution map in 1980−2020
图 4 1980—2020年研究区生态系统服务价值分布示意图
Figure 4 Ecosystem service value distribution map in 1980−2020
图 5 景观生态风险和生态系统服务价值时空热点聚类示意图
Figure 5 Spatial and temporal hotspot analysis of landscape ecological risk and ecosystem service
图 6 1980—2020年景观生态风险与生态系统价值关系聚类示意图
Figure 6 Cluster map of the relationship between landscape ecological risk and ecosystem value in 1980−2020
表 1 土地利用主导功能分类
Table 1. Land use function classification
一级地类 二级地类 类型 赋分 生产空间 农业生产空间 水田、旱地 6 工业生产空间 工交建设用地 3 生活空间 城市生活空间 城镇用地 1 乡村生活空间 农村居民点 2 生态空间 草地生态空间 高覆盖度草地、中覆盖度草地、低覆盖度草地 4 水域生态空间 河流、湖泊、水库坑塘等 8 林地生态空间 有林地、灌木林、疏林
地、其他林地5 其他生态空间 盐碱地、沙地、裸土地等 7  下载: 导出CSV
下载: 导出CSV
表 2 1980—2020年研究区“三生空间”面积与分布比例统计
Table 2. Statistics on the area and distribution ratio of the “production-living-ecological space” in the study area in 1980−2020
一级
地类二级
地类1980年 1990年 2000年 2010年 2020年 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 面积/km2 矿内/% 矿外/% 生产
空间农业 673.87 97.79 99.10 668.83 97.48 99.03 766.09 99.58 99.17 732.65 97.57 98.74 726.15 88.44 95.72 工业 8.37 2.21 0.90 9.11 2.52 0.97 9.11 0.42 0.83 11.17 2.43 1.26 45.31 11.31 4.28 生活
空间城市 10.25 0.12 15.19 16.91 0.12 22.38 16.91 0.11 23.09 28.42 8.18 30.75 30.53 8.04 29.73 乡村 80.10 99.88 72.35 90.39 99.88 72.24 93.82 99.89 76.91 94.04 91.82 69.25 102.54 91.96 70.27 生态
空间水域 213.28 7.03 12.28 293.65 14.06 15.72 272.94 12.02 17.09 268.78 11.44 15.75 272.56 3.65 17.20 草地 1229.57 53.76 65.89 925.63 42.60 50.06 833.10 42.15 50.35 855.32 43.49 47.83 804.85 40.35 45.24 林地 19.24 0.31 1.20 94.49 0.82 6.17 11.75 0.98 7.12 97.63 0.92 6.76 121.25 1.05 8.54 其他 460.67 38.90 19.58 600.97 47.11 26.70 607.12 48.42 30.78 606.85 47.51 28.57 592.15 49.77 27.90
下载: 导出CSV
表 3 1980—2020年研究区“三生空间”面积转移与分布比例统计
Table 3. Statistics on the area transfer and distribution ratio of the “production-living-ecological space” in the study area in 1980−2020
时段 面积 生产—生活 生产—生态 生活—生产 生活—生态 生态—生产 生态—生活 1980—1990年 矿外面积比例/% 62.54 86.38 8.17 59.35 96.53 48.81 矿内面积比例/% 37.46 13.62 91.83 40.65 3.47 51.19 转移总面积/km2 12.07 62.68 0.66 0.11 79.48 1.18 1990—2000年 矿外面积比例/% 1.62 87.46 33.33 0 90.69 0.74 矿内面积比例/% 98.38 12.54 66.67 0 9.31 99.26 转移总面积/km2 1.61 5.12 0 0 97.38 0.12 2000—2010年 矿外面积比例/% 69.27 65.29 59.70 4.44 87.70 99.37 矿内面积比例/% 30.73 34.71 40.30 95.56 12.30 0.63 转移总面积/km2 5.19 64.75 3.54 36.15 49.55 5.59 2010—2020年 矿外面积比例/% 100 98.59 0 0 34.91 14.87 矿内面积比例/% 0 1.41 0 0 65.09 85.13 转移总面积/km2 2.49 25.54 0 0 33.94 44.96
下载: 导出CSV
-
[1] 陈晓红, 许晓庆, 刘艳军, 等. 基于三生空间质量的哈长城市群城市脆弱性时空演变格局及驱动力研究[J]. 生态学报, 2022, 42(15): 6395 − 6405. CHEN Xiaohong, XU Xiaoqing, LIU Yanjun, et al. Patterns and driving forces of the temporal-spatial evolution of urban vulnerability in Harbin-Changchun urban agglomeration based on the production-living-ecological spatial quality [J]. Acta Ecologica Sinica, 2022, 42(15): 6395 − 6405. [2] 李江苏, 孙威, 余建辉. 黄河流域三生空间的演变与区域差异——基于资源型与非资源型城市的对比[J]. 资源科学, 2020, 42(12): 2285 − 2299. LI Jiangsu, SUN Wei, YU Jianhui. Change and regional differences of production-living-ecological space in the Yellow River Basin: based on comparative analysis of resource-based and non-resource-based cities [J]. Resources Science, 2020, 42(12): 2285 − 2299. [3] 黄天能, 张云兰. 基于“三生空间”的土地利用功能演变及生态环境响应——以桂西资源富集区为例[J]. 生态学报, 2021, 41(1): 348 − 359. HUANG Tianneng, ZHANG Yunlan. Transformation of land use function and response of eco-environment based on“production-life-ecology space”: a case study of resource-rich area in western Guangxi [J]. Acta Ecologica Sinica, 2021, 41(1): 348 − 359. [4] 管晶, 焦华富. 煤炭资源型城市城乡空间结构演变及影响因素——以安徽省淮北市为例[J]. 自然资源学报, 2021, 36(11): 2836 − 2852. GUAN Jing, JIAO Huafu. The evolution process and influencing factors of urban-rural spatial structure in coal resource-based city: a case study of Huaibei City in Anhui Province [J]. Journal of Natural Resources, 2021, 36(11): 2836 − 2852. [5] 潘卫涛, 岳邦瑞, 姚龙杰, 等. 耦合风险与服务的市域生态安全格局构建——以陕西省咸阳市为例[J]. 应用生态学报, 2023, 34(1): 178 − 186. PAN Weitao, YUE Bangrui, YAO Longjie, et al. Urban ecological security pattern construction coupled with risk and service: a case study of Xianyang City, Shaanxi Province, China [J]. Chinese Journal of Applied Ecology, 2023, 34(1): 178 − 186. [6] 徐超平, 夏斌. 资源型城市土地利用变化及其对生态系统服务价值的影响[J]. 生态环境学报, 2010, 19(12): 2887 − 2891. XU Chaoping, XIA Bin. Land use changes and its influences on ecosystem service value of resources-based city [J]. Ecology and Environmental Sciences, 2010, 19(12): 2887 − 2891. [7] 于淑会, 康园园, 邓伟, 等. 太行山东部县域“三生”用地转型与景观生态风险分析——以河北省平山县为例[J]. 中国生态农业学报, 2022, 30(7): 1113 − 1122. YU Shuhui, KANG Yuanyuan, DENG Wei, et al. Analysis of “production-living-ecological” land transformation and landscape ecological risk in the eastern counties of the Taihang Mountain: a case study in Pingshan County [J]. Chinese Journal of Eco-Agriculture, 2022, 30(7): 1113 − 1122. [8] 刘长峰, 侯鹰, 陈卫平, 等. 基于生态系统服务的城市化区域生态风险表征方法研究[J]. 生态学报, 2021, 41(9): 3343 − 3353. LIU Changfeng, HOU Ying, CHEN Weiping, et al. Research on ecological risk characterization methods for urbanized areas based on ecosystem services [J]. Acta Ecologica Sinica, 2021, 41(9): 3343 − 3353. [9] 曹祺文, 张曦文, 马洪坤, 等. 景观生态风险研究进展及基于生态系统服务的评价框架: ESRISK[J]. 地理学报, 2018, 73(5): 843 − 855. CAO Qiwen, ZHANG Xiwen, MA Hongkun, et al. Review of landscape ecological risk and an assessment framework based on ecological services: ESRISK [J]. Acta Geographica Sinica, 2018, 73(5): 843 − 855. [10] 李骊, 张青青, 王雅梅, 等. 2000—2018年克孜河流域生态系统脆弱性、服务功能价值及风险评价[J]. 中国沙漠, 2021, 41(2): 164 − 172. LI Li, ZHANG Qingqing, WANG Yamei, et al. Comprehensive assessment of ecosystem vulnerability, the value of service function and risk in Kezi River Basin in 2000−2018 [J]. Journal of Desert Research, 2021, 41(2): 164 − 172. [11] 朱润苗, 陈松林. 1980—2020年福建省景观生态风险与生态系统服务价值的空间关系[J]. 应用生态学报, 2022, 33(6): 1599 − 1607. ZHU Runmiao, CHEN Songlin. Spatial relationship between landscape ecological risk and ecosystem service value in Fujian Province, China during 1980−2020 [J]. Chinese Journal of Applied Ecology, 2022, 33(6): 1599 − 1607. [12] 谢丽霞, 白永平, 车磊, 等. 基于价值-风险的黄河上游生态功能区生态分区建设[J]. 自然资源学报, 2021, 36(1): 196 − 207. XIE Lixia, BAI Yongping, CHE Lei, et al. Construction of ecological zone based on value-risk ecological function area in the Upper Yellow River [J]. Journal of Natural Resources, 2021, 36(1): 196 − 207. [13] 徐维艺, 阚瑶川, 王国文, 等. 基于价值-风险的阜平县生态分区研究[J]. 林业与生态科学, 2022, 37(1): 46 − 56. XU Weiyi, KAN Yaochuan, WANG Guowen, et al. Study on ecological regionalization of Fuping County based on value-risk [J]. Forestry and Ecological Sciences, 2022, 37(1): 46 − 56. [14] 勾蒙蒙, 刘常富, 李乐, 等. “三生空间”视角下三峡库区土地利用转型的生态系统服务价值效应[J]. 应用生态学报, 2021, 32(11): 3933 − 3941. GOU Mengmeng, LIU Changfu, LI Le, et al. Ecosystem service value effects of the Three Gorges Reservoir Area land use transformation under the perspective of “production-living-ecological” space [J]. Chinese Journal of Applied Ecology, 2021, 32(11): 3933 − 3941. [15] 逯承鹏, 纪薇, 刘志良, 等. 黄河流域甘肃段县域“三生”功能空间时空格局及影响因素识别[J]. 地理科学, 2022, 42(4): 579 − 588. LU Chengpeng, JI Wei, LIU Zhiliang, et al. Spatial-temporal pattern and influencing factors of the “Production-LivingEcological” functional space of the Yellow River Basin at county level in Gansu, China [J]. Scientia Geographica Sinica, 2022, 42(4): 579 − 588. [16] WANG Hong, LIU Xingming, ZHAO Chuanyan, et al. Spatial-temporal pattern analysis of landscape ecological risk assessment based on land use/land cover change in Baishuijiang National nature reserve in Gansu Province, China [J/OL]. Ecological Indicators, 2021, 124: 107454[2022-10-16]. doi: 10.1016/j.ecolind.2021.107454. [17] 杨庚, 张振佳, 曹银贵, 等. 晋北大型露天矿区景观生态风险时空异质性[J]. 生态学杂志, 2021, 40(1): 187 − 198. YANG Geng, ZHANG Zhenjia, CAO Yingui, et al. Spatial-temporal heterogeneity of landscape ecological risk of large-scale open-pit mining area in north Shanxi [J]. Journal of Ecology, 2021, 40(1): 187 − 198. [18] 常小燕, 李新举, 李西灿, 等. 矿区土地利用生态风险的时空异质性[J]. 生态学报, 2019, 39(9): 3075 − 3088. CHANG Xiaoyan, LI Xinju, LI Xican, et al. Spatial-temporal heterogeneity of ecological risk of land use in mining areas [J]. Acta Ecologica Sinica, 2019, 39(9): 3075 − 3088. [19] 陈斌, 徐尚昭, 周阳阳, 等. “三生空间”视角下宜昌市景观生态安全评价及其耦合特征分析[J]. 水土保持研究, 2022, 29(4): 344 − 351. CHEN Bin, XU Shangzhao, ZHOU Yangyang, et al. Evaluation and coupling coordination analysis of landscape ecological security of Yichang from the perspective of production-life-ecological space [J]. Research of Soil and Water Conservation, 2022, 29(4): 344 − 351. [20] 王娟娟, 毋兆鹏. 乌鲁木齐市2000—2018年“三生”空间格局演变及其风险评价[J]. 水土保持通报, 2021, 41(6): 318 − 326, 335. WANG Juanjuan, WU Zhaopeng. Evolution on patterns and risk assessment of productive-living-ecological space in Urumqi City during 2000−2018 [J]. Bulletin of Soil and Water Conservation, 2021, 41(6): 318 − 326, 335. [21] 李少玲, 谢苗苗, 李汉廷, 等. 资源型城市景观生态风险的时空分异: 以乌海市为例[J]. 地学前缘, 2021, 28(4): 100 − 109. LI Shaoling, XIE Miaomiao, LI Hanting, et al. Spatio-temporal dynamics of landscape ecological risk in resource-based cities: a case study of Wuhai [J]. Earth Science Frontiers, 2021, 28(4): 100 − 109. [22] HAN Jiazheng, HU Zhenqi, WANG Peijun, et al. Spatio-temporal evolution and optimization analysis of ecosystem service value-a case study of coal resource-based city group in Shandong, China [J/OL]. Journal of Cleaner Production, 2022, 363: 132602[2022-10-16]. doi: 10.1016/j.jclepro.2022.132602. [23] 谢高地, 张彩霞, 张昌顺, 等. 中国生态系统服务的价值[J]. 资源科学, 2015, 37(9): 1740 − 1746. XIE Gaodi, ZHANG Caixia, ZHANG Changshun, et al. The value of ecosystem services in China [J]. Resource Sciences, 2015, 37(9): 1740 − 1746. [24] 苏宁, 丁国栋, 杜林芳, 等. 人类活动对资源型城市生态系统服务价值的影响——以鄂尔多斯为例[J]. 生态学报, 2022, 42(16): 6543 − 6553. SU Ning, DING Guodong, DU Linfang, et al. Impact of human activities on ecosystem services value of resource-based city: a case study in Ordos, the Inner Mongolia [J]. Acta Ecologica Sinica, 2022, 42(16): 6543 − 6553. [25] 李锋, 陈春. “三生”用地功能动态变化及其生态系统服务价值响应——以顺德区为例[J]. 水土保持研究, 2021, 28(1): 250 − 257. LI Feng, CHEN Chun. Dynamic change of ecological-living-industrial land and the response to ecosystem service value−a case study of Shunde [J]. Research of Soil and Water Conservation, 2021, 28(1): 250 − 257. [26] 郭彦君, 郭文炯. “三生空间”视角下山西中部盆地城市群景观生态风险分析[J]. 生态学杂志, 2022, 41(9): 1813 − 1824. GUO Yanjun, GUO Wenjiong. Landscape ecological risk analysis of urban agglomeration in the central basin of Shanxi from the perspective of “production-living-ecological spaces” [J]. Chinese Journal of Ecology, 2022, 41(9): 1813 − 1824. [27] MOANGA D, BIGING G, RADKE J, et al. The space-time cube as an approach to quantifying future wildfires in California [J]. International Journal of Wildland Fire, 2021, 30(2): 139 − 153. [28] XU Dehe, ZHANG Qi, DING Yan, et al. Spatiotemporal pattern mining of drought in the last 40 years in China based on the SPEI and space-time cube [J]. Journal of Applied Meteorology and Climatology, 2021, 60(9): 1219 − 1230. [29] KE Xiaoling, WANG Xingyue, GUO Haixiang, et al. Urban ecological security evaluation and spatial correlation research−based on data analysis of 16 cities in Hubei Province of China [J/OL]. Journal of Cleaner Production, 2021, 311: 127613[2022-10-18]. doi: 10.1016/j.jclepro.2021.127613. [30] GE Fengjian, CHEN Wanxu, ZENG Yuanyuan, et al. The nexus between urbanization and traffic accessibility in the middle reaches of the Yangtze River urban agglomerations, China[J]. International Journal of Environmental Research and Public Health, 2021, 18(7): 3823[2022-10-18]. doi: 10.3390/ijerph18073828. [31] 张小虎, 张合兵, 雷国平. 大庆市土地利用变化及驱动机制分析[J]. 干旱区资源与环境, 2012, 26(9): 109 − 113. ZHANG Xiaohu, ZHANG Hebing, LEI Guoping. Land use change and driving mechanism in Daqing City [J]. Journal of Arid Land Resources and Environment, 2012, 26(9): 109 − 113. [32] 张豪, 冯长春, 郭永沛. 城市边缘区“三生”空间格局演变与驱动因素分析——以北京市朝阳区为例[J]. 北京大学学报(自然科学版), 2023, 59(3): 478 − 488. ZHANG Hao, FENG Changchun, GUO Yongpei. Evolution and driving factors of “Production-Living-Ecological Space” in rural-urban fringe of Beijing [J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 2023, 59(3): 478 − 488. [33] 翟羽娟, 张艳红, 姜琦刚, 等. 吉林省西部农牧交错区“三生空间”时空演变[J]. 吉林大学学报(地球科学版), 2022, 52(3): 1016 − 1026. ZHAI Yujuan, ZHANG Yanhong, JIANG Qigang, et al. Spatial-temporal evolution pattern of “Production-Living-Ecological Space” in agro-pastoral ecotone of western Jilin Province [J]. Journal of Jilin University, 2022, 52(3): 1016 − 1026. [34] 窦睿音, 张生玲, 刘学敏. 中国资源型城市“三生系统”耦合协调发展研究[J]. 统计与决策, 2021, 37(2): 98 − 102. DOU Ruiyin, ZHANG Shengling, LIU Xuemin. Research on the coupled coordinated development of the “production system-life system-ecosystem” in resource-based cities in China [J]. Statistics &Decision, 2021, 37(2): 98 − 102. [35] 窦睿音, 张生玲, 刘学敏. 中国资源型城市“三生系统”耦合协调时空分异演变及其影响因素分析[J]. 北京师范大学学报(自然科学版), 2021, 57(3): 363 − 371. DOU Ruiyin, ZHANG Shengling, LIU Xuemin. Spatial and temporal diversity patterns and influencing factors in “production system-life system-ecosystem” coupled coordination in resource-based cities in China [J]. Journal of Beijing Normal University (Natural Science), 2021, 57(3): 363 − 371. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220666
 点击查看大图
点击查看大图
计量
- 文章访问数: 2209
- HTML全文浏览量: 420
- PDF下载量: 22
- 被引次数: 0
