





-
森林生态系统是陆地生态系统的重要组成部分,具有丰富的生物多样性[1−3]。受全球气候变化和人类活动的双重影响,森林生态系统生物多样性下降,生态功能也随之退化[4−5]。20世纪50年代以来,生态学领域学者对植物多样性有了比较深入的研究,ANTONIO等[6]分析了海拔1 500~2 500 m的植物多样性变化,发现在0.1 hm2内的植物多样性最丰富,同时随海拔上升物种丰富度出现下降的趋势。针对植物多样性的变化和形成机制有多种假说,包括生产力和空间规模,群落内干扰与斑块动态、物种共进化与异质性形成规律、群落内物种的共存机制等[7−10],但仍没有完善的理论体系。因此,多样性的形成机制成为研究森林物种多样性的主要方向之一。对植物多样性的研究主要在生物多样性编目、信息系统的制定和就地保护等方面[11−12];尤其在物种多样性变化规律[13−14],多样性形成机制[15]等方面积累了大量研究成果。物种多样性变化研究主要从时间尺度(年份、季节),空间尺度(经纬度、海拔)及生境变化等方面探究物种多样性的动态。植物多样性形成机制研究主要从环境因子(地形、土壤、气候等),人类干扰等方面探讨物种多样性存在差异,但对于森林不同死亡梯度下植物多样性的研究尚属空白。曾楠[16]研究发现:在20世纪末到21世纪初内蒙古罕山次生林,包括赛罕乌拉国家级自然保护区、特金罕山自然保护区、黄岗梁自然保护区、高格斯台罕乌拉自然保护区和青山国家级自然保护区等,因干旱发生了较强的斑块性大面积山杨Populus davidiana死亡现象。在此背景下,山杨死亡后森林结构和环境的变化对林下植被更新和植物多样性的影响值得关注。本研究通过罕山次生林北段青山国家级自然保护区山杨林树木不同死亡梯度状态(树木发生死亡即活立木到枯立木到倒木的发生状态,倒木、枯立木及死木质物统称为木质残体)来划分不同死亡阶段的样地,对死亡后林下植物种多样性进行研究,探讨山杨林不同死亡梯度下植物多样性的特点,为研究区森林生物多样性保护和次生林演替规律提供参考。
-
研究区位于内蒙古自治区兴安盟科右前旗内蒙古青山国家级自然保护区(45°53′33″~46°09′27″N,121°11′27″~121°32′31″E,以下简称青山保护区),位于内蒙古大兴安岭中段东坡,是内蒙古大兴安岭南部罕山次生林的北段。青山保护区的土壤可分为暗棕壤、黑钙土、灰色森林土、草甸土、棕色针叶林土。植被类型以落叶阔叶林为主,有山杨林、白桦Betula platyphylla林、蒙古栎Quercus mongolica林等;灌丛植被主要有照山白Rhododendron micranthum、土庄绣线菊Spiraea pubescens等;草本植物主要有地榆Sanguisorba officinalis、歪头菜Vicia unijuga等。
-
样地选在山杨次生林内,按照林分不同死亡及退化梯度,设置了轻度死亡样地(山杨基本正常生长,少量倒木和死树),中度死亡样地(多以枯稍、将死或已经死亡枯立的山杨为主),重度死亡样地(大部分山杨已经死亡且已形成大量倒木)等3个不同死亡梯度的样地,样地大小为20 m×20 m,每个梯度设置3个重复,共计9块样地(表1)。每块样地内采用五点法设置5个灌木样方,规格大小为5 m×5 m,分别在4个角和正中间,共计45个灌木样方。同样方法,每块样地内设置5个1 m×l m草本样方,共计45个草本样方。沿每个样地对角线设置3块5 m×5 m的更新样方,调查苗木更新情况,共计27块样方。
表 1 样地基本信息
Table 1. Basic information of test plots
样地 纬度(N) 经度(E) 海拔/m 坡向 坡度/(°) 倒木密度/(株·hm−2) 枯立木密度/(株·hm−2) L1 45°57′13.78″ 121.21′26.53″ 668 西北 12 588±53 308±29 L2 45°57′12.82″ 121°21′27.51″ 710 西北 10 783±72 259±16 L3 45°57′13.23″ 121°21′26.35″ 721 西北 13 975±68 346±25 K1 45°57′12.67″ 121°21′29.44″ 708 西北 16 958±36 392±69 K2 45°57′12.77″ 121°21′30.41″ 686 西北 19 1053±88 352±59 K3 45°57′13.69″ 121°21′32.03″ 696 西北 12 896±65 453±73 D1 45°59′49.32″ 121°22′56.48″ 432 西北 18 1375±46 350±43 D2 45°59′52.47″ 121°22′58.04″ 772 西北 17 1029±69 296±69 D3 45°59′52.75″ 121°22′59.42″ 770 西北 23 1698±63 398±73 说明:L为轻度死亡林分,K为中度死亡林分,D为重度死亡林分。1、2、3为样地重复编号。 -
在样地内进行每木检尺,调查样地内胸径1 cm以上(胸径1 cm以下的乔木定为幼苗)的所有树木胸径、树高、冠幅、郁闭度和林下更新情况,利用抬头望法和冠层分析仪相结合调查森林郁闭度;灌木和草本分别调查高度、盖度、株丛数、多样性。记录每个样地的相对光照强度(森林中的光照/全光照)、坡度、坡位、坡向、凋落物厚度等,并通过国际倒木分类和调查研究体系进行倒木基础特征调查,如表1和表2所示。
表 2 不同死亡梯度林分因子、环境因子特征
Table 2. Characteristics of forest factors and environmental factors in different regions
样地 凋落物厚度/cm 萌蘖更新/(株·hm−2) 活立木密度/(株·hm−2) 木质残体密度/(株·hm−2) 死亡率/% 郁闭度 L1 6.27±0.45 6 950 733.25±62.92 a 318.00±13.30 a 17.39 0.63 L2 5.87±0.45 3 975 756.43±54.56 a 329.00±18.30 a 16.65 0.66 L3 5.38±0.45 5 325 693.32±48.52 a 315.00±16.37 a 15.55 0.60 K1 6.48±0.43 10 125 588.56±56.76 b 483.35±187.65 a 46.54 0.69 K2 6.48±0.43 15 741 575.00±86.50 b 495.32±167.38 a 48.76 0.71 K3 6.48±0.43 15 158 538.53±67.63 b 465.54±187.64 a 45.25 0.78 D1 3.26±0.42 18 400 242.56±74.00 c 1467.65±343.15 b 43.93 0.55 D2 2.26±0.23 3 941 250.67±63.00 c 1591.46±245.31 b 45.17 0.56 D3 3.56±0.45 1 975 268.54±68.00 c 1591.85±413.36 b 42.83 0.61 说明:不同小写字母表示不同样地间差异显著(P<0.05)。 -
通过野外调查数据,采用Margalef丰富度指数(D)、Shannon-Wiener 多样性指数(H)、Pielou均匀度指数(J)和Simpson优势度指数(H’ )4类指标,重要值(IV)和Sorensen群落相似系数(SI)来综合分析评价青山保护区林下不同植被措施的植物物种多样性及其差异。
数据采用Excel 2010整理,方差分析在SPSS 23中进行,相关分析与多元线性回归方程通过R语言程序进行处理,绘图在SigmaPlot 14.0中进行。对不同死亡梯度样地内的植物群落进行主坐标分析(PCoA),使研究数据相似性或差异性呈现可视化坐标。利用VennDiagram绘制维恩(Venn)图。
-
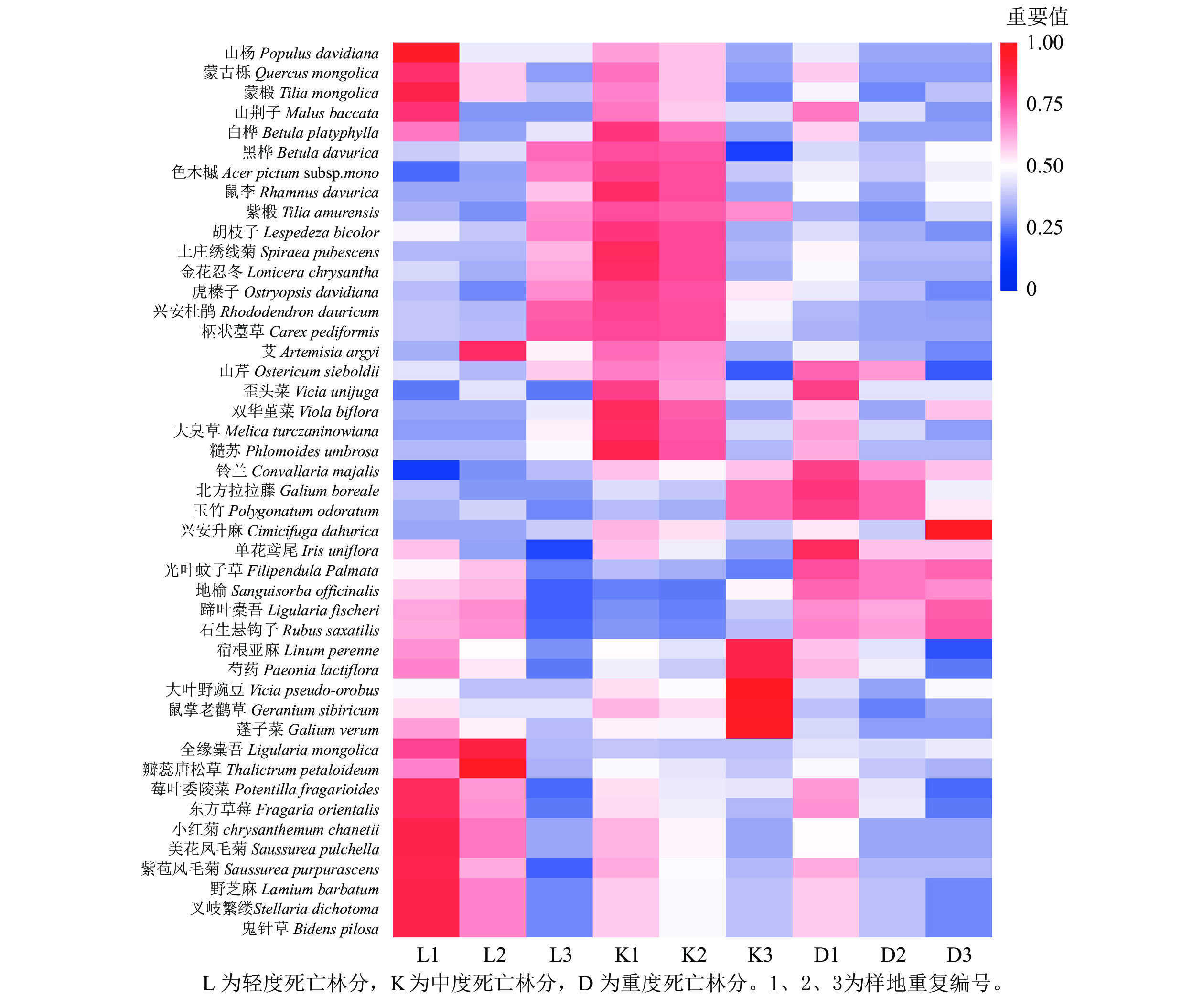
青山保护区森林不同死亡梯度样地共发现野生植物45种。其中,乔木层9种、灌木层5种、草本层31种。轻度死亡样地群落共有30种植物。其中,乔木物种共有6种植物,占群落总物种数的15.64%。如图1所示:山杨和蒙古栎为群落优势种,色木槭Acer mono和黑桦Betula davurica为伴生种。灌木有3种,占群落总物种数的9.38%,群落优势种为土庄绣线菊,重要值为0.59,金花忍冬Lonicera chrysantha为伴生种。草本植物21种,占群落总物种数的81.25%,重要值最大的分别是柄状薹草Caex pediformis和玉竹Polygonatum odoratum,为群落优势种。
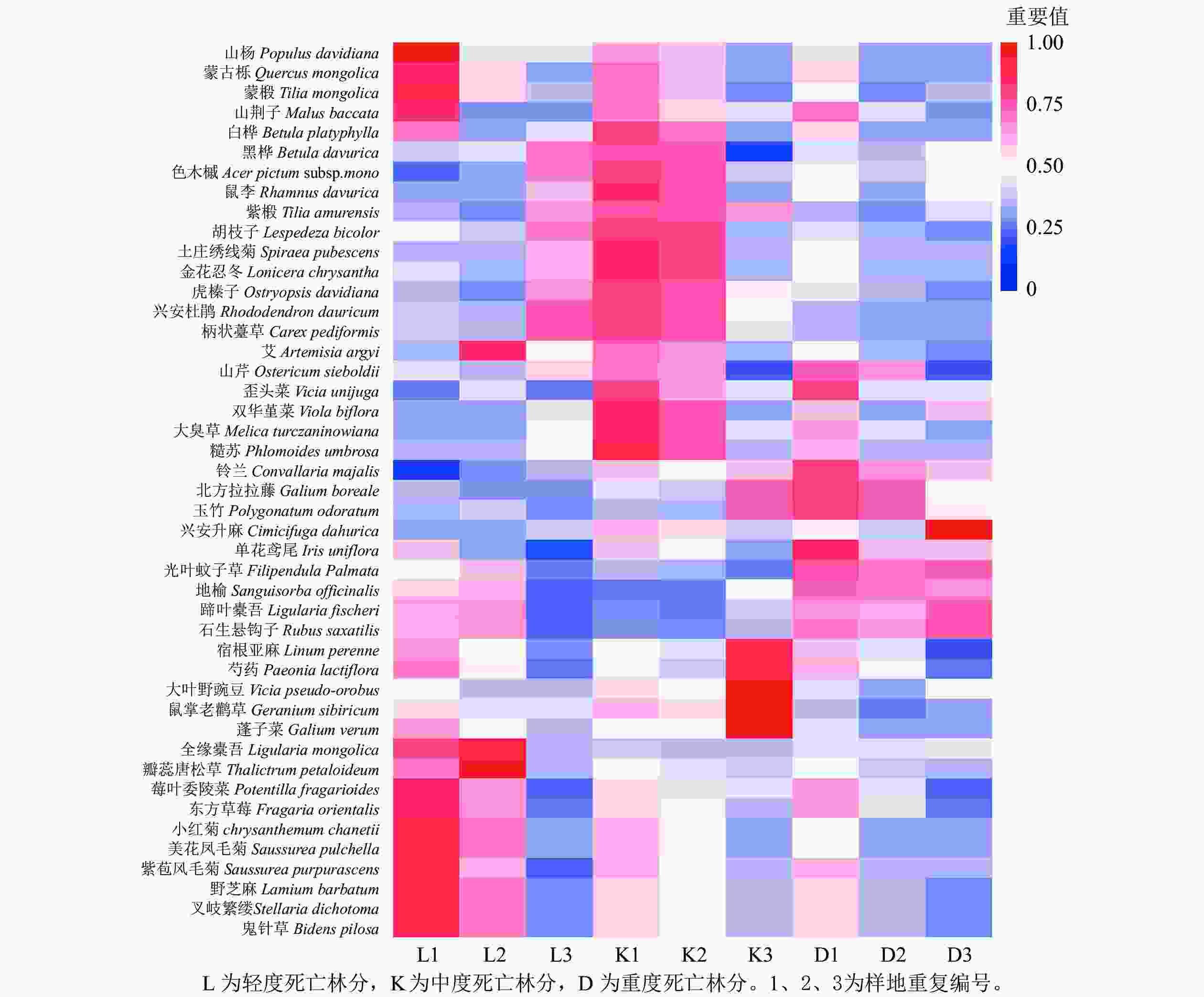
图 1 不同死亡梯度林下乔灌草重要值分布
Figure 1. Distribution of important values of arbor, shrub and grass under forests with different death gradients
中度死亡样地共发现植物24种。其中,乔木物种有7种植物,占群落总物种数的22.22%。重要值最大的是山杨和蒙古栎,是该群落的优势种,山荆子Malus baccata、蒙椴Tilia mongolica、白桦、色木槭为伴生种。灌木物种有2种,占群落总物种数的11.11%,优势种为土庄绣线菊和金花忍冬,分别为0.65和0.34。草本植物15种,占群落总物种数的66.67%,重要值最大的分别是柄状薹草和玉竹,重要值分别为0.43、0.10,为该层优势种。
重度死亡样地共发现植物30种。其中,乔木物种有8种,占群落总物种数的23.08%,山杨和紫椴Tilia amurensis 为优势种,重要值分别是0.39、0.28,蒙古栎、蒙椴等为伴生种。灌木物种有5种,占群落总物种数的19.23%,土庄绣线菊和虎榛子Ostryopsis davidiana为优势种,重要值分别为0.38和0.43。胡枝子Lespedeza bicolor、金花忍冬和兴安杜鹃Rhododendron dauricum为伴生种。草本植物17种,占群落总物种数的57.69%,柄状薹草和大臭草Melica turczaninowiana为群落优势种,重要值分别为0.55和0.13。
-
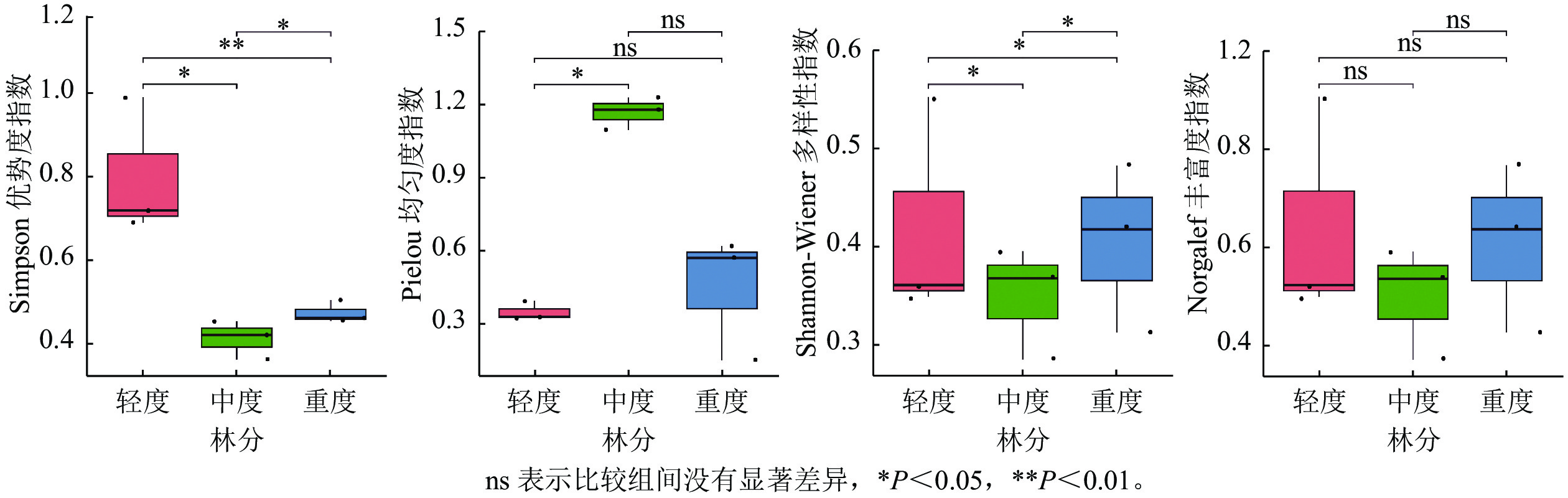
对不同死亡梯度林分群落各层α多样性进行分析(图2),乔木层的Simpson指数从大到小依次为重度死亡林分、中度死亡林分、轻度死亡林分,不同林分乔木层Simpson指数为0.28~0.36。灌木层的Simpson指数从大到小依次为重度死亡林分、轻度死亡林分、中度死亡林分。草本层的Simpson指数从大到小依次为轻度死亡林分、中度死亡林分、重度死亡林分,不同林分草本层Simpson指数为0.46~0.68。乔木层Simpson指数在森林死亡梯度由轻到重逐渐增加,灌木层随森林死亡梯度由轻到重先减小后增加,只有草本层Simpson指数由轻到重逐渐降低;Shannon-Wiener指数与Simpson指数趋势相同。不同林分乔木层Shannon-Wiener指数为0.47~0.63,不同林分灌木层Shannon-Wiener指数为0.41~0.66;样地内的丰富度指数均从大到小依次为轻度死亡林分、重度死亡林分、中度死亡林分,不同林分草本层丰富度指数为0.71~1.23,灌木层丰富度指数为0.28~0.47,乔木层丰富度指数为0.36~0.58。不同死亡样地乔木层均匀度指数从大到小依次为中度死亡林分、重度死亡林分、轻度死亡林分,不同林分乔木层均匀度指数为0.44~0.62。灌木层Pielou均匀度指数呈现出重度死亡林分、轻度死亡林分、中度死亡林分,不同林分灌木层Pielou均匀度指数为0.75~0.86。不同死亡梯度草本层均匀度指数从大到小依次为轻度死亡林分、重度死亡林分、中度死亡林分。总体而言,植物多样性从大到小依次为轻度死亡林分、重度死亡林分、中度死亡林分。

图 2 不同死亡梯度下植物群落α多样性指数
Figure 2. Alpha diversity index of vegetation communities under forests with different death gradients
-
β多样性分析表示多组样本之间的差异,这种差异可以通过群落样本之间的距离来表现,群落之间组成成分越相似,它们之间距离就越小。本研究对不同死亡梯度样地内的植物群落进行PCoA分析,如图3所示:群落之间的样本各自聚为一类,轻度死亡样地内的植物群落差异最大,第1主坐标可以解释样本总差异的43.66%,第2主坐标可以解释样本总差异的33.46%,说明3种样地在群落多样性之间存在显著差异(P<0.05)。
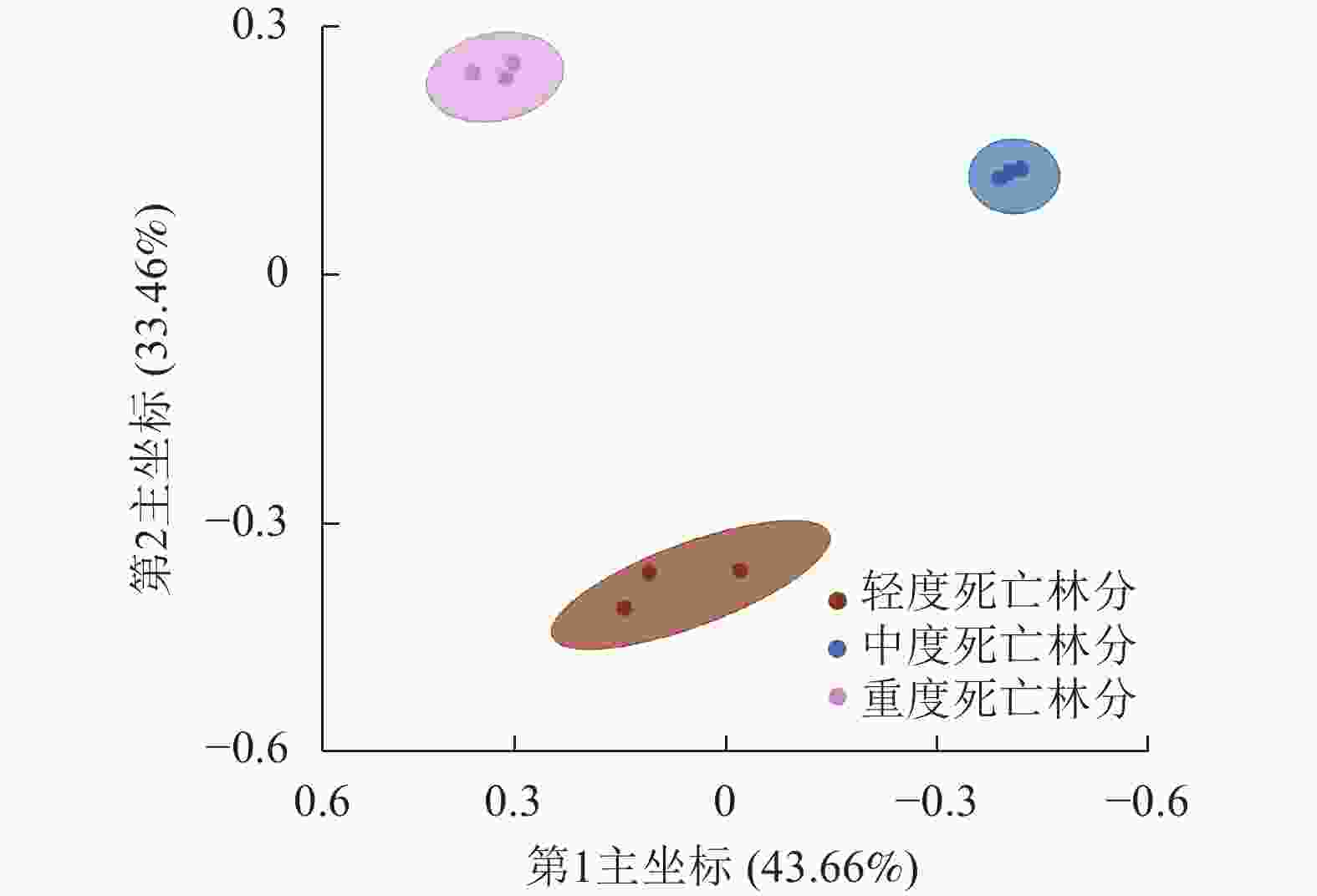
图 3 不同死亡梯度下植物群落主坐标分析
Figure 3. PCoA of vegetation communities under forests with different death gradients
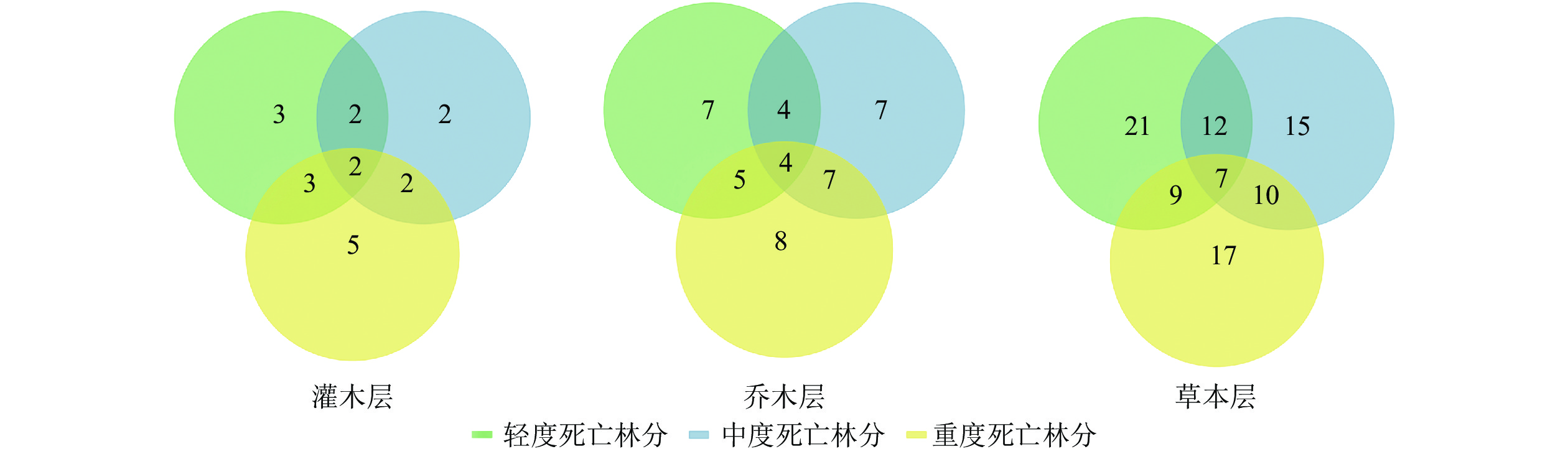
利用维恩图(图4)可清晰地看到不同群落之间的联系,统计发现样地内的植物物种数从大到小依次为轻度死亡林分、重度死亡林分、中度死亡林分。可以看出,从森林死亡梯度由轻到重,其共有种和植物物种数在中度死亡林分内最少,重度和轻度死亡林分内物种数量相同,3个层次的共有种共计13种植物。
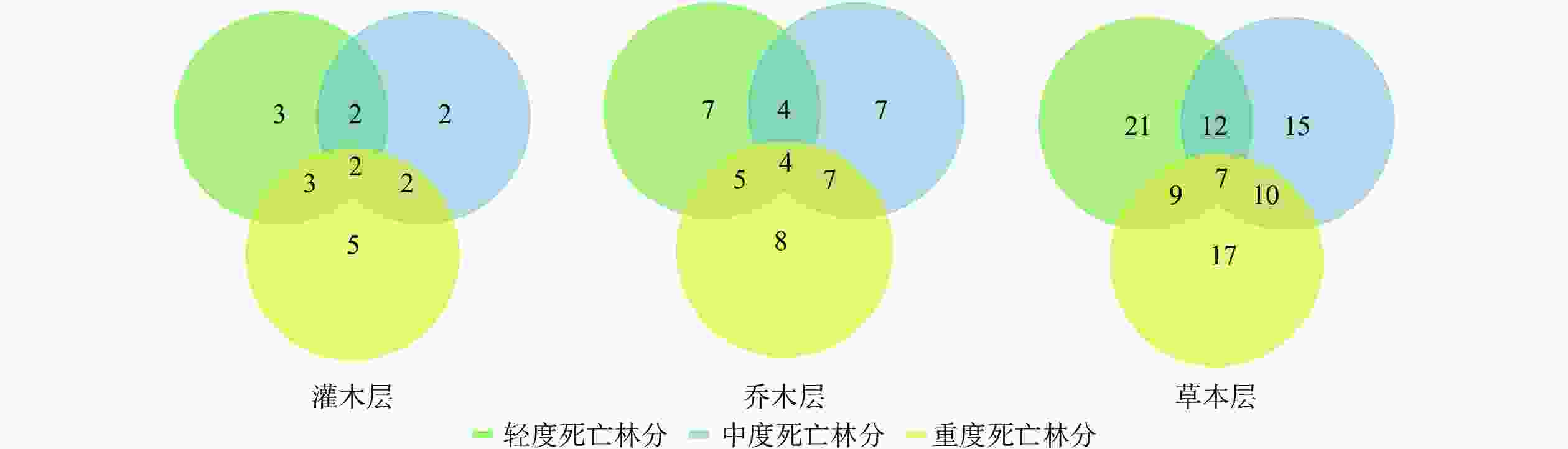
图 4 不同死亡梯度林下的植被群落物种数
Figure 4. Species number of vegetation community under forests with different death gradients
如表3所示:不同死亡梯度的干扰影响了林下群落植被组成的分布,不同死亡梯度林分内共有种为13种,SI指数为55%~61%。总体而言,乔木层和灌木层对相对系数的影响并不大,真正导致SI指数变化的是草本层,所以,不同的死亡程度对草本层的影响显著,森林死亡梯度由轻到重,其共有种和植物物种的相似系数逐渐降低,随着山杨死亡程度的增加植被组成差异也不断增加。
表 3 不同死亡梯度林分共有种和植物物种的相似系数
Table 3. Similarity coefficients of common species and plant species in different forest stands with different death gradients
死亡梯度 中度死
亡林分重度死
亡林分轻度死亡林分 55 61 中度死亡林分 54 -
植物多样性由群落内物种间关系和环境条件决定,植物多样性的差异在一定程度上也能够反映环境的变化[17]。本研究发现,所有样地内乔木层的山杨重要值随着死亡梯度由轻到重降低。在对样地内植物种类及数量的研究中发现:重度死亡样地内的乔木物种数较多,导致山杨在样地内的重要值下降。灌木层重要值最大的是中度死亡样地,林内大量乔木发生死亡导致灌木成为该群落的资源优胜者。草本层重要值最大的是重度死亡样地,森林死亡后形成林隙改变林下微环境,促使林下植物多样性及森林更新发生变化。以往研究发现:森林在死亡及形成倒木的过程中形成的林隙能够促进和加速原生老林的更新[18],并促使灌木得到生长释放[19−20],因此灌木在中度死亡林分重要值最大。在内蒙古大兴安岭南部林区,林隙更新是森林更新的主要方式,倒木和其他物质在森林中形成天然苗床,为幼苗发育提供栖息地和养分。林隙促使林内光照改善,温湿度较为稳定,使林木更新的机会增加[21]。倒木可以为森林植物提供养分,间接改善森林内的植物多样性[22]。森林死亡改变了林地微环境,驱动乔灌草不同层次的植物多样性,并且总体趋于多样性变差。
-
本研究中所有生物多样性指数均在轻度死亡样地最大。草本层的各多样性指数均大于灌木层和乔木层,与布买丽娅木·吐如汗[23]研究库车山区种子植物多样性海拔梯度格局的结果一致。草本层多样性指数高的原因可能是研究区属于林草过渡区,同时次生林区林分密度小,林下水分条件好,郁闭度低,提高了林下生境的异质性和植物多样性。每个生物群落都属于一个多优势种群落,这种优势会在以后的植被演替中进一步表现出来[24−25],因此群落具有较高的稳定性。但是由于森林死亡改变林下微环境等问题,打破了区域生态系统的稳定性,使生物多样性产生了波动性变化。本研究对不同死亡梯度林分群落各层的α多样性进行分析,发现草本层居于森林结构的最下层,其Simpson指数、Shannon-Wiener指数在轻度死亡林分最大,重度死亡林分最小。原因一是因为重度死亡样地的倒木对草本层有一定的地表遮荫拦截作用,致使草本生长受限,二可能是因为倒木倾倒破坏了植被,导致多样性降低。而中度死亡样地多数树木死亡,样地内的环境条件受限,从而草本植物也受到影响;灌木层的Simpson指数、Shannon-Wiener指数、丰富度指数均从大到小依次为重度死亡林分、轻度死亡林分、中度死亡林分。可能是因为重度死亡样地的倒木经过一定时间的分解增加了土壤养分,促进灌木生长,另外,倒木倾倒后光照增强,有利于灌木生长。而中度死亡样地多样性指数最小,可能是因为样地内环境条件受限,发生树木死亡,从而灌木生长也受到影响。丰富度指数和Pielou物种多样性指数也是轻度死亡样地最大,因为树木死亡会影响物种的平衡。β多样性轻度死亡样地内的物种差异最大,产生这种现象可能是因为随森林死亡时间的推移,林内积累了更多的凋落物,从而促进了草本植物的生长,这与刘兵兵[26]所得森林死亡后林下植物多样与凋落物厚度呈显著负相关的结论一致。但是林分不同垂直结构的物种生态位、生活习性(耐阴、喜阳)和繁殖更新方式等的差异,导致群落植物多样性对森林死亡梯度的响应差异明显。
-
青山保护区不同死亡梯度样地共有45种植物,其中乔木9种,灌木5种,草本31种,总体而言不同死亡梯度林分群落各层植物总数从大到小依次为轻度死亡林分、中度死亡林分、重度死亡林分, SI指数为55%~61%。森林死亡改变了林地微环境,从而驱动了乔灌草不同层次的物种组成变化,并且总体趋于多样性变差。
森林死亡总体降低了内蒙古罕山次生林区青山保护区生物多样性,尤其草本植物多样性的降低明显;灌木植物多样性在死亡初期降低,长时间尺度上有升高的趋势。森林死亡驱动乔木实现快速更新,提高了乔木植物多样性,但林内的更新能否形成混交林有待进一步研究。
Undergrowth plant diversity in the northern part of secondary forest in Hanshan Mountain, Inner Mongolia
-
摘要:
目的 青山保护区是内蒙古罕山次生林北段的典型区域,了解内蒙古罕山次生林北段森林死亡梯度下植物多样性特征,为科学保护和利用森林自生植物群落,探索更适宜森林发展的植物群落提供依据。 方法 以青山保护区山杨Populus davidiana次生林为研究对象,采用样地调查法对不同死亡梯度林下植物组成进行调查,使用生物多样性指数和群落相似系数对植物组成及多样性的差异进行分析。 结果 内蒙古罕山次生林区北段不同死亡梯度下样地植物有45种,其中,乔木9种,灌木5种,草本31种,样地内的植物物种数从大到小依次为轻度死亡样地、重度死亡样地、中度死亡样地。不同样地内的Simpson和Shannon-Wiener指数从大到小依次为轻度死亡林分、重度死亡林分、中度死亡林分。中度死亡样地的丰富度指数和Pielou均匀度指数最高。通过主坐标分析(PCoA)发现轻度死亡样地内的群落差异最大。Sorensen群落相似系数(SI)为55%~61%,不同死亡梯度林分内共有种为13种,乔木层和灌木层对SI的差异影响并不大,草本多样性的降低效益明显,导致不同死亡梯度林分SI差异变大。总体而言,随着山杨死亡程度的增加植被组成差异性也不断增加。 结论 森林死亡导致植物多样性总体趋于降低,森林大量死亡抑制了草本植物多样性,但是促进了乔木和灌木的更新。研究区山杨优势度出现衰退,林内更新能否形成混交林有待进一步研究。图4表3参26 Abstract:Objective Qingshan Reserve is a typical area in the northern section of the secondary forest in Hanshan, Inner Mongolia, in order to understand the characteristics of plant diversity under the forest death gradient in the northern section of the Hanshan secondary forest in Inner Mongolia, and to provide a basis for scientific protection and utilization of forest autogenous plant communities and explore plant communities more suitable for forest development. Method Taking the secondary forest of Populus davidiana in Qingshan Reserve as the research object, the plant composition of different dead gradients was investigated by sample survey method, and the differences in plant composition and diversity were analyzed by using biodiversity index and community similarity coefficient. Result There were 45 plant species in the northern section of Hanshan secondary forest area, Inner Mongolia, including 9 species of trees, 5 species of shrubs and 31 species of herbs, and the number of plant species in the sample plot was as follows: mild death plot>severe death plot>moderate death plot. The Simpson and Shannon-Wiener indices in the same field showed mildly dead stands>severely dead stands>moderately dead stands. Moderately dead plots had the highest richness and Pielou uniformity index. The greatest variation in communities within mildly fatal plots was found by PCoA analysis. The SI index was in the range of 55%−61%, there were 13 species in different death gradient forest stands, the difference between the relative coefficient of the tree layer and the shrub layer was not large, and the reduction benefit of herbaceous diversity was obvious, resulting in the difference of SI index of different death gradient forest stands became larger, and overall, with the increase of aspen mortality degree, the difference in vegetation composition also increased. Conclusion The mass forest mortality inhibits the development of herbaceous plant diversity, but promotes the renewal of trees and shrubs. The dominance of aspen in the study area declined, and whether the diversity of forest renewal could form mixed forests needed to be further studied. [Ch, 4 fig. 3 tab. 26 ref.] -
Key words:
- Daxinganling /
- Hanshan secondary forest /
- plant diversity /
- community /
- forest death
-
图 1 不同死亡梯度林下乔灌草重要值分布
Figure 1 Distribution of important values of arbor, shrub and grass under forests with different death gradients
图 2 不同死亡梯度下植物群落α多样性指数
Figure 2 Alpha diversity index of vegetation communities under forests with different death gradients
图 3 不同死亡梯度下植物群落主坐标分析
Figure 3 PCoA of vegetation communities under forests with different death gradients
图 4 不同死亡梯度林下的植被群落物种数
Figure 4 Species number of vegetation community under forests with different death gradients
表 1 样地基本信息
Table 1. Basic information of test plots
样地 纬度(N) 经度(E) 海拔/m 坡向 坡度/(°) 倒木密度/(株·hm−2) 枯立木密度/(株·hm−2) L1 45°57′13.78″ 121.21′26.53″ 668 西北 12 588±53 308±29 L2 45°57′12.82″ 121°21′27.51″ 710 西北 10 783±72 259±16 L3 45°57′13.23″ 121°21′26.35″ 721 西北 13 975±68 346±25 K1 45°57′12.67″ 121°21′29.44″ 708 西北 16 958±36 392±69 K2 45°57′12.77″ 121°21′30.41″ 686 西北 19 1053±88 352±59 K3 45°57′13.69″ 121°21′32.03″ 696 西北 12 896±65 453±73 D1 45°59′49.32″ 121°22′56.48″ 432 西北 18 1375±46 350±43 D2 45°59′52.47″ 121°22′58.04″ 772 西北 17 1029±69 296±69 D3 45°59′52.75″ 121°22′59.42″ 770 西北 23 1698±63 398±73 说明:L为轻度死亡林分,K为中度死亡林分,D为重度死亡林分。1、2、3为样地重复编号。  下载: 导出CSV
下载: 导出CSV
表 2 不同死亡梯度林分因子、环境因子特征
Table 2. Characteristics of forest factors and environmental factors in different regions
样地 凋落物厚度/cm 萌蘖更新/(株·hm−2) 活立木密度/(株·hm−2) 木质残体密度/(株·hm−2) 死亡率/% 郁闭度 L1 6.27±0.45 6 950 733.25±62.92 a 318.00±13.30 a 17.39 0.63 L2 5.87±0.45 3 975 756.43±54.56 a 329.00±18.30 a 16.65 0.66 L3 5.38±0.45 5 325 693.32±48.52 a 315.00±16.37 a 15.55 0.60 K1 6.48±0.43 10 125 588.56±56.76 b 483.35±187.65 a 46.54 0.69 K2 6.48±0.43 15 741 575.00±86.50 b 495.32±167.38 a 48.76 0.71 K3 6.48±0.43 15 158 538.53±67.63 b 465.54±187.64 a 45.25 0.78 D1 3.26±0.42 18 400 242.56±74.00 c 1467.65±343.15 b 43.93 0.55 D2 2.26±0.23 3 941 250.67±63.00 c 1591.46±245.31 b 45.17 0.56 D3 3.56±0.45 1 975 268.54±68.00 c 1591.85±413.36 b 42.83 0.61 说明:不同小写字母表示不同样地间差异显著(P<0.05)。
下载: 导出CSV
表 3 不同死亡梯度林分共有种和植物物种的相似系数
Table 3. Similarity coefficients of common species and plant species in different forest stands with different death gradients
死亡梯度 中度死
亡林分重度死
亡林分轻度死亡林分 55 61 中度死亡林分 54
下载: 导出CSV
-
[1] 吕康婷, 张二山, 李思颖, 等. 黄山松人工林林分空间结构对林下植物多样性的影响[J]. 浙江农林大学学报, 2022, 39(6): 1257 − 1266. LÜ Kangting, ZHANG Ershan, LI Siying, et al. The impact of the spatial structure of Pinus hwangshanensis plantation on undergrowth plant diversity [J]. Journal of Zhejiang A&F University, 2022, 39(6): 1257 − 1266. [2] CURRIE D J, PAQUIN V. Large-scale biogeographical patterns of species richness of trees [J]. Nature, 1987, 239: 326 − 327. [3] 张鸽, 王琦枝, 何媛, 等. 北川滑坡迹地植物群落多样性变化及环境解释[J]. 浙江农林大学学报, 2022, 39(1): 50 − 59. ZHANG Ge, WANG Qizhi, HE Yuan, et al. Change of plant community diversity and environmental interpretation of Beichuan landslide footprint [J]. Journal of Zhejiang A&F University, 2022, 39(1): 50 − 59. [4] 李敏, 王艳红, 唐荣, 等. 不同微生境下季风常绿阔叶林附生维管植物多样性特征[J]. 西北植物学报, 2021, 41(12): 2133 − 2141. LI Min, WANG Yanhong, TANG Rong, et al. Diversity characteristics of epiphytic vascular plant in monsoon tropical and subtropical moist broadleaf forests under different microhabitats [J]. Acta Botanica Sinica, 2021, 41(12): 2133 − 2141. [5] LU Jingzhao, LU Hongwei, BRUSSEAU M L, et al. Interaction of climate change, potentially toxic elements (PTEs), and topography on plant diversity and ecosystem functions in a high-altitude mountainous region of the Tibetan Plateau [J/OL]. Chemosphere, 2021, 275: 130099[2023-02-15]. doi:10.1016/j.chemosphere.2021.13009. [6] ANTONIO V G, THOMAS J G. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán [J]. Journal of Ecology, 1998, 86(6): 999 − 2010. [7] WEIHER E, WERF A, THOMPSON K, et al. Challenging theophrastus: a common core list of plant traits for functional ecology [J]. Journal of Vegetation Science, 1999, 10(5): 609 − 620. [8] SHEZI T, O’ CONNOR T, WITKOWSKI E. Impact of livestock grazing intensity on plant diversity of montane grassland in the northern Drakensberg, South Africa [J]. African Journal of Range &Forage Science, 2021, 38(1): 67 − 79. [9] LERZMAN K P, SUTHERLAND G D, INSELBERG A, et al. Canopy gaps and the landscape mosaic in a coastal Temperate rain forest [J]. Ecology, 1996, 77(4): 1254 − 1207. [10] 夏威, 安明态, 陈龙, 等. 贵州北盘江不同强度石漠化区木本植物多样性及生态功能群研究[J]. 西北植物学报, 2020, 40(3): 524 − 531. XIA Wei, AN Mingtai, CHEN Long, et al. Study on woody plant diversity and ecological functional groups in rocky desertification areas with different intensities in Beipan River, Guizhou [J]. Journal of Botany of Northwest China, 2020, 40(3): 524 − 531. [11] 陈灵芝. 生物多样性保护现状及其研究[J]. 植物杂志, 1993(5): 7 − 9. CHEN Lingzhi. Current status and research on biodiversity conservation [J]. Journal of Botany, 1993(5): 7 − 9. [12] 刘斌, 张参参, 汪金松, 等. 江西九连山不同恢复模式林分的物种多样性特征[J]. 林业科学研究, 2020, 33(4): 42 − 52. LIU Bin, ZHANG Shenshen, WANG Jinsong, et al. Characteristics of species diversity of different restoration model stands in Jiulian Mountain, Jiangxi Province [J]. Forestry Science Research, 2020, 33(4): 42 − 52. [13] 赵中华, 白登忠, 惠刚盈, 等. 小陇山不同经营措施下次生锐齿栎天然林物种多样性研究[J]. 林业科学研究, 2013, 26(3): 326 − 331. ZHAO Zhonghua, BAI Dengzhong, HUI Gangying, et al. Species diversity of Quercus aliena var. acuteserrata natural forest with different management measures at Xiaolongshan [J]. Forestry Science Research, 2013, 26(3): 326 − 331. [14] 高贤明, 马克平, 陈灵芝. 暖温带若干落叶阔叶林群落物种多样性及其与群落动的关系[J]. 植物生态学报, 2001, 25(3): 283 − 290. GAO Xianming, MA Keping, CHEN Lingzhi. Species diversity of some deciduous broad-leaved forest communities in warm temperate zone and its relationship with community animals [J]. Journal of Plant Ecology, 2001, 25(3): 283 − 290. [15] 王兵, 郑秋红, 郭浩. 基于Shannon-Wiener指数的中国森林物种多样性保育价值评估方法[J]. 林业科学研究, 2008, 21(2): 268 − 274. WANG Bing, ZHENG Qiuhong, GUO Hao. Evaluation method for conservation value of species diversity of forests in China based on Shannon Wiener index [J]. Forestry Science Research, 2008, 21(2): 268 − 274. [16] 曾楠. 大兴安岭南段阔叶林乔木死亡动态与生态系统响应[D]. 呼和浩特: 内蒙古农业大学, 2017. ZENG Nan. Dynamics of Tree Death and Ecosystem Response of Broad-Leaved Forest in the Southern Section of Greater Khingan [D]. Huhhot: Inner Mongolia Agricultural University, 2017. [17] 葛红杰, 张峰. 汾河中下游湿地植被物种多样性研究[J]. 西部林业科学, 2008, 37(2): 29 − 33. GE Hongjie, ZHANG Feng. Study on species diversity of wetland vegetation in the middle and lower reaches of Fenhe River [J]. Western Forestry Science, 2008, 37(2): 29 − 33. [18] 康玲. 内蒙古大兴安岭兴安落叶松与白桦倒木分解研究[D]. 呼和浩特: 内蒙古农业大学, 2012. KANG Ling. Study on the Decomposition of Fallen Trees of Larix gmelinii and Betula platyphylla in Greater Khingan, Inner Mongolia [D]. Huhhot: Inner Mongolia Agricultural University, 2012. [19] 刘少冲, 段文标, 冯静, 等. 林隙对小兴安岭阔叶红松林树种更新及物种多样性的影响[J]. 应用生态学报, 2011, 22(6): 1381 − 1388. LIU Shaochong, DUAN Wenbiao, FENG Jing, et al. The impact of gaps on species regeneration and species diversity of broad-leaved korean pine forest in Lesser Khingan [J]. Journal of Applied Ecology, 2011, 22(6): 1381 − 1388. [20] 刘兵兵, 赵鹏武, 周梅, 等. 林窗对大兴安岭南段杨桦次生林林下更新特征的影响[J]. 林业资源管理, 2019(4): 31 − 36. LIU Bingbing, ZHAO Pengwu, ZHOU Mei, et al. The impact of forest gaps on the regeneration characteristics of poplar birch secondary forests in the southern section of Greater Khingan [J]. Forestry Resources Management, 2019(4): 31 − 36. [21] 赵鹏武, 海龙, 宋彩玲, 等. 大兴安岭北部兴安落叶松原始林倒木研究[J]. 干旱区资源与环境, 2010, 24(3): 173 − 177. ZHAO Pengwu, HAI Long, SONG Cailing, et al. Study on the fallen wood of Xing’ an Larix gmelinii primitive forest in the north of Greater Khingan [J]. Resources and Environment in Arid Areas, 2010, 24(3): 173 − 177. [22] DOMINIQUE F, LOUIS I, PIERRE D, et al. Small mammal responses to coarse woody debris distribution at different spatial scales in managed and unmanaged boreal forests [J]. Forest Ecology and Management, 2012, 266(2): 194 − 205. [23] 布买丽娅木·吐如汗. 库车山区种子植物多样性海拔梯度格局研究[D]. 乌鲁木齐: 新疆师范大学, 2017. Bumaliyamu Turuhan. Study on the Altitude Gradient Pattern of Spermatophyte Diversity in Kuqa Mountain Area [D]. Urumqi: Xinjiang Normal University, 2017. [24] 郑昌龙, 王健铭, 李景文. 北京松山国家级自然保护区不同林龄胡桃楸林下植物多样性[J]. 浙江农林大学学报, 2017, 34(5): 825 − 832. ZHENG Changlong, WANG Jianming, LI Jingwen. Diversity of undergrowth plants of Juglans mandshurica at different ages in Songshan National Nature Reserve [J]. Journal of Zhejiang A&F University, 2017, 34(5): 825 − 832. [25] 毛齐正, 马克明, 邬建国, 等. 城市生物多样性分布格局研究进展[J]. 生态学报, 2013, 33(4): 1051 − 1064. MAO Qizheng, MA Keming, WU Jianguo, et al. Research progress on the distribution pattern of urban biodiversity [J]. Journal of Ecology, 2013, 33(4): 1051 − 1064. [26] 刘兵兵. 内蒙古罕山次生林区山杨死亡后物种组成及山杨更新特征[D]. 呼和浩特: 内蒙古农业大学, 2020. LIU Bingbing. Species Composition and Regeneration Characteristics of Populus davidiana after Death in Hanshan Secondary Forest Area, Inner Mongolia [D]. Huhhot: Inner Mongolia Agricultural University, 2020. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230204
 点击查看大图
点击查看大图
计量
- 文章访问数: 1342
- HTML全文浏览量: 372
- PDF下载量: 47
- 被引次数: 0
