





-
生态化学计量学是通过研究不同比例的化学元素以揭示生态过程和作用的学科。利用生态化学计量学,可以研究生态系统中各种化学元素[特别是碳(C)、氮(N)、磷(P)]的平衡关系。碳为构成有机物的最基本元素,是植物生长发育过程中不可缺少的一部分。氮和磷为蛋白质和核酸的特征元素,常常是植物生长的限制性元素。因此,通过研究植物与土壤的化学计量关系,可以了解植物所需营养元素和土壤中有效元素之间的比例关系。开展植物、凋落物和土壤的生态化学计量研究,能为判断植物限制性元素、土壤营养的供给能力、养分的有效性及碳、氮、磷对环境变化的适应和反馈调节能力提供重要科学依据。在气候变化背景下,晋西黄土区普遍存在植被退化、土地退化、水分亏缺等多种生态问题[1],因此,研究植被恢复生态系统,可为改善晋西黄土区土壤质量、植被覆盖率等多种生态问题提供科学指导[2−4]。研究表明:植被恢复在全球碳循环中扮演着重要角色,是全球气候变化的重要组成部分和陆地生态系统碳循环过程的关键因素[5−7]。采用生态化学计量学的原理和方法,围绕不同植被恢复类型对土壤养分的影响,研究生态系统中碳、氮、磷元素的变化与平衡[8],为理解生态系统中生物体与自然环境相互作用关系及养分循环过程提供了有效手段。国内学者开展了大量的试验研究,并在陆地生态系统中植物和土壤属性关系方面取得了显著成就。土壤中碳、氮、磷质量分数可决定植被生长状态,而植被通过凋落物和根系分泌物的输入影响土壤养分[9]。近年来,大量研究探讨了草原、农田和森林陆地生态系统中C∶N∶P化学计量特征。JIANG等[10]对鄱阳湖农田土壤化学计量特征研究发现:氮是决定土壤碳氮比和氮磷比的主要因素,土壤碳磷比主要受磷的影响。杨阔等[11]研究发现:青藏高原植物叶片氮、磷质量分数呈显著正相关,且氮磷比是判断植物生长受限的关键因子。SONG等[12]研究发现:在植被演替过程中,土壤有机碳、全氮质量分数总是同步增加,而土壤全磷质量分数则无显著变化。这些研究都是针对植物叶片或土壤的生态化学计量特征进行的,但对土壤-植物系统中的生态化学计量特征的研究较少,主要集中于区域尺度上叶片与土壤化学计量特征,对于植物其他器官如树干、树皮、地上部分、地下部分的化学计量特征及其与土壤关系研究较为缺乏,需进一步研究[13]。
植被恢复生态系统的化学计量关系的形成是一个长期过程,不同地区和不同类型的生态系统具有不同的植被组成和环境条件,需要高质量的数据支撑研究。目前,植被恢复生态系统的化学计量关系还缺乏综合性的研究和长期观察。同时,对于不同植被恢复类型下生态系统的恢复时间和恢复能力的研究也比较有限。经过不断治理,黄土高原形成了以自然恢复植被和人工恢复植被为主的多种植被类型[14]。自然恢复状态下,植被生态系统中的植被和土壤之间形成了一个复杂的养分循环过程[15−16]。
为了解黄土高原植被恢复中植物与土壤的变化特征,前人对不同植被恢复类型下土壤理化性质、植被特征以及各层次的化学计量特征等进行了系统的研究,但关于晋西黄土区植被和土壤养分循环与生态系统乔灌草层次植被属性相互关系的研究甚少。基于此,本研究在晋西黄土区开展野外定位试验,研究植被恢复类型对生态系统植物层次和土壤化学计量的综合影响,为黄土高原植被恢复实践提供理论参考和科学支撑。
-
研究区设在山西吉县森林生态系统国家野外科学观测研究站,位于黄土高原东部山西省临汾市内,35°53′~36°21′N,110°27′~111°07′E,属于暖温带半湿润地区。该区属黄土高原残垣沟壑区,年平均气温为9.9 ℃,年平均日照时数为2 563.8 h,年平均降水量为579.1 mm,降水集中在6—9月,年平均蒸发量为1 729.0 mm,年平均无霜期为172.0 d。
研究区地势东高西低,海拔为440~1 820 m。海拔1 350 m以下为典型黄土高原侵蚀地貌。土壤类型为褐土,可分3个亚类:丘陵褐土、普通褐土和淋溶褐土。常见的木本植物有194种,分属49科;草本植物180种,分属44种(不包括农作物)。天然次生林植被主要有辽东栎Quercus mongolica、山杨Populus davidiana、旱柳Salix matsudana等;人工林植被主要有油松Pinus tabuliformis、刺槐Robinia pseudoacacia、侧柏Platycladus orientalis等;农作物主要有玉米Zea mays、小麦Triticum aestivum等;下层灌木有沙棘Hippophae rhamnoides、山桃Prunus davidiana、胡枝子Lespedeza bicolor、三裂绣线菊Spiraea trilobata等。
-
在2017年8月采样,依据代表性和典型性原则,在研究区内选择油松人工林、刺槐人工林、侧柏人工林、辽东栎天然林等4种典型植被恢复类型。在4种植被恢复类型中,选择具代表性的区域,分散随机选择5个试验小区,共20个试验小区。试验小区基本特征见表1。
表 1 不同植被恢复类型下试验小区基本情况特征
Table 1. Basic characteristics of experimental plot under different vegetation restoration types
植被恢复类型 坐标 海拔/
m坡度/
(°)坡向 林龄/
a林分密度/
(株·hm−2)林下优势植被 油松人工林 36°16′ 24.36″N,110°45′ 29.65″E 1148 26 南 30 1650 沙棘、山桃、低矮薹草等 侧柏人工林 36°16′ 39.00″N,110°44′ 57.10″E 1210 25 南 30 1800 黄荆、山桃、白颖薹草、细裂叶莲蒿等 刺槐人工林 36°16′ 23.11″N,110°45′ 41.14″E 1135 24 南 30 1850 沙棘、山桃、泥胡菜、白颖薹草等 辽东栎天然林 36°44′ 39.34″N,110°44′ 38.80″E 1081 23 南 32 1050 沙棘、山桃、黄荆、低矮薹草、细裂叶莲蒿等 说明:低矮薹草Carex humilis;黄荆Vitex negundo;白颖薹草Carex duriuscula subsp. rigescens;细裂叶莲蒿Artemisia stechmanniana;泥胡菜Hemisteptia lyrata。 植被取样及测定方法:在试验小区中,通过分析种-面积曲线,确定各层次物种多样性增长趋缓的样地尺寸,同时考虑到研究的可行性,最终选定的样地尺寸能较好地反映区域的生物多样性[17]。在试验小区内,乔木层随机设置4个10 m×10 m的调查样方,记录树种、生物量、树高、胸径、冠幅等因子;灌木层随机设置4个5 m×5 m的调查样方,记录灌木的生物量、株高、基径、盖度等因子;草本层随机设置4个1 m×1 m的调查样方,调查草本的生物量、株高、基径、盖度等因子。
乔木层采样:采用标准木解析法。在所选调查样方内,选取胸径在平均值附近的3株立木作为平均标准木。将解析木在距离地面0.3 m处伐倒,并测定胸径、枝下高、树高和树高1/4、1/2、3/4处的直径。测定树高的同时,将树干分成若干段,采用中央断面区分求积法在每个区分段的中点位置截取圆盘(其中乔木按树干、树枝、树根截取),并测定每个圆盘的带皮鲜质量和去皮鲜质量。同时,分离地上部分的叶、枝、皮等部分,分别称取鲜质量后,随机抽取一定量圆盘,装袋带回实验室,70~80 ℃烘干至恒量后称量,经研磨、过筛后测定元素质量分数。
灌木层采样:采取全收获法,选择生物量生长高峰期月份采集。选取4个调查样方,采用每木检尺法在样方内按平均直径、株高等选取5株形态大小接近的优势种标准株(丛),分别齐面收割,并挖出地下部分,随机抽取1 kg样品带回实验室,70~80 ℃烘干后,经研磨、过筛后进行元素测定。
草本层采样:采用全收获法,选择生物量生长高峰期月份采集。在调查样方内,按照植物物种,收割每一样地的草本植物,并称取其质量。取混合样1 kg带回烘干,经研磨、过筛后进行元素测定。
土壤样品收集与测定方法:在试验小区中,分散随机选择5处土壤剖面,使用土钻法采集土壤样品,在0~100 cm设置6个土层(0~10、10~20、20~40、40~60、60~80、80~100 cm),用100 cm3环刀在每层中间取原状土样。每次取样后用取样袋带回实验室进行理化性质测定。以上样品经105 ℃烘干12 h,研磨、过筛等以备元素测定。植物和土壤全碳质量分数采用重铬酸钾外加热法测定,植物和土壤全氮质量分数采用凯氏蒸馏法测定,植物和土壤全磷质量分数采用氢氧化钠熔融-钼锑抗比色法测定。
-
测定4种植被恢复类型下土壤各土层的水分和碳、氮、磷质量分数并进行化学计量分析,利用SPSS 25.0进行单因素方差分析(ANOVA),采用描述性统计和方差分析等方法处理数据,相关数据采用平均值±标准差表示,同一因素不同水平间差异显著性采用最小显著极差法(LSD)检验,显著性水平为P<0.05。Sigma Plot 12.5绘图。
建立土壤-植被结构矩阵和土壤-植被功能属性矩阵,并使用Mantel Test (MT)分析典型植被生态系统植物要素和土壤要素之间的相互影响机制和构效关系。采用R软件进行冗余分析(RDA)排序,分析植被属性与土壤、植被元素的相互关系。
-
从表2可以看出:不同植被恢复类型下各层次植被属性存在显著差异,但各属性总体上呈波动性和表聚性。在乔木层中,油松人工林地上生物量、胸径和冠幅显著高于其余3种植被(P<0.05),分别为(125.79±3.63) t·hm−2、(18.19±1.16) cm、(28.38±1.59) dm;油松人工林高度低于刺槐人工林和辽东栎天然林,略高于侧柏人工林,为(6.91±0.21) m;地下生物量显著高于辽东栎天然林(P<0.05),为(24.28±1.86) t·hm−2,但与刺槐人工林和侧柏人工林无显著差异。侧柏人工林地下生物量与油松人工林相似,为(25.86±1.68) t·hm−2;乔木层高度、胸径和冠幅在4种植被中最小,分别为(6.37±0.43) m、(8.48±0.88) cm、(10.83±1.80) dm。刺槐人工林和辽东栎天然林的植被属性较为相似,表现为地上生物量、乔木层高度、胸径无显著差异,而辽东栎天然林的地下生物量和冠幅均低于刺槐人工林,为(18.74±0.70) t·hm−2、(14.67±2.06) dm。
表 2 不同植被恢复类型下各层次植被属性比较
Table 2. Comparison of vegetation attributes at different levels under different vegetation restoration types
林层 植被恢复类型 地上生物量/(t·hm−2) 地下生物量/(t·hm−2) 各层次高度/m 胸径或基径/cm 冠幅/dm 乔木层 油松人工林 125.79±3.63 a 24.28±1.86 a 6.91±0.21 b 18.19±1.16 a 28.38±1.59 a 刺槐人工林 71.25±2.25 c 25.09±2.02 a 8.60±0.42 a 11.19±0.64 b 17.30±3.09 b 侧柏人工林 78.26±3.91 b 25.86±1.68 a 6.37±0.43 a 8.48±0.88 b 10.83±1.80 b 辽东栎天然林 71.87±1.55 c 18.74±0.70 b 8.91±1.03 b 11.16±2.64 c 14.67±2.06 c 灌木层 油松人工林 4.74±0.16 d 2.91±0.11 b 1.25±0.44 ab 1.38±0.94 a 5.44±3.63 a 刺槐人工林 6.60±0.16 c 1.09±0.06 d 1.09±0.24 ab 1.22±0.23 a 4.51±2.23 a 侧柏人工林 7.52±0.72 b 1.92±0.10 c 0.91±0.21 b 0.73±0.19 a 4.57±3.09 a 辽东栎天然林 8.62±0.78 a 5.91±0.18 a 1.59±0.29 a 1.17±0.16 a 6.02±2.48 a 草本层 油松人工林 1.55±0.05 d 0.61±0.03 d 0.13±0.03 b 1.61±1.73 ab 1.88±1.46 b 刺槐人工林 3.06±0.24 b 1.43±0.06 b 0.16±0.03 b 3.24±1.07 a 3.42±0.93b 侧柏人工林 2.43±0.23 c 1.03±0.08 c 0.16±0.03 b 0.65±0.62 b 2.85±2.22 b 辽东栎天然林 3.65±0.36 a 3.54±0.25 a 0.44±0.14 a 1.65±1.05 ab 9.07±4.24 a 说明:不同植被恢复类型下各层次植被属性均为平均值±标准差;不同小写字母表示相同林层不同植被恢复类型之间差异显著(P<0.05)。 在灌木层中,油松人工林的地上生物量显著低于其余3种植被(P<0.05),为(4.74±0.16) t·hm−2;地下生物量显著低于辽东栎天然林(P<0.05),略高于刺槐人工林和侧柏人工林,为(2.91±0.11) t·hm−2;灌木层高度和基径与其余3种植被无显著差异。刺槐人工林和侧柏人工林植被的地下生物量和冠幅低于其余2种植被,分别为(1.09±0.06) t·hm−2、(4.57±3.09) dm;灌木层高度和基径与其余2种植被无显著差异。辽东栎天然林的植被属性较为突出,表现为除基径、灌木层高度、冠幅与其余3种植被无显著差异外,地上和地下生物量均显著高于其余3种植被(P<0.05),分别为(8.62±0.78) t·hm−2、(5.91±0.18) t·hm−2。
在草本层中,油松人工林的地上生物量、地下生物量、草本层高度和冠幅在4种植被中最小,分别为(1.55±0.05) t·hm−2、(0.61±0.03) t·hm−2、(0.13±0.03) m、(1.88±1.46) dm,基径与辽东栎天然林无显著差异。刺槐人工林和侧柏人工林,除基径外,地上生物量、草本层高度和冠幅均无显著差异,而刺槐人工林的基径显著高于其余2组植被(P<0.05),为(3.24±1.07) cm。侧柏人工林的基径显著低于刺槐人工林(P<0.05),为(0.65±0.62) cm。辽东栎天然林的植被属性较为突出,表现为除基径与油松人工林无显著差异外,地上生物量、地下生物量、草本层高度和冠幅均显著高于其他3种植被(P<0.05),分别为(3.65±0.36) t·hm−2、(3.54±0.25) t·hm−2、(0.44±0.14) m、(9.07±4.24) dm。
-
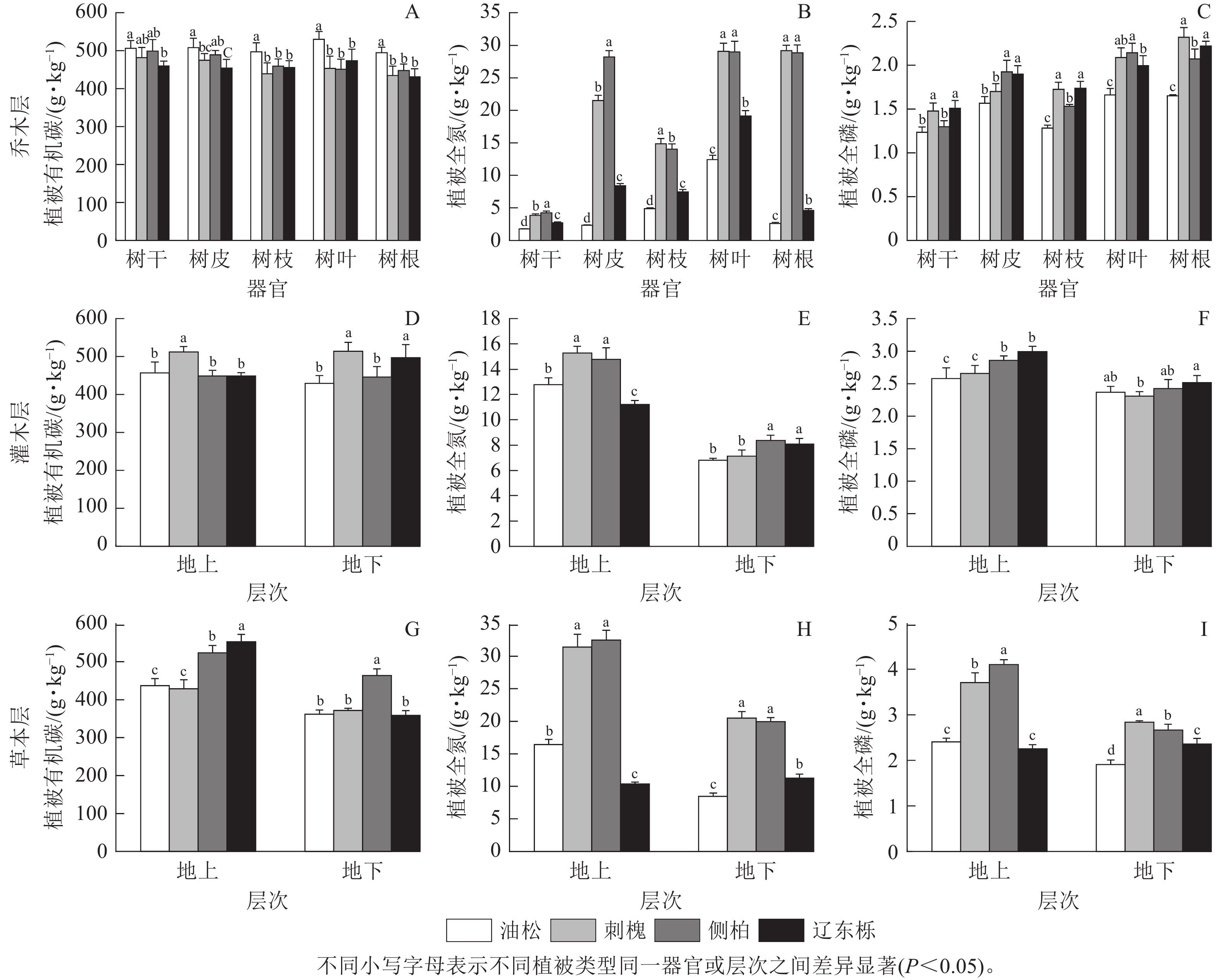
由图1可见:在乔木层中,不同植被恢复类型下,各器官的乔木层有机碳质量分数、全磷质量分数基本无显著差异;树干、树皮、树枝的全氮质量分数均存在显著差异(P<0.05)。

图 1 不同植被恢复类型不同层次下器官养分性质比较
Figure 1. Comparison of nutrient properties of organs under different layers of different vegetation restoration types
由图1 A~C可知:在乔木层中,油松人工林树枝、树叶和树根的有机碳质量分数显著高于其他植被恢复类型(P<0.05),分别为 (497.13±23.56)、(529.99±20.57)、(494.84±14.50) g·kg−1;在树干和树皮中,侧柏人工林的全氮质量分数显著高于其他植被恢复类型(P<0.05),分别为(4.25±0.26)、(28.21±0.96) g·kg−1,在树叶和树根中,刺槐人工林和侧柏人工林的全氮质量分数显著高于其他植被恢复类型(P<0.05)。在树枝中,刺槐人工林和辽东栎天然林的全磷质量分数均显著高于其他植被恢复类型(P<0.05),分别为(1.73±0.03)、(1.67±0.03) g·kg−1。
由图1 D~F可知:在灌木层中,刺槐人工林的地上部分有机碳质量分数均显著高于其他植被恢复类型(P<0.05),为(512.25±14.36) g·kg−1。刺槐人工林和侧柏人工林的地上部分全氮质量分数显著高于其他植被恢复类型(P<0.05),分别为(15.29±0.54) 、(14.67±0.11) g·kg−1;侧柏人工林和辽东栎天然林的地下部分全氮质量分数显著高于其他植被恢复类型(P<0.05),分别为(8.31±0.02) 、(7.90±0.05) g·kg−1。辽东栎天然林的地下部分全磷质量分数显著高于刺槐人工林(P<0.05),为(2.53±0.01) g·kg−1。
由图1 G~I可知:在草本层中,辽东栎天然林地上部分有机碳质量分数显著高于其他植被恢复类型(P<0.05),为(552.90±19.33) g·kg−1;侧柏人工林地下部分有机碳质量分数显著高于其他植被恢复类型(P<0.05),为(463.97±17.15) g·kg−1。刺槐人工林和侧柏人工林地上部分和地下部分全氮质量分数均显著高于其他植被恢复类型(P<0.05),刺槐人工林和侧柏人工林地上部分全氮质量分数分别为(31.50±0.10)、(33.39±0.08) g·kg−1 ,刺槐人工林和侧柏人工林地下部分全氮质量分数分别为(20.71±0.03)、(19.27±0.02) g·kg−1 。侧柏人工林地上部分全磷质量分数显著高于其他植被恢复类型(P<0.05),为(4.10±0.10) g·kg−1;刺槐人工林地下部分全磷质量分数显著高于其他植被恢复类型(P<0.05),为(2.84±0.03) g·kg−1。
-
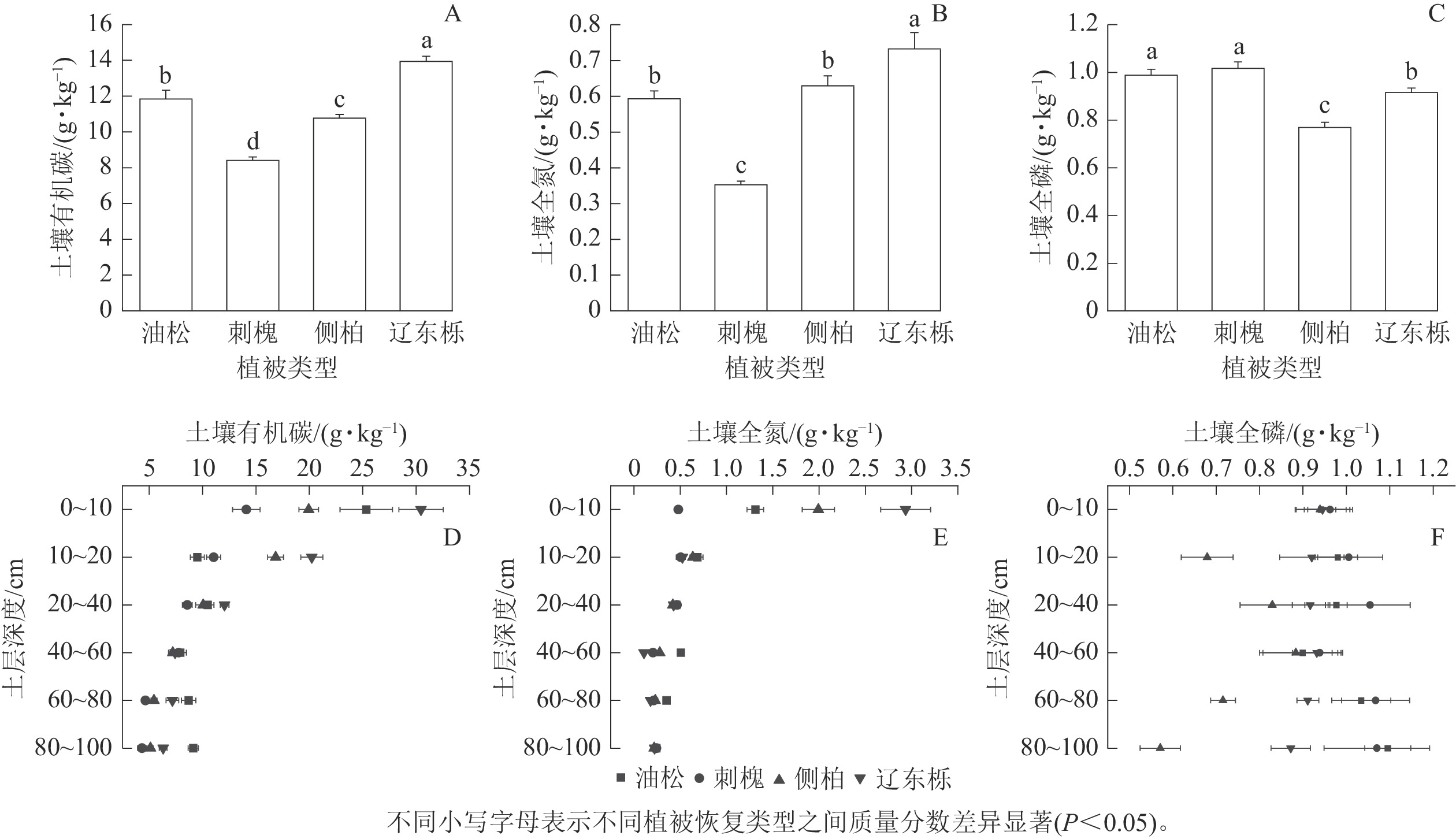
由图2可知:土壤有机碳、全氮、全磷质量分数在4种典型植被恢复类型间存在显著差异。辽东栎天然林土壤有机碳、全氮质量分数显著高于人工林(P<0.05),刺槐人工林土壤有机碳、全氮质量分数显著低于其他2种植被恢复类型(P<0.05,图2A~B)。油松人工林和刺槐人工林土壤全磷质量分数显著高于侧柏人工林和辽东栎天然林,但彼此间无显著差异(图2C)。
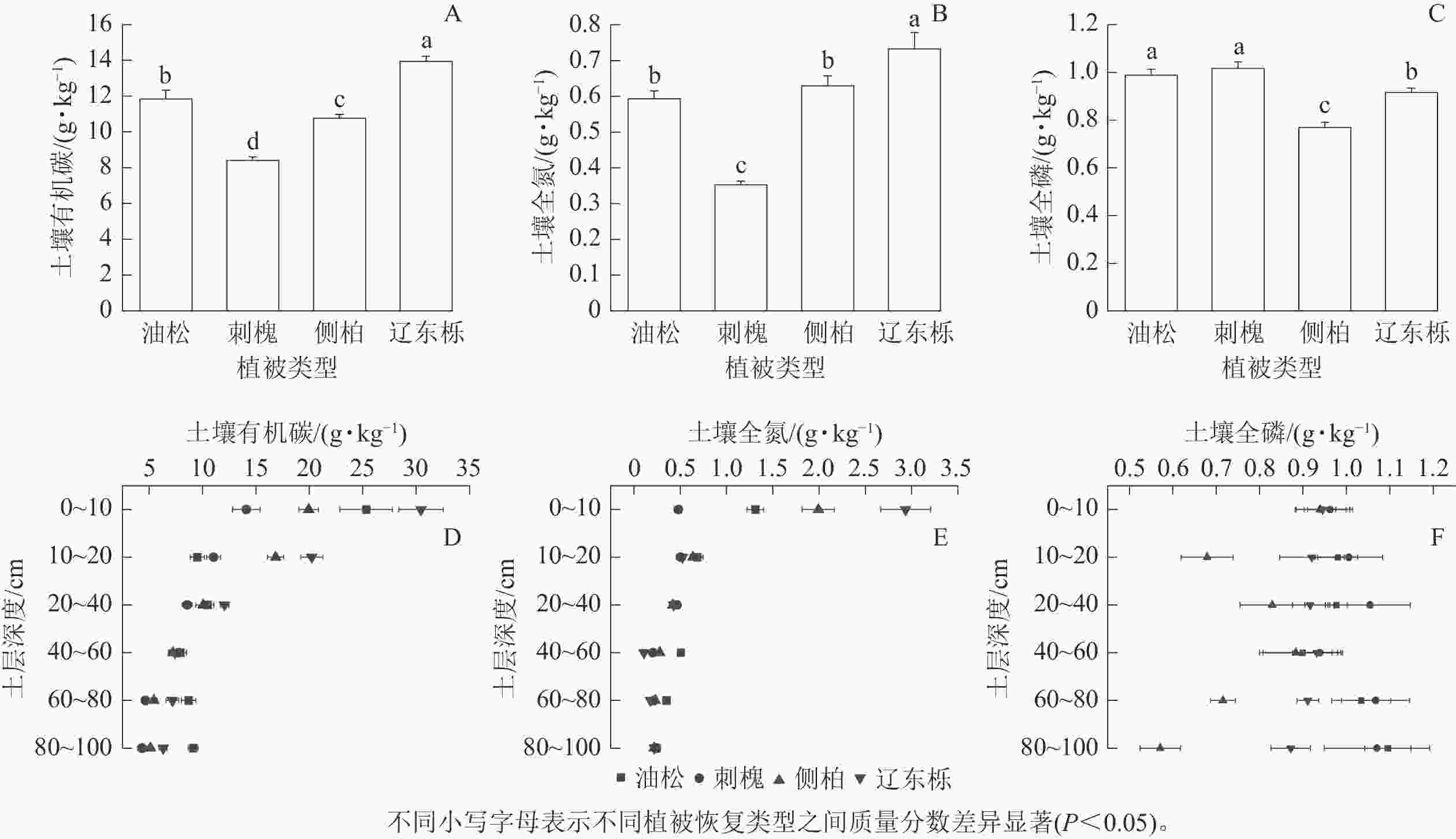
图 2 不同植被恢复类型及不同土层深度土壤有机碳、全氮、全磷质量分数
Figure 2. SOC, TN and TP mass fractions of different vegetation restoration types and different soil depth
由图2D~F可知:4种植被恢复类型0~20 cm处土壤有机碳高于20~60 cm处,60~100 cm土壤有机碳质量分数又有所增加,但不高于0~20 cm。可见各土层土壤有机碳质量分数从高到低依次为0~20、20~60、60~100 cm。4种植被恢复类型0~20 cm土壤全氮质量分数相对较高,但在20~100 cm,全氮质量分数总体呈现逐渐降低的趋势,全氮质量分数从高到低依次为0~20、20~60、60~100 cm。与土壤有机碳和全氮相比,4种植被恢复类型土壤中全磷质量分数在各土层变化较为复杂,可能会随着深度而增加或降低,且不同树种变化趋势也可能不同。
由图3可知:土壤化学计量比在4种植被恢复类型间存在显著差异。由图3A和图3B可知:刺槐人工林的碳氮比显著高于其他植被恢复类型(P<0.05),辽东栎天然林的碳磷比显著高于其他植被恢复类型(P<0.05)。由图3C可知:辽东栎天然林和侧柏人工林土壤氮磷比显著高于油松人工林和刺槐人工林(P<0.05),但彼此间无显著差异,刺槐人工林显著低于其他3种植被恢复类型(P<0.05)。
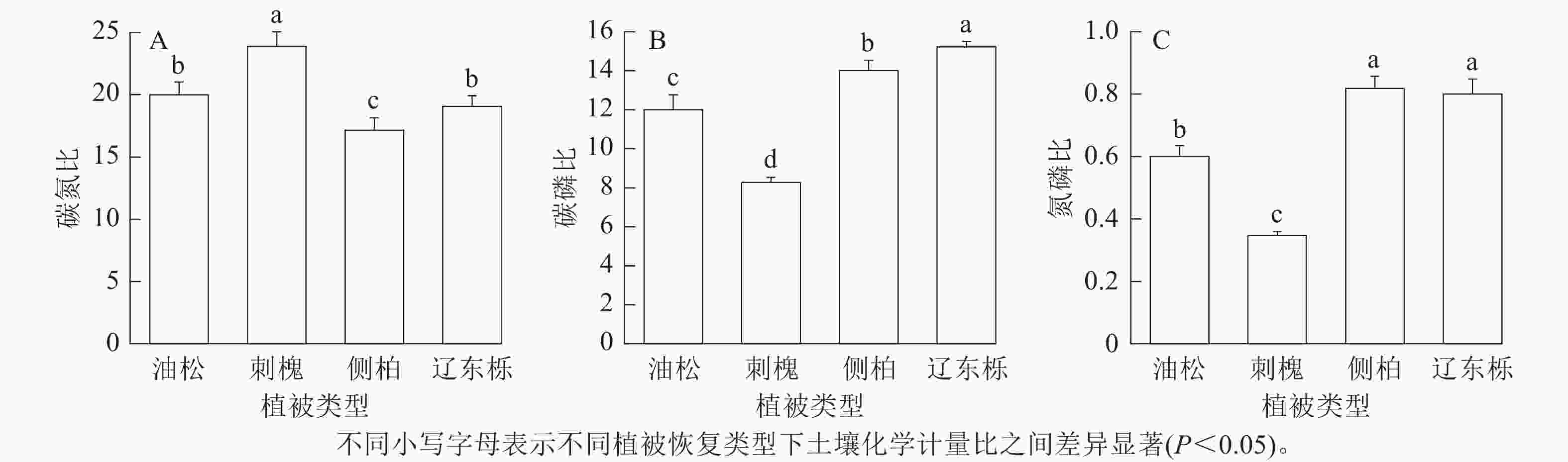
图 3 不同植被恢复类型下土壤化学计量比
Figure 3. Comparison of soil stoichiometric characteristics under different vegetation restoration types
-
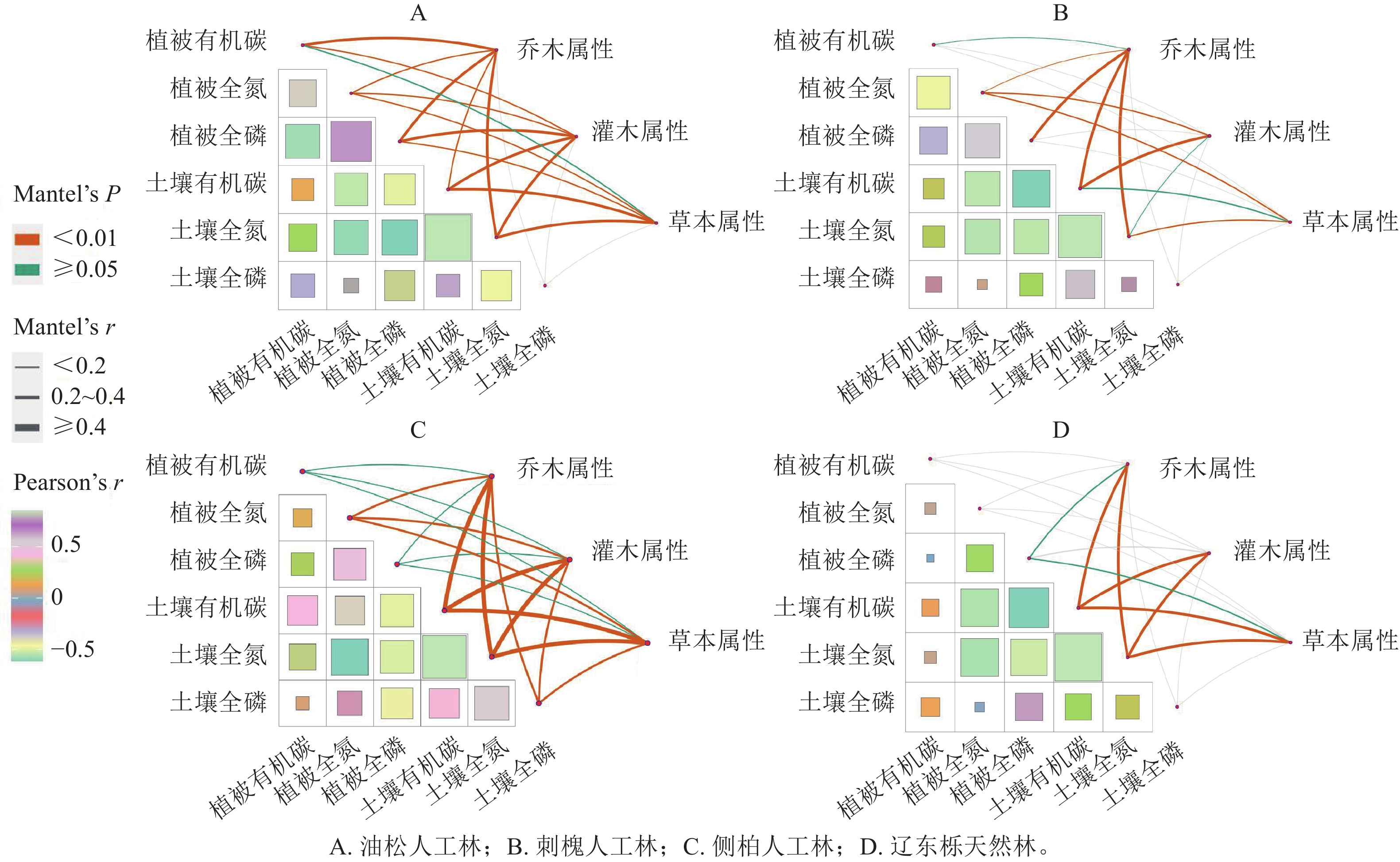
通过Pearson相关分析发现:在4种植被恢复类型条件下,植物全氮质量分数与土壤全氮质量分数显著相关(P<0.05),但由图4A~C比较图4D可知:人工林的各相关系数要显著高于辽东栎天然林。人工林植被有机碳质量分数与土壤有机碳质量分数的相关系数也显著高于辽东栎天然林(P<0.05)。另外,人工林植被全磷与土壤全氮之间的相关系数显著高于辽东栎天然林(P<0.05)。
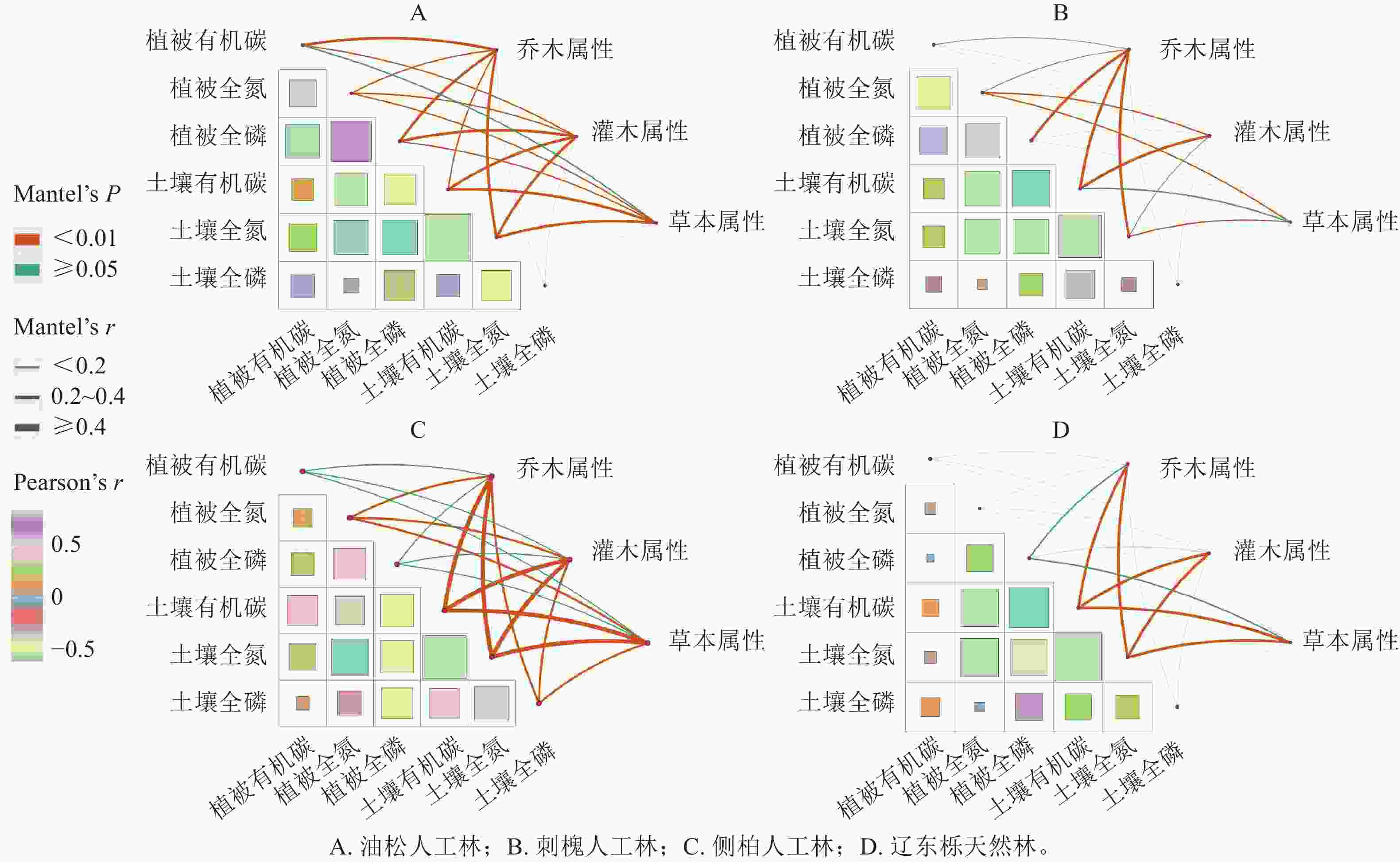
图 4 不同植被恢复类型下植被和土壤养分与各层次植被属性相互关系
Figure 4. Relationship between vegetation and soil nutrients and vegetation attributes of each layer of ecosystem under different vegetation restoration types
由图4可知:人工林各植被层次对养分分布的影响均较强,其中,乔木层对养分分布的影响最强,而人工林各植被层对土壤全磷和植物全磷仅有微量影响。在辽东栎天然林中,土壤有机碳、土壤全氮、土壤全磷的关系相对密切,特别是土壤全氮和土壤有机碳呈显著正相关(P<0.05)。同时,除了植物全磷与土壤有机碳、土壤全氮呈负相关外,植物有机碳、植物全氮和植物全磷与土壤对应指标相关性不显著。从生态系统乔、灌、草各植被层次对植被和土壤养分分布的影响角度可以看出:辽东栎天然林乔木层对养分分布的影响弱于人工林,只对土壤有机碳、土壤全磷和植物全磷有一定影响。
-
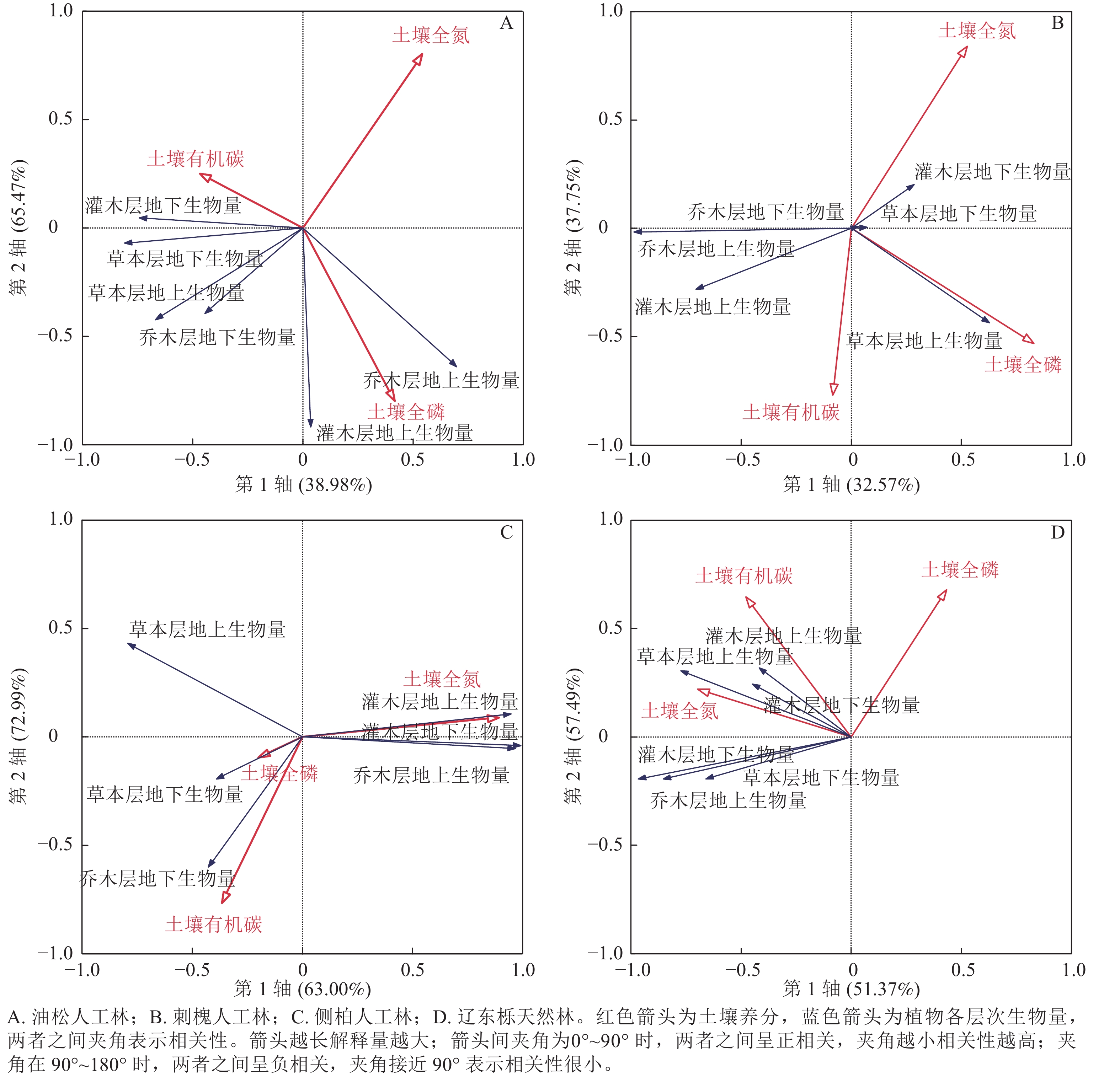
RDA结果(图5)显示:油松人工林乔木层的地上生物量与土壤全磷呈正相关,与土壤有机碳呈负相关,与土壤全氮相关性较小;油松人工林地下生物量与土壤全氮呈负相关,与土壤有机碳、土壤全磷相关性较小。灌木层地上和地下生物量与土壤全氮呈负相关,灌木层的地上生物量与土壤有机碳呈正相关,与土壤全磷呈负相关,灌木层的地下生物量则相反。草本层的地上和地下生物量与土壤全氮的关系与灌木层相同,草本层地下生物量与土壤有机碳呈正相关,与土壤全磷相关性较小,草本地上生物量与土壤有机碳、土壤全磷相关性较小。
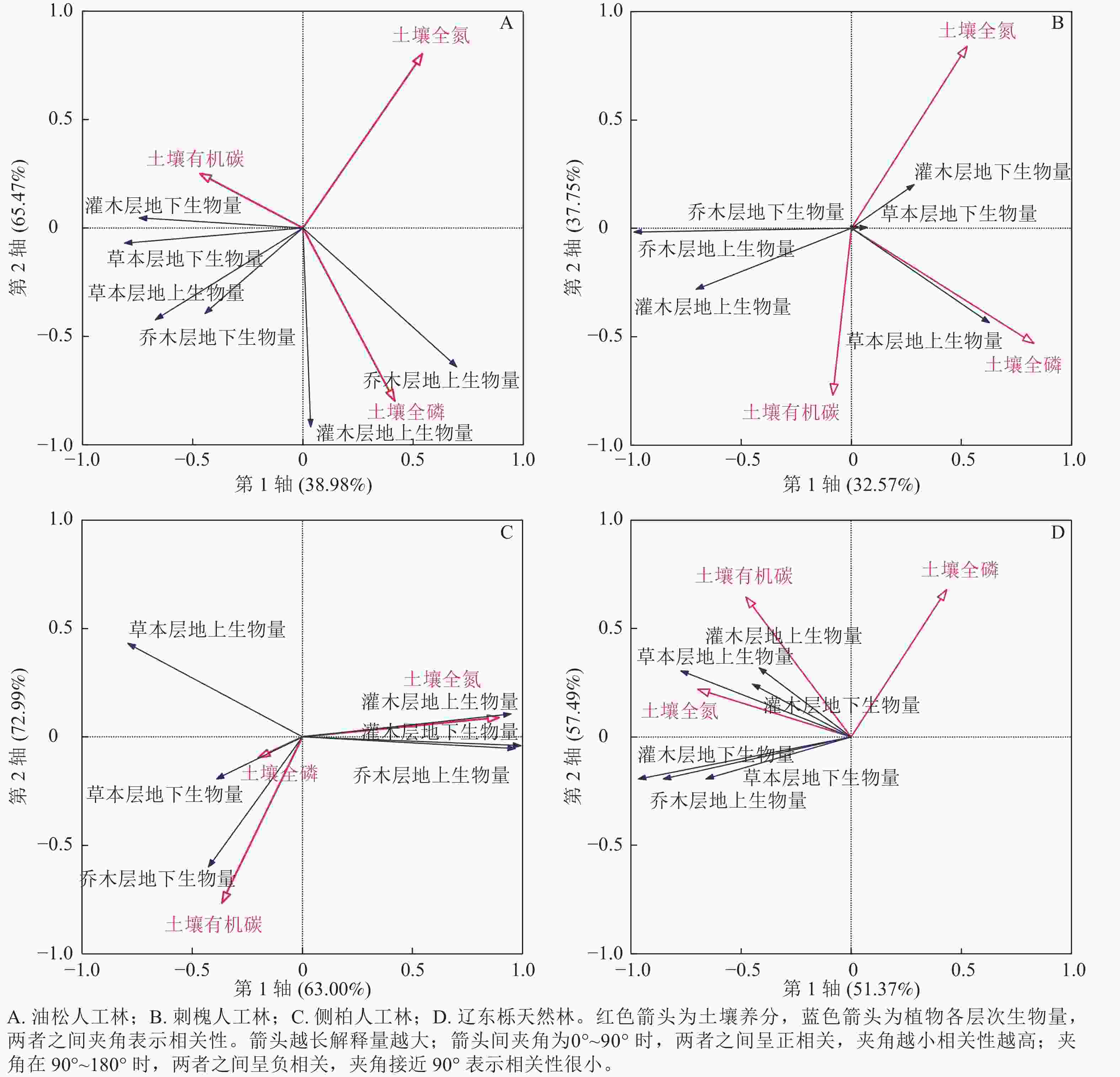
图 5 不同植被恢复类型下生物量与土壤养分的相互关系
Figure 5. Relationship between biomass and soil nutrients under different vegetation restoration types
刺槐人工林乔木层的地上生物量与土壤全磷呈负相关,与土壤有机碳、土壤全氮相关性较小,地下生物量与土壤养分相关性较小。灌木层的地上生物量与土壤有机碳呈正相关,与土壤全氮、土壤全磷呈负相关,地下生物量则相反。草本层的地上生物量与土壤有机碳、土壤全磷呈正相关,与土壤全氮呈负相关,但相关性较小,地下生物量与土壤养分相关性较小。
侧柏人工林乔木层地上生物量与土壤全氮呈正相关,与土壤全磷呈负相关,与土壤有机碳相关性较小,地下生物量与土壤有机碳、土壤全磷呈正相关,与土壤全氮呈负相关。灌木层地上和地下生物量与土壤全氮呈正相关,与土壤有机碳、全磷呈负相关。草本层地上和地下生物量与土壤全氮呈负相关,地上生物量与土壤有机碳、土壤全磷的相关性较小,地下生物量则与土壤有机碳、土壤全磷呈正相关。
辽东栎天然林各层地上和地下生物量均与土壤全氮、土壤有机碳呈正相关,其中灌木层的生物量与土壤全氮、土壤有机碳的相关性最强。乔木层的地上和地下生物量、草本层的地下生物量与土壤全磷呈负相关,灌木层的地上和地下生物量、草本层的地上生物量与土壤全磷相关性较小。
-
研究植物有机碳、植物全氮、植物全磷质量分数可以了解植物养分利用能力和营养状况,从而揭示植物生长的养分限制[18−20]。本研究发现:植物有机碳、植物全氮、植物全磷质量分数在4种不同植被恢复类型间存在一定差异,这与ZENG等[18]对黄土高原不同植被群落和李慧等[21]对晋西不同树种的研究结果相吻合。本研究发现:在辽东栎天然林草本层植物有机碳质量分数显著高于其他植被类型。这是由于辽东栎天然林中主要生长的是乔木或灌木,它们相对较长的生长周期导致植物有机碳积累时间更长。然而,相比其他植被恢复类型,辽东栎天然林草本层中植物全氮质量分数较低。这可能是由于油松人工林、刺槐人工林、侧柏人工林等树种在生长过程中需要大量的氮进行生物合成和代谢,导致它们具有较高的氮吸收和积累能力,因此土壤中相对较多的氮素被用于支持树木的生长[22]。
器官的养分储存功能存在差异,同一类型植被不同器官的植物有机碳、植物全氮、植物全磷质量分数不同。在本研究中,树叶的有机碳、全氮、全磷质量分数最高,这与郭颖等[23]对中国陆地生态系统土壤-植物生态化学计量学研究结果相符合。这是由于叶片是进行光合作用的主要器官,在光合作用过程中,植物吸收二氧化碳和其他无机物质并将其转化为有机物质、蛋白质、核酸等多种化合物,这些有机物首先供养叶片。不同植被恢复类型下,不同器官的有机碳、全氮、全磷质量分数分配和化学计量特征存在显著差异,反映了植物在不同器官间的营养分配策略与环境适应性的变化。此外,这也反映了植物对养分元素需求和利用方式的不同。
-
本研究中,辽东栎天然林土壤有机碳和土壤全氮质量分数显著高于其他3种人工林植被。辽东栎天然林地上、地下生物量高于其他3种人工林植被,原因可能是辽东栎天然林主要是乔木,植被种类丰富,物种多样性较高,凋落物种类较为丰富,同时,土壤中微生物量较高、活性较强,有利于土壤表层有机质的分解[24],使得土壤的碳氮质量分数增高。武小刚等[25]研究发现:植被群落决定凋落物的产生。辽东栎天然林凋落物更易分解,地上生物量及腐殖质等贡献增加,对碳和氮的积累产生直接影响。在本研究中,4种植被恢复类型下,碳、氮元素都主要聚集在土壤0~10 cm土层中,与陈晓萍等[26]在武夷山的研究结果一致。在本研究中,不同土壤深度的磷质量分数并没有表现出较大差异,这可能是因为植物能利用的磷元素形式较少,因此磷元素难以在土层之间转移[27]。在不同植被土壤的磷元素质量分数具有差异,这可能是土壤母质不同所产生的结果,这与LIU等[28]在祁连山的研究结果相似。
土壤碳氮比通常可以反应植被凋落物和根系残留物对土壤碳和氮的积累[29]。以往研究表明[30]:土壤碳氮比越高,土壤有机质的分解速率越慢。在本研究中,刺槐人工林土壤碳氮比最高,而辽东栎天然林和侧柏人工林土壤碳氮比较低,说明辽东栎天然林和侧柏人工林中的土壤有机质分解速率较快。土壤氮磷比可以诊断土壤养分限制类型。在本研究中,4种植被恢复类型下,土壤的氮磷比为0.3~0.8,显著低于全球森林的平均水平14.6[31],说明该区域土壤存在氮限制。研究结果显示:4种植被恢复类型下,土壤的碳磷比与氮磷比呈正相关,这与白永飞等[32]的研究结果一致。本研究的土壤氮磷比与土壤氮质量分数显著正相关,与土壤磷质量分数相关性较小,进一步验证了该区土壤存在氮限制。
-
不同植被恢复类型下,土壤有机碳和土壤全氮展现出了较高的相关性,表明土壤中的养分与植被密切相关。然而,天然林养分与土壤养分之间的关系不太紧密,这表明土壤中养分丰缺程度对植被的生长和养分分布影响较小。在生态系统中,相比于人工林,辽东栎天然林的乔木层对土壤和植被养分的分布影响明显减弱。此外,土壤养分更受到生态系统各个层面的植被属性影响,而不是只受土壤取样深度的影响。辽东栎天然林中,土壤有机碳、土壤全氮、土壤全磷的关系相对密切,特别是土壤全氮和土壤有机碳呈显著正相关。这可能是因为土壤有机碳与土壤全氮均来源于植被凋落物和根系残留物分解合成的有机质[29]。对植被生物量与土壤养分因子的冗余分析结果表明:辽东栎天然林的各层生物量与土壤有机碳、土壤全氮呈正相关,这与赵护兵等[33]对植被土壤养分特征的研究结果较为一致。这可能是因为辽东栎天然林的林分密度相对较低,而其余3种人工林种植密度较高,树木之间的根系相互交错,乔木层之间的竞争比较激烈,使得乔木层对土壤和植被养分分布的影响更加明显。在辽东栎天然林中,自然生长情况下林分密度相对较低,树木之间的竞争关系较弱,因此乔木层对土壤和植被养分分布的影响相对减弱。其次,辽东栎天然林的植被层次多样性较高,且植被层次之间的竞争关系相对平衡[34]。因此,辽东栎天然林中植被层次之间的养分分布相对均衡,乔木层对土壤和植被养分的分布影响相对减弱。最后,辽东栎天然林的土壤养分较为丰富。在人工林中,由于种植密度较高,养分利用率较高,土壤养分相对较低,人工林乔木层对土壤和植物养分分布的影响更加明显[35]。而在辽东栎天然林中,由于生态系统的自然恢复和土地退化的逆转,土壤中的养分含量较高,乔木层对土壤和植物养分分布的影响相对减弱[36]。因此,辽东栎天然林的植被层次之间的竞争关系相对平衡,土壤养分相对丰富,乔木层对土壤和植物养分分布的影响相对减弱,这也是辽东栎天然林生态系统相对于人工林生态系统更为平衡和稳定的重要原因。
在本研究中,辽东栎天然林生态系统的植被养分与土壤养分关系不紧密,这表明在辽东栎天然林中,植被的生长和养分分布不仅受土壤养分水平的影响,还受到其他环境因素的影响,例如水分、光照、温度等[37],这与高宁大[38]的研究结果相同。相比之下,人工林通常采取高密度植被、定期抚育管理等措施,导致植被群落对土壤养分的利用更加集中和高效[39]。这种策略可以在较短时间内加快人工林的生长速度,增加产量,但也容易导致土壤肥力的快速耗竭和生态系统的不稳定。另一方面,相比人工林,辽东栎天然林的植被群落采用的养分利用策略更加合理[40]。辽东栎天然林中的植被群落通常具有多样性和分层结构,可以充分利用不同高度和不同层次的光、水、营养等资源,使得植被对土壤中养分的利用更加高效和均衡[41],植被和土壤之间形成了复杂的养分循环过程,生态系统依靠植被和土壤之间的养分循环过程,充分利用各养分元素以进行自然恢复。因此,在生态系统恢复和管理中,应该注重保护和恢复天然林生态系统,尽可能减少人为干预和破坏,保持生态系统的稳定性和可持续性。
-
本研究探究了晋西黄土区4种植被恢复类型(油松人工林、刺槐人工林、侧柏人工林、辽东栎天然林)不同层次植物碳、氮、磷质量分数,探究不同植被恢复类型下的土壤碳、氮、磷质量分数及化学计量特征。结果表明:①不同植被恢复类型下乔灌草的生物量和植被属性具有显著差异,其中乔木层中油松人工林生物量最高,地上和地下生物量分别为(125.79±3.63)、(24.28±1.86) t·hm−2,灌木层和草本层中辽东栎天然林的生物量最高,灌木层辽东栎天然林地上和地下生物量分别为(8.62±0.78)、(5.91±0.18) t·hm−2,草本层样地辽东栎天然林地上和地下生物量分别为(3.65±0.36)、(3.54±0.25) t·hm−2。在乔木层中,辽东栎天然林的树干和树枝的磷质量分数显著高于其他植被,分别为(1.51±0.09)、(1.74±0.07) g·kg−1;在灌木层中,辽东栎天然林地上部分和地下部分的磷质量分数均显著高于其他3种人工林植被,分别为(2.99±0.32)、(2.52±0.11) g·kg−1;在草本层中,辽东栎天然林碳质量分数显著高于其他植被,为(552.90±19.33) g·kg−1。②土壤全氮、有机碳具有明显的表聚性。辽东栎天然林的土壤养分高于其他林地,土壤全磷、土壤全氧、土壤有机碳质量分数分别平均为(0.92±0.02)、(0.73±0.05)、(13.94±0.28) g·kg−1。对于化学计量特征,辽东栎林的碳磷比高于其他植被,为15.22±0.28,表明辽东栎天然林生长速率较快,但养分利用效率更高。③Mantel Test分析和冗余分析结果显示:不同植被恢复类型下,土壤有机碳和土壤全氮存在较高的相关性;辽东栎天然林的植物养分与土壤养分相关性低于其他3种人工林。同时,土壤植物养分分布的影响人工林乔木层显著高于东栎天然林。土壤养分相比植物养分更受生态系统各层次植被属性的影响。
本研究探索了晋西黄土高原区不同植被恢复类型对生态化学计量的影响及差异,发现辽东栎天然林灌木层和草本层生物量高于其他3种人工林植被,因此种植辽东栎可以在相对较短的时间内形成初步的生态系统,实现水土保持、防风固沙、生态景观等多种功能。
-
感谢山西吉县森林生态系统国家野外科学观测研究站的实验平台及站内水文组、土壤组、气象组、生物组的支持。
Relationship between stoichiometry and ecological factors at various levels of typical vegetation restoration ecosystem in the loess area of western Shanxi Province
-
摘要:
目的 植被恢复是改善黄土高原水土流失和生态环境的重要手段。评估植被恢复对生态系统养分循环和生态服务功能的影响需进一步明晰林地植被与土壤的化学元素计量和分布特征,这对合理开展植被恢复类型筛选具有重要意义。 方法 选取晋西黄土区4种典型植被恢复类型(油松Pinus tabuliformis人工林、刺槐Robinia pseudoacacia人工林、侧柏Platycladus orientalis人工林、辽东栎Quercus mongolica天然林),分别测定乔木层的树枝、树叶、树干、树皮、树根和灌木层、草本层的地上、地下部分以及0~100 cm土壤各层的碳、氮、磷质量分数,结合单因素方差分析法、最小显著差法、冗余分析法等分析植被恢复类型对生态系统养分循环和生态化学计量的影响。 结果 ①相比其他植被恢复类型,辽东栎天然林样地的灌木层和草本层生物量最高,其地上部分生物量分别为(8.62±0.78)、(5.91±0.18) t·hm−2,地下部分生物量分别为(3.65±0.36)、(3.54±0.25) t·hm−2。②辽东栎天然林草本层碳质量分数最高,为(552.90±19.33) g·kg−1,氮质量分数最低,为(10.36±0.30) g·kg−1。③植物与土壤的碳、氮、磷质量分数具有显著相关关系(P<0.05);辽东栎天然林的土壤碳磷比最高,为15.22±0.28,而刺槐人工林土壤碳磷比最低,为8.28±0.26。④Mantel Test和冗余分析结果表明:人工林中,土壤和植被中的养分相关关系显著(P<0.05),然而在辽东栎天然林中两者相关性相对较小,而且辽东栎天然林的乔木层对土壤和植被化学计量特征的影响相对较弱。 结论 近自然造林可以提高植被多样性,并提供更好的生态系统服务。图5表2参41 Abstract:Objective Vegetation restoration is an important method of improving soil erosion and ecological environment in the Loess Plateau. This study aims to assess the impact of vegetation restoration on nutrient cycling and ecological service functions in ecosystem, and further clarify the stoichiometry and distribution characteristics of chemical elements in forest vegetation and soil, which is of great importance for selecting rational models of vegetation restoration. Method 4 typical vegetation restoration types were selected in the loess area of western Shanxi Province, namely Pinus tabuliformis plantation, Robinia pseudoacacia plantation, Platycladus orientalis plantation, and Quercus mongolica natural forest. The contents of carbon (C), nitrogen (N), phosphorus (P) in branches, leaves, trunks, barks, roots of the tree layer, as well as those in the aboveground and underground parts of the shrub layer, herbaceous layer, and 0 − 100 cm soil layers were determined respectively. The effects of vegetation restoration types on nutrient cycling and eco-stoichiometry were analyzed by single factor analysis of variance (ANOVA), minimum significant difference (MSD) and redundancy analysis (RDA). Result (1) Compared with other vegetation restoration types, the shrub layer and herbaceous layer in Q. mongolica forest had the highest biomass, with aboveground biomass of (8.62±0.78) and (5.91±0.18) t·hm−2 and underground biomass of (3.65±0.36) and (3.54±0.25) t·hm−2, respectively. (2) Q. mongolica natural forest had the highest C content, which was (552.90±19.33) g·kg−1, and the lowest N content, which was (10.36±0.30) g·kg−1. (3) There was a significant correlation (P<0.05) between C, N and P contents in plants and soil. Soil C and P ratio in Q. mongolica was the highest (15.22±0.28), while that in R. pseudoacacia was the lowest (8.28±0.26). (4) Mantel Test and RDA showed that there was a significant correlation between nutrients in soil and vegetation in plantation forests (P<0.05), but in Q. mongolica natural forest, the correlation was relatively small and the influence of tree layer on soil and vegetation stoichiometric characteristics was relatively weak. Conclusion Near natural afforestation can promote vegetation diversity and provide better ecosystem services. [Ch, 5 fig. 2 tab. 41 ref.] -
Key words:
- Loess Plateau /
- plant organs /
- vegetation restoration type /
- ecological stoichiometry
-
图 1 不同植被恢复类型不同层次下器官养分性质比较
Figure 1 Comparison of nutrient properties of organs under different layers of different vegetation restoration types
图 2 不同植被恢复类型及不同土层深度土壤有机碳、全氮、全磷质量分数
Figure 2 SOC, TN and TP mass fractions of different vegetation restoration types and different soil depth
图 3 不同植被恢复类型下土壤化学计量比
Figure 3 Comparison of soil stoichiometric characteristics under different vegetation restoration types
图 4 不同植被恢复类型下植被和土壤养分与各层次植被属性相互关系
Figure 4 Relationship between vegetation and soil nutrients and vegetation attributes of each layer of ecosystem under different vegetation restoration types
图 5 不同植被恢复类型下生物量与土壤养分的相互关系
Figure 5 Relationship between biomass and soil nutrients under different vegetation restoration types
表 1 不同植被恢复类型下试验小区基本情况特征
Table 1. Basic characteristics of experimental plot under different vegetation restoration types
植被恢复类型 坐标 海拔/
m坡度/
(°)坡向 林龄/
a林分密度/
(株·hm−2)林下优势植被 油松人工林 36°16′ 24.36″N,110°45′ 29.65″E 1148 26 南 30 1650 沙棘、山桃、低矮薹草等 侧柏人工林 36°16′ 39.00″N,110°44′ 57.10″E 1210 25 南 30 1800 黄荆、山桃、白颖薹草、细裂叶莲蒿等 刺槐人工林 36°16′ 23.11″N,110°45′ 41.14″E 1135 24 南 30 1850 沙棘、山桃、泥胡菜、白颖薹草等 辽东栎天然林 36°44′ 39.34″N,110°44′ 38.80″E 1081 23 南 32 1050 沙棘、山桃、黄荆、低矮薹草、细裂叶莲蒿等 说明:低矮薹草Carex humilis;黄荆Vitex negundo;白颖薹草Carex duriuscula subsp. rigescens;细裂叶莲蒿Artemisia stechmanniana;泥胡菜Hemisteptia lyrata。  下载: 导出CSV
下载: 导出CSV
表 2 不同植被恢复类型下各层次植被属性比较
Table 2. Comparison of vegetation attributes at different levels under different vegetation restoration types
林层 植被恢复类型 地上生物量/(t·hm−2) 地下生物量/(t·hm−2) 各层次高度/m 胸径或基径/cm 冠幅/dm 乔木层 油松人工林 125.79±3.63 a 24.28±1.86 a 6.91±0.21 b 18.19±1.16 a 28.38±1.59 a 刺槐人工林 71.25±2.25 c 25.09±2.02 a 8.60±0.42 a 11.19±0.64 b 17.30±3.09 b 侧柏人工林 78.26±3.91 b 25.86±1.68 a 6.37±0.43 a 8.48±0.88 b 10.83±1.80 b 辽东栎天然林 71.87±1.55 c 18.74±0.70 b 8.91±1.03 b 11.16±2.64 c 14.67±2.06 c 灌木层 油松人工林 4.74±0.16 d 2.91±0.11 b 1.25±0.44 ab 1.38±0.94 a 5.44±3.63 a 刺槐人工林 6.60±0.16 c 1.09±0.06 d 1.09±0.24 ab 1.22±0.23 a 4.51±2.23 a 侧柏人工林 7.52±0.72 b 1.92±0.10 c 0.91±0.21 b 0.73±0.19 a 4.57±3.09 a 辽东栎天然林 8.62±0.78 a 5.91±0.18 a 1.59±0.29 a 1.17±0.16 a 6.02±2.48 a 草本层 油松人工林 1.55±0.05 d 0.61±0.03 d 0.13±0.03 b 1.61±1.73 ab 1.88±1.46 b 刺槐人工林 3.06±0.24 b 1.43±0.06 b 0.16±0.03 b 3.24±1.07 a 3.42±0.93b 侧柏人工林 2.43±0.23 c 1.03±0.08 c 0.16±0.03 b 0.65±0.62 b 2.85±2.22 b 辽东栎天然林 3.65±0.36 a 3.54±0.25 a 0.44±0.14 a 1.65±1.05 ab 9.07±4.24 a 说明:不同植被恢复类型下各层次植被属性均为平均值±标准差;不同小写字母表示相同林层不同植被恢复类型之间差异显著(P<0.05)。
下载: 导出CSV
-
[1] 李威. 植物生态化学计量学的全球格局综述[J]. 南昌工程学院学报, 2016, 35(6): 6 − 10. LI Wei. Global pattern of plant ecological stoichiometrics characteristics [J]. Journal of Nanchang Institute of Technology, 2016, 35(6): 6 − 10. [2] 谭清月. 黄土高原植被恢复对主要生态功能及文化服务的影响[D]. 杨凌: 西北农林科技大学, 2022. TAN Qingyue. Effects of Vegetation Restoration on Main Ecological Functions and Cultural Services on China’s Loess Plateau [D]. Yangling: Northwest A&F University, 2022. [3] 薛凯. 2000年以来黄土高原植被恢复状况与驱动力分析[D]. 聊城: 聊城大学, 2022. XUE Kai. Vegetation Restoration Status and Its Driving Forces in the Loess Plateau Since 2000 [D]. Liaocheng: Liaocheng University, 2022. [4] 李家豪. 黄土高原不同植被恢复措施对草本群落和土壤碳氮的影响[D]. 西安: 西北大学, 2022. LI Jiahao. Effects of Different Vegetation Restoration Types on Herbaceous Communities and Soil Carbon and Nitrogen in the Loess Plateau [D]. Xi’an: Xibei University, 2022. [5] 杨阳, 刘良旭, 童永平, 等. 黄土高原植被恢复过程中土壤碳储量及影响因素研究进展[J]. 地球环境学报, 2023, 14(6): 649 − 662. YANG Yang, LIU Liangxu, TONG Yongping, et al. Advances and driving factors in soil organic carbon storage during vegetation restoration in the Loess Plateau, China [J]. Journal of Earth Environment, 2023, 14(6): 649 − 662. [6] 王世军, 杨磊, 段兴武, 等. 黄土高原小流域植被恢复的土壤水分和养分权衡效应研究[J]. 土壤通报, 2022, 53(2): 356 − 365. WANG Shijun, YANG Lei, DUAN Xingwu, et al. Trade-off effects of soil moisture and soil nutrients under vegetation restoration in a small watershed on the Loess Plateau, China [J]. Chinese Journal of Soil Science, 2022, 53(2): 356 − 365. [7] 王俊, 郭金龙, 张永旺, 等. 黄土高原自然植被恢复过程中土壤温度和水分的相关性[J]. 水土保持学报, 2022, 36(2): 130 − 137. WANG Jun, GUO Jinlong, ZHANG Yongwang, et al. The correlation between soil temperature and water content during the natural vegetation restoration on the Loess Plateau [J]. Journal of Soil and Water Conservation, 2022, 36(2): 130 − 137. [8] 邓健, 张丹, 张伟, 等. 黄土丘陵区刺槐叶片-土壤-微生物碳氮磷化学计量学及其稳态性特征[J]. 生态学报, 2019, 39(15): 5527 − 5535. DENG Jian, ZHANG Dan, ZHANG Wei, et al. Carbon, nitrogen, and phosphorus stoichiometry and homeostasis characteristics of leaves, soil, and microbial biomass of Robinia pseudoacacia forests in the Loess Hilly Region of China [J]. Acta Ecologica Sinica, 2019, 39(15): 5527 − 5535. [9] 闫丽娟, 王海燕, 李广, 等. 黄土丘陵区4种典型植被对土壤养分及酶活性的影响[J]. 水土保持学报, 2019, 33(5): 190 − 196, 204. YAN Lijuan, WANG Haiyan, LI Guang, et al. Effects of four typical vegetations on soil nutrient and enzymes activities in Loess Hilly Region [J]. Journal of Soil and Water Conservation, 2019, 33(5): 190 − 196, 204. [10] JIANG Yefeng, RAO Lei, SUN Kai, et al. Spatio-temporal distribution of soil nitrogen in Poyang lake ecological economic zone (South-China) [J]. Science of the Total Environment, 2018, 626: 235 − 243. [11] 杨阔, 黄建辉, 董丹, 等. 青藏高原草地植物群落冠层叶片氮磷化学计量学分析[J]. 植物生态学报, 2010, 34(1): 17 − 22. YANG Kuo, HUANG Jianhui, DONG Dan, et al. Canopy leaf N and P stoichiometry in grassland communities of Qinghai-Tibetan Plateau, China [J]. Chinese Journal of Plant Ecology, 2010, 34(1): 17 − 22. [12] SONG Min, PENG Wanxia, DU Hu, et al. Responses of soil and microbial C∶N∶P stoichiometry to vegetation succession in a karst region of southwest China [J/OL]. Forests, 2019, 10(9): 755[2023-07-27]. doi:10.3390/f10090755. [13] 卢建男, 刘凯军, 王瑞雄, 等. 中国荒漠植物-土壤系统生态化学计量学研究进展[J]. 中国沙漠, 2020, 42(2): 173 − 182. LU Jiannan, LIU Kaijun, WANG Ruixiong, et al. Research advances in stoichiometry of desert plant-soil system in China [J]. Journal of Desert Research, 2020, 42(2): 173 − 182. [14] 梁楚欣, 范弢, 陈培云. 滇东石漠化坡地不同恢复模式下云南松林土壤碳氮磷化学计量特征及其影响因子[J]. 浙江农林大学学报, 2023, 40(3): 511 − 519. LIANG Chuxin, FAN Tao, CHEN Peiyun. Stoichiometric characteristics and influencing factors of soil C, N and P in Pinus yunnanensis forests under different restoration modes on rocky desertification slope land in eastern Yunnan [J]. Journal of Zhejiang A&F University, 2023, 40(3): 511 − 519. [15] 刘玉林, 朱广宇, 邓蕾, 等. 黄土高原植被自然恢复和人工造林对土壤碳氮储量的影响[J]. 应用生态学报, 2018, 29(7): 2163 − 2172. LIU Yulin, ZHU Guangyu, DENG Lei, et al. Effects of natural vegetation restoration and afforestation on soil carbon and nitrogen storage in the Loess Plateau, China [J]. Chinese Journal of Applied Ecology, 2018, 29(7): 2163 − 2172. [16] 赵育民, 牛树奎, 王军邦, 等. 植被光能利用率研究进展[J]. 生态学杂志, 2007, 26(9): 1471 − 1477. ZHAO Yuming, NIU Shukui, WANG Junbang, et al. Research progress on vegetation light energy utilization [J]. Chinese Journal of Ecology, 2007, 26(9): 1471 − 1477. [17] 王彬, 魏天兴, 刘钊. 黄土丘陵区华北落叶松人工林生态系统生物量与养分循环特征[J]. 水土保持研究, 2017, 24(6): 45 − 51. WANG Bin, WEI Tianxing, LIU Zhao. Characteristics of nutrient cycling and ecosystem structure of Larix principis-rupprechtii in Hilly Loess Plateau [J]. Research of Soil and Water Conservation, 2017, 24(6): 45 − 51. [18] ZENG Quanchao, LI Xing, DONG Yanghong, et al. Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China [J]. Catena, 2016, 147: 481 − 488. [19] 刘莉, 王明浩, 杨蔚, 等. 不同干扰类型下滇西北高寒草甸土壤化学计量特征[J]. 草业科学, 2022, 39(4): 634 − 644. LIU Li, WANG Minghao, YANG Wei, et al. Soil stoichiometric characteristics of alpine meadow in northwest Yunnan under different disturbance types [J]. Pratacultural Science, 2022, 39(4): 634 − 644. [20] KHAN A, KONG Xiangjun, ULLAH N, et al. Planting density induced changes in cotton biomass yield, fiber quality, and phosphorus distribution under Beta Growth Model [J/OL]. Agronomy, 2019, 9(9): 500[2023-07-27]. doi:10.3390/agronomy9090500. [21] 李慧, 王百田, 刘涛. 晋西黄土区不同森林树种及其林地土壤养分含量的变化[J]. 林业科学研究, 2016, 29(4): 587 − 595. LI Hui, WANG Baitian, LIU Tao. The nutrient content variations of different forest species and the forest soil in Loess Region of Western Shanxi [J]. Forest Research, 2016, 29(4): 587 − 595. [22] 董莉丽, 郑粉莉. 黄土丘陵区不同土地利用类型下土壤酶活性和养分特征[J]. 生态环境, 2008, 17(5): 2050 − 2058. DONG Lili, ZHENG Fenli. Characteristics of soil enzyme activities and nutrients under various landuse in the Loessial hilly-gully region [J]. Ecology and Environment, 2008, 17(5): 2050 − 2058. [23] 郭颖, 李军, 张亚亚, 等. 我国陆地生态系统土壤·植物生态化学计量学研究进展[J]. 安徽农业科学, 2016, 44(16): 1 − 6. GUO Ying, LI Jun, ZHANG Yaya, et al. Research advances in soil and plant stoichiometry in terrestrial ecosystem in China [J]. Journal of Anhui Agricultural Sciences, 2016, 44(16): 1 − 6. [24] SINGH C, MISHRA D, SINGH K, et al. Geographical settings and tree diversity influenced soil carbon storage in diferent forest types in Rajasthan, India [J/OL]. Catena, 2022, 209: 105856[2023-07-27]. doi: 10.1016/j.catena.2021.105856. [25] 武小钢, 郭晋平, 杨秀云, 等. 芦芽山典型植被土壤有机碳剖面分布特征及碳储量[J]. 生态学报, 2011, 31(11): 3009 − 3019. WU Xiaogang, GUO Jinping, YANG Xiuyun, et al. Soil organic carbon storage and profile inventory in the different vegetation types of Luya Mountain [J]. Acta Ecologica Sinica, 2011, 31(11): 3009 − 3019. [26] 陈晓萍, 郭炳桥, 钟全林, 等. 武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应[J]. 生态学报, 2018, 38(1): 273 − 281. CHEN Xiaoping, GUO Bingqiao, ZHONG Quanlin, et al. Response of fine root carbon, nitrogen, and phosphorus stoichiometry to soil nutrients in Pinus taiwanensis along an elevation gradient in the Wuyi Mountains [J]. Acta Ecologica Sinica, 2018, 38(1): 273 − 281. [27] 朱都庆. 我国干旱与半干旱地区土壤氮磷元素空间分布格局及其驱动因素[D]. 兰州: 兰州大学, 2021. ZHU Duqing. Spatial Distribution Patterns of Soil Nitrogen and Phosphorus and Their Driving Factors in Arid and Semiarid Zones of China [D]. Lanzhou: Lanzhou University, 2021. [28] LIU Jianguo, GOU Xiaohua, ZHANG Fen, et al. Spatial patterns in the C∶N∶P stoichiometry in Qinghai spruce and the soil across the Qilian Mountains, China [J/OL]. Catena, 2021, 196: 104814[2023-07-27]. doi: 10.1016/j.catena.2020.104814. [29] RADOČAJ D, JURIŠIĆ M, ANTONIĆ O. Determination of soil C∶N suitability zones for organic farming using an unsupervised classification in eastern Croatia [J/OL]. Ecological Indicators, 2021, 123: 107382[2023-07-27]. doi: 10.1016/j.ecolind.2021.107382. [30] CLEVELAND C C, LIPTZIN D. C∶N∶P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? [J]. Biogeochemistry, 2007, 85: 235 − 252. [31] MELILLO J M, FIELD C B, MOLDAN B. Interactions of the Major Biogeochemical Cycles: Global Change and Human Impacts [M]. Washington D C: Island Press, 2003: 117 − 134. [32] 白永飞, 李凌浩, 王其兵, 等. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J]. 植物生态学报, 2000, 24(6): 667 − 673. BAI Yongfei, LI Linghao, WANG Qibing, et al. Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River basin, Inner Mongolia [J]. Acta Phytoecologica Sinica, 2000, 24(6): 667 − 673. [33] 赵护兵, 刘国彬, 侯喜禄. 黄土丘陵区流域主要植被类型养分循环特征[J]. 草业学报, 2006, 15(3): 63 − 69. ZHAO Hubing, LIU Guobin, HOU Xilu. Characteristics of nutrient cycling of different vegetation types in the Zhifanggou Watershed on the Loess Hilly Region [J]. Acta Prataculturae Sinica, 2006, 15(3): 63 − 69. [34] 赵一娉, 曹扬, 陈云明, 等. 黄土丘陵沟壑区森林生态系统生态化学计量特征[J]. 生态学报, 2017, 37(16): 5451 − 5460. ZHAO Yiping, CAO Yang, CHEN Yunming, et al. Ecological stoichiometry in a forest ecosystem in the hilly-gully area of the Loess Plateau [J]. Acta Ecologica Sinica, 2017, 37(16): 5451 − 5460. [35] MA Rentian, HU Feinan, LIU Jingfang, et al. Shifts in soil nutrient concentrations and C∶N∶P stoichiometry during long-term natural vegetation restoration [J/OL]. PeerJ, 2020, 8: e8382[2023-07-27]. doi: 10.7717/peerj.8382. [36] 李张敏. 九连山人工和自然恢复森林物种多样性特征及其环境解释[D]. 南昌: 江西农业大学, 2019. LI Zhangmin. Characteristics of Species Diversity in Artificial and Natural Restored Forests in Jiulian Mountains and Their Environmental Explanations [D]. Nanchang: Jiangxi Agricultural University, 2019. [37] 王子婷, 柴春山, 张洋东, 等. 半干旱黄土区柠条生长与环境因子的关系研究进展[J]. 中国水土保持, 2021(1): 49 − 52. WANG Ziting, CAI Chunshan, ZHANG Yangdong, et al. Research progress on the relationship between the growth of caragana and environmental factors in semi-arid loess region [J]. Soil and Water Conservation in China, 2021(1): 49 − 52. [38] 高宁大. 土壤水分和养分对树木细根生物量、生产力的影响分析[J]. 现代园艺, 2023, 46(6): 39 − 41. GAO Ningda. Effects of soil moisture and nutrients on fine root biomass and productivity of trees [J]. Contemporary Horticulture, 2023, 46(6): 39 − 41. [39] 张勇强, 李智超, 厚凌宇, 等. 林分密度对杉木人工林下物种多样性和土壤养分的影响[J]. 土壤学报, 2020, 57(1): 239 − 250. ZHANG Yongqiang, LI Zhichao, HOU Lingyu, et al. Effects of stand density on understory species diversity and soil nutrients in Chinese fir plantation [J]. Acta Pedologica Sinica, 2020, 57(1): 239 − 250. [40] 张平究, 潘根兴. 不同植被群落下喀斯特土壤养分及生物化学性质特征[J]. 水土保持学报, 2012, 26(1): 77 − 80, 84. ZHANG Pingjiu, PAN Genxing. Characteristics of soil nutrients and biochemical properties under different vegetation communities in Karst area [J]. Journal of Soil and Water Conservation, 2012, 26(1): 77 − 80, 84. [41] 熊瑛楠, 冯天骄, 王平, 等. 晋西黄土区长期人工林恢复对土壤水分和养分性质的影响[J]. 水土保持学报, 2022, 36(2): 228 − 237, 246. XIONG Yingnan, FENG Tianjiao, WANG Ping, et al. Effeets of long-term artificial forest restoration on soil moisture and nutrient characteristics in the Loess Area of Western Shanxi Province, China [J]. Journal of Soil and Water Conservation, 2022, 36(2): 228 − 237, 246. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230448
 点击查看大图
点击查看大图
计量
- 文章访问数: 1360
- HTML全文浏览量: 367
- PDF下载量: 24
- 被引次数: 0
