




-
海拔高度与昆虫的物种多样性关系密切,不同海拔高度的同类昆虫物种多样性各不相同[1]。鳞翅目Lepidopte蛾类昆虫经过长时间的协同进化,已成为维持生态平衡的重要指示类群之一,是具有重要科研价值的一类昆虫[2]。大蛾类作为翅展大于25 mm的蛾类,是重要的访花和传粉昆虫[3],同时大蛾类的部分昆虫也是主要的农林害虫,如枯叶蛾科Lasiocampidae的松毛虫属Dendrolimus昆虫每年对松林面积的破坏性大,已成为全世界森林中危害最严重的一类害虫[4−6]。鉴于此,本研究在浙江天目山国家级自然保护区内对不同海拔高度的大蛾类种群多样性及时间动态进行了研究,以期为天目山保护区昆虫资源数据提供参考。
-
浙江天目山国家级自然保护区位于杭州市临安区(30°18′~30°24′N,119°23′~119°29′E),最高峰仙人顶海拔为1 506.0 m。该区属亚热带季风气候,年均气温为8.1~14.3 ℃,年均降水量为1 100.0~1 900.0 mm。区内植被类型多样且随着海拔梯度的变化而发生显著变化[7]。
-
在研究区内,每隔200 m的海拔梯度设置1个样点,共设置8个样点,分别为海拔0~200、200~400、400~600、600~800、800~1 000、1 000~1 200、1 200~1 400、1 400~1 506 m。白天时段在各个海拔梯度通过网捕法捕捉大蛾类幼虫和成虫,幼虫放在室内饲养再根据成虫的形态特征进行鉴定。夜晚时段(20:00—24:00)在各个海拔梯度中选取距林缘50~100 m、地势相对平坦的空地,放置1个灯诱帐篷[规格大小为1.2 m×1.2 m×1.8 m,光源为2 000 W (12 000 mA)续航75 h的户外充电照明灯]进行灯诱,并收集大蛾类昆虫。从5月开始,每周对各个样点的大蛾类标本收集并统计,共持续4个月,以每个月为单位对该月所采集的大蛾类进行分析、鉴定和统计。
-
采用Simpson指数、Shannon指数、Pielou均匀度和Margalef丰富度指数进行大蛾类种群多样性分析,统计每个样地中出现的物种数和个体数。多样性指数计算参考文献[8−9]。
-
利用对数正态分布模型对不同海拔梯度上大蛾类种群的种-多度进行分析,模型表达式为${{{S}}_{{{(R)}}}} = {S _0}\exp ( - {\alpha ^2}{R^2})$。其中:S(R)是R倍程中的物种数量,S0是对数正态分布的众数倍程物种数,α是与分布有关的参数[10]。
-
种群相似性用Jaccard相似性系数和系统聚类进行表征。①Jaccard相似性系数计算公式为C=c/(a+b+c)。其中:C为种群Jaccard相似性系数,a、b分别为2个生境所具有的物种数,c为2个生境中共有的物种数[11]。②系统聚类计算公式为${{d}}_{{ij}}=\sqrt{\left[{{\displaystyle \sum ({{X}}_{{ik}}-{{X}}_{{jk}})}}^{\text{2}}\right]}$。其中:dij为i种群与j种群之间的欧氏距离,Xik为i种群中第k个指标的值,Xjk为j种群中第k个指标的值[12]。
-
用Excel对大蛾类昆虫进行统计,并进行多样性分析,通过SPSS 21对大蛾类种群的物种数进行系统聚类和差异显著性检验,用Origin 2022对物种进行时间动态和种-多度关系分析。
-
从8个海拔梯度中共采集大蛾类昆虫标本3 367只,隶属25科269种。草螟科Crambidae、尺蛾科Geometridae、夜蛾科Noctuidae和天蛾科Sphingidae的个体数均超过总标本数的10%,其中尺蛾科个体数最多,有615只,占总个体数的18.3%。尺蛾科、天蛾科在每个海拔梯度上占总数比例都较多。箩纹蛾科Brahmaeidae的个体数量最少。
从表1可见:不同海拔的物种数各不相同,从低海拔到高海拔总体呈先增加后减小的趋势。随着海拔的升高,物种数和个体数逐渐增加,在800 m处达到最大值,随后随着海拔的升高,物种数和个体数逐渐减少。
表 1 天目山不同海拔下大蛾类种群的组成
Table 1. Composition of large moth species at different altitudes of Mount Tianmu
科名 0~200 m 200~400 m 400~600 m 600~800 m 800~1 000 m 1 000~1 200 m 1 200~1 400 m 1 400~1 600 m 个体总
数/只S N S N S N S N S N S N S N S N 斑蛾科Zygaenidae 1 2 0 0 1 6 1 12 1 5 0 0 0 0 0 0 25 波纹蛾科Thyatiridae 1 9 1 11 2 10 2 25 2 20 1 6 1 1 0 0 82 蚕蛾科Bombycidae 1 3 1 8 1 3 2 12 1 4 1 5 1 1 0 0 36 草螟科Crambidae 15 29 16 43 25 53 26 69 23 71 15 44 7 21 3 8 338 尺蛾科Geometridae 19 49 20 65 31 89 53 116 57 122 34 93 22 53 11 28 615 刺蛾科Limacodidae 1 2 1 4 3 15 4 12 3 10 1 1 0 0 0 0 44 大蚕蛾科Saturniidae 2 8 4 15 7 30 8 45 7 30 4 14 5 14 0 0 156 带蛾科Thaumetopoeidae 0 0 1 2 1 4 1 9 1 7 0 0 0 0 0 0 22 灯蛾科Arctiidae 1 3 1 12 1 7 2 25 2 18 1 1 0 0 1 2 68 毒蛾科Lymantriidae 2 6 2 13 3 20 4 31 3 15 2 6 1 1 0 0 92 凤蛾科Epicopeiidae 1 2 0 0 1 1 1 1 1 4 0 0 0 0 0 0 8 钩蛾科Drepanidae 3 7 4 8 7 16 9 27 7 22 1 4 2 3 0 0 87 卷蛾科Tortricidae 1 9 1 7 0 0 1 7 1 3 0 0 0 0 0 0 26 枯叶蛾科Lasiocampidae 1 3 3 13 8 23 9 31 6 21 5 16 2 5 0 0 112 瘤蛾科Nolidae 1 3 2 6 3 13 3 22 3 13 2 4 0 0 0 0 61 箩纹蛾科Brahmaeidae 0 0 0 0 0 0 0 1 1 2 0 0 0 0 0 0 3 螟蛾科Pyralidae 2 7 4 15 5 23 6 56 5 47 6 27 2 8 3 8 191 木蠹蛾科Cossidae 2 10 1 2 2 8 3 9 1 8 1 1 0 0 0 0 38 裳蛾科Erebidae 2 7 4 15 7 25 11 40 9 27 6 13 1 3 0 0 130 苔蛾科Lithosiidae 1 2 1 10 1 20 1 43 1 28 1 34 1 6 0 0 143 天蛾科Sphingidae 12 28 13 35 24 56 31 112 30 113 26 80 8 20 5 13 457 网蛾科Thyrididae 1 4 2 11 3 20 3 20 2 14 1 4 1 1 0 0 74 燕蛾科Uraniidae 0 0 1 2 2 10 2 18 1 8 0 0 0 0 0 0 38 夜蛾科Noctuidae 5 16 7 22 11 47 35 79 40 78 23 59 17 36 2 11 348 舟蛾科Notodontidae 2 8 4 13 3 13 11 55 14 61 2 15 1 7 1 1 173 S合计 77 94 152 229 222 131 72 26 269 N合计 217 332 512 877 751 427 180 71 3367 S占比/% 28.62 34.94 56.51 85.13 82.53 49.44 26.77 9.67 N占比/% 6.44 9.86 15.21 26.05 22.30 12.68 5.35 2.10 说明:S. 物种数(种);N. 个体数(只)。各海拔梯度之间有相同物种。 -
从表2可见:不同海拔梯度下,大蛾类种群的Simpson指数和Shannon指数从高到低依次为海拔600~800、400~600、800~1 000、200~400、0~200、1 000~1 200、1 200~1 400、1 400~1 600 m,在海拔 1 000 m以上差异显著(P<0.05)。Pielou均匀度在海拔800 m以上差异显著(P<0.05)。Margalef丰富度在海拔1 000 m以上差异显著(P<0.05)。
表 2 天目山不同海拔梯度下大蛾类种群多样性指数
Table 2. Large moth population diversity index under different altitude gradients of Mount Tianmu
海拔/m Simpson指数 Shannon指数 Pielou均匀度 Margalef丰富度 0~200 0.869±0.040 bc 2.322±0.310 bc 0.345±0.040 b 3.903±0.400 a 200~400 0.901±0.010 abc 2.565±0.120 ab 0.381±0.010 a 3.617±0.160 a 400~600 0.910±0.010 ab 2.634±0.080 a 0.391±0.010 a 3.527±0.200 a 600~800 0.922±0.000 a 2.769±0.200 a 0.411±0.020 a 3.542±0.180 a 800~1 000 0.902±0.010 abc 2.605±0.090 ab 0.387±0.010 a 3.625±0.180 a 1 000~1 200 0.857±0.030 c 2.195±0.160 c 0.326±0.020 b 2.972±0.190 b 1 200~1 400 0.791±0.060 d 1.816±0.310 d 0.269±0.040 c 2.696±0.230 b 1 400~1 600 0.646±0.030 e 1.242±0.070 e 0.188±0.010 d 1.408±0.110 c 说明:数值为平均值±标准差。同列不同字母表示同一指数在不同海拔间差异显著(P<0.05)。 -
8个海拔梯度的种-多度曲线中,海拔600~800、800~1 000、1 000~1 200 m趋向于对数正态分布模型(图1),拟合以上3个海拔梯度,发现海拔在800~1 000 m处的拟合度最高,说明在海拔800~1 000 m处的环境条件最优越,即优势类群和稀有类群都较少,常见种较多。其他海拔梯度符合生态位优先占领假说。

图 1 天目山不同海拔高度大蛾类种-多度曲线
Figure 1. Large moth species-multidegree curves at different altitudes of Mount Tianmu
-
从表3可见:8个海拔梯度下大蛾类种群呈现中等相似,且差异不显著。其中海拔600~1 000 m的大蛾类种群与其他海拔的大蛾类种群相似性最高,而海拔1 400~1 600 m的大蛾类种群与其他海拔大蛾类种群相似性低。表明高海拔地区对大蛾类种群的影响较明显。
表 3 不同海拔天目山大蛾类种群相似性系数
Table 3. Similarity coefficients of large moth population at different altitudes of Mount Tianmu
海拔/m 0~200 m 200~400 m 400~600 m 600~800 m 800~1 000 m 1 000~1 200 m 1 200~1 400 m 1 400~1 600 m 0~200 1.000 0.833 0.875 0.880 0.880 0.864 0.682 0.318 200~400 1.000 0.875 0.880 0.880 0.864 0.682 0.318 400~600 1.000 0.657 0.657 0.826 0.652 0.304 600~800 1.000 0.900 0.760 0.600 0.280 800~1 000 1.000 0.760 0.600 0.280 1 000~1 200 1.000 0.789 0.368 1 200~1 400 1.000 0.467 1 400~1 600 1.000 -
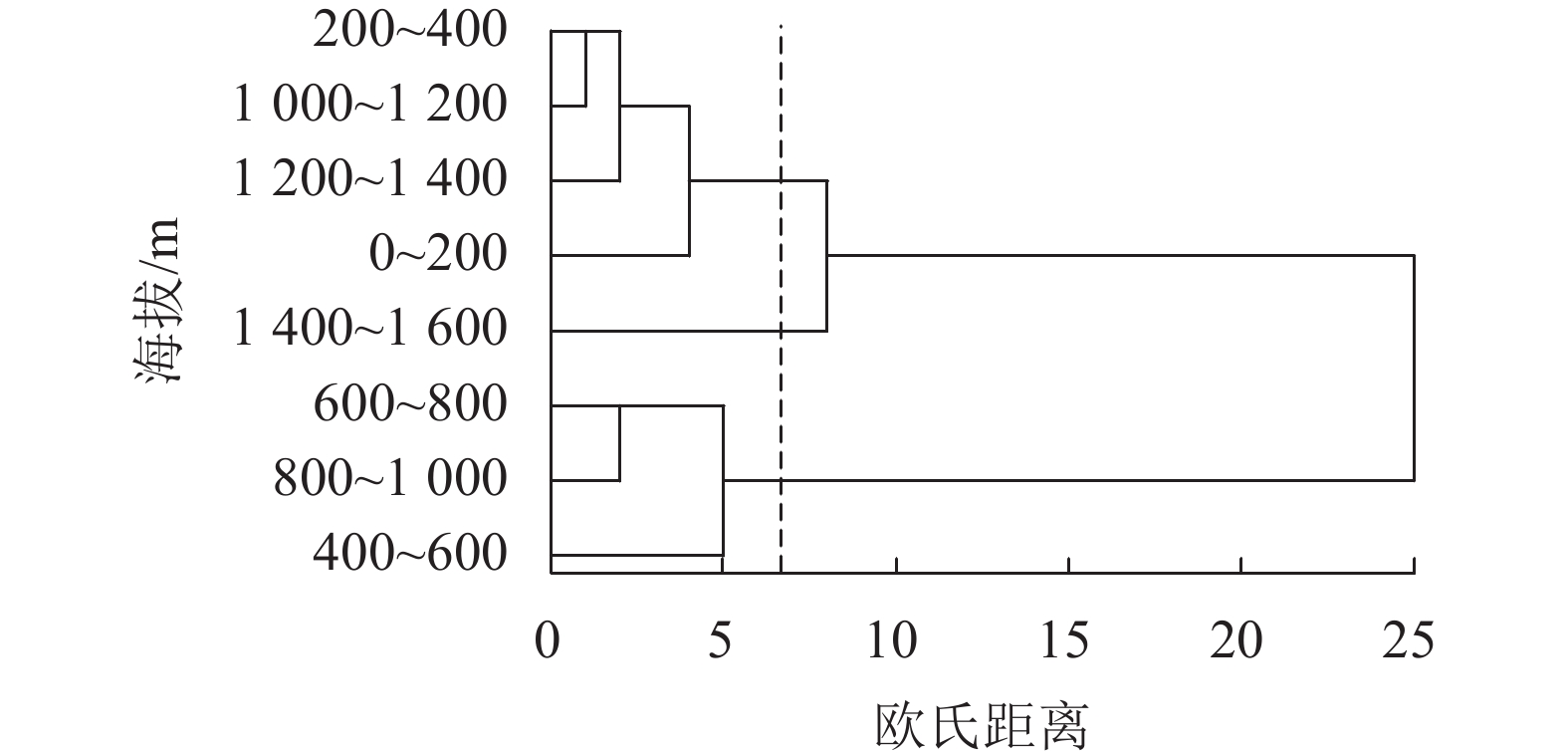
从图2可见:海拔200~400、1 000~1 400 m的大蛾类种群聚为一类;海拔0~200、1 400~1 600 m的种群聚为一类;海拔400~1 000 m的种群聚为一类。可见,海拔400~1 000 m处的环境保护相对较好,种群结构稳定,不同海拔高度的大蛾类种群系统聚类与相似性分析结果类似。

图 2 天目山不同海拔系统聚类分析
Figure 2. Continental distance clustering analysis at different altitudes of Mount Tianmu
-
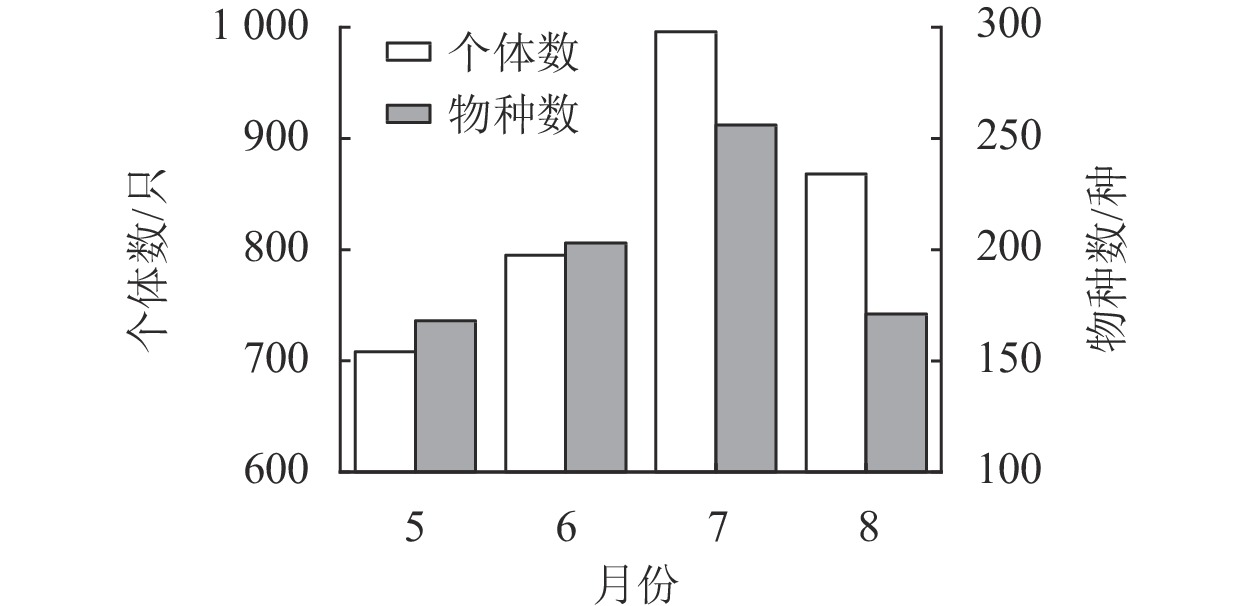
如图3所示:5月共采集大蛾类标本708只,隶属22科168种,6月共采集大蛾类标本795只,隶属24科203种,7月共采集大蛾类标本996只,隶属24科256种,8月共采集大蛾类标本868只,隶属23科171种。通过对比分析发现:7月大蛾类的物种数和个体数均最大,5月最小。其中,7和8月大蛾类种群的物种数和个体数存在差异。

图 3 不同月份大蛾类的物种数和个体数
Figure 3. Number of species and individuals of large moth in different months
-
如表4所示:天目山大蛾类种群的Simpson指数、Shannon指数、Pielou均匀度和Margalef丰富度的变化趋势一致,均在5和6月逐渐递增,7月最大,之后开始下降。说明大蛾类种群的物种数和个体数在均匀度指数上都是先增加后减少,均匀度越高则多样性越稳定,均匀度从大到小依次为7、6、5、8月。
表 4 不同月份大蛾类种群多样性
Table 4. New diversity of large moth species in different months
月份 个体
数/只Simpson
指数Shannon
指数Pielou
均匀度Margalef
丰富度5 708 0.798 2.162 0.329 2.668 6 795 0.819 2.263 0.339 3.231 7 996 0.819 2.438 0.353 3.534 8 868 0.788 2.210 0.327 2.876 -
本研究表明:天目山大蛾类种群多样性随着海拔高度的上升呈先增加后减少的趋势,在海拔600~800 m达到最大值。这与张瀚文等[1]的研究结果不一致,但与孙儒泳[13]的研究结果类似。这可能与天目山不同海拔的森林环境效益有关[14]。海拔0~400 m由于在保护区外,人为干扰严重,导致天目山生物多样性降低。海拔400~1 000 m在保护区内,物种多样性增加[15]。海拔1 000~1 600 m由于人为防火道的设立,再加上高大乔木的植被类型逐渐转换为落叶矮林和毛竹Phyllostachys edulis林,大蛾类种群的生物多样性降低。
除此之外,本研究还发现许多西部以及中部地区分布的大蛾类,例如粉红边尺蛾Leptomiza crenularia、灰沙黄碟尺蛾Thinopteryx delectans和白肩天蛾Rhagastis mongoliana mongoliana等[16]。此外还发现了青球箩纹蛾Brahmaea hearseyi、胖夜蛾Orthogonia sera、修虎蛾Sarbanissa transiens、平背天蛾Cechenena minor、椴六点天蛾Marumba dyras、金双点螟Orybina regalis等常见且种群数较少的大蛾类种类,这些物种对生境质量要求较高,森林环境变化对该大蛾类多样性影响明显[17]。由此可见天目山大蛾类物种丰富,生态环境较好,为绝大多数生物提供了较好的栖息地[18]。
-
中度干扰假说是指中等条件有利于提高大蛾类种群稳定性[19],对天目山8个不同海拔梯度的大蛾类种群多样性分析发现:人为环境的干扰也是影响大蛾类种群稳定的重要因素。在海拔600~1 000 m的梯度上,由于受到轻度人为干扰,大蛾类个体数量、物种数和多样性指数较高,大蛾类种群较稳定。海拔0~600、1 200~1 600 m梯度上分别受到重度人为干扰和无人为干扰,大蛾类个体数、物种数和多样性指数较低,大蛾类种群不稳定,这也印证了中度干扰有利于提高大蛾类种群稳定性。
-
天目山大蛾类种群不同月份的个体数和物种数的变化趋势基本一致。这种现象可能与天目山气温变化和植被群落丰富度有关[20]。7月的物种数和个体数最多,其次是6、5、8月,这可能是7月是常绿阔叶林植被生长最为茂盛的阶段,此阶段更适宜大蛾类种群的发育繁殖。表明大蛾类种群对环境变化敏感,物种数差距大的同时多样性指数和均匀度指数也存在一定的差异,因此将大蛾类种群作为指示生物,对天目山海拔梯度的比较分析具有一定的代表性。
-
浙江天目山国家级自然保护区管理局杨淑贞教授级高级工程师对本研究给予大力支持,在此感谢!
Population diversity and temporal dynamics of large moths at different altitudes in Mount Tianmu
-
摘要:
目的 阐明浙江天目山大蛾类种群在不同海拔上的关系以及时间动态变化,为天目山昆虫调查提供理论依据。 方法 采用灯诱法,于2022年5—8月在浙江天目山国家级自然保护区内,对不同海拔高度的大蛾类昆虫进行采集,并对大蛾类种群及时间动态进行了研究。 结果 共采集大蛾类昆虫3 367只,隶属25科269种。优势类群为尺蛾科Geometridae、草螟科Crambidae、天蛾科Sphingidae和夜蛾科Noctuidae。在海拔600~1 200 m内,大蛾类种群的Simpson指数、Shannon指数、Margalef丰富度指数和Pielou均匀度指数均较高,总体呈正相关。在海拔600~1 200 m梯度上,大蛾类的种-多度曲线趋向对数正态分布,其余海拔梯度符合生态位优先占领假说。7月的大蛾类物种数和个体数最多,稳定性较强。 结论 天目山的大蛾类种群对时间敏感,5—8月大蛾类物种和个体数先增加后减少;大蛾类种群多样性随着海拔的升高先增加后减少,中低海拔梯度间的相似性系数呈中等相似和极相似水平。图3表4参20 Abstract:Objective With the clarification of the relationships and temporal dynamics of large moth species in Mount Tianmu in Zhejiang Province, this paper aims to provide a theoretical basis for the insect investigation in Mount Tianmu. Method Large moth specimens were collected from Zhejiang Mount Tianmu National Nature Reserve at different altitudes during the four-month period from May to August in 2022 before a systematic analysis was conducted of them and their population temporal dynamics. Result A total of 3367 large moth specimens were collected, belonging to 269 species of 25 families. The dominant taxa were the families Geometidae, Crambidae, Sphingidae and Noctuidae. At the altitude ranges of 600 − 1 200 m, the Simpson index, Shannon index, Margalef richness index and Pielou uniformity index were high, and the overall correlation was positive. The species-multidegree curve of large moths tended to be lognormal distribution at this altitude gradient, and the rest of altitude gradients were consistent with the niche priority occupation hypothesis. The number of large moth species was the largest with the strongest stability in July. Conclusion The large moth species were sensitive to time in Mount Tianmu with the number of species and individuals of them increasing at first and then decreasing from May to August. Species diversity of large moths increased at first and then decreased with the increase of altitude, and the similarity coefficient between medium and low altitude gradients was at medium and high levels. [Ch, 3 fig. 4 tab. 20 ref.] -
Key words:
- altitude /
- large moth population /
- diversity analysis /
- temporal dynamics /
- Mount Tianmu
-
图 1 天目山不同海拔高度大蛾类种-多度曲线
Figure 1 Large moth species-multidegree curves at different altitudes of Mount Tianmu
图 2 天目山不同海拔系统聚类分析
Figure 2 Continental distance clustering analysis at different altitudes of Mount Tianmu
图 3 不同月份大蛾类的物种数和个体数
Figure 3 Number of species and individuals of large moth in different months
表 1 天目山不同海拔下大蛾类种群的组成
Table 1. Composition of large moth species at different altitudes of Mount Tianmu
科名 0~200 m 200~400 m 400~600 m 600~800 m 800~1 000 m 1 000~1 200 m 1 200~1 400 m 1 400~1 600 m 个体总
数/只S N S N S N S N S N S N S N S N 斑蛾科Zygaenidae 1 2 0 0 1 6 1 12 1 5 0 0 0 0 0 0 25 波纹蛾科Thyatiridae 1 9 1 11 2 10 2 25 2 20 1 6 1 1 0 0 82 蚕蛾科Bombycidae 1 3 1 8 1 3 2 12 1 4 1 5 1 1 0 0 36 草螟科Crambidae 15 29 16 43 25 53 26 69 23 71 15 44 7 21 3 8 338 尺蛾科Geometridae 19 49 20 65 31 89 53 116 57 122 34 93 22 53 11 28 615 刺蛾科Limacodidae 1 2 1 4 3 15 4 12 3 10 1 1 0 0 0 0 44 大蚕蛾科Saturniidae 2 8 4 15 7 30 8 45 7 30 4 14 5 14 0 0 156 带蛾科Thaumetopoeidae 0 0 1 2 1 4 1 9 1 7 0 0 0 0 0 0 22 灯蛾科Arctiidae 1 3 1 12 1 7 2 25 2 18 1 1 0 0 1 2 68 毒蛾科Lymantriidae 2 6 2 13 3 20 4 31 3 15 2 6 1 1 0 0 92 凤蛾科Epicopeiidae 1 2 0 0 1 1 1 1 1 4 0 0 0 0 0 0 8 钩蛾科Drepanidae 3 7 4 8 7 16 9 27 7 22 1 4 2 3 0 0 87 卷蛾科Tortricidae 1 9 1 7 0 0 1 7 1 3 0 0 0 0 0 0 26 枯叶蛾科Lasiocampidae 1 3 3 13 8 23 9 31 6 21 5 16 2 5 0 0 112 瘤蛾科Nolidae 1 3 2 6 3 13 3 22 3 13 2 4 0 0 0 0 61 箩纹蛾科Brahmaeidae 0 0 0 0 0 0 0 1 1 2 0 0 0 0 0 0 3 螟蛾科Pyralidae 2 7 4 15 5 23 6 56 5 47 6 27 2 8 3 8 191 木蠹蛾科Cossidae 2 10 1 2 2 8 3 9 1 8 1 1 0 0 0 0 38 裳蛾科Erebidae 2 7 4 15 7 25 11 40 9 27 6 13 1 3 0 0 130 苔蛾科Lithosiidae 1 2 1 10 1 20 1 43 1 28 1 34 1 6 0 0 143 天蛾科Sphingidae 12 28 13 35 24 56 31 112 30 113 26 80 8 20 5 13 457 网蛾科Thyrididae 1 4 2 11 3 20 3 20 2 14 1 4 1 1 0 0 74 燕蛾科Uraniidae 0 0 1 2 2 10 2 18 1 8 0 0 0 0 0 0 38 夜蛾科Noctuidae 5 16 7 22 11 47 35 79 40 78 23 59 17 36 2 11 348 舟蛾科Notodontidae 2 8 4 13 3 13 11 55 14 61 2 15 1 7 1 1 173 S合计 77 94 152 229 222 131 72 26 269 N合计 217 332 512 877 751 427 180 71 3367 S占比/% 28.62 34.94 56.51 85.13 82.53 49.44 26.77 9.67 N占比/% 6.44 9.86 15.21 26.05 22.30 12.68 5.35 2.10 说明:S. 物种数(种);N. 个体数(只)。各海拔梯度之间有相同物种。  下载: 导出CSV
下载: 导出CSV
表 2 天目山不同海拔梯度下大蛾类种群多样性指数
Table 2. Large moth population diversity index under different altitude gradients of Mount Tianmu
海拔/m Simpson指数 Shannon指数 Pielou均匀度 Margalef丰富度 0~200 0.869±0.040 bc 2.322±0.310 bc 0.345±0.040 b 3.903±0.400 a 200~400 0.901±0.010 abc 2.565±0.120 ab 0.381±0.010 a 3.617±0.160 a 400~600 0.910±0.010 ab 2.634±0.080 a 0.391±0.010 a 3.527±0.200 a 600~800 0.922±0.000 a 2.769±0.200 a 0.411±0.020 a 3.542±0.180 a 800~1 000 0.902±0.010 abc 2.605±0.090 ab 0.387±0.010 a 3.625±0.180 a 1 000~1 200 0.857±0.030 c 2.195±0.160 c 0.326±0.020 b 2.972±0.190 b 1 200~1 400 0.791±0.060 d 1.816±0.310 d 0.269±0.040 c 2.696±0.230 b 1 400~1 600 0.646±0.030 e 1.242±0.070 e 0.188±0.010 d 1.408±0.110 c 说明:数值为平均值±标准差。同列不同字母表示同一指数在不同海拔间差异显著(P<0.05)。
下载: 导出CSV
表 3 不同海拔天目山大蛾类种群相似性系数
Table 3. Similarity coefficients of large moth population at different altitudes of Mount Tianmu
海拔/m 0~200 m 200~400 m 400~600 m 600~800 m 800~1 000 m 1 000~1 200 m 1 200~1 400 m 1 400~1 600 m 0~200 1.000 0.833 0.875 0.880 0.880 0.864 0.682 0.318 200~400 1.000 0.875 0.880 0.880 0.864 0.682 0.318 400~600 1.000 0.657 0.657 0.826 0.652 0.304 600~800 1.000 0.900 0.760 0.600 0.280 800~1 000 1.000 0.760 0.600 0.280 1 000~1 200 1.000 0.789 0.368 1 200~1 400 1.000 0.467 1 400~1 600 1.000
下载: 导出CSV
表 4 不同月份大蛾类种群多样性
Table 4. New diversity of large moth species in different months
月份 个体
数/只Simpson
指数Shannon
指数Pielou
均匀度Margalef
丰富度5 708 0.798 2.162 0.329 2.668 6 795 0.819 2.263 0.339 3.231 7 996 0.819 2.438 0.353 3.534 8 868 0.788 2.210 0.327 2.876
下载: 导出CSV
-
[1] 张瀚文, 王敏, 李婧, 等. 不同海拔高度对夹金山草本层植物物种多样性与生物量的影响[J]. 应用与环境生物学报, 2023, 29(5): 1125 − 1131. ZHANG Hanwen, WANG Min, LI Jing, et al. Effects of different altitudes on herbaceous plant species diversity and biomass in Jiajin Mountain [J]. Chinese Journal of Applied and Environmental Biology, 2023, 29(5): 1125 − 1131. [2] 杨远亮, 孙鸿鸥, 曾全, 等. 四川九寨沟国家级自然保护区蛾类多样性调查[J]. 四川林业科技, 2023, 44(6): 95 − 100. YANG Yuanliang, SUN Hongou, ZENG Quan, et al. Investigation on moth diversity in Jiuzhaigou National Nature Reserve, Sichuan Province [J]. Journal of Sichuan Forestry Science and Technology, 2023, 44(6): 95 − 100. [3] 金哲雅, 汤健, 武庚风, 等. 镜泊湖大蛾类昆虫物种多样性研究[J]. 安徽农业科学, 2014, 42(4): 1063 − 1065. JIN Zheya, Tang Jian, WU Gengfeng, et al. Study on the diversity of big moths from Jingpohu [J]. Journal of Anhui Agricultural Sciences, 2014, 42(4): 1063 − 1065. [4] 陈梦悦. 湖南金童山国家级自然保护区双江口工区蛾类及年发生动态[D]. 长沙: 湖南农业大学, 2023. CHEN Mengyue. Moths and its Annual Occurrence Dynamics in Shuangjiangkou Area of Jintongshan National Nature Reserve in Hunan Province [D]. Changsha: Hunan Agricultural University, 2023. [5] 徐华潮, 吴鸿, 杨淑贞, 等. 浙江天目山昆虫物种多样性研究[J]. 浙江林学院学报, 2002, 19(4): 350 − 355. XU Huachao, WU Hong, YANG Shuzhen, et al. Insect species diversity of Mount Tianmu in China [J]. Journal of Zhejiang Forestry College, 2002, 19(4): 350 − 355. [6] 曹春婧, 何建龙, 王占军, 等. 宁夏不同区域欧李园昆虫群落多样性[J]. 浙江农林大学学报, 2021, 38(6): 1253 − 1260. CAO Chunjing, HE Jianlong, WANG Zhanjun, et al. Insect community diversity of Cerasus humilis orchards in different regions of Ningxia [J]. Journal of Zhejiang A&F University, 2021, 38(6): 1253 − 1260. [7] 张宏伟. 外来植物对浙江天目山植物多样性海拔梯度格局的影响[D]. 上海: 华东师范大学, 2022. ZHANG Hongwei. The Effects of Alien Species on Plants Diversity Patterns along an Elevational Gradient in Mt. Tianmu [D]. Shanghai: East China Normal University, 2022. [8] 周阳, 权子豪, 段曼微, 等. 基于林分昆虫多样性调查的传统灯诱法改进效果分析[J]. 生态学报, 2023, 43(16): 6851 − 6861. ZHOU Yang, QUAN Zihao, DUAN Manwei, et al. Effect of the improvement of traditional light trapping method based on the investigation of insect diversity in different forest stands [J]. Acta Ecological Sinica, 2023, 43(16): 6851 − 6861. [9] 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ: α多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231 − 239. MA Keping, LIU Yuming. Methods for measuring community diversity I: α methods for measuring diversity (Part II) [J]. Biodiversity Science, 1994, 2(4): 231 − 239. [10] 唐启义. DPS数据处理系统-实验设计, 统计分析及数据挖掘[M]. 北京: 科学出版社, 2010. TANG Qiyi. DPS Data Processing System-experimental Design, Statistical Analysis and Data Mining [M]. Beijing: Science Press, 2010. [11] 任雪毓, 庞岩, 王梅, 等. 北方蒙古栎和栓皮栎林蛾类多样性比较分析[J]. 林业科学研究, 2022, 35(6): 143 − 150. REN Xueyu, PANG Yan, WANG Mei, et al. Analysis of moth diversity in oak forests, Quercus variabilis and Q. mongolica in Northern China [J]. Forest Research, 2022, 35(6): 143 − 150. [12] 段曼微, 李香, 周阳, 等. 基于蛾类多样性研究人工林斑块的边缘效应[J]. 生物多样性, 2023, 31(5): 69 − 79. DUAN Manwei, LI Xiang, ZHOU Yang, et al. Edge effect in plantation patches based on moth diversity [J]. Biodiversity Science, 2023, 31(5): 69 − 79. [13] 孙儒泳. 动物生态学原理[M]. 北京: 北京师范大学出版社, 2001. SUN Ruyong. Principles of Animal Ecology [M]. Beijing: Beijing Normal University Press, 2001. [14] 王义平, 吴鸿, 徐华潮. 以昆虫作为指示生物评估森林健康的生物学与生态学基础[J]. 应用生态学报, 2008, 19(7): 1625 − 1630. WANG Yiping, WU Hong, XU Huachao. Biological and ecological basis for assessing forest health using insects as indicator organisms [J]. Chinese Journal of Applied Ecology, 2008, 19(7): 1625 − 1630. [15] 楼一恺, 范忆, 戴其林, 等. 天目山常绿落叶阔叶林群落垂直结构与群落整体物种多样性的关系[J]. 生态学报, 2021, 41(21): 8568 − 8577. LOU Yikai, FAN Yi, DAI Qilin, et al. Relationship between vertical structure and overall species diversity in an evergreen deciduous broad-leaved forest community of Tianmu Mountain Natural Reserve [J]. Acta Ecological Sinica, 2021, 41(21): 8568 − 8577. [16] 孙媛, 蓝陈仪航, 章振, 等. 桃潜叶蛾生物学特性及防治研究进展[J]. 浙江农林大学学报, 2022, 39(3): 687 − 694. SUN Yuan, LANCHEN Yihang, ZHANG Zhen, et al. Advances in biological characteristics and control of Lyonetia clerkella [J]. Journal of Zhejiang A&F University, 2022, 39(3): 687 − 694. [17] 吴鸿, 潘承文. 天目山昆虫[M]. 北京: 科学出版社, 2001. WU Hong, PAN Chengwen. Insects of Tianmushan National Nature Reserve [M]. Beijing: Science Press, 2001. [18] 来锡福, 焦旭东, 王多文, 等. 武威地区苗圃地灯下蛾类昆虫多样性及时序动态分析[J]. 中国农学通报, 2022, 38(22): 104 − 109. LAI Xifu, JIAO Xudong, WANG Duowen, et al. Diversity and temporal dynamics of moths under an ultraviolet lamp in nursery in Wuwei Area [J]. Chinese Agricultural Science Bulletin, 2022, 38(22): 104 − 109. [19] 杨美霞, 党蕊, 丁昌萍, 等. 陕西子午岭国家级自然保护区昆虫多样性调查[J]. 延安大学学报(自然科学版), 2021, 40(4): 102 − 107. YANG Meixia, DANG Rui, DING Changping, et al. Compare the molecular interaction force of organic compounds based on smartphone temperature sensor [J]. Journal of Yan’an University (Natural Science Edition), 2021, 40(4): 102 − 107. [20] 居峰, 董丽娜, 陈希, 等. 不同森林植被类型蛾类群落结构及其多样性研究[J]. 江苏林业科技, 2011, 38(1): 1 − 6, 11. JU Feng, DONG Lina, CHEN Xi, et al. The community structure and diversity study of moths at the different vegetation types in Zijin mountain area [J]. Jiangsu Forestry Science and Technology, 2011, 38(1): 1 − 6, 11. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230469
 点击查看大图
点击查看大图
计量
- 文章访问数: 1951
- HTML全文浏览量: 358
- PDF下载量: 71
- 被引次数: 0
