




-
香椿属Toona是楝科Meliaceae中一个较为重要的属,该属树种材质优良、纹理直、具光泽,为上等家具及室内装饰用材,被称为“中国桃花心木”[1]。其中香椿T. sinensis是中国特有的珍贵速生用材树种,落叶乔木,树冠庞大,树干通直,生长迅速,一般10 a左右即可成材,其木材细密富有弹性,纹理通直美观、色泽红润,耐腐。同时,香椿嫩叶具有浓郁的香气,在中国具有悠久的栽培历史[2],早在明代徐光启就将其作为救饥植物载入《农政全书》,是国人喜食的山珍野菜。香椿经济价值较高,具有极大的开发利用潜力。
目前,对香椿的研究集中在分类[3−4]、栽培繁育[5−6]、营养价值应用[7−8]、药用[9]和遗传改良[10−14]等方面。香椿作为珍贵优质阔叶用材树种,育种研究起步较晚。20世纪80年代,孙鸿有等[10]开展了香椿种源试验,综合选育出浙江丽水、湖南邵阳等6个优良种源。随后河南[11]、江苏[12]、湖北[13]和陕西[14]等省陆续开展了香椿遗传改良的研究,在推动香椿人工林发展中起到了重要作用。香椿分布区较广,蕴藏着丰富的遗传变异。前期通过全面收集香椿分布区24个种源种子,开展了苗期测定,筛选出苗期表现比较优异的7个种源[15]。
种源和家系选择是林木遗传改良的重要手段之一。国内外开展的主要造林树种种源试验,证明种源选择能取得良好的改良效果。国内前期虽营建了多个香椿种源试验林,但香椿育种周期长,多数试验林没有得到有效保存和维护,未对其种源试验结果进行长期跟踪评价。鉴于此,本研究利用浙江省开化县林场保存完好的10年生香椿种源试验林,研究其生长和形质性状的地理种源变异,通过聚类分析开展种源区划分,并筛选速生优质的香椿种源,为香椿的种质资源选择评价提供理论基础,也为今后的香椿良种选育工作提供参考。
-
研究区位于浙江省开化县林场国家杉木良种基地。该区属中亚热带季风气候区,四季分明,冬夏长,春秋短。年平均气温为16.6 ℃,无霜期为254.0 d,年平均降水量为1 830.8 mm。土壤以红壤为主,质地中壤至轻黏,酸性。造林地前期为杉木Cunninghamia lanceolata林,于2011年被砍伐。2012年春季营建香椿种源试验林。试验采用随机区组设计,10株单列小区,3次重复,即每个种源30株,株行距为2.0 m×2.5 m。
-
研究材料包括香椿种源24个(表1)。2021年12月初对试验林进行全林调查,测定树高、胸径、枝下高、冠幅等性状,并计算获得树干圆满度(用树干下部3 m高处直径与胸径之比)和树冠圆满度(冠幅与其树冠长度之比) [16−17]。
表 1 香椿24个种源的地理位置
Table 1. Locations of the twenty-four provenances of T. sinensis
种源编号 种源 经度(E) 纬度(N) 海拔/m 种源编号 种源 经度(E) 纬度(N) 海拔/m 1 三门峡 111.047° 34.054° 779 15 峨眉山 103.480° 29.590° 877 2 郴州 113.032° 25.793° 402 20 凭祥 106.755° 22.105° 348 3 湘西 109.601° 27.948° 389 21 晴隆 105.218° 25.834° 1102 4 武汉 114.298° 30.584° 26 23 潍坊 118.544° 36.512° 492 5 新宁 110.851° 26.429° 859 24 聊城 117.635° 26.265° 361 8 邯郸 114.490° 36.612° 91 25 杭州 119.960° 30.048° 100 9 九江 115.992° 29.712° 17 26 阜阳 115.621° 33.160° 34 10 南京 118.767° 32.041° 14 27 运城 112.549° 37.857° 885 11 新乡 113.805° 35.190° 76 34 恩施 109.479° 30.295° 718 12 南阳 111.481° 33.297° 376 36 乳源 113.280° 23.125° 120 13 伏牛山 119.419° 32.042° 33 38 元谋 101.877° 25.704° 1 652 14 天水 105.724° 34.578° 1 355 42 安康 109.760° 32.804° 526 -
从瑞士联邦森林、雪和景观研究所网站(Chelsa Climate, http://chelsa-climate. org/ downloads/)下载高分辨率气象因子。土壤数据采自 HWSD-V 1.2 数据。
-
材积计算公式如下:
$$ V= 0.000\; 052\; 76D ^{1.882 \;161}H^{ 1.009\; 317}。 $$ (1) 式(1)中:V为单株材积(m3) ;D为胸径(cm);H为树高(m)。统计分析以小区平均数为单位,按常规统计方法进行。计算采用DPS软件在计算机上进行,对测定的性状值以及数据转换值进行单因素方差分析。线性模型为:
$$ X_{i j}=\mu+\alpha_i+\varepsilon_{i j} 。 $$ (2) 式(2)中:Xij为第i个种源某性状的第j个观察值,µ为总体平均值,αi为种源的某性状效应, εij为随机误差。对各性状的广义遗传力进行估算,公式为:
$$ h^{ 2} = 1-1/F。 $$ (3) 式(3)中:h 2为表型遗传力,F为方差分析中的F值。优良种源评价及选择参考李广友等[18]和胡兴峰等[19]的综合选择指数方程,公式为:
$$ {I}_{i}=\sum _{i=1}^{n}{w}_{i}{h}^{2}{P}_{n} 。 $$ (4) 式(4):Ii为种源选择指数值,wi为某性状的经济权重,h 2为性状遗传力,Pn为某种源第n个性状的表型值。遗传增益计算公式为:
$$ \Delta G_i=\frac{s h^2}{\overline X} \times 100 \%。 $$ (5) 式(5)中:ΔGi为第i个种源的遗传增益;s为选择差;h 2为性状遗传力;$ \overline{X} $为种源表型均值。
-
主成分、相关分析等采用R语言corrplot、FactoMineR、factoextra等程序包进行。
-
香椿种源生长和形质性状方差分析结果(表2)显示:树高、胸径、枝下高、冠幅和材积在重复间差异显著(P<0.05)或极显著(P<0.01);不同种源间树高、胸径、树干圆满度和材积差异极显著(P<0.01),这说明其地理种源间的遗传变异丰富,优良种源选择的潜力很大。10年生时种源树高平均为8.86 m,变幅为6.10~10.90 m;胸径均值为8.07 cm,变幅为5.57~10.61 cm,胸径生长量最大种源是最小种源的1.9倍。种源单株材积均值为0.026 m3,变幅为0.008~0.042 m3,其变异幅度较大,材积生长量最大种源是最小种源的5.25倍。香椿不同种源间树干圆满度差异极显著(P<0.01),而树冠圆满度在种源间差异不显著。树高、胸径、冠幅、枝下高、树干圆满度、树冠圆满度和材积等7个性状变异系数分别为9.5%、13.1%、12.3%、15.7%、2.9%、12.3%和35.9%,从变异系数也可看出:材积的变异最大,其次为枝下高和胸径。以上7个性状的广义遗传力分别为0.65、0.69、0.33、0.42、0.76、0.42和0.57,说明香椿树高、胸径、树干圆满度和材积等性状在种源水平上受较强的遗传控制。
表 2 香椿种源生长和形质性状的方差分析及主要遗传参数估计
Table 2. Variance analysis and main genetic parameters of growth and stem-form of T. sinensis
性状 变异来源 遗传参数 重复 种源 误差 均值 变幅 变异系数/% 树高 46.285 7** 60.273 2** 42.210 8.86 6.10~10.90 9.5 胸径 72.938 1** 93.919 4** 57.900 8.07 5.57~10.61 13.1 冠幅 1.389 9* 5.865 2 7.851 2.97 2.19~3.51 12.3 枝下高 10.944 8** 20.060 2 23.200 3.89 2.51~4.66 15.7 树干圆满度 0.036 3** 0.561 2** 0.028 0.80 0.56~0.93 2.9 树冠圆满度 0.080 4 0.540 6 0.626 0.62 0.46~0.79 12.3 材积 0.004 7** 0.005 1** 0.004 0.026 0.008~0.042 35.9 说明:*. P<0.05;**. P<0.01。遗传参数中的树高、胸径、冠幅、枝下高和材积的单位分别是m、cm、m、m和m3。 -
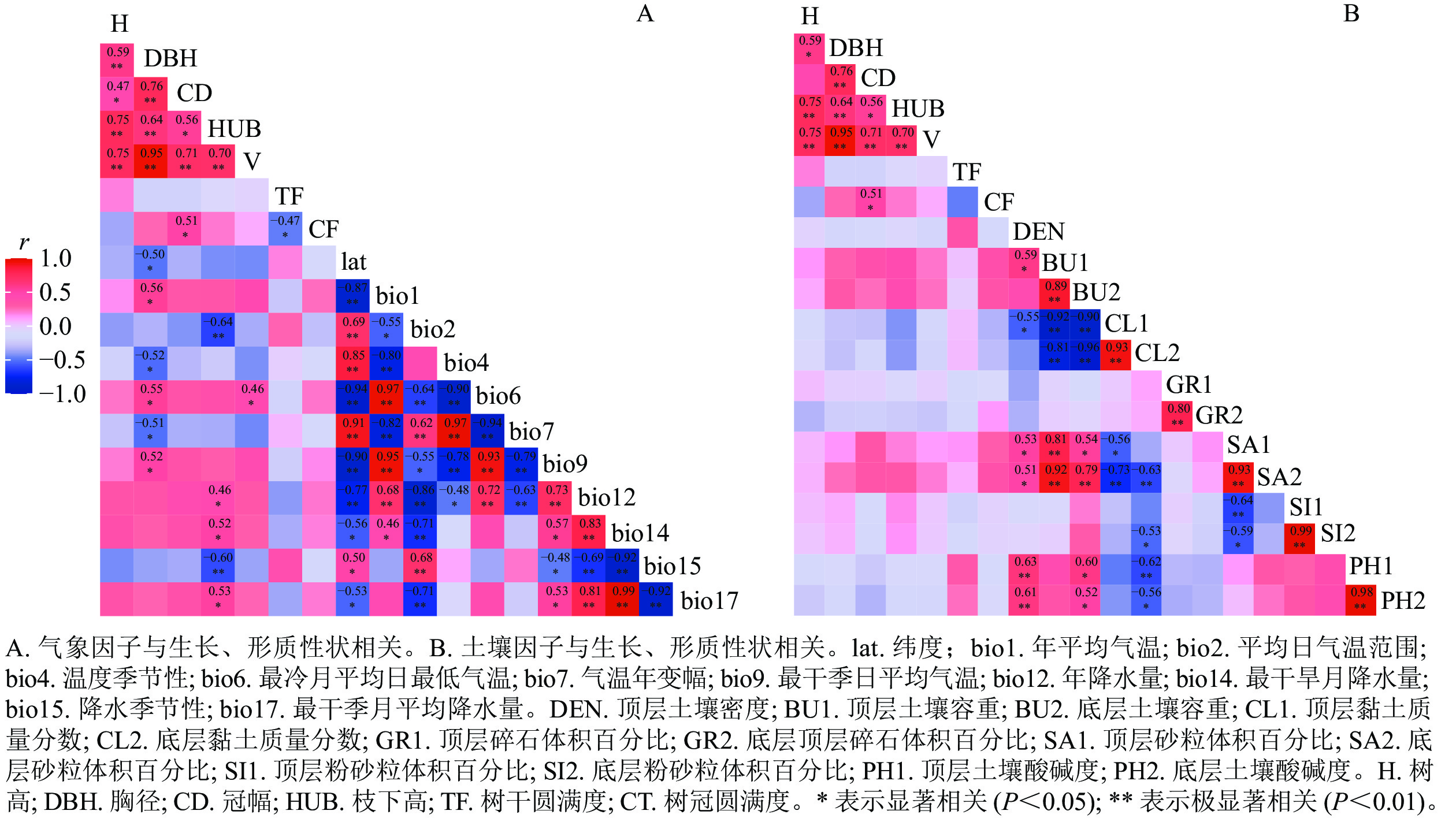
从香椿不同种源树高、胸径、冠幅、枝下高、树干圆满度、树冠圆满度和材积等生长和形质性状与产地生态因子之间的相关关系(图1A)可知:香椿种源胸径与年平均气温、最冷月平均日最低气温和最干季日平均气温显著(P<0.05)正相关,与纬度、温度季节性、气温年变幅显著(P<0.05)负相关,说明香椿种源以上这些生长、形质性状呈现典型的纬度变异模式,但冠幅圆满度与纬度相关性不显著,表明其为随机的地理变异模式。香椿种源枝下高与年降水量、最干旱月降水量(bio14)、最干季月平均降水量显著(P<0.05)正相关,而与平均日气温范围和降水季节性极显著(P<0.01)负相关。香椿种源材积与最冷月平均日最低气温(bio6)显著(P<0.05)正相关。香椿种源生长、形质性状与产地土壤因子相关不显著,说明产地土壤对香椿种源生长影响较小(图1B)。
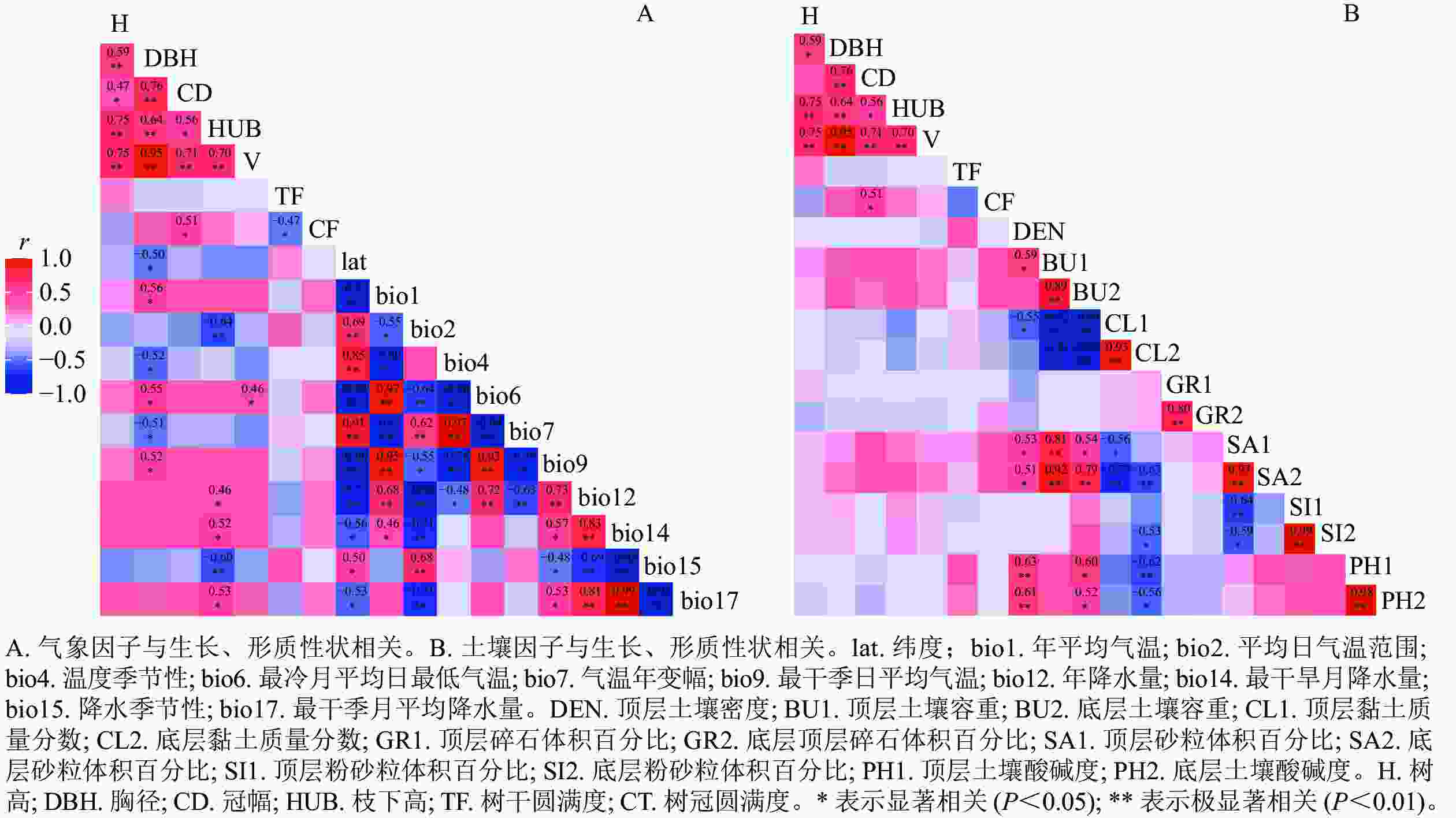
图 1 香椿种源生长、形质性状与产地生态因子的相关分析
Figure 1. Correlation between growth and stem-form quality of different provenances and the ecological factors of the provenances
-
香椿种源生长和形质性状表型相关和遗传相关分析发现(表3):10年生香椿种源胸径与冠幅、枝下高和材积之间的遗传相关和表型相关均呈极显著(P<0.01)或显著(P<0.05)正相关,说明在进行香椿种源胸径改良的同时,可以改良其冠幅、枝下高和材积等生长性状。
表 3 香椿种源生长、形质性状表型相关和遗传相关
Table 3. Phenotypic and genetic correlation between growth and stem-form qualities traits of T. sinensis
性状 树高 胸径 冠幅 枝下高 材积 树干圆满度 树冠圆满度 树高 1 0.569 8 0.549 7 0.990 7** 0.755 9 0.214 8 −0.394 5 胸径 0.614 8* 1 0.965 8** 0.988 8** 0.990 9** −0.276 5 0.605 9 冠幅 0.491 7 0.750 4** 1 0.976 9** 0.970 4** −0.370 5 0.752 8 枝下高 0.752 5** 0.651 2* 0.576 0 1 0.990 8** −0.198 1 0.025 6 材积 0.772 0** 0.951 9** 0.695 9* 0.712 7** 1 −0.091 7 0.367 3 树干圆满度 0.187 6 −0.185 9 −0.181 5 −0.080 3 −0.017 1 1 −0.762 1 树冠圆满度 −0.360 3 0.232 5 0.489 0 0.223 1 0.049 5 −0.460 5 1 说明:对角线下为表型相关系数,对角线上为遗传相关系数;*. P<0.05;**. P<0.01。 -
香椿生长和形质性状主成分分析结果(表4)显示:主成分1特征值为3.822 8,贡献率为54.611%,其中树高、胸径、冠幅、枝下高和材积的特征值的绝对值相对较高,分别为0.390 8、0.474 7、0.428 0、0.427 1和0.482 2。主成分2的特征值为1.791 5,贡献率为25.593%,其中树冠圆满度和树干圆满度的特征值绝对值相对较高,分别为0.659 1和0.555 8。因此,主成分1可代表生长性状,主成分2可代表形质性状,能够综合反映香椿种源生长和形质性状的大部分信息。以特征值大于1为标准提取的前2个主成分,累计贡献率达80.204%,说明这2个主成分可以包含所有性状的80.204%信息。由表4中各生长和形质性状的载荷量可以得到主成分的线性方程,用 x1、x2、x3、x4、x5、x6、x7 分别表示树高、胸径、冠幅、枝下高、材积、树干圆满度和树冠圆满度,用 y1、y2分别表示主成分1和主成分2,得到如下线性方程:y1=0.390 8x1+0.474 7x2+0.428 0x3+0.427 1x4+0.482 2x5−0.078 3 x6+0.133 0x7;y2=0.433 5x1−0.059 2x2−0.219 3x3+0.073 7 x4+0.108 3x5+0.555 8x6−0.659 1x7。
表 4 香椿种源各性状主成分分析
Table 4. Principal component analysis of different traits for provenances of T. sinensis
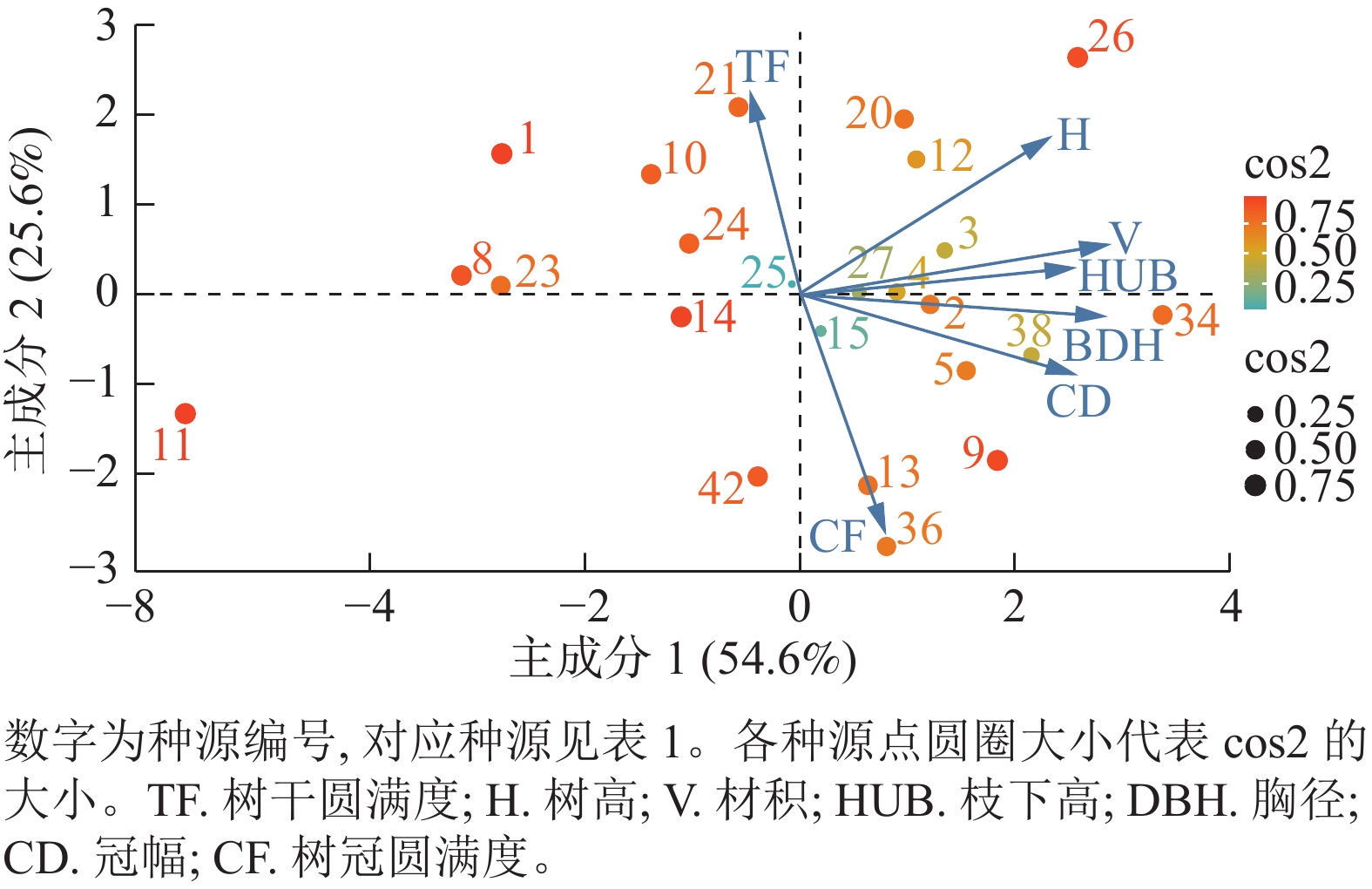
主成分 树高 胸径 冠幅 枝下高 材积 树干圆满度 树冠圆满度 特征值 贡献率/% 累积贡献率/% 主成分1 0.390 8 0.474 7 0.428 0 0.427 1 0.482 2 −0.078 3 0.133 0 3.822 8 54.611 54.611 主成分2 0.433 5 −0.059 2 −0.219 3 0.073 7 0.108 3 0.555 8 −0.659 1 1.791 5 25.593 80.204 主成分3 0.221 5 0.013 6 −0.343 0 0.095 8 0.008 9 −0.809 0 −0.411 5 0.635 9 9.084 89.287 主成分4 0.181 8 −0.432 6 −0.138 8 0.755 5 −0.335 2 0.047 7 0.274 2 0.477 3 6.820 96.106 主成分5 0.349 3 −0.372 8 0.722 1 −0.232 0 −0.325 1 −0.153 4 −0.185 7 0.247 0 3.529 99.635 主成分6 0.154 7 −0.630 2 −0.105 5 −0.194 5 0.686 5 −0.058 5 0.235 1 0.021 0 0.299 99.934 主成分7 0.656 8 0.218 5 −0.315 0 −0.374 9 −0.257 4 0.034 8 0.462 2 0.004 6 0.066 100.000 以主成分1和主成分2绘制了香椿所有种源主成分样点图以及性状与主成分的关系图(图2)。结合表2和图2可以看出:树高、胸径、材积、枝下高和冠幅5个性状距离x轴更接近,说明主成分1的差异主要是这5个性状的贡献,其中材积的贡献最大,其次为胸径。树干圆满度和树冠圆满度2个性状与y轴接近,说明主成分2的差异主要是这2个性状的贡献。从各种源某个主成分代表性的特征值(cos2)和分布来看,元谋、恩施和太和等种源具有速生的特点,新乡、邯郸和四川等种源生长速度较慢。

图 2 香椿种源生长和形质性状主成分分析图
Figure 2. Principal component analysis of different traits for provenances of T. sinensis
-
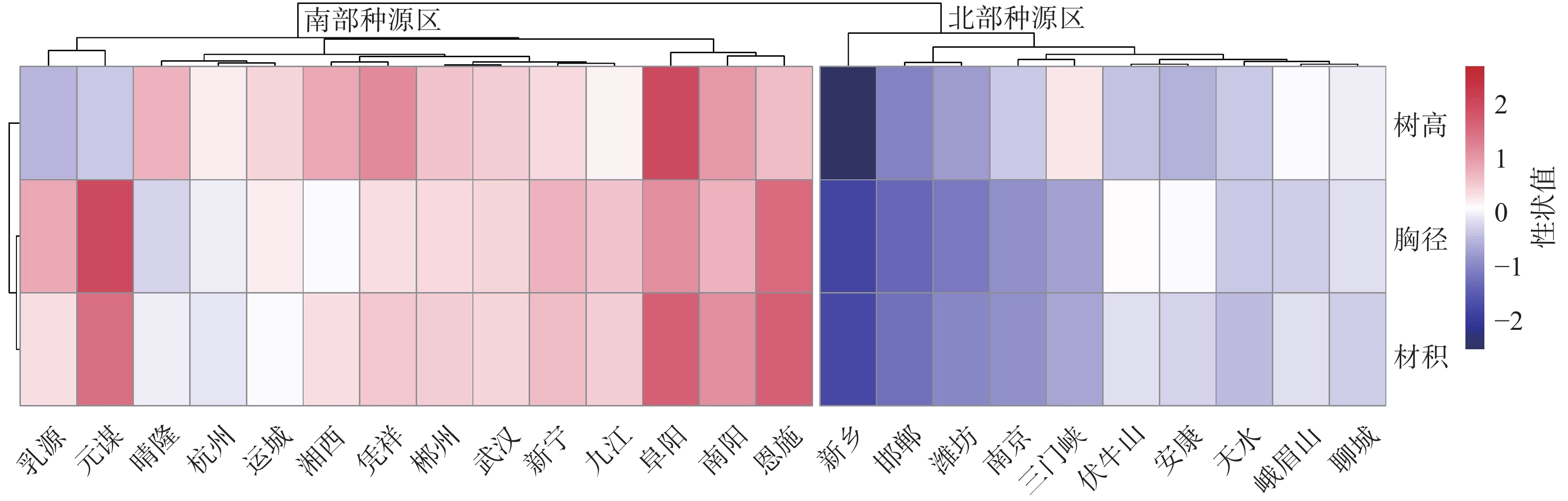
采用树高、胸径和材积3个生长性状的种源均值,对所有种源进行聚类(图3),可以将24个种源划分为2个组,即南部种源区和北部种源区。南部种源区可划分为3个亚组:亚组SouthⅠ包括了华南和西南区域的凭祥、晴隆、元谋和乳源等,亚组SouthⅡ包括了华中和长江以南华东区域的郴州和湘西、九江和恩施等,亚组SouthⅢ包括了伏牛山以南和皖北淮河流域的阜阳和南阳等。北部种源区也可划分为3个亚组:亚组NorthⅠ包括西北区域的天水等,亚组NorthⅡ包括了秦岭山区的安康等,亚组NorthⅢ包括伏牛山区、太行山区和山东丘陵区域的三门峡和新乡、邯郸和潍坊等。

图 3 香椿种源聚类热图
Figure 3. Cluster analysis heatmap for provenances of T. sinensis
-
把主成分1和主成分2的贡献率作为权重(y1, y2),计算各种源综合评价得分。由主成分分析结果可知:前2个主成分的权重分别为54.611%和25.593%,所以,综合评价得分(W)计算公式为:W=54.611%×y1+25.593%×y2。利用该计算公式,获得香椿各种源的综合评价得分(表5)。
表 5 香椿种源综合评价得分和排名表
Table 5. Comprehensive evaluation scores and ranking of provenances
种源 综合评价得分 排名 种源 综合评价得分 排名 种源 综合评价得分 排名 阜阳 7.496 1 九江 6.732 9 南京 6.166 17 恩施 7.253 2 武汉 6.721 10 聊城 6.166 18 凭祥 6.977 3 运城 6.566 11 安康 6.087 19 南阳 6.913 4 晴隆 6.423 12 天水 6.008 20 元谋 6.883 5 杭州 6.372 13 三门峡 5.729 21 湘西 6.844 6 乳源 6.355 14 潍坊 5.418 22 新宁 6.827 7 峨眉山 6.351 15 邯郸 5.350 23 郴州 6.752 8 伏牛山 6.348 16 新乡 4.376 24 基于主成分分析和各种源综合评价得分结果,筛选出元谋、恩施和阜阳等3个优良种源,其树高、胸径、材积、冠幅、枝下高、树干圆满度和树冠圆满度获得的平均遗传增益分别为5.21%、16.50%、32.88%、3.56%、3.13%、6.65%和4.74% (表6)。
表 6 香椿3个优良种源各性状均值和遗传增益
Table 6. Means and genetic gain of various traits of the top 3 provenances of T. sinensis
种源 树高/m 胸径/cm 材积/m3 冠幅/m 枝下高/m 树干圆满度 树冠圆满度 元谋 8.4 10.61 0.039 8 3.25 3.92 0.87 0.71 恩施 9.4 9.95 0.042 2 3.51 4.43 0.88 0.72 阜阳 10.9 9.43 0.042 1 3.10 4.19 0.85 0.65 总体均值 8.86 8.07 0.0260 2.97 3.89 0.80 0.62 入选种源平均值 9.57 10.00 0.041 0 3.29 4.18 0.87 0.69 遗传增益/% 5.21 16.50 32.88 3.56 3.13 6.65 4.74 -
本研究利用已达半个轮伐期的香椿地理种源试验发现:10年生香椿种源间树高、胸径、树干圆满度和材积等性状差异显著。树高、胸径、冠幅、枝下高和材积等性状均与纬度呈负相关,其中胸径与纬度显著负相关,南方种源的生产力明显高于北方种源。这个结果与香椿早期研究结果一致[10 , 15],证实了香椿地理种源生长性状基本上呈现纬度变异形式。生长、 形质性状与种源产地生态因子的相关分析表明:树高、胸径、冠幅、枝下高和树干圆满度等纬度变异主要与产地气温和降水有关,而枝下高还与土壤容重和黏土质量分数有关。这与马尾松Pinus massoniana[19]、文冠果Xanthoceras sorbifolia [20]、杉木 [21]的研究结果相似,香椿种源生长性状与产地水热条件相关。在对杉木[21]研究发现:影响种源生长的主要因素是冬季平均气温和秋冬季降水,而本研究发现香椿种源产地年平均气温、最冷月平均日最低气温和最干季日平均气温等生态因子是影响胸径生长的主要因素。SILVESTRO等[22]对云杉Picea asperata研究也发现:产地温度升高1 ℃,其生长速度增加0.1 cm·d−1。但这有异于MADSEN等[23]的相关研究结果:栎Quercus树的生存主要取决于产地的气温,而生长更多地依赖于产地的降水。从香椿种源生长、形质性状与产地土壤因子相关不显著来看,其生长和形质性状主要与造林地土壤相关理化因子有关。
本研究发现:香椿种源胸径与冠幅、枝下高和材积之间呈极显著的正遗传相关,这与香椿已有的研究结果一致[10]。对香椿胸径进行选择,同时也可对冠幅、枝下高和材积等性状进行改良,但对形质性状没有明显的改良效果。所以今后对香椿进行材积改良时,胸径可以作为重要的目标性状。
林木种源区划分能指导种子的合理调拨,提高林木的生产力[24−25]。本研究根据10年生香椿的生长性状将香椿种源划分为南部和北部两大种源区,基本上以秦岭—淮河为南、北部种源区分界线。南部种源区包括了乳源、元谋、凭祥和晴隆等华南和西南区域;郴州、湘西、新宁、武汉、恩施和杭州等华中和长江以南华东区域,南阳和阜阳等伏牛山以南和皖北淮河流域。北部种源区包括了天水等西北区域;安康、峨眉山等秦巴山区;新乡、三门峡、邯郸、潍坊和运城等伏牛山区、太行山区和山东丘陵区域。本研究根据种源性状主成分分析和综合评价得分,选出了3个优良种源,均来自南部种源区。孙鸿有等[10]确定的优良种源也是来自南部种源区,由于采样地点不同,种源来源地略有差异,但本研究结果与刘军等[15]的苗期种源选择结果差异较大。
Provenance variation and selection in growth, shape, and quality traits of 10-year-old Toona sinensis
-
摘要:
目的 通过分析10年生香椿Toona sinensis生长与形质性状的种源变异,揭示不同性状在地理种源上的变异规律,并进行种源区划及优良种源选择。 方法 以浙江开化县林场的10年生香椿种源林为试材,测定其树高、胸径、枝下高、冠幅、材积、树干圆满度和树冠圆满度,获取各种源生态环境因子,进行方差、主成分、相关性和聚类等分析,计算广义遗传力、综合选择指数和遗传增益,阐明生长和形质性状地理变异模式及与产地生态因子关系。 结果 树高、胸径、树干圆满度和材积存在极显著(P<0.01)种源差异,种源胸径、树高、冠幅、枝下高和材积呈典型的纬向变异模式。种源胸径与冠幅、枝下高和材积之间均呈极显著(P<0.01)正相关。胸径与产地年平均气温、最冷月平均日最低气温和最干燥季节日平均气温等显著(P<0.05)正相关。根据种源聚类结果,可将香椿划分为南部和北部2个种源区,并筛选出3个优良种源,分别为元谋、恩施和太和。 结论 10年生香椿种源间差异显著,并且树高、胸径、材积等性状存在从南向北逐渐减小的变异规律,对其进行选择时胸径是主要的遗传改良性状,可为材用型香椿良种选择提供可靠依据。图3表6参25 Abstract:Objective The purpose of this study is to examine the provenance variations in growth, shape and quality traits of 10-year-old Toona sinensis, to reveal the variation of different traits in geographical provenance, and to carry out provenance division and selection. Method Using the 10-year-old T. sinensis provenance test in Kaihua County, Zhejiang Province as material, the tree height, diameter at breast height (DBH), height under the branch, crown width, volume, trunk completeness, and crown completeness were measured, and various ecological and environmental factors were obtained. Variance, principal component, correlation, and cluster analysis were applied to calculate the general heritability, comprehensive selection index, and genetic gain to elucidate the geographical variation patterns of growth, shape, and quality traits and their relationship with ecological factors of different origin. Result There were significant provenance differences (P<0.01) in tree height, DBH, trunk completeness, and volume. The DBH, tree height, crown width, height under branches, and volume of the provenances displayed typical zonal variation patterns. There was a highly significant positive correlation (P<0.01) between DBH and crown width, height under branches and volume. DBH was positively correlated (P<0.05) with the annual mean temperature, the average daily minimum temperature in the coldest month, and the average daily mean temperature in the driest season. According to the clustering results of all provenances, T. sinensis could be divided into two provenance zones: south and north. Three excellent provenances were selected, namely Yuanmou, Enshi and Taihe. Conclusion There are significant differences among the provenances of 10-year-old T. sinensis, with a gradual decrease in the variation of tree height, DBH, and volume from south to north. DBH is the main genetic improvement trait during selection, which can provide a reliable basis for the selection of high-quality T. sinensis varieties for timber use. [Ch, 3 fig. 6 tab. 25 ref.] -
Key words:
- Toona sinensis /
- genetic variation /
- latitude variation /
- temperature /
- provenance zone
-
图 1 香椿种源生长、形质性状与产地生态因子的相关分析
Figure 1 Correlation between growth and stem-form quality of different provenances and the ecological factors of the provenances
图 2 香椿种源生长和形质性状主成分分析图
Figure 2 Principal component analysis of different traits for provenances of T. sinensis
表 1 香椿24个种源的地理位置
Table 1. Locations of the twenty-four provenances of T. sinensis
种源编号 种源 经度(E) 纬度(N) 海拔/m 种源编号 种源 经度(E) 纬度(N) 海拔/m 1 三门峡 111.047° 34.054° 779 15 峨眉山 103.480° 29.590° 877 2 郴州 113.032° 25.793° 402 20 凭祥 106.755° 22.105° 348 3 湘西 109.601° 27.948° 389 21 晴隆 105.218° 25.834° 1102 4 武汉 114.298° 30.584° 26 23 潍坊 118.544° 36.512° 492 5 新宁 110.851° 26.429° 859 24 聊城 117.635° 26.265° 361 8 邯郸 114.490° 36.612° 91 25 杭州 119.960° 30.048° 100 9 九江 115.992° 29.712° 17 26 阜阳 115.621° 33.160° 34 10 南京 118.767° 32.041° 14 27 运城 112.549° 37.857° 885 11 新乡 113.805° 35.190° 76 34 恩施 109.479° 30.295° 718 12 南阳 111.481° 33.297° 376 36 乳源 113.280° 23.125° 120 13 伏牛山 119.419° 32.042° 33 38 元谋 101.877° 25.704° 1 652 14 天水 105.724° 34.578° 1 355 42 安康 109.760° 32.804° 526  下载: 导出CSV
下载: 导出CSV
表 2 香椿种源生长和形质性状的方差分析及主要遗传参数估计
Table 2. Variance analysis and main genetic parameters of growth and stem-form of T. sinensis
性状 变异来源 遗传参数 重复 种源 误差 均值 变幅 变异系数/% 树高 46.285 7** 60.273 2** 42.210 8.86 6.10~10.90 9.5 胸径 72.938 1** 93.919 4** 57.900 8.07 5.57~10.61 13.1 冠幅 1.389 9* 5.865 2 7.851 2.97 2.19~3.51 12.3 枝下高 10.944 8** 20.060 2 23.200 3.89 2.51~4.66 15.7 树干圆满度 0.036 3** 0.561 2** 0.028 0.80 0.56~0.93 2.9 树冠圆满度 0.080 4 0.540 6 0.626 0.62 0.46~0.79 12.3 材积 0.004 7** 0.005 1** 0.004 0.026 0.008~0.042 35.9 说明:*. P<0.05;**. P<0.01。遗传参数中的树高、胸径、冠幅、枝下高和材积的单位分别是m、cm、m、m和m3。
下载: 导出CSV
表 3 香椿种源生长、形质性状表型相关和遗传相关
Table 3. Phenotypic and genetic correlation between growth and stem-form qualities traits of T. sinensis
性状 树高 胸径 冠幅 枝下高 材积 树干圆满度 树冠圆满度 树高 1 0.569 8 0.549 7 0.990 7** 0.755 9 0.214 8 −0.394 5 胸径 0.614 8* 1 0.965 8** 0.988 8** 0.990 9** −0.276 5 0.605 9 冠幅 0.491 7 0.750 4** 1 0.976 9** 0.970 4** −0.370 5 0.752 8 枝下高 0.752 5** 0.651 2* 0.576 0 1 0.990 8** −0.198 1 0.025 6 材积 0.772 0** 0.951 9** 0.695 9* 0.712 7** 1 −0.091 7 0.367 3 树干圆满度 0.187 6 −0.185 9 −0.181 5 −0.080 3 −0.017 1 1 −0.762 1 树冠圆满度 −0.360 3 0.232 5 0.489 0 0.223 1 0.049 5 −0.460 5 1 说明:对角线下为表型相关系数,对角线上为遗传相关系数;*. P<0.05;**. P<0.01。
下载: 导出CSV
表 4 香椿种源各性状主成分分析
Table 4. Principal component analysis of different traits for provenances of T. sinensis
主成分 树高 胸径 冠幅 枝下高 材积 树干圆满度 树冠圆满度 特征值 贡献率/% 累积贡献率/% 主成分1 0.390 8 0.474 7 0.428 0 0.427 1 0.482 2 −0.078 3 0.133 0 3.822 8 54.611 54.611 主成分2 0.433 5 −0.059 2 −0.219 3 0.073 7 0.108 3 0.555 8 −0.659 1 1.791 5 25.593 80.204 主成分3 0.221 5 0.013 6 −0.343 0 0.095 8 0.008 9 −0.809 0 −0.411 5 0.635 9 9.084 89.287 主成分4 0.181 8 −0.432 6 −0.138 8 0.755 5 −0.335 2 0.047 7 0.274 2 0.477 3 6.820 96.106 主成分5 0.349 3 −0.372 8 0.722 1 −0.232 0 −0.325 1 −0.153 4 −0.185 7 0.247 0 3.529 99.635 主成分6 0.154 7 −0.630 2 −0.105 5 −0.194 5 0.686 5 −0.058 5 0.235 1 0.021 0 0.299 99.934 主成分7 0.656 8 0.218 5 −0.315 0 −0.374 9 −0.257 4 0.034 8 0.462 2 0.004 6 0.066 100.000
下载: 导出CSV
表 5 香椿种源综合评价得分和排名表
Table 5. Comprehensive evaluation scores and ranking of provenances
种源 综合评价得分 排名 种源 综合评价得分 排名 种源 综合评价得分 排名 阜阳 7.496 1 九江 6.732 9 南京 6.166 17 恩施 7.253 2 武汉 6.721 10 聊城 6.166 18 凭祥 6.977 3 运城 6.566 11 安康 6.087 19 南阳 6.913 4 晴隆 6.423 12 天水 6.008 20 元谋 6.883 5 杭州 6.372 13 三门峡 5.729 21 湘西 6.844 6 乳源 6.355 14 潍坊 5.418 22 新宁 6.827 7 峨眉山 6.351 15 邯郸 5.350 23 郴州 6.752 8 伏牛山 6.348 16 新乡 4.376 24
下载: 导出CSV
表 6 香椿3个优良种源各性状均值和遗传增益
Table 6. Means and genetic gain of various traits of the top 3 provenances of T. sinensis
种源 树高/m 胸径/cm 材积/m3 冠幅/m 枝下高/m 树干圆满度 树冠圆满度 元谋 8.4 10.61 0.039 8 3.25 3.92 0.87 0.71 恩施 9.4 9.95 0.042 2 3.51 4.43 0.88 0.72 阜阳 10.9 9.43 0.042 1 3.10 4.19 0.85 0.65 总体均值 8.86 8.07 0.0260 2.97 3.89 0.80 0.62 入选种源平均值 9.57 10.00 0.041 0 3.29 4.18 0.87 0.69 遗传增益/% 5.21 16.50 32.88 3.56 3.13 6.65 4.74
下载: 导出CSV
-
[1] 周翔宇. 中国香椿属的研究[D]. 南京: 南京林业大学, 2005. ZHOU Xiangyu. Study on Toona in China [D]. Nanjing: Nanjing Forestry University, 2005. [2] 彭方仁, 梁有旺. 香椿的生物学特性及开发利用前景[J]. 林业科技开发, 2005, 19(3): 3 − 6. PENG Fangren, LIANG Youwang. Biological characteristics and development and utilization prospects of Toona sinensis [J]. China Forestry Science and Technology, 2005, 19(3): 3 − 6. [3] 陈锡沐, 梁宝汉, 李秉滔. 广东楝科植物分类的初步研究[J]. 武汉植物学研究, 1986, 4(2): 167 − 194. CHEN Ximu, LIAGN Baohan, LI Bingtao. A preliminary taxonomic study on Meliaceae in Guangdong [J]. Journal of Wuhan Botanical Research, 1986, 4(2): 167 − 194. [4] 陆长旬, 张德纯, 王德槟. 香椿起源和分类地位的研究[J]. 植物研究, 2001, 21(2): 195 − 199. LU Changxun, ZHANG Dechun, WANG Debin. Origin and taxonomic position of Chinese Toon [Toona sinensis (A. Juss. ) Roem. ] [J]. Bulletin of Botanical Research, 2001, 21(2): 195 − 199. [5] 王希英, 王海宏, 乔勇进. 香椿优质丰产栽培关键技术[J]. 防护林科技, 2008(5): 133 − 134. WANG Xiying, WANG Haihong, QIAO Yongjin. Key techniques of high quality and high yield cultivation of Toona sinensis [J]. Protection Forest Science and Technology, 2008(5): 133 − 134. [6] MROGINSKI E, REY H Y, MROGINSKI L A. In vitro plantlet regeneration from Australian Red Cedar (Toona ciliata, Meliaceae) [J]. New Forests, 2003, 25(3): 177 − 184. [7] 杨玉珍, 彭方仁, 李洪岩. 不同种源香椿芽菜营养成分的变化研究[J]. 河南农业科学, 2007(4): 83 − 85. YANG Yuzhen, PENG Fangren, LI Hongyan. Changes of the nutritive compositions of Toona sinenesis bud in different provenances [J]. Journal of Henan Agricultural Sciences, 2007(4): 83 − 85. [8] 葛多云, 邹盛勤. 香椿叶中氨基酸和营养元素分析[J]. 微量元素与健康研究, 2005, 22(6): 23 − 24. GE Duoyun, ZOU Shengqin. Determination of amino acids and nutrient elements in Toona sinensis Roem. leaves [J]. Studies of Trace Elements and Health, 2005, 22(6): 23 − 24. [9] WANG Peihwei, TSAI M J, HSU C Y, et al. Toona sinensis Roem (Meliaceae) leaf extract alleviates hyperglycemia via altering adipose glucose transporter 4 [J]. Food and Chemical Toxicology, 2008, 46(7): 2554 − 2560. [10] 孙鸿有, 王鹏飞, 方炳法, 等. 香椿地理变异与种源选择[J]. 浙江林学院学报, 1992, 9(3): 237 − 245. SUN Hongyou, WANG Pengfei, FANG Bingfa, et al. Geographic variation and provenance selection of Chinese mahogany [J]. Journal of Zhejiang Forestry College, 1992, 9(3): 237 − 245. [11] 李淑玲, 桑玉强, 王平, 等. 不同种源香椿性状遗传分析[J]. 河南农业大学学报, 2000, 34(4): 363 − 366. LI Shuling, SANG Yuqiang, WANG Ping, et al. The genetic analysis on different species of Toona sinensis [J]. Journal of Henan Agricultural University, 2000, 34(4): 363 − 366. [12] 梁有旺, 彭方仁, 陈德平. 不同种源香椿苗期生长差异比较[J]. 林业科技开发, 2007, 21(2): 38 − 41. LIANG Youwang, PENG Fangren, CHEN Deping. The variations of seedling growth among provenances in Toona sinensis [J]. China Forestry Science and Technology, 2007, 21(2): 38 − 41. [13] 雷小华, 涂炳坤, 王茂丽, 等. 主成分分析在香椿性状评价和优良单株选择中的运用[J]. 华中农业大学学报, 2006, 25(4): 441 − 444. LEI Xiaohua, TU Bingkun, WANG Maoli, et al. Character evaluation and superior selection of Toona sinensis by principal component analysis [J]. Journal of Huazhong Agricultural University, 2006, 25(4): 441 − 444. [14] 崔宏安, 陈铁山, 范龙霞, 等. 陕西省种源香椿天然类型叶的结构与抗逆性研究[J]. 西北林学院学报, 2008, 23(2): 39 − 41. CUI Hongan, CHEN Tieshan, FAN Longxia, et al. Natural types of leaf structure and resistance of Toona sinensis Shaanxi Provenance [J]. Journal of Northwest Forestry University, 2008, 23(2): 39 − 41. [15] 刘军, 陈益泰, 姜景民, 等. 香椿种源苗期性状变异与原产地生态因子典型相关分析[J]. 东北林业大学学报, 2010, 38(11): 27 − 29. LIU Jun, CHEN Yitai, JIANG Jingmin, et al. Canonical correlation analysis between trait variation of Toona sinensis seedlings from different provenances and ecological factors in their original region [J]. Journal of Northeast Forestry University, 2010, 38(11): 27 − 29. [16] 徐成立, 张景兰, 陈东来. 树冠圆满度对树木生长的影响及作用研究[J]. 河北农业大学学报, 2005, 28(3): 45 − 48. XU Chengli, ZHANG Jinglan, CHEN Donglai. Study on the affect of the crown-fullness ratio on the growth of the tree and the form of the tree-trunk [J]. Journal of Agricultural Universtiy of Hebei, 2005, 28(3): 45 − 48. [17] 刘青华, 金国庆, 张蕊, 等. 24 年生马尾松生长、形质和木材基本密度的种源变异与种源区划[J]. 林业科学, 2009, 45(10): 55 − 61. LIU Qinghua, JIN Guoqing, ZHANG Rui, et al. Provenance variation in growth, stem-form and wood density of masson pine at 24-year-old and the provenance division [J]. Scientia Silvae Sinicae, 2009, 45(10): 55 − 61. [18] 李光友, 徐建民, 陆钊华, 等. 尾叶桉二代测定林家系的综合评选[J]. 林业科学研究, 2005, 18(1): 57 − 61. LI Guangyou, XU Jianmin, LU Zhaohua, et al. Studies on index selections of Eucalyptus urophylla families [J]. Forest Research, 2005, 18(1): 57 − 61. [19] 胡兴峰, 吴帆, 孙晓波, 等. 38年生马尾松种源生长及材性联合分析[J]. 南京林业大学学报(自然科学版), 2022, 46(3): 203 − 212. HU Xingfeng, WU Fan, SUN Xiaobo, et al. Joint analysis of growth and wood property of 38-year-old Pinus massoniana from 55 provenances [J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2022, 46(3): 203 − 212. [20] 张毅, 敖妍, 刘觉非, 等. 不同分布区文冠果的生长性状差异及其与地理-气候因子的相关性分析[J]. 植物资源与环境学报, 2019, 28(3): 44 − 50, 57. ZHANG Yi, AO Yan, LIU Juefei, et al. Differences in growth characters of Xanthoceras sorbifolium from different distribution areas and analysis on its correlation with geographical-climatic factors [J]. Journal of Plant Resources and Environment, 2019, 28(3): 44 − 50, 57. [21] WU Hanbin, DUAN Aiguan, ZHANG Jianguo. Long-term growth variation and selection of geographical provenances of Cunninghamia lanceolata (Lamb. ) Hook [J/OL]. Forests, 2019, 10: 876[2023-08-21]. doi:10.3390/f10100876. [22] SILVESTRO R, BRASSEUR S, KLISZ M, et al. Bioclimatic distance and performance of apical shoot extension: disentangling the role of growth rate and duration in ecotypic differentiation [J/OL]. Forest Ecology and Management, 2020, 477: 118483[2023-08-20]. doi: 10.1016/j. foreco.2020.118483. [23] MADSEN C L, KJARE D, RABILD A. Climatic criteria for successful introduction of Quercus species identified by use of Arboretum data [J]. Forestry:An International Journal of Forest Research, 2021, 94(4): 526 − 537. [24] 杨传平, 杨书文, 夏德安, 等. 兴安落叶松种源试验研究 (Ⅲ)种源区划[J]. 东北林业大学学报, 1991, 19(增刊 2): 77 − 83. YANG Chuanping, YANG Shuwen, XIA De’an, et al. Study on the provenance test of Larix gmelinii (Ⅲ) the provenance division [J]. Journal of Northeast Forest University, 1991, 19(suppl 2): 77 − 83. [25] 王军辉, 顾万春, 李斌, 等. 桤木优良种源/家系的选择研究——生长的适应性和遗传稳定性分析[J]. 林业科学, 2000, 36(3): 59 − 66. WANG Junhui, GU Wanchun, LI Bin, et al. Study on selection of Alnus cremastogyne provenance/family-analysis of growth adaptation and genetic stability [J]. Scientia Silvae Sinicae, 2000, 36(3): 59 − 66. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20230481
 点击查看大图
点击查看大图
计量
- 文章访问数: 1270
- HTML全文浏览量: 515
- PDF下载量: 35
- 被引次数: 0
