




-
菊花脑Chrysanthemum nankingense,又名菊花叶、路边黄等,是菊科Asteraceae草本植物[1]。原产于中国长江中下游流域,主要分布在华东地区,适应性较强,常见于沟渠边、田埂及林缘等湿润环境[2]。菊花脑富含次生代谢物,风味独特,药食兼用,是江苏特色保健蔬菜[3]。此外,二倍体菊花脑与多倍体观赏菊花Chrysanthemum×morifolium品种及药用菊花品种具有密切亲缘关系[4]。因此,菊花脑可作为模式植物,用于菊花遗传性状改良研究,尤其是在非生物胁迫耐受性领域,常用于筛选潜在的有益基因[5]。
海藻糖(trehalose)是一种非还原性双糖,在植物逆境响应中发挥重要作用。它能够调节气孔开度、维持细胞渗透。海藻糖通过海藻糖-6-磷酸磷酸酶(trehalose-6-phosphate phosphatase,TPP) 催化合成过程参与植物碳代谢调控及非生物胁迫响应[6]。海藻糖-6-磷酸磷酸酶在植物生长发育和逆境响应中发挥关键作用。海藻糖-6-磷酸磷酸酶基因通过催化海藻糖-6-磷酸(trehalose-6-phosphate,Tre6P) 脱磷酸生成海藻糖,不仅参与碳代谢调控,还能优化能量分配,从而增强植物在干旱、盐和低温等逆境条件下的耐受性[7]。在拟南芥Arabidopsis thaliana中,TPP 家族基因的表达受脱落酸(ABA) 及应激信号调控,利用AtTPPI 和 AtTPPF 的过表达可改善根系结构和气孔调控,提高水分利用率,从而增强抗旱性[8]。tppi 突变体对盐胁迫敏感,提示该基因参与耐盐相关通路[9]。DREB1A作为AP2/ERF家族转录因子,在干旱条件下诱导表达并作为转录激活因子结合 AtTPPF 启动子,从而提高其转录水平,调控海藻糖-6-磷酸水平及活性氧(ROS)积累以响应抗旱[10]。AtTPPD 表达依赖叶绿体定位,在盐胁迫下的表达有助于维持叶绿体功能[11]。此外,在拟南芥中TPP基因表达还受昼夜节律和环境压力调控,如 AtTPPA 在拟南芥的莲座叶中表现出显著昼夜节律,其转录本在黎明达到峰值,而 AtTPPH 在中午表达最高[12]。在农作物中,过表达 OsTPP1 的转基因玉米Zea mays在小穗发育阶段可提高籽粒结实率和收获指数,在田间试验中产量比对照组提高 9%~49%,严重干旱条件下产量可提高 31%~123%[13]。已有研究表明:在菊科菊属Chrysanthemum植物中,增加植株体内海藻糖含量或进行外源海藻糖处理,均可显著提高植株茎尖冷冻保存后的再生能力,并增强细胞对低温胁迫的耐受性[14],但对海藻糖-6-磷酸磷酸酶分子特征及逆境胁迫条件下的表达变化尚缺乏分析。
目前,关于 TPP 及 Tre6P 信号通路的研究主要集中在拟南芥、玉米等模式植物和农作物中,并已在碳代谢调控及非生物胁迫响应方面取得了一定进展。研究表明:TPP 在植物发育过程中同样具有重要调控功能。以玉米为例,TPP 基因家族成员RAMOSA3 (RA3)编码海藻糖-6-磷酸磷酸酶,在花序分生组织中表达,并通过调控转录因子 RAMOSA1 (RA1),影响糖信号向花序分生组织的输入,从而调控花序分枝形成[15]。此外,玉米中 TPP4 与 RA3 在花序分枝调控中具有功能冗余,且该作用不依赖其酶活性[16]。在干旱、盐和低温等非生物胁迫条件下,菊花脑的转录水平和生理响应显著改变。例如,低温和渗透胁迫处理可诱导菊花脑特定转录因子基因家族表达变化,并影响抗氧化酶活性和激素水平的积累。此外,利用 聚乙二醇(PEG)模拟干旱条件,揭示了菊花脑在水分胁迫下的生理调控特征[17]。尽管这些研究揭示了菊花脑对多种胁迫的响应机制,但针对具体候选基因的系统功能研究尚不充分。本研究以菊花脑为材料,利用与其有较高的序列保守性和同源关系的玉米RA3进行生物信息学分析,克隆得到同源基因 CnRAMOSA3 (CnRA3)。通过进一步分析该基因在不同组织及非生物胁迫条件下的表达模式,揭示TPP基因在菊花脑中的生物学功能及在非生物胁迫响应中的潜在作用,以期为后续功能研究提供理论基础。
-
本研究所用材料为菊花脑,取自南京农业大学中国菊花种质资源保存中心。植株在人工气候室条件下培养,光照/黑暗周期为16 h/8 h,培养温度为22 ℃,相对湿度保持在65%~70%。
-
分别采集长日照条件下(营养生长期)长势一致的18~20片叶龄的成熟植株的根、茎、叶(从上往下数第3片/第4片完全叶)、茎尖,及移入短日照(光照8 h、黑夜16 h)至现蕾(生殖生长期)状态后植株的根、茎、叶(从上往下数第3片完全叶)、花蕾。每个样品3个生物学重复,液氮速冻后于−80 ℃保存备用。
-
根据玉米中RAMOSA3 (RA3,蛋白序列号为ABD92779.1)序列,从菊花网站(https://cgd.njau.edu.cn/)获得菊花脑中同源基因CnRA3的序列(Cn0437030),并以此设计全长扩增引物(表1)。以菊花脑叶片cDNA为模板,按照Phusion高保真DNA聚合酶(TaKaRa)说明书的聚合酶链式反应(PCR)体系和程序进行全长扩增。通过琼脂糖凝胶电泳对PCR产物进行检测,将条带大小正确的胶块切下,并按SanPrep柱式DNA胶回收试剂盒(TaKaRa)方法纯化,连接至pEASY-Blunt Simple载体(北京全式金生物),随后通过热激法转化大肠埃希菌Escherichia coli DH5α;挑取单克隆阳性菌落检测后,将跑胶条带正确的送至生物公司进行测序。
表 1 本研究所用引物
Table 1. Primers used in this study
引物 序列(5′→3′) 用途 CnRA3-F ATGACTAACCCTAATGCAGTTG CnRA3全长扩增 CnRA3-R CTAATGATTGCGCGCAACTG 扩增CnRA3的cDNA全长 qCnRA3-F TGGTCGGATGGCGAAAAAT CnRA3定量正向引物 qCnRA3-R TCATTTTCGCGCGGCACTCAT CnRA3定量反向引物 CnActin-F AGCTTGCATATGTTGCTCTTGA CnRA3定量内参正向引物 CnActin-R TTACCGTAAAGGTCCTTCCTGA CnRA3定量内参反向引物 -
通过SOPMA (https://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL在线网站(https://swissmodel.expasy.org)对蛋白的二级结构、三级结构进行预测。使用美国国家生物技术信息中心(NCBI)数据库(https://www.ncbi.nlm.nih.gov/) Batch CD-Search工具预测蛋白保守结构域。利用Expasy在线网站(https://web.expasy.org/protparam)预测蛋白分子量、氨基酸种类和所占比例等基本理化性质。分别利用Expasy-ProtScale (https://web.expasy.org/protscale/)和TMHMM-2.0 (https://services.healthtech.dtu.dk/services/TMHMM-2.0/)以及SignalP-4.1 (https://services.healthtech.dtu.dk/services/SignalP-4.1/)在线网站预测蛋白的亲疏水性、跨膜区和信号肽。使用DNAMAN 10.0软件对不同物种氨基酸序列进行比对。利用MEGA 11.0软件构建系统发育树。利用菊花基因组公共数据库(https://cgd.njau.edu.cn/)和PlantCARE网站(https://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对CnRA3基因启动子序列进行查找和分析。
-
为探究CnRA3在菊花脑不同生长时期和不同组织中的表达情况,分别取有8~10片展开叶,处于营养生长时期和生殖生长时期的菊花脑的根、茎、叶片(从上至下第5片叶),营养生长时期的顶芽和生殖生长时期的花蕾。每个样本设3个生物学重复,液氮速冻并研磨后按照植物RNA提取试剂盒(SteadyPure Plant RNA Extraction Kit,艾科瑞)提取总RNA,经1.0%琼脂糖凝胶电泳检测合格后,利用Evo M-MLV反转录预混型试剂盒(艾科瑞)将其反转录为cDNA。cDNA保存于−20 ℃备用。
-
将生长状况良好的菊花脑无菌茎段于MS培养基上培养。人工气候培养室:光照/黑暗时长为16 h/8 h,温度为22 ℃,相对湿度为65%~70%。待菊花脑长至10片叶时,选取长势一致的植株随机分为3组(ABA组、NaCl组、PEG-
6000 组),每组3个生物学重复(每重复3株)。预培养7 d后进行胁迫处理:ABA组浇灌50 μmol·L−1 ABA溶液;NaCl组浇灌200 mmol·L−1 NaCl溶液;PEG组浇灌20%(w/v) PEG-6000 溶液,以胁迫处理开始为0 h,分别在0、3、6 h取样。取样部位为从上至下第3片叶,3个生物学重复,液氮速冻并手动研磨后按照植物RNA提取试剂盒(SteadyPure Plant RNA Extraction Kit,艾科瑞)提取总RNA,经1.0%琼脂糖凝胶电泳检测合格后,利用Evo M-MLV反转录预混型试剂盒(艾科瑞)将其反转录为cDNA。cDNA保存于-20 ℃,备用。 -
根据测序所得CnRA3全长序列设计荧光定量PCR (RT-qPCR)引物,以1.2.4、1.2.5节中反转录所得cDNA模板,进行PCR扩增以鉴定定量引物是否可用。使用Eppendorf Real Time PCR荧光定量仪,以CnActin[2]的扩增产物作为内参,设3个技术重复,分析CnRA3的相对表达量。结果分析采用2−△△CT的计算方法。
-
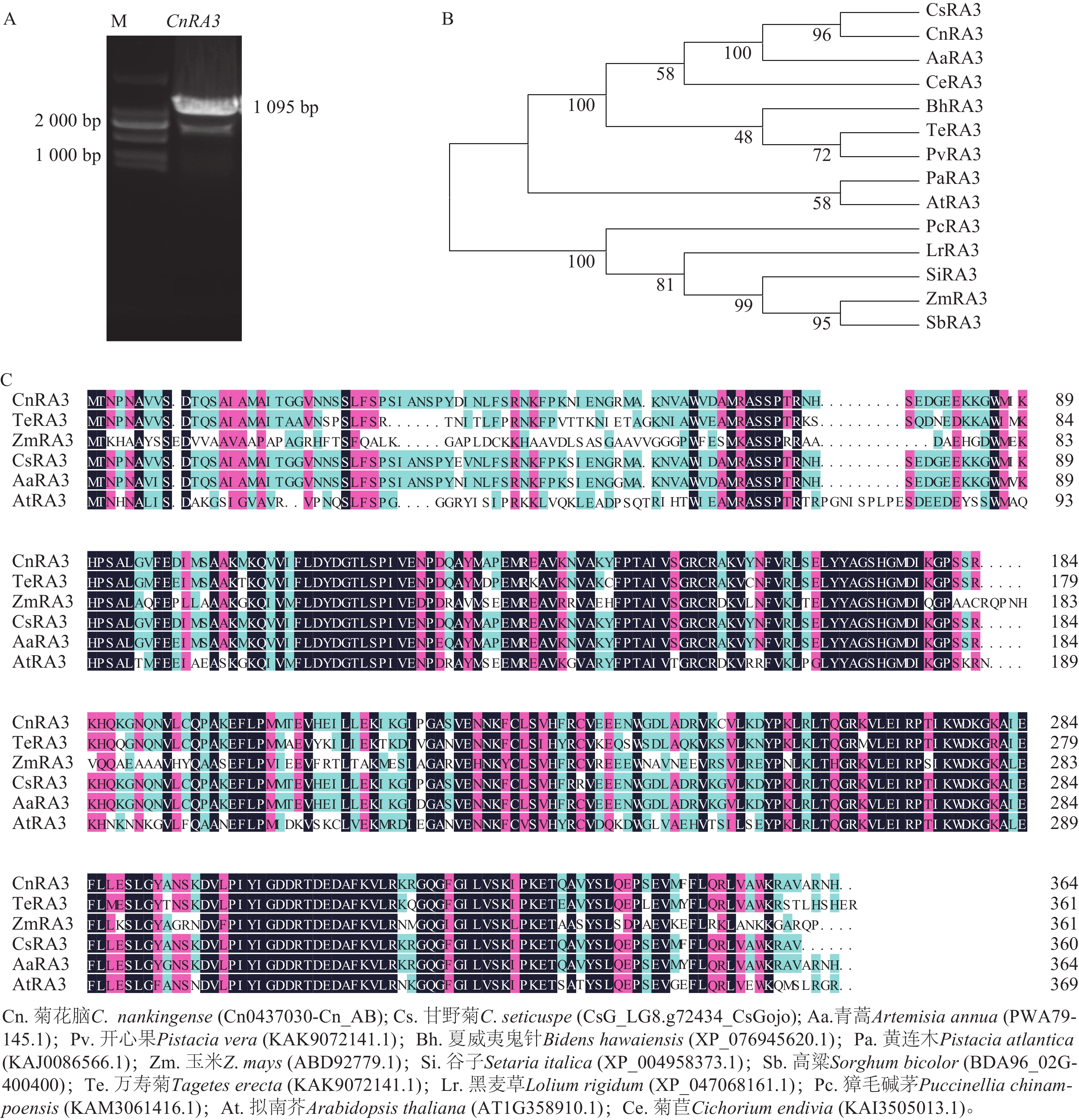
根据玉米中RA3 序列和菊花网站的序列信息,通过PCR扩增菊花脑同源基因Cn0437030,根据同源性,将其命名为CnRA3。CnRA3的完整开放阅读框(open reading frame,ORF)长度为1 095 bp (图1A),编码364个氨基酸。菊花脑CnRA3与其他13个物种的RA3蛋白进化分析结果(图1B)表明:CnRA3与甘野菊Chrysanthemum seticuspe的蛋白亲缘关系最近,其次是青蒿Artemisia annua,而与拟南芥Arabidopsis thaliana关系较远,暗示TPP结构域在菊科植物中进化较为保守。氨基酸序列比对结果显示(图1C):不同植物来源的 RA3 蛋白具有较高的氨基酸相似性。双子叶植物 CnRA3和 AaRA3 之间的相似度高达 98.6%,与 TeRA3 的相似度约为 90.4%。相比之下,RA3 蛋白与拟南芥 AtRA3 和玉米 ZmRA3 的相似度相对较低,分别为约 78.2% 和70.6%,表明 RA3 在进化过程中总体保守,但在单、双子叶植物间发生了一定程度的分化,表明CnRA3蛋白与其他物种的RA3蛋白具有较高的保守性。
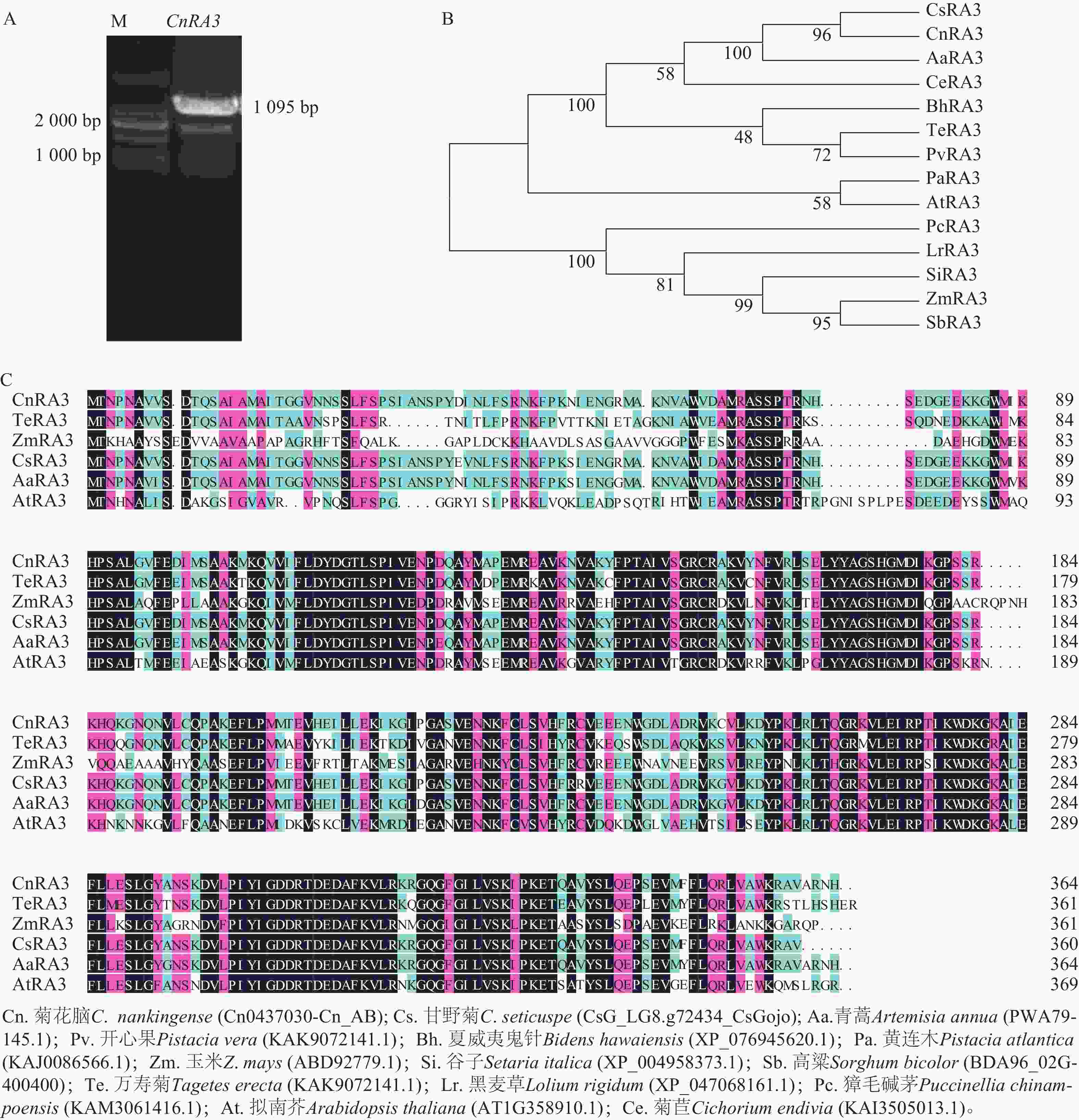
图 1 菊花脑CnRA3基因的克隆(A)与不同物种中RA3进化树构建(B)和同源性分析(C)
Figure 1. Cloning of CnRA3 gene from C. nankingense (A), phylogenetic tree construction of RA3 in different species (B), and homology analysis (C)
-
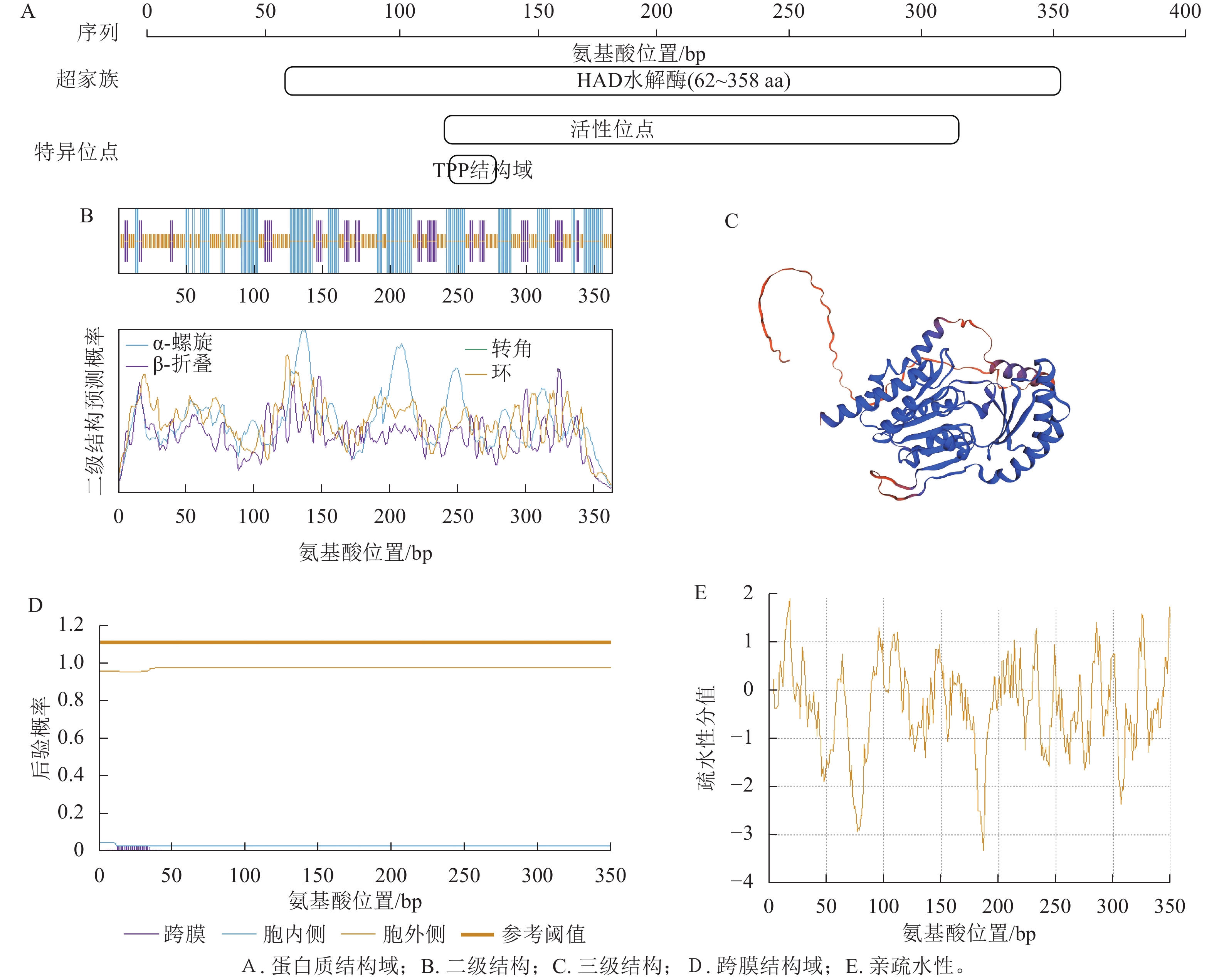
NCBI数据库Batch CD-Search工具预测结果显示:CnRA3蛋白含TPP结构域,属于HAD超家族成员(图2A)。运用SOPMA网站对CnRA3蛋白的二级结构进行预测(图2B),结果显示:该蛋白主要由无规则卷曲、延伸链和α-螺旋构成,其中无规则卷曲的占比最高,达48.35%,α-螺旋占35.44%,延伸链占比最低,为16.21%。SWISS-MODEL预测显示(图2C),TPP蛋白三级结构预测呈典型 α/β折叠构象,α-螺旋和 β-折叠主要集中于中部,形成稳定功能核心区。
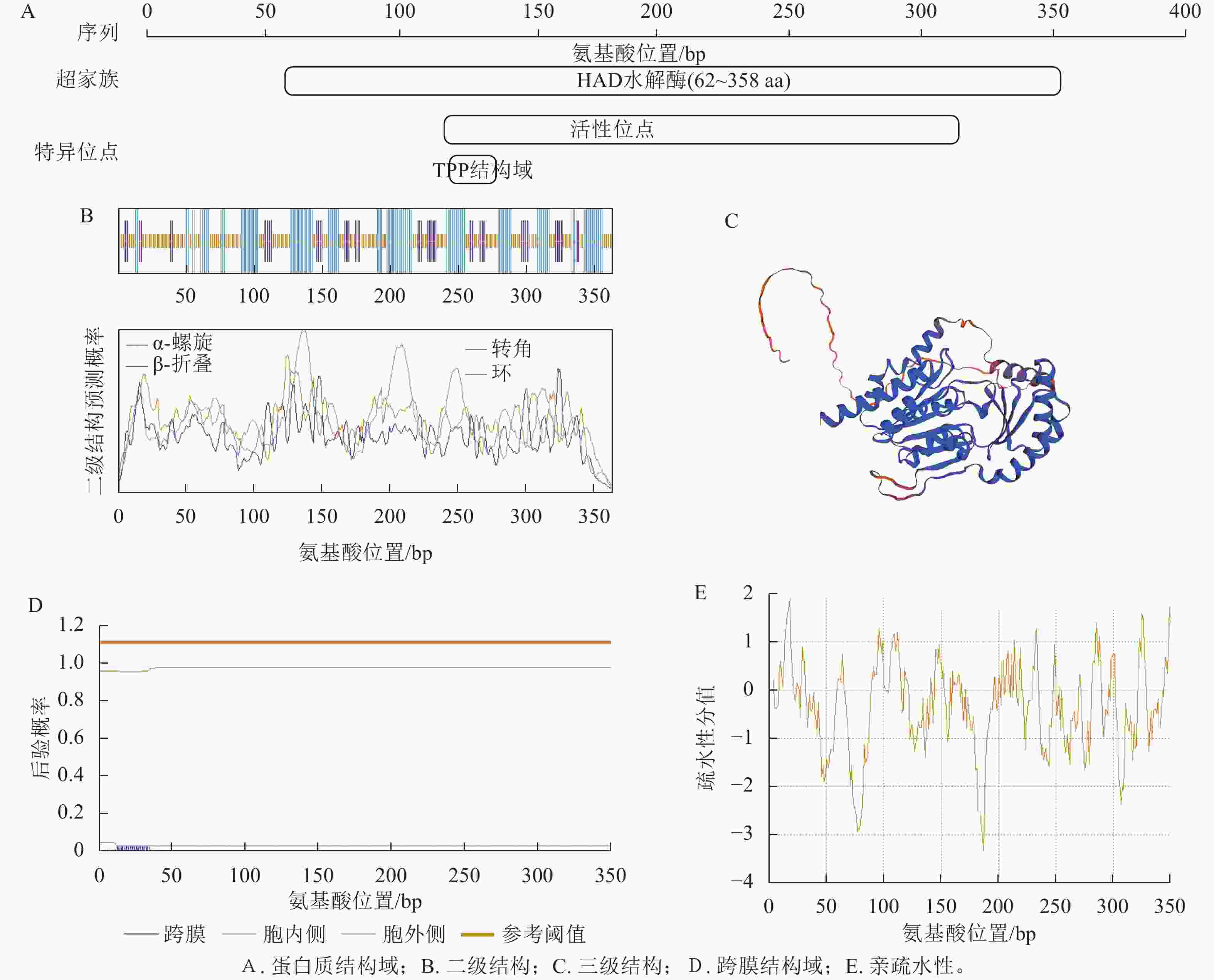
图 2 CnRA3蛋白序列分析预测结果
Figure 2. Bioinformation analysis and prediction results of CnRA3
分析CnRA3基本理化性质和结构可知:该蛋白的分子量约为40.04 kDa,理论等电点为9.14,为碱性蛋白。该蛋白共包含364个氨基酸,其中谷氨酸质量分数最高,为6.7%,带负电荷的天冬氨酸和谷氨酸分别有18和24个,带正电荷的精氨酸和赖氨酸分别有20和31个。不稳定系数、脂肪系数(AI)分别为41.43和81.43,预测为稳定蛋白。SignalP-4.1和TMHMM-2.0网站预测结果显示:CnRA3蛋白不存在信号肽和跨膜域(图2D)。平均亲水指数为−0.363, Expasy-ProtScale网站预测为亲水性蛋白(图2E)。
-
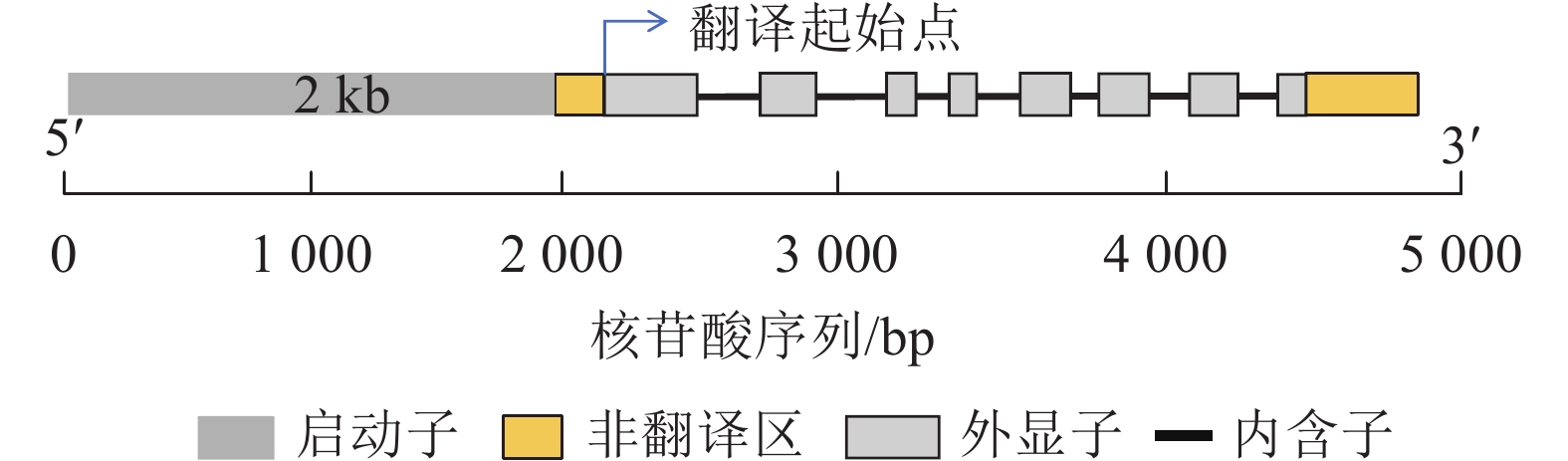
利用PlantCare网站进行CnRA3基因启动子的调控元件预测。对CnRA3基因的翻译起始位点上游2 kb序列分析发现(图3,表2):该基因含有厌氧诱导、生长素、赤霉素、脱落酸、茉莉酸甲酯、低温响应元件和典型的CAAT-box和TATA-box元件。上述分析结果表明:多个激素响应元件的共存提示 CnRA3 可能受不同信号途径的交互调控,因此,本研究从干旱和盐胁迫2个方面,结合ABA诱导响应实验对其在非生物胁迫响应中的抗逆功能进行了实验分析,并根据已有相关文献,对其在低温胁迫下的潜在作用进行了探讨。
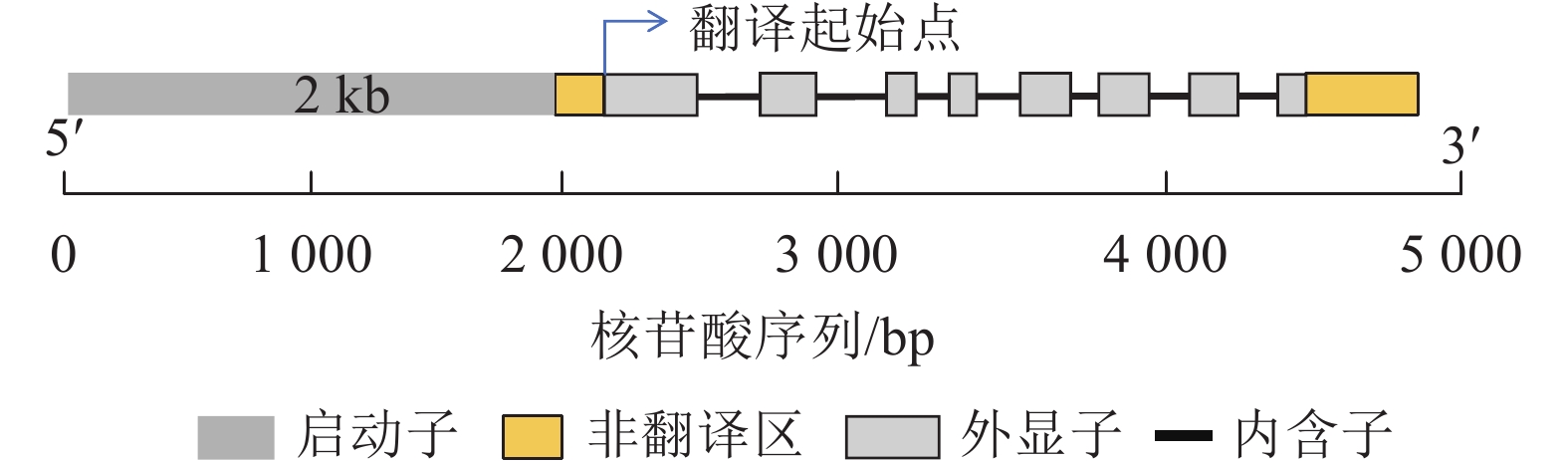
图 3 CnRA3基因序列结构图
Figure 3. Gene structure of CnRA3
表 2 CnRA3基因启动子所含胁迫与响应元件分析
Table 2. Analysis of stress-responsive elements contained in the promoter of the CnRA3 gene
顺式作用元件 基序 序列 位置 链 功能 G-Box CACGTT 1 585 6 − 参与光响应的顺式作用调控元件 G-box CACGTC 43 6 − G-box TACGTG 1 883 6 + ARE AAACCA 110 6 + 厌氧诱导必需的顺式作用调控元件 ARE AAACCA 1 228 6 − ARE AAACCA 1 453 6 − ABRE ACGTG 44 5 + 参与脱落酸响应的顺式作用元件 ABRE ACGTG 1 586 5 + ABRE ACGTG 1 884 5 + TGACG-motif TGACG 1 111 5 + 参与茉莉酸响应的顺式作用调控元件 TGACG-motif TGACG 1 325 5 + TGA-element AACGAC 440 6 + 生长素响应元件 TGA-element AACGAC 1 342 6 + 3-AF3 binding site CACTATCTAAC 1 388 10 + 保守DNA模块阵列(CMA3)的一部分 TC-rich repeats GTTTTCTTAC 82 9 − 参与防御与胁迫响应的顺式作用元件 -
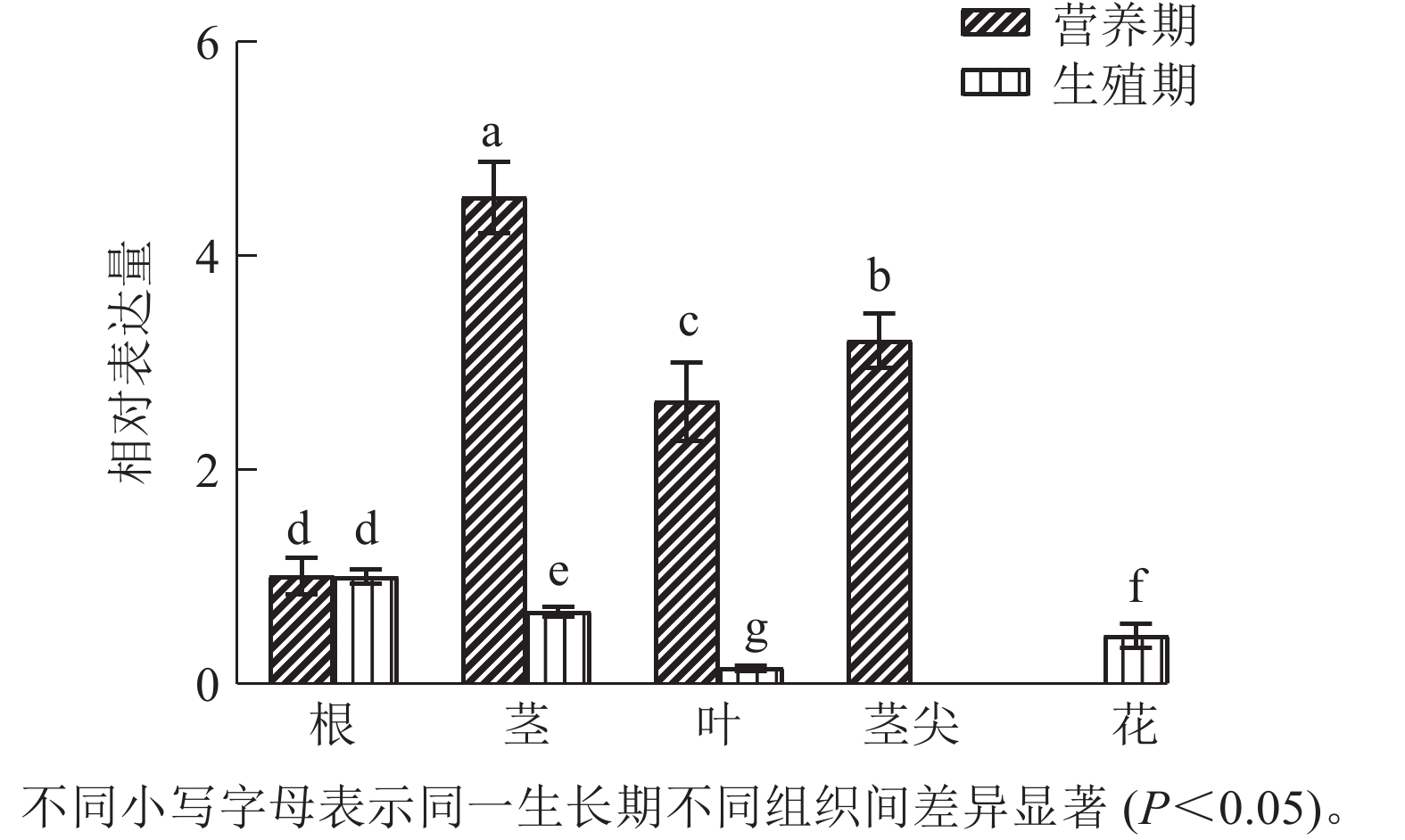
运用RT-qPCR分析CnRA3在菊花脑不同组织中的表达差异(图4)。结果发现:在营养生长时期,CnRA3在不同组织中相对表达量由高到低依次为茎、茎尖、叶、根,不同组织间表达差异达到显著水平(P<0.05);在生殖生长时期,不同组织中相对表达量由高到低依次为根、茎、花、叶,且不同组织间表达差异也达到显著水平(P<0.05)。同时,CnRA3 表达存在生长阶段差异。在营养生长期,CnRA3 在茎和茎尖的相对表达量最高,其次为叶和根,提示该基因可能在茎的生长调控中发挥作用。而在生殖生长期,根的相对表达量最高,其次为茎、花和叶,表明 CnRA3 在生殖阶段可能更多参与根部的生理活动及茎的发育。
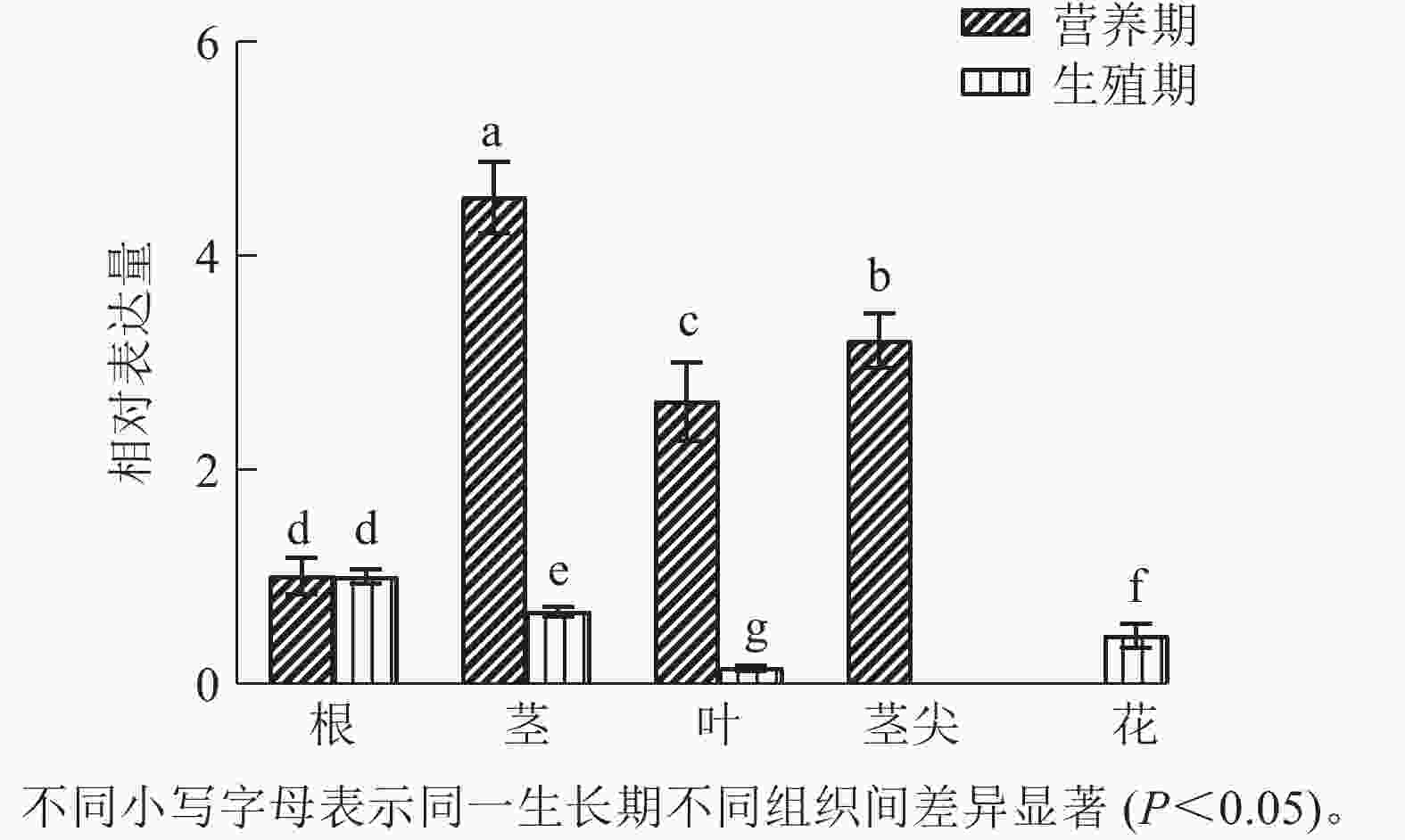
图 4 CnRA3基因在不同组织的表达量变化
Figure 4. Expression changes of CnRA3 in different tissues
-
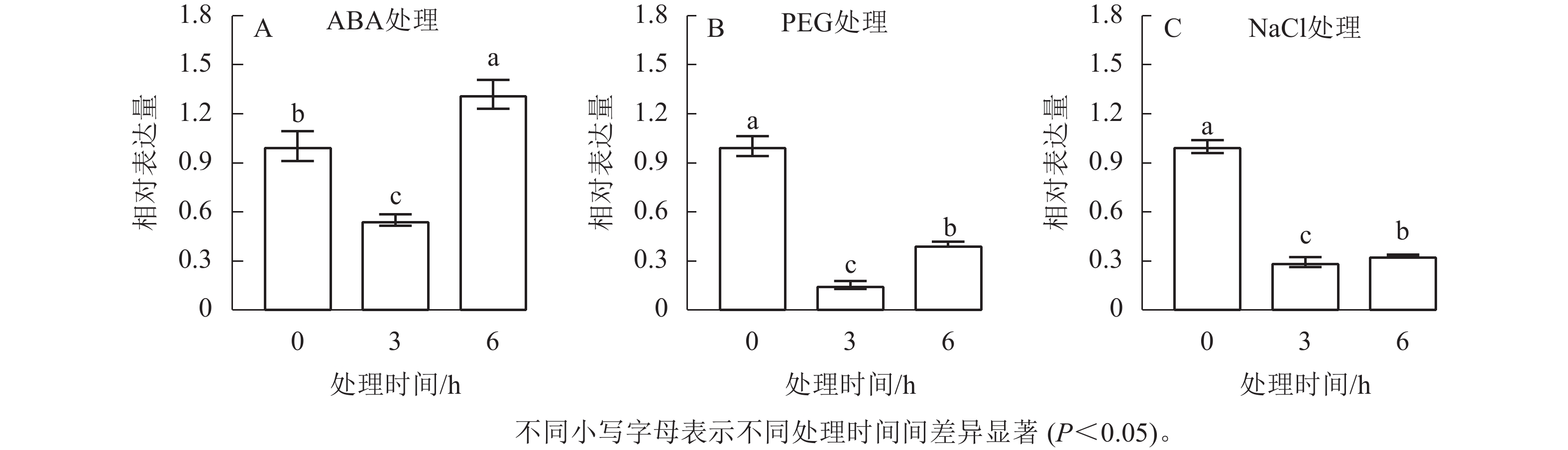
启动子序列的顺式作用元件分析结果显示:在CnRA3基因起始密码子上游2.0 kb区域内,存在对干旱、高渗环境及ABA应答相关的元件。因此,本研究进一步探究了CnRA3基因在模拟干旱、NaCl溶液处理、外源ABA处理条件下的表达模式。由图5可知:在不同处理下,CnRA3表达水平均呈现出随时间的延长呈现先下降后上升的趋势。其中在 ABA 处理下,CnRA3的表达水平随处理时间发生显著变化(图5A),其中 3 h 时相对表达量显著降低,约为处理前的 0.55 倍,而在处理 6 h 后其表达水平显著回升,达到处理前的 1.32 倍,差异达到显著水平(P<0.05),表明CnRA3对 ABA 信号具有阶段性响应特征。进一步分析 PEG 和 NaCl 处理下的表达情况(图5B、C)发现:两者均在 3 h 时显著抑制CnRA3的表达(P<0.05),其中 PEG 和 NaCl 处理后的相对表达量分别降至约 0.29 和 0.15倍,显著低于 0 h 水平(P<0.05);随着处理时间延长至 6 h,CnRA3表达量均出现一定程度回升,但仍低于初始水平。上述结果表明:CnRA3基因对干旱胁迫和NaCl胁迫均表现出明显的抑制响应,而其在 ABA 处理下的动态变化暗示该基因可能通过 ABA 相关信号调控途径参与菊花脑对非生物胁迫的响应过程。
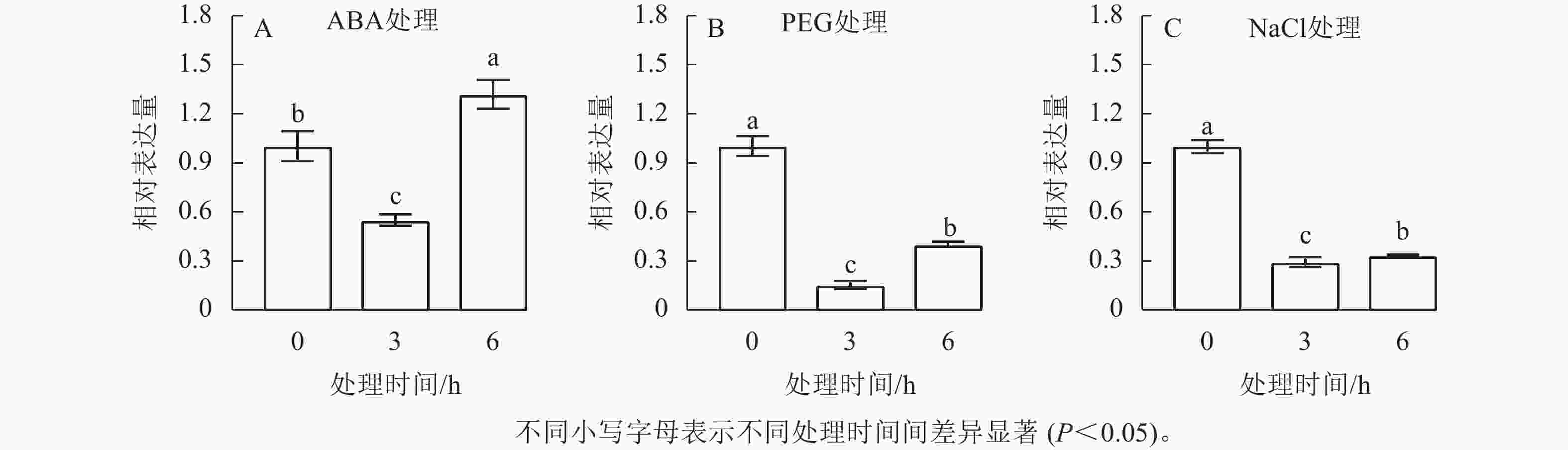
图 5 CnRA3基因在外源ABA(A)、PEG(B)和NaCl处理(C)下的表达变化
Figure 5. Relative expression level of the CnRA3 gene under ABA treatment (A), PEG treatment, and NaCl treatment (B)
-
海藻糖代谢及其介导的糖信号在植物生长发育和非生物胁迫响应中发挥重要调控作用[18]。其中,T6P被认为是反映植物体内碳状态的重要信号分子,而 TPP通过催化 Tre6P 去磷酸化,在维持 Tre6P 稳态及调控碳代谢平衡中处于关键地位[19]。在拟南芥等模式植物中,TPP 基因家族成员可整合糖信号、ABA 信号及能量感知通路,参与植物对干旱、低温和渗透胁迫的适应过程。在拟南芥中,部分 AtTPP 基因在 ABA 处理及逆境条件下被显著诱导,其功能被认为与SnRK1信号通路介导的能量重分配和生长抑制反应密切相关[20]。这类研究揭示了 TPP 在调控中的重要作用,也为非模式植物中 TPP 基因功能研究提供了参考。
菊花作为重要的观赏和经济作物,其生长发育及观赏品质极易受到干旱、低温和盐胁迫等环境因素的影响[21]。然而,目前关于菊花TPP基因的功能研究仍是空白。本研究成功克隆并鉴定了菊花脑 CnRA3 基因,其编码蛋白含有典型TPP结构域,不含跨膜区。系统进化分析表明,CnRA3 基因与菊科RA3 蛋白具有较高同源性,提示该基因在进化过程中具有一定的保守性。蛋白结构分析结果进一步支持 CnRA3 具备典型 TPP 蛋白的结构特征,为其发挥酶学功能提供了结构基础。
启动子分析显示:CnRA3 上游区域含有多种与激素和逆境响应相关的顺式作用元件。其中,ABRE 元件与 ABA 信号调控密切相关,与 CnRA3 在外源 ABA 处理下的表达变化趋势相一致。ARE 元件通常参与渗透胁迫和逆境响应过程,可能与该基因在 PEG 模拟干旱和盐胁迫处理下的表达变化有关。TC-rich repeats 元件与防御和逆境响应相关,CnRA3 的转录调控可能受多种胁迫信号共同影响。REN等[22]研究表明:CnRA3的转录本Unigene70569_S2_3在低温胁迫条件下显著上调,提示CnRA3可能参与对低温胁迫的调控过程,从而进一步支持CnRA3在非生物胁迫响应中的潜在功能。在拟南芥中,TPP基因家族成员也被报道参与低温胁迫响应,其表达受到低温诱导,提高植物的低温适应能力[23]。组织定量表达分析表明:CnRA3在不同组织中的相对表达量呈显著差异。营养生长期以茎和茎尖表达量较高,而生殖生长期则以根表达量最高,其次为茎、花和叶,提示CnRA3可能在不同生长阶段的特定组织中发挥作用。拟南芥TPP家族成员在不同组织和发育阶段中呈现出明显的组织特异性表达,其中有10个TPP基因分别在分生组织、根系及花器官中具有较高表达水平,在顶端分生组织维持、根系结构调控以及花器官分化等发育过程中发挥重要作用,从而影响拟南芥的整体生长和形态建成[24]。CnRA3在茎尖、根和花等组织中的差异表达,推测该基因可能在菊花脑不同发育阶段的器官发育和生长调控中发挥重要作用。表达分析结果表明:CnRA3 在 ABA 处理下呈现阶段性表达变化,在 PEG 模拟干旱和盐胁迫条件下早期表达受到抑制,随后出现一定程度的恢复,且在根和叶中表达水平相对较高。该表达特征在响应趋势上与拟南芥 AtTPPF 在 ABA 信号及非生物胁迫条件下的表达调控模式具有一定相似性[25]。在拟南芥中,TPP基因在干旱胁迫条件下被诱导表达,并与植株抗旱性增强相关[26]。这些基因通过调节气孔活动、根系生长及组织适应性反应,帮助植物更好地应对逆境胁迫。结合本研究的组织定量分析结果,推测CnRA3可能在不同生长阶段的特定组织中发挥作用。本研究重点分析了CnRA3在干旱和盐胁迫下的抗逆功能,并结合发表的转录组数据,对其在低温胁迫中的潜在作用进行了初步推测。
-
本研究结果表明:CnRA3蛋白具有典型 TPP 结构特征,在菊科植物中具有较高的进化保守性,其启动子区域富含 ABA 和逆境响应相关顺式作用元件。
表达分析显示:CnRA3 对 ABA、PEG 模拟干旱及盐胁迫具有明显响应特征,且在根和叶等组织中表达较高,提示其可能参与 ABA 介导的非生物胁迫响应过程。CnRA3 在不同生长阶段的特定组织中呈现差异表达,其组织特异性表达特征可能通过调控关键组织的生理响应,从而在干旱、盐胁迫等逆境条件下增强菊花脑的适应能力,并可能在低温胁迫响应中发挥潜在作用。
尽管本研究从结构特征、启动子元件及表达模式等方面对 CnRA3 进行了初步解析,但其具体生物学功能仍有待通过遗传转化或基因编辑等手段进一步验证。未来可结合低温胁迫处理及功能验证实验,进一步明确CnRA3在低温响应中的调控作用。本研究为深入揭示 TPP 介导的糖信号在菊花逆境响应中的作用机制提供了重要基础,也为菊花抗逆性分子育种提供了潜在候选基因。
Cloning and expression analysis of the CnRA3 gene in Chrysanthemum nankingense
-
摘要:
目的 克隆菊花脑Chrysanthemum nankingense海藻糖-6-磷酸磷酸酶编码基因CnRAMOSA3 (CnRA3),并对其进行生物信息学分析和表达模式分析,鉴定菊花脑RAMOSA3同源基因的结构特征及在非生物胁迫响应中的潜在功能。 方法 以菊花脑为实验材料,依据其转录组数据库信息,克隆获得CnRA3基因全长编码序列,采用生物信息学方法分析其系统进化关系、蛋白结构特征及启动子顺式作用元件,并利用实时荧光定量聚合酶链式反应(RT-qPCR)分析该基因在不同组织中的表达模式,及在外源脱落酸(ABA)、聚乙二醇(PEG)模拟干旱和盐胁迫处理下的表达变化。 结果 CnRA3开放阅读框(ORF)长度为 1095 bp,编码364个氨基酸。编码蛋白含有典型的海藻糖-6-磷酸磷酸酶(TPP)结构域,不含跨膜结构,整体呈亲水性特征。系统进化分析表明:CnRA3在菊科Asteraceae植物中进化较为保守,其启动子区域富含ABA及逆境响应相关顺式作用元件。RT-qPCR分析显示:CnRA3 在菊花脑营养生长时期和生殖生长时期的各组织中均有表达,但其表达水平具有明显的组织特异性和发育阶段差异。在营养生长时期,CnRA3 主要在茎和茎尖中表达;在生殖生长时期,主要在根和茎部位表达。在ABA、PEG和盐胁迫处理下,CnRA3相对表达量显著变化(P<0.05),表达水平均呈现出随时间的延长先下降后上升的趋势。结论 CnRA3 可能在干旱胁迫和盐胁迫条件下表现出抑制性反应,并参与 ABA 介导的非生物胁迫响应过程。图5表2参26 -
关键词:
- 菊花脑 /
- CnRAMOSA3 (CnRA3) /
- 生物信息学分析 /
- 非生物胁迫 /
- 基因表达
Abstract:Objective The trehalose-6-phosphate phosphatase encoding gene CnRAMOSA3 (CnRA3) was cloned from Chrysanthemum nankingense, and its bioinformatics characteristics and expression patterns were analyzed to characterize its structural features and explore its potential roles in abiotic stress responses. Method Using C. nankingense as the experimental material, the full-length coding sequence of the CnRA3 gene was cloned based on transcriptome database information. Bioinformatics analysis were conducted to investigate its phylogenetic relationships, protein structural characteristics, and cis-acting elements in the promoter region. In addition, real-time quantitative PCR (RT-qPCR) was employed to analyze the expression patterns of CnRA3 under exogenous abscisic acid (ABA), PEG-simulated drought, and salt stress treatments, as well as in different tissues. Result The open reading frame (ORF) of CnRA3 was 1095 bp in length, encoding 364 amino acids. The encoded protein contained a typical trehalose-6-phosphate phosphatase (TPP) domain, without transmembrane structure, and exhibited overall hydrophilic characteristics. Phylogenetic analysis showed that CnRA3 was conserved in the Asteraceae family, with its promoter enriched in ABA- and stress-responsive cis-acting elements. RT-qPCR analysis revealed that CnRA3 was expressed in multiple tissues with distinct tissue specificity and developmental stage differences. During the vegetative growth stage, CnRA3 was primarily expressed in stems and shoot tips, while during the reproductive growth stage, it was mainly expressed in roots and stems. Under ABA, PEG, and salt stress treatments, the expression level of CnRA3 changed significantly, exhibiting a time-dependent decrease followed by an increase.Conclusion CnRA3 may exhibit an inhibitory response under drought and salt stress conditions and participate in ABA-mediated abiotic stress responses. [Ch, 5 fig. 2 tab. 26 ref.] -
Key words:
- Chrysanthemum nankingense /
- CnRAMOSA3 (CnRA3) /
- bioinformatics analysis /
- abiotic /
- gene expression
-
图 1 菊花脑CnRA3基因的克隆(A)与不同物种中RA3进化树构建(B)和同源性分析(C)
Figure 1 Cloning of CnRA3 gene from C. nankingense (A), phylogenetic tree construction of RA3 in different species (B), and homology analysis (C)
图 5 CnRA3基因在外源ABA(A)、PEG(B)和NaCl处理(C)下的表达变化
Figure 5 Relative expression level of the CnRA3 gene under ABA treatment (A), PEG treatment, and NaCl treatment (B)
表 1 本研究所用引物
Table 1. Primers used in this study
引物 序列(5′→3′) 用途 CnRA3-F ATGACTAACCCTAATGCAGTTG CnRA3全长扩增 CnRA3-R CTAATGATTGCGCGCAACTG 扩增CnRA3的cDNA全长 qCnRA3-F TGGTCGGATGGCGAAAAAT CnRA3定量正向引物 qCnRA3-R TCATTTTCGCGCGGCACTCAT CnRA3定量反向引物 CnActin-F AGCTTGCATATGTTGCTCTTGA CnRA3定量内参正向引物 CnActin-R TTACCGTAAAGGTCCTTCCTGA CnRA3定量内参反向引物  下载: 导出CSV
下载: 导出CSV
表 2 CnRA3基因启动子所含胁迫与响应元件分析
Table 2. Analysis of stress-responsive elements contained in the promoter of the CnRA3 gene
顺式作用元件 基序 序列 位置 链 功能 G-Box CACGTT 1 585 6 − 参与光响应的顺式作用调控元件 G-box CACGTC 43 6 − G-box TACGTG 1 883 6 + ARE AAACCA 110 6 + 厌氧诱导必需的顺式作用调控元件 ARE AAACCA 1 228 6 − ARE AAACCA 1 453 6 − ABRE ACGTG 44 5 + 参与脱落酸响应的顺式作用元件 ABRE ACGTG 1 586 5 + ABRE ACGTG 1 884 5 + TGACG-motif TGACG 1 111 5 + 参与茉莉酸响应的顺式作用调控元件 TGACG-motif TGACG 1 325 5 + TGA-element AACGAC 440 6 + 生长素响应元件 TGA-element AACGAC 1 342 6 + 3-AF3 binding site CACTATCTAAC 1 388 10 + 保守DNA模块阵列(CMA3)的一部分 TC-rich repeats GTTTTCTTAC 82 9 − 参与防御与胁迫响应的顺式作用元件
下载: 导出CSV
-
[1] AI Penghui, XUE Jundong, SHI Zhongya, et al. Genome-wide characterization and expression analysis of MYB transcription factors in Chrysanthemum nankingense[J]. BMC Plant Biology, 2023, 23(1): 140. DOI: 10.1186/s12870-023-04137-7. [2] SONG Chi, LIU Yifei, SONG Aiping, et al. The Chrysanthemum nankingense genome provides insights into the evolution and diversification of Chrysanthemum flowers and medicinal traits[J]. Molecular Plant, 2018, 11(12): 1482−1491. DOI: 10.1016/j.molp.2018.10.003. [3] WANG Chengxiang, SU Yong, WEI Min, et al. Decoding flavonoid metabolism for nutritional enhancement: a transcriptome-metabolome integration study of biosynthesis in edible Chrysanthemum indicum L.[J]. Foods, 2025, 14(11): 1896. DOI: 10.3390/foods14111896. [4] HAO Dacheng, SONG Yanjun, XIAO Peigen, et al. The genus Chrysanthemum: phylogeny, biodiversity, phytometabolites, and chemodiversity[J]. Frontiers in Plant Science, 2022, 13: 973197. DOI: 10.3389/fpls.2022.973197. [5] DU Xinping, LI Wenyan, SHENG Liping, et al. Over-expression of Chrysanthemum CmDREB6 enhanced tolerance of Chrysanthemum to heat stress[J]. BMC Plant Biology, 2018, 18(1): 178. DOI: 10.1186/s12870-018-1400-8. [6] YANG Yan, YAO Yandong, LI Jing, et al. Trehalose alleviated salt stress in tomato by regulating ROS metabolism, photosynthesis, osmolyte synthesis, and trehalose metabolic pathways[J]. Frontiers in Plant Science, 2022, 13: 772948. DOI: 10.3389/fpls.2022.772948. [7] FICHTNER F, LUNN J E. The role of trehalose 6-phosphate (Tre6P) in plant metabolism and development[J]. Annual Review of Plant Biology, 2021, 72: 737−760. DOI: 10.1146/annurev-arplant-050718-095929. [8] WANG Wenjing, CHEN Qingbin, XU Shouming, et al. Trehalose-6-phosphate phosphatase E modulates ABA-controlled root growth and stomatal movement in Arabidopsis[J]. Journal of Integrative Plant Biology, 2020, 62(10): 1518−1534. DOI: 10.1111/jipb.12925. [9] KATAYA A R A, ELSHOBAKY A, HEIDARI B, et al. Multi-targeted trehalose-6-phosphate phosphatase Ⅰ harbors a novel peroxisomal targeting signal 1 and is essential for flowering and development[J]. Planta, 2020, 251(5): 98. DOI: 10.1007/s00425-020-03389-z. [10] LIN Qingfang, YANG Jiao, WANG Qiongli, et al. Overexpression of the trehalose-6-phosphate phosphatase family gene AtTPPF improves the drought tolerance of Arabidopsis thaliana[J]. BMC Plant Biology, 2019, 19(1): 381. DOI: 10.1186/s12870-019-1986-5. [11] KRASENSKY J, BROYART C, RABANAL F A, et al. The redox-sensitive chloroplast trehalose-6-phosphate phosphatase AtTPPD regulates salt stress tolerance[J]. Antioxidants & Redox Signaling, 2014, 21(9): 1289−1304. DOI: 10.1089/ars.2013.5693. [12] LIN Qingfang, WANG Song, DAO Yihang, et al. Arabidopsis thaliana trehalose-6-phosphate phosphatase gene TPPI enhances drought tolerance by regulating stomatal apertures[J]. Journal of Experimental Botany, 2020, 71(14): 4285−4297. DOI: 10.1093/jxb/eraa173. [13] NUCCIO M L, WU J, MOWERS R, et al. Expression of trehalose-6-phosphate phosphatase in maize ears improves yield in well-watered and drought conditions[J]. Nature Biotechnology, 2015, 33(8): 862−869. DOI: 10.1038/nbt.3277. [14] OSORIO-SAENZ A, MASCORRO-GALLARDO J O, GONZÁLEZ-ARNAO M T, et al. Evaluation of endogenous and exogenous effect of trehalose on regeneration of cryopreserved Chrysanthemum (Dendranthema grandiflorum Kitam.) shoot-tips[J]. Acta Horticulturae, 2014, 1039: 99−105. DOI: 10.17660/actahortic.2014.1039.11. [15] CLAEYS H, VI S L, XU Xiaosa, et al. Control of meristem determinacy by trehalose 6-phosphate phosphatases is uncoupled from enzymatic activity[J]. Nature Plants, 2019, 5(4): 352−357. DOI: 10.1038/s41477-019-0394-z. [16] SATOH-NAGASAWA N, NAGASAWA N, MALCOMBER S, et al. A trehalose metabolic enzyme controls inflorescence architecture in maize[J]. Nature, 2006, 441(7090): 227−230. DOI: 10.1038/nature04725. [17] ZHANG Yi, GU Jing, XIA Xiaolong, et al. Contrasting responses to drought stress between Chrysanthemum japonense and C. nankingense[J]. Ornamental Plant Research, 2022, 2(1): 1−11. DOI: 10.48130/opr-2022-0016. [18] PAUL M J, PRIMAVESI L F, JHURREEA D, et al. Trehalose metabolism and signaling[J]. Annual Review of Plant Biology, 2008, 59: 417−441. DOI: 10.1146/annurev.arplant.59.032607.092945. [19] KERBLER S M, ARMIJOS-JARAMILLO V, LUNN J E, et al. The trehalose 6-phosphate phosphatase family in plants[J]. Physiologia Plantarum, 2023, 175(6): e14096. DOI: 10.1111/ppl.14096. [20] TSAI A Y, GAZZARRINI S. Trehalose-6-phosphate and SnRK1 kinases in plant development and signaling: the emerging picture[J]. Frontiers in Plant Science, 2014, 5: 119. DOI: 10.3389/fpls.2014.00119. [21] GAO Wenjie, HE Miao, LIU Jie, et al. Overexpression of Chrysanthemum lavandulifolium ClCBF1 in Chrysanthemum morifolium ‘White Snow’ improves the level of salinity and drought tolerance[J]. Plant Physiology and Biochemistry, 2018, 124: 50−58. DOI: 10.1016/j.plaphy.2018.01.004. [22] REN Liping, SUN Jing, CHEN Sumei, et al. A transcriptomic analysis of Chrysanthemum nankingense provides insights into the basis of low temperature tolerance[J]. BMC Genomics, 2014, 15(1): 844. DOI: 10.1186/1471-2164-15-844. [23] LIN Qingfang, WANG Jianyong, GONG Jiaxin, et al. The Arabidopsis thaliana trehalose-6-phosphate phosphatase gene AtTPPI improve chilling tolerance through accumulating soluble sugar and JA[J]. Environmental and Experimental Botany, 2023, 205: 105117. DOI: 10.1016/j.envexpbot.2022.105117. [24] SCHLUEPMANN H, PELLNY T, van DIJKEN A, et al. Trehalose 6-phosphate is indispensable for carbohydrate utilization and growth in Arabidopsis thaliana[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(11): 6849−6854. DOI: 10.1073/pnas.1132018100. [25] MORALES-HERRERA S, JOURQUIN J, COPPÉ F, et al. Trehalose-6-phosphate signaling regulates lateral root formation in Arabidopsis thaliana[J]. Proceedings of the National Academy of Sciences of the United States of America, 2023, 120(40): e2302996120. DOI: 10.1073/pnas.2302996120. [26] DELATTE T L, SEDIJANI P, KONDOU Y, et al. Growth arrest by trehalose-6-phosphate: an astonishing case of primary metabolite control over growth by way of the SnRK1 signaling pathway[J]. Plant Physiology, 2011, 157(1): 160−174. DOI: 10.1104/pp.111.180422. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20260166
 点击查看大图
点击查看大图
计量
- 文章访问数: 151
- HTML全文浏览量: 55
- PDF下载量: 21
- 被引次数: 0
