










-
生物多样性是人类赖以生存的重要支撑,对城市的可持续发展具有重要意义[1]。鸟类是城市中最为常见且易于接近的动物,有利于促进城市居民身心健康[2]。但是,在城市化等剧烈人类活动干扰下,鸟类赖以生存的栖息地不断减少,鸟类生物多样性急剧下降,引起了国内外学者的广泛关注[3]。长江三角洲城市群是中国经济发展最快的地区,城市化水平高,鸟类栖息地破碎化严重,研究该地区鸟类栖息环境营建的理论和方法具有重要的实践应用价值。城市鸟类主要食物来自植物[4],园林植物的果实是城市鸟类植物性食物的主要组成部分[5],合理配置挂果树种对吸引更多鸟类具有重要意义。同时由于食物的可获得性直接影响鸟类对生境的选择[6],因此对鸟类取食与食源树种果实特性关系的分析是研究城市鸟类栖息环境构建的重要基础之一。目前涉及鸟类取食与果实特性关系的研究可归纳为果实所处植物空间、果实外特性和果实内特性3个方面:果实所处空间的高度特征对鸟类取食有明显影响[7],鸟类取食时其形态特征与果实生态特征存在一定对应性[8-9],鸟类对不同果实类型的取食(如肉质核果、梨果等)存在一定偏好 [10],同时受进化演替的影响,果实颜色、大小与鸟类取食存在相关性[11-13],鸟类更倾向于高糖分、低蛋白的果实,并且不同鸟类对果实脂肪含量的选择倾向存在差异[14-15]。长江三角洲地区进入秋冬季后鸟类食源整体减少,但鸟类数量却有所增加,这使得植物果实供给压力增大。因此,本研究以杭州市临安区绿地中常见的秋冬季鸟类食源树种为研究对象,分析果实特性中的果实所处植物空间、果实形态变化、果实内含物与鸟类取食之间的关系,为长江三角洲地区城市生物多样性规划和引鸟植物景观营建提供理论支撑。
-
本研究在浙江农林大学东湖校区内开展。该校区位于浙江省杭州市临安区,占地面积约200 hm2,属于中亚热带季风气候,全年降水量为1 628.6 mm,全年平均气温为16.4 ℃。校区以校园和植物园“两园合一”理念规划建设,校园内拥有湖泊、湿地、缓坡林地和草地,保存植物3 300种(含品种),吸引鸟类超过400种[16],生物多样性丰富,观测区块为城市近自然生境,具有足够代表性和典型性。
-
选择杭州地区园林绿化植物中常见的秋冬季鸟类食源树种10种(表1),所选研究对象以2~5株的规模分布于3处近自然生境中,且环境相似,人为主动干扰少(图1)。在秋冬季常见鸟类中,体型较大的有白头鹎Pycnonotus sinensis、领雀嘴鹎Spizixos semitorques、乌鸫Turdus merula、山斑鸠Streptopelia orientalis、黑脸噪鹛Garrulax perspicillatus等,体型较小的鸟类有丝光椋鸟Sturnus sericeus、燕雀Fringilla montifringilla、红胁蓝尾鸲Tarsiger cyanurus、北红尾鸲Phoenicurus auroreus、红头长尾山雀Aegithalos concinnus、大山雀Parus major、灰头鹀Emberiza spodocephala等。
表 1 10种鸟类食源树种概况
Table 1. Overview of 10 tree species as food source
序号 树种 科名 属名 株高/m 观赏特性 1 垂丝海棠 Malus halliana 蔷薇科Rosaceae 苹果属 Malus 2.0~2.5 花期3—4月,花粉红色,花量大 2 火棘 Pyracantha fortuneana 蔷薇科 火棘属 Pyracantha 0.8~1.2 花期3—4月,花白色,花量大;秋冬季红果 3 石楠 Photinia serratifolia 蔷薇科 石楠属 Photinia 3.0~3.5 花期5—7月,花白色,花量大 4 中华石楠 Photinia beauverdiana 蔷薇科 石楠属 3.0~3.5 花期5月,花白色,花量大;秋季红果 5 女贞 Ligustrum lucidum 木犀科Oleaceae 女贞属 Ligustrum 4.0~4.5 花期5—7月,花淡紫色 6 小叶女贞 Ligustrum quihoui 木犀科 女贞属 4.0~4.5 花期5—7月,花淡紫色 7 冬青 Ilex chinensis 冬青科Aquifoliaceae 冬青属 Ilex 4.5~5.0 花期4—6月,花紫红色或淡紫色;秋冬季红果 8 樟树 Cinnamomum camphora 樟科Lauraceae 樟属 Cinnamomum 14.0~16.0 树形优美,冠大荫浓 9 乌桕 Sapium sebiferum 大戟科Euphorbiaceae 乌桕属 Sapium 9.0~10.0 叶形独特,秋季红叶,秋冬季白果 10 苦楝 Melia azedarach 楝科Meliaceae 楝属 Melia 9.0~10.0 花期4—5月,花淡紫色;羽叶舒展秀丽 
图 1 10种树种生境
Figure 1. Habitats of 10 tree species
-
在2021年11月10—23日对鸟类取食情况进行预观测,对研究地鸟类食源树种进行调查,将2021年11月25日至2022年1月24日作为观测周期对鸟类取食情况进行系统观测,并记录相关数据。主要研究方法如下:①鸟类取食行为观测。每种植物各选2株作为固定观测对象,每间隔1 d,在8:30—10:00和14:30—16:00共2个时间段,使用胜途望远镜10×24标准款对食果鸟类进行观测,记录取食鸟类的种类、数量和取食对象。②拍照记录。拍照记录果实所处植物环境和果实形态变化。③果实内含物检测。在果实被鸟类大量取食的时间段内,每10 d摘取果实进行内含物测定,选择植物果实所共有的、在成熟过程中具有代表性的4个生理指标,即可滴定酸(氢氧化钠滴定法)、可溶性糖(蒽酮比色法)、淀粉(高氯酸蒽酮比色法)和可溶性蛋白(考马斯亮蓝G-250染色法)[17-18]。 ④相关性分析。用SPSS 26软件对取食鸟类数量与果实内含物进行相关性和显著性分析。
-
不同的植物在枝、叶和两者间构成的植物空间上展现不同的形态,使其果实所处的环境具备不同的特征。通过对10种树种形态的观测记录(表2,图2),将其果实所处的环境特征分为3类(图3):①常绿且枝硬舒展,区分明显内外空间,如石楠、女贞、冬青、樟树;②常绿且枝软密集,稍区分内外空间,如火棘;③落叶且枝硬舒展,不区分内外空间,如垂丝海棠、中华石楠、小叶女贞、乌桕、苦楝。
表 2 果实所处植物空间
Table 2. Spatial features of 10 tree species
类别 树种 叶 枝 植物空间 ① 石楠 全叶 枝舒展,比较硬朗 枝叶形成内部空间,空间较舒朗,视线较通透 女贞 全叶 枝舒展比较硬朗 冬青 全叶 枝舒展硬朗 樟树 全叶 枝舒展硬朗 ② 火棘 少叶 枝密集柔软,带尖刺 空间逼仄,视线较通透 ③ 垂丝海棠 无叶 枝舒展硬朗,带长钝刺类枝 空间舒朗,视线通透 中华石楠 无叶 枝舒展硬朗 小叶女贞 少叶 枝舒展比较硬朗 乌桕 无叶 枝舒展硬朗 苦楝 无叶 枝舒展硬朗 
图 2 果实所处植物空间
Figure 2. Tree spatial features with fruits

图 3 果实所处植物空间特征类型
Figure 3. Plant spatial features
-
在不同果熟阶段,果实的颜色、光泽和干枯状态等会产生变化。本研究时间段内10种树种果实的变化特征可分为2类(表3):①果实形态在果熟后60 d内基本不变,包括火棘、石楠、女贞、小叶女贞、冬青和樟树;②果实形态在果熟后60 d内随时间增加而逐渐枯萎,包括垂丝海棠、中华石楠、乌桕和苦楝。部分果实在40 d后因掉落或被取食而枝头无挂果,其余存留的部分果实在形态上无显著变化。其中垂丝海棠果实在同一植株同一时间段内同时存在3种果实状态(图4)。
表 3 10种树种果实形态变化特征
Table 3. Fruits morphology of 10 tree species
类别 树种 果实形态变化 备注 ① 火棘 颜色稍微加深变暗,整体维持在亮橘红色,果实形态基本不变 果实形态在果熟后60 d内基本不变 石楠 颜色稍微加深变暗,整体维持在暗红色,果实形态基本不变 女贞 颜色逐渐加深,由紫至紫黑,果实形态基本不变 小叶女贞 颜色逐渐加深,由紫至紫黑,果实形态基本不变 冬青 颜色基本不变,果实形态基本不变 樟树 颜色基本不变,表皮在后期微微变皱,失去光泽,果实形态基本不变 ② 垂丝海棠 颜色逐渐变红加深,由橘黄色至红褐色至暗褐色,果实形态萎缩干枯较快 果实形态在果熟后60 d内逐渐枯萎 中华石楠 颜色逐渐加深变暗,整体维持在亮红色,表皮逐渐干枯变皱,
果实形态在末期快速干枯变黑乌桕 颜色逐渐发黄发黑,表面出现脏黑杂物 苦楝 颜色逐渐加深,由黄绿变橙黄,果实形态逐渐变皱 
图 4 10种树种果实形态变化特征记录
Figure 4. Changing of fruit morphology of 10 tree species
-
不同果实的4类内含物质量分数均存在差异(表4)。果实的可滴定酸质量分数以苦楝最高,为(18.1±1.7) mg·g−1,樟树最低,为(2.3±0.2) mg·g−1;淀粉质量分数以苦楝最高,为(45.1±0.9) mg·g−1,石楠最低,为(4.3±0.3) mg·g−1;可溶性糖质量分数也以苦楝最高,为(247.00±4.05) mg·g−1,乌桕最低,为(2.23±0.86) mg·g−1;可溶性糖质量分数以樟树最高,为(13.77±0.08) mg·g−1,垂丝海棠最低,为(0.22±0.06) mg·g−1。在所选10种树种果实中,蔷薇科4种果实的可溶性糖和可溶性蛋白质量分数处于较低水平;木犀科2种果实中,小叶女贞果实的淀粉、可溶性糖和可溶性蛋白质量分数均高于女贞,处于较高水平;冬青果实的淀粉、可溶性糖和可溶性蛋白质量分数均处于较高水平;樟树的淀粉和可溶性蛋白质量分数处于较高水平;乌桕、苦楝的4类内含物质量分数均处于较高水平。
表 4 10种树种果实内含物质量分数
Table 4. Inclusions’contents in fruits of 10 tree species
序号 树种 科名 可滴定酸/(mg·g−1) 淀粉/(mg·g−1) 可溶性糖/(mg·g−1) 可溶性蛋白/(mg·g−1) 1 垂丝海棠 蔷薇科 14.6±2.5 b 5.2±0.3 de 41.84±0.51 e 0.22±0.06 i 2 火棘 蔷薇科 6.6±0.2 c 12.5±0.2 b 30.80±0.56 f 1.22±0.06 h 3 石楠 蔷薇科 5.9±0.2 cd 4.3±0.3 e 23.44±0.40 g 3.15±0.07 f 4 中华石楠 蔷薇科 3.8±1.0 ef 4.6±0.1 de 26.38±0.40 g 6.10±0.13 e 5 女贞 木犀科 4.5±0.4 de 5.8±0.2 d 68.85±2.40 d 6.51±0.03 d 6 小叶女贞 木犀科 4.9±0.1 cde 9.7±0.3 c 108.01±2.80 c 10.42±0.20 b 7 冬青 冬青科 6.4±0.6 cd 9.3±0.1 c 196.91±1.06 b 7.30±0.03 c 8 樟树 樟科 2.3±0.2 f 9.3±1.7 c 32.48±0.89 f 13.77±0.08 a 9 乌桕 大戟科 3.0±0.7 ef 11.8±1.4 b 2.23±0.86 h 1.64±0.10 g 10 苦楝 楝科 18.1±1.7 a 45.1±0.9 a 247.00±4.05 a 7.47±0.19 c 说明:同列不同字母表示不同植物间差异显著(P<0.05) -
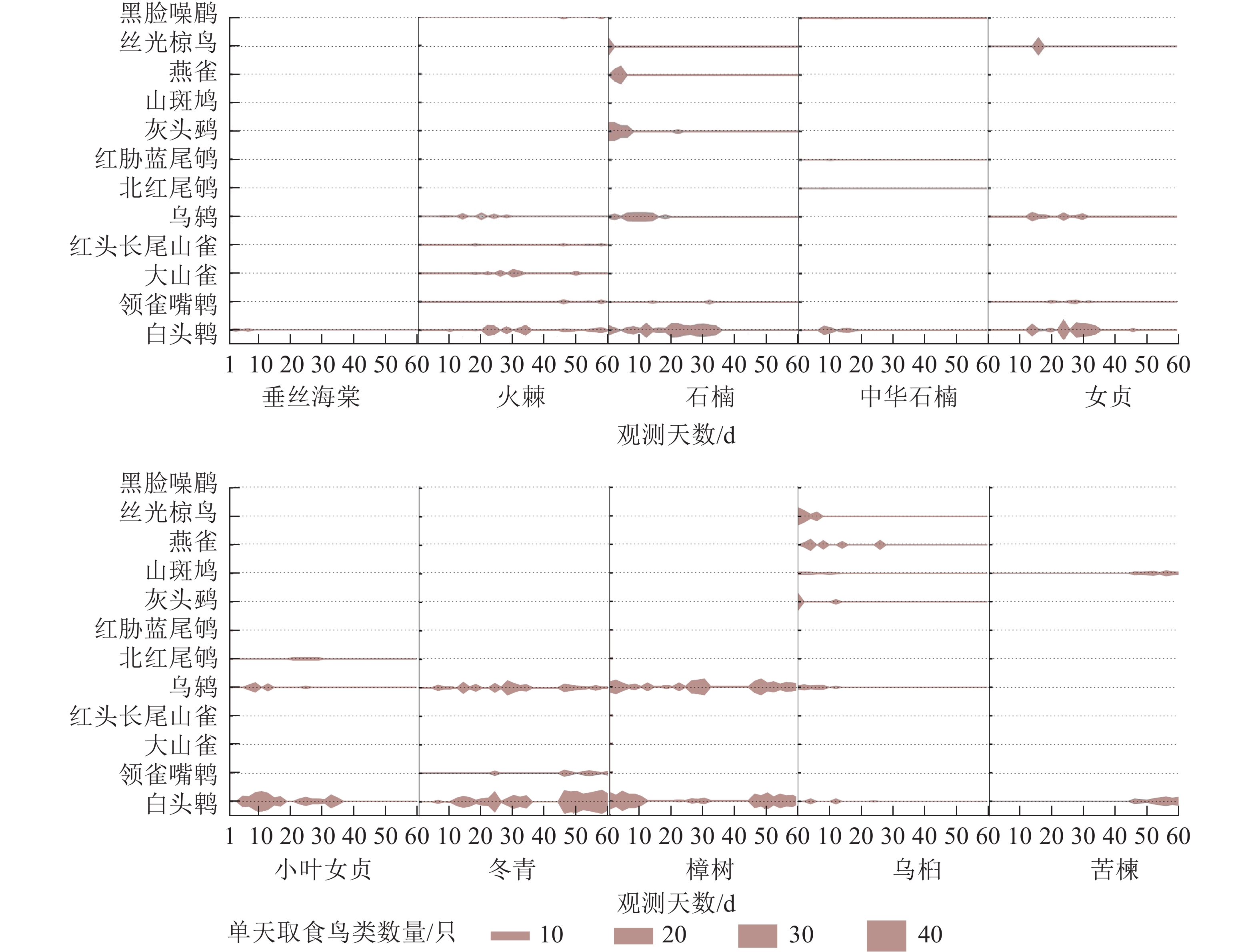
如图5所示:从不同鸟类对果实的选择看,白头鹎食性最广,对10种树种均有取食行为,且其取食的数量最多,占总量的55.2%;乌鸫对8种树种有取食行为,其取食数量占总量的21.3%;其他鸟类的取食对象多为1~3种树种,且数量相对较少。
图 5 10种树种与鸟类取食的相关性特征
Figure 5. Correlation between trees species and birds eating features of 10 tree species
从不同树种对鸟类的吸引方面,在引鸟数量上,石楠、冬青和樟树的引鸟数量最多,火棘、女贞、小叶女贞和乌桕次之,垂丝海棠、中华石楠和苦楝最少;在引鸟种类丰富度上,以火棘、石楠和乌桕最为突出,吸引了6种鸟类,其次是中华石楠和女贞,都只有4种,最少的是垂丝海棠,仅有白头鹎1种鸟类取食;在引鸟持续时间上,冬青和樟树最久,整个观测周期内都有取食,垂丝海棠最短,仅初期10 d左右被少量取食;火棘、石楠、女贞、小叶女贞和苦楝等取食时间均在40 d以上,中华石楠(15 d左右)和乌桕(30 d以下)的取食时间则相对较短。
-
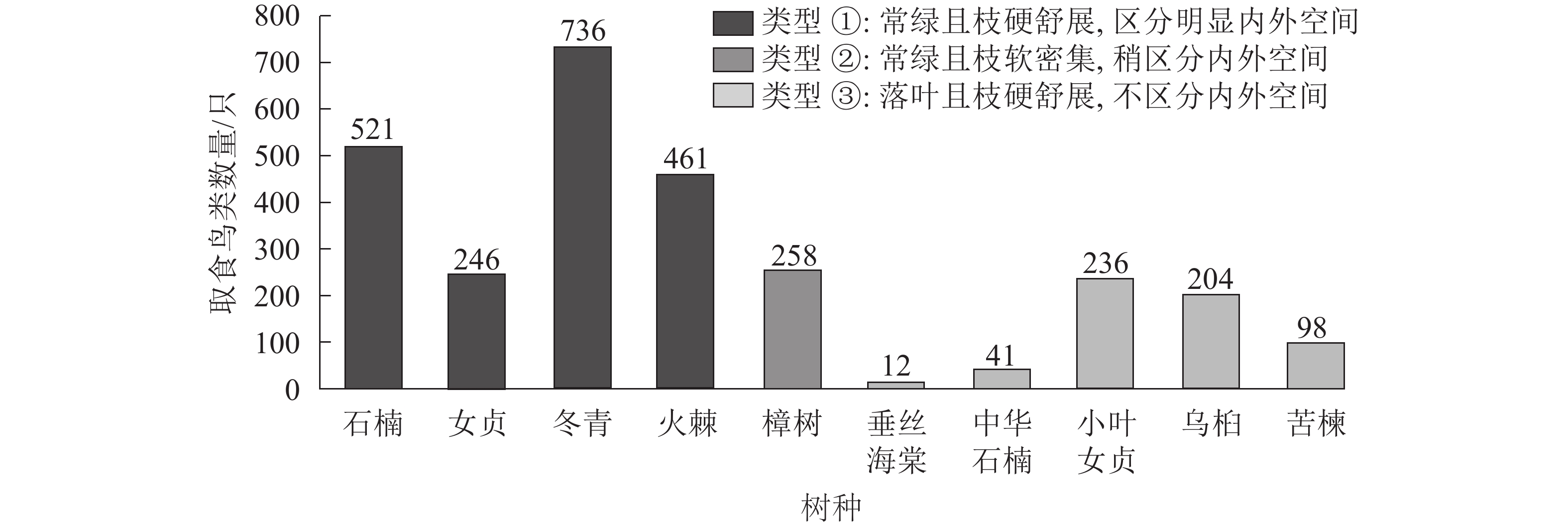
图6表明:处于第1类环境(常绿且枝硬舒朗、区分明显内外植物空间)的树种果实被鸟类取食的数量显著高于处在第2、3类植物空间中的果实,且在第3类环境中,小叶女贞、乌桕和苦楝果实的取食要多于垂丝海棠和中华石楠。

图 6 10种树种的果实所处植物空间类型与取食鸟类数量的关系
Figure 6. Relationship of 10 tree species’ spatial feature and bird numbers
果实处于第2类环境(枝软密集而内外空间略有分隔)的植物仅有火棘,不同鸟类对其取食方式有所不同(图7)。体形较大且活动于植物上层空间的鸟类如白头鹎、领雀嘴鹎等,往往停留于植物顶端且枝条相对硬朗的地方进行取食;体形较大而喜活动于地面空间的鸟类如黑脸噪鹛、乌鸫等,往往只取食垂落近地面枝条上的果实或地面落果;只有体形较小的鸟类如红头长尾山雀、大山雀等,会进入到火棘内部空间取食,但是该取食现象也相对较少。因此,在经过一段时间的取食后,火棘顶部和底部的果实被取食的较为干净,而中部和内部的果实可以保留很长时间。

图 7 火棘被鸟类取食情况
Figure 7. Birds eating fruits of P. fortuneana
-
研究(图8)发现:鸟类对第1类果实(形态在果熟后60 d内基本不变)的取食持续时间较长,一般持续到枝头被取食干净且几乎无挂果为止。但是,不同鸟类对第2类果实(形态在果熟后60 d内随时间增加而逐渐枯萎)的取食则有所不同。在观察白头鹎对垂丝海棠的取食行为中发现:由于垂丝海棠果实在同一株植物上同时存在不同的状态,白头鹎往往优先取食成熟度较高、颜色相对较深的果实,而相对青涩且颜色较浅的,以及开始萎缩干枯的果实则几乎没有被取食(图9)。在观察其他鸟类对中华石楠和乌桕果实的取食行为中发现:在果实处于饱满、新鲜状态时取食较多,在果实开始干枯后取食变少至消失,枝头仍有果实保留。
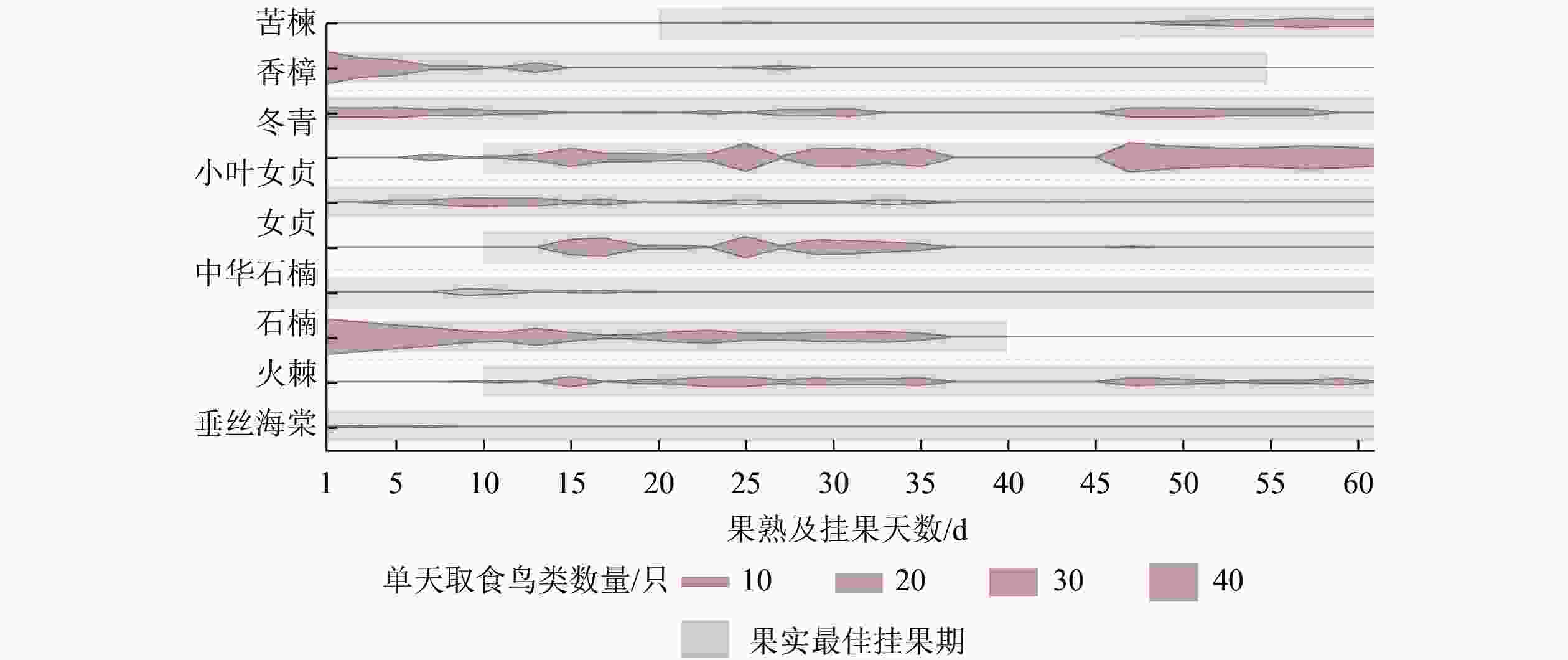
图 8 鸟类取食时段与植物果熟及挂果时段比较
Figure 8. Comparison of bird feeding periods with fruits ripening and hanging periods

图 9 垂丝海棠被取食现象
Figure 9. Feeding phenomenons of M. halliana
-
在选择相关性分析的研究对象时,应在取食鸟类、取食时间和食源树种3个方面进行甄别。第一,在所观测的食果鸟类中,白头鹎和乌鸫是主要种群,占据观测记录总数约75%,同时这2种鸟类的食性最广,分别取食了10种和8种果实,在食果鸟类中具有代表性。第二,在研究对象的时间段选择上,应考虑上述2种鸟类对各果实持续取食的时间,通过分析图5,认为在观测记录期第11~40天这2种鸟类对大部分植物果实都有持续性取食行为,相对比较适宜。第三,在食源树种选择中,应考虑果实所处植物空间特征和果实形态变化特征相近的植物,同时这2种鸟类都应具有对其取食的行为。综上,选定白头鹎和乌鸫,石楠、樟树和冬青作为研究对象,以第11~40天为研究时间段,统计此期间取食各果实的2种鸟类数量,以此为研究数据分析鸟类取食与果实内含物的相关性。
表5表明:虽然白头鹎和乌鸫的取食数量与4类内含物没有显著相关性,但是存在一定的影响关系。①可滴定酸质量分数与白头鹎和乌鸫取食数量的相关系数分别为0.882和0.211,呈正相关,其中可滴定酸与乌鸫取食数量的相关性较弱;②淀粉质量分数与白头鹎和乌鸫取食数量的相关系数分别为−0.775和−0.020,呈负相关,其中淀粉质量分数与乌鸫取食数量的相关性微弱;③可溶性糖质量分数与白头鹎和乌鸫取食数量的相关系数分别为0.941和0.866,呈正相关,相关性强;④可溶性蛋白质量分数与白头鹎取食数量的相关系数为−0.326,呈负相关,与乌鸫取食数量的相关系数为0.509,呈正相关。综上可知,只有可溶性糖质量分数与2种鸟类取食数量的相关性最强,对鸟类取食存在积极影响。
表 5 白头鹎、乌鸫取食数量与石楠、樟树、冬青果实各内含物质量分数的相关性分析
Table 5. Correlation analysis between feeding counts of P. sinensis, T. merula feeding and fruits contents of Ph. serratifolia, C. camphora, I. chinensis
生理指标 白头鹎数量 乌鸫数量 可滴定酸质量分数 0.882 0.211 淀粉质量分数 −0.775 −0.020 可溶性糖质量分数 0.941 0.866 可溶性蛋白质量分数 −0.326 0.509 说明:*表示在 0.05 (双尾)水平上显著相关 -
在鸟类生存的各类生境中,其种群多样性最高的是林地,其次是灌草丛和湖泊洼地,居民区最少,这与受人类活动干扰的影响有关[19-20]。在受干扰程度较高时,多数鸟类喜欢在高大的树木上栖息和取食[21-22],这与本次观测结果基本相同,即树木高大且隐蔽性较好的植物前来取食的鸟类数量较多。在常绿与落叶植物组合中,石楠、樟树等形成的植物空间比中华石楠、垂丝海棠等隐蔽。在落叶植物中,小叶女贞是长势较高的小乔木,乌桕和苦楝是大乔木且挂果位置远离地面,而中华石楠和垂丝海棠都是灌木或近小乔木,挂果位置离地面近。这2组植物中,每组的前者相比于后者,都能够为鸟类提供低干扰、高安全的取食环境,因此取食鸟类数量相对较多。同时鸟类为了规避潜在干扰和风险,对果实所处植物空间的起落便捷性也有较高要求,这与观测结果中鸟类对火棘的取食行为方式基本一致。由于火棘枝条柔软、生长紧密、互相穿插,且枝条上长有1~2 cm的尖刺,使得植物内部空间狭小错乱,同时火棘果实又簇生垂挂于枝条上,所处植物空间不利于鸟类停留起飞和活动,因此形成了顶端和底部的果实被大量取食而中间几乎无取食的现象。由此可见,植物果实所处的空间环境特征对鸟类取食具有显著影响,包含了安全隐蔽性和起落便捷性2个方面原因,安全隐蔽性越高、起落越便捷,鸟类的取食倾向就越明显,而在受人为干扰更大的城市环境中,这一现象可能会愈加显著。
鸟类对果实的选择也依赖于植物果实的基本特征。其中一个重要因素是果实的成熟期,鸟类往往喜欢取食成熟度较高的果实[23]。而果实成熟的本质是内含物发生改变,其主要特征之一是糖分的不断积累,结合本研究果实内含物与鸟类取食数量的相关性分析结果,说明鸟类更喜取食含糖量高的果实。另一个因素是果实成熟的形态特征,这是因为鸟类色觉敏锐,更喜颜色鲜艳的果实[24-25],同时果实的挂果状态及保持能力体现了果实成熟后的新鲜程度和内含物稳定程度。结合本研究观测结果可知:在果实颜色明亮、色泽饱满时,鸟类取食倾向性高,在果实成熟过后开始枯萎而颜色暗淡时,鸟类取食倾向性变低。
此外,植物果实中还存在较多因素影响着鸟类的取食倾向。如火棘,虽然果实所处空间不利于鸟类取食,果实含糖量比其他种类果实要低,但是在引鸟数量和引鸟种类多样性上效果都较好,除开果小和颜色鲜艳明亮外,火棘果实氨基酸种类和含量均丰富[26],这可能是其受鸟类青睐的重要因素。再如乌桕,果实在同一时间段的含糖量比其他种类果实要低,但其油脂含量超高,这可能是促使其被大量取食的原因。相反,苦楝果实的含糖量相比于其他种类果实要高,但是果实含有毒成分[27],这可能是只有在后期果实减少的情况下才记录到其被取食现象的原因。
本次研究结果可为长江三角洲地区城市引鸟植物景观营建提供以下建议:在食源树种选择上,应考虑具有较好挂果保持能力和较高含糖量果实的树种,重点考虑常绿或高大乔木类挂果树种;在食源树种种植位置上,应充分考虑其隐蔽性,特别是涉及有落叶灌木或小乔木时,应种植在人为干扰较小的区域或与高大乔木合理搭配,为鸟类提供安全的取食空间。
Preference of birds to fruit characteristics of common landscaping trees in campus green spaces in Hangzhou
-
摘要:
目的 分析果实特性,探究鸟类取食行为与秋冬季食源树种果实特性的相关性,为城市引鸟植物景观营建和生物多样性规划提供理论支撑。 方法 定点定时对鸟类取食行为进行周期性观测记录,根据前期观测结果,选择鸟类最为喜好的10种常见园林树种,拍照记录其果实所处的环境和果实形态变化,检测被取食期间果实的可滴定酸、可溶性糖、淀粉和可溶性蛋白的质量分数,并进行相关性分析。 结果 果实所处植物空间可分为3类:①常绿且枝硬舒展,区分明显内外空间;②常绿且枝软密集,稍区分内外空间等;③落叶且枝硬舒展,不区分内外空间。果实所处植物空间特征的安全隐蔽性和起落便捷性会直接影响鸟类是否倾向于取食该果实,同时果实空间环境的内部特点会影响鸟类取食该果实的方式,针对枝软且复杂逼仄的环境,体形较大的鸟类例如领雀嘴鹎Spizixos semitorques等会选择停留于植物顶部取食,体形较小的鸟类例如大山雀Parus major等会选择进入植物内部取食。果实的形态变化特征可分为2类:①果实形态在果熟后60 d内基本不变;②果实形态在果熟后60 d内随时间增加而逐渐枯萎。其果实挂果状态会影响鸟类的取食倾向,新鲜饱满的果实对鸟类更具吸引力,挂果状态保持较差的树种如垂丝海棠Malus halliana等只在果实成熟的早期阶段被白头鹎Pycnonotus sinensis取食,挂果状态保持良好的树种如樟树Cinnamomum camphora等则被白头鹎和乌鸫Turdus merula长期持续取食。果实的内含物会影响鸟类的取食倾向,含糖量高的果实更易吸引白头鹎和乌鸫,白头鹎更倾向于酸质量分数较高的果实而乌鸫更倾向于蛋白质质量分数较高的果实。 结论 为了提升城市绿地中的鸟类多样性,应保证食源树种种植位置的隐蔽性,重点选择具有较好挂果状态保持能力和较高含糖量的常绿高大乔木作为其食源树种。图9表5参27 Abstract:Objective Analyzing the fruit characteristics and investigating the correlation between bird feeding behavior and fruit characteristics in autumn and winter aims to provide theoretical insights for urban bird-attracting plant landscape construction and biodiversity planning. Method The periodic investigation of birds feeding behavior was carried out at three sites in the East Lake campus of Zhejiang A&F University. The spatial environments that fruits located in and the changes of fruit external morphology about 10 common landscaping tree species were recorded. The contents of fruits including titratable acid, soluble sugar, starch and soluble protein were gauged during the birds feeding period. Result The spatial environments that fruits located in could be divided into three categories: (1) evergreen trees that inner and outer space could be distinguished easily with hard and spreading branches; (2) evergreen trees that inner and outer space could be distinguished slightly with soft and dense branches; (3) deciduous trees that inner and outer space could not be distinguished with hard and spreading branches. The security and concealment of spatial environments that fruits located in and the convenience for bird landing and taking-off would directly affect the tendency of bird feeding, and the internal characteristics of spatial environment would affect approaches of bird feeding. When it came soft branches and complex spatial environment, larger birds such as Spizixos semitorques chose to feed on the top of plants , while smaller birds such as Parus major chose to feed inside. Fruits’external morphological characteristic could be divided into two categories: (1) the fruits with unchanged external morphology within 60 days; (2) the fruits that external morphology gradually withered as time flowed within 60 days. Fruit hanging status affected the feeding tendency of birds as fresh and plump fruits were more attractive to birds. Fruits that were poorly maintained, such as Malus halliana, were only eaten by Pycnonotus sinensis at the early stage of fruit ripening, while fruits that were well maintained, such as Cinnamomum camphora, were continuously eaten by P. sinensis and Turdus merula for a long time. The contents of the fruit affected the feeding tendency of the birds. Fruits with high sugar content were more attractive to P. sinensis and T. merula, while the P. sinensis prefered fruits with higher acid content and the T. merula prefered fruits with higher protein content. Conclusion In order to improve bird diversity in urban green space, the concealing of food source tree should be ensured, and the tall evergreen trees owning fruits with high retention ability and high sugar content should be selected as the food source trees. [Ch, 9 fig. 5 tab. 27 ref.] -
Key words:
- landscape trees /
- bird feeding /
- plant space /
- fruit morphology /
- fruit sugar content
-
图 5 10种树种与鸟类取食的相关性特征
Figure 5 Correlation between trees species and birds eating features of 10 tree species
图 6 10种树种的果实所处植物空间类型与取食鸟类数量的关系
Figure 6 Relationship of 10 tree species’ spatial feature and bird numbers
图 8 鸟类取食时段与植物果熟及挂果时段比较
Figure 8 Comparison of bird feeding periods with fruits ripening and hanging periods
表 1 10种鸟类食源树种概况
Table 1. Overview of 10 tree species as food source
序号 树种 科名 属名 株高/m 观赏特性 1 垂丝海棠 Malus halliana 蔷薇科Rosaceae 苹果属 Malus 2.0~2.5 花期3—4月,花粉红色,花量大 2 火棘 Pyracantha fortuneana 蔷薇科 火棘属 Pyracantha 0.8~1.2 花期3—4月,花白色,花量大;秋冬季红果 3 石楠 Photinia serratifolia 蔷薇科 石楠属 Photinia 3.0~3.5 花期5—7月,花白色,花量大 4 中华石楠 Photinia beauverdiana 蔷薇科 石楠属 3.0~3.5 花期5月,花白色,花量大;秋季红果 5 女贞 Ligustrum lucidum 木犀科Oleaceae 女贞属 Ligustrum 4.0~4.5 花期5—7月,花淡紫色 6 小叶女贞 Ligustrum quihoui 木犀科 女贞属 4.0~4.5 花期5—7月,花淡紫色 7 冬青 Ilex chinensis 冬青科Aquifoliaceae 冬青属 Ilex 4.5~5.0 花期4—6月,花紫红色或淡紫色;秋冬季红果 8 樟树 Cinnamomum camphora 樟科Lauraceae 樟属 Cinnamomum 14.0~16.0 树形优美,冠大荫浓 9 乌桕 Sapium sebiferum 大戟科Euphorbiaceae 乌桕属 Sapium 9.0~10.0 叶形独特,秋季红叶,秋冬季白果 10 苦楝 Melia azedarach 楝科Meliaceae 楝属 Melia 9.0~10.0 花期4—5月,花淡紫色;羽叶舒展秀丽  下载: 导出CSV
下载: 导出CSV
表 2 果实所处植物空间
Table 2. Spatial features of 10 tree species
类别 树种 叶 枝 植物空间 ① 石楠 全叶 枝舒展,比较硬朗 枝叶形成内部空间,空间较舒朗,视线较通透 女贞 全叶 枝舒展比较硬朗 冬青 全叶 枝舒展硬朗 樟树 全叶 枝舒展硬朗 ② 火棘 少叶 枝密集柔软,带尖刺 空间逼仄,视线较通透 ③ 垂丝海棠 无叶 枝舒展硬朗,带长钝刺类枝 空间舒朗,视线通透 中华石楠 无叶 枝舒展硬朗 小叶女贞 少叶 枝舒展比较硬朗 乌桕 无叶 枝舒展硬朗 苦楝 无叶 枝舒展硬朗
下载: 导出CSV
表 3 10种树种果实形态变化特征
Table 3. Fruits morphology of 10 tree species
类别 树种 果实形态变化 备注 ① 火棘 颜色稍微加深变暗,整体维持在亮橘红色,果实形态基本不变 果实形态在果熟后60 d内基本不变 石楠 颜色稍微加深变暗,整体维持在暗红色,果实形态基本不变 女贞 颜色逐渐加深,由紫至紫黑,果实形态基本不变 小叶女贞 颜色逐渐加深,由紫至紫黑,果实形态基本不变 冬青 颜色基本不变,果实形态基本不变 樟树 颜色基本不变,表皮在后期微微变皱,失去光泽,果实形态基本不变 ② 垂丝海棠 颜色逐渐变红加深,由橘黄色至红褐色至暗褐色,果实形态萎缩干枯较快 果实形态在果熟后60 d内逐渐枯萎 中华石楠 颜色逐渐加深变暗,整体维持在亮红色,表皮逐渐干枯变皱,
果实形态在末期快速干枯变黑乌桕 颜色逐渐发黄发黑,表面出现脏黑杂物 苦楝 颜色逐渐加深,由黄绿变橙黄,果实形态逐渐变皱
下载: 导出CSV
表 4 10种树种果实内含物质量分数
Table 4. Inclusions’contents in fruits of 10 tree species
序号 树种 科名 可滴定酸/(mg·g−1) 淀粉/(mg·g−1) 可溶性糖/(mg·g−1) 可溶性蛋白/(mg·g−1) 1 垂丝海棠 蔷薇科 14.6±2.5 b 5.2±0.3 de 41.84±0.51 e 0.22±0.06 i 2 火棘 蔷薇科 6.6±0.2 c 12.5±0.2 b 30.80±0.56 f 1.22±0.06 h 3 石楠 蔷薇科 5.9±0.2 cd 4.3±0.3 e 23.44±0.40 g 3.15±0.07 f 4 中华石楠 蔷薇科 3.8±1.0 ef 4.6±0.1 de 26.38±0.40 g 6.10±0.13 e 5 女贞 木犀科 4.5±0.4 de 5.8±0.2 d 68.85±2.40 d 6.51±0.03 d 6 小叶女贞 木犀科 4.9±0.1 cde 9.7±0.3 c 108.01±2.80 c 10.42±0.20 b 7 冬青 冬青科 6.4±0.6 cd 9.3±0.1 c 196.91±1.06 b 7.30±0.03 c 8 樟树 樟科 2.3±0.2 f 9.3±1.7 c 32.48±0.89 f 13.77±0.08 a 9 乌桕 大戟科 3.0±0.7 ef 11.8±1.4 b 2.23±0.86 h 1.64±0.10 g 10 苦楝 楝科 18.1±1.7 a 45.1±0.9 a 247.00±4.05 a 7.47±0.19 c 说明:同列不同字母表示不同植物间差异显著(P<0.05)
下载: 导出CSV
表 5 白头鹎、乌鸫取食数量与石楠、樟树、冬青果实各内含物质量分数的相关性分析
Table 5. Correlation analysis between feeding counts of P. sinensis, T. merula feeding and fruits contents of Ph. serratifolia, C. camphora, I. chinensis
生理指标 白头鹎数量 乌鸫数量 可滴定酸质量分数 0.882 0.211 淀粉质量分数 −0.775 −0.020 可溶性糖质量分数 0.941 0.866 可溶性蛋白质量分数 −0.326 0.509 说明:*表示在 0.05 (双尾)水平上显著相关
下载: 导出CSV
-
[1] 马远, 李锋, 杨锐. 城市化对生物多样性的影响与调控对策[J]. 中国园林, 2021, 37(5): 6 − 13. MA Yuan, LI Feng, YANG Rui. The impact of urbanization on biodiversity and its regulation countermeasures [J]. Chin Landscape Archit, 2021, 37(5): 6 − 13. [2] SHWARTZ A, TURBE A, SIMON L, et al. Enhancing urban biodiversity and its influence on city-dwellers: an experiment [J]. Biol Conserv, 2014, 171: 82 − 90. [3] 张征恺, 黄甘霖. 中国城市鸟类学研究进展[J]. 生态学报, 2018, 38(10): 3357 − 3367. ZHANG Zhengkai, HUANG Ganlin. Urban ornithological research in China: a review [J]. Acta Ecol Sin, 2018, 38(10): 3357 − 3367. [4] 赵伊琳, 王成, 白梓彤, 等. 城市化鸟类群落变化及其与城市植被的关系[J]. 生态学报, 2021, 41(2): 479 − 489. ZHAO Yilin, WANG Cheng, BAI Zitong, et al. Changes of bird community under urbanization and its relationship with urban vegetation [J]. Acta Ecol Sin, 2021, 41(2): 479 − 489. [5] 陆彩虹, 鲁长虎. 南京中山植物园鸟类对香樟果实(种子)的取食[J]. 动物学杂志, 2019, 54(6): 784 − 792. LU Caihong, LU Changhu. Bird feeding on the fruits (seeds) of camphor trees in Nanjing Zhongshan Botanical Garden, China [J]. Chin J Zool, 2019, 54(6): 784 − 792. [6] 邓娇, 晏玉莹, 张志强, 等. 城市化对长沙市区城市公园繁殖期鸟类物种多样性的影响[J]. 生态学杂志, 2014, 33(7): 1853 − 1859. DENG Jiao, YAN Yuying, ZHANG Zhiqiang, et al. Influence of urbanization on bird species diversity in urban parks in Changsha, Hunan Province during the breeding period [J]. Chin J Ecol, 2014, 33(7): 1853 − 1859. [7] 周放. 鼎湖山森林鸟类群落的集团结构[J]. 生态学报, 1987, 7(2): 176 − 184. ZHOU Fang. Guild structure of the forest bird community in Dinghushan [J]. Acta Ecol Sin, 1987, 7(2): 176 − 184. [8] 王玲, 丁志锋, 胡君梅, 等. 广州城市绿地中鸟类对食源树种的偏好[J]. 四川动物, 2016, 35(6): 838 − 844. WANG Ling, DING Zhifeng, HU Junmei, et al. Bird preference to food source trees in urban green space, Guangzhou, China [J]. Sichuan J Zool, 2016, 35(6): 838 − 844. [9] DONOSO I, SCHLEUNING M, GARCIA D. et al. Defaunation effects on plant recruitment depend on size matching and size trade-offs in seed-dispersal networks[J/OL]. Proc Biol Sci, 2017, 284(1855): 20162664[2022-04-02]. doi: 10.1098/rspb.2016.2664. [10] 陆彩虹. 中山植物园鸟播植物果实特征及食果鸟类的取食行为[D]. 南京: 南京林业大学, 2020. LU Caihong. Fruit Characteristics of Bird-seeded Plants in Zhongshan Botanical Garden and Their Feeding Behaviors[D]. Nanjing: Nanjing Forest University, 2020. [11] DUAN Qiong, GOODALE E, QUAN Ruichang. Bird fruit preferences match the frequency of fruit colors in tropical Asia[J/OL]. Sci Rep, 2014, 4(1) : 5627[2022−04−02]. doi: 10.1038/srep05627. [12] ALMAZAN-NUNEZ R C, ALVAREZ-ALVAREZ E A, SIERRA-MORALES P, et al. Fruit size and structure of zoochorous trees: identifying drivers for the foraging preferences of fruit-eating birds in a Mexican successional dry forest[J/OL]. Animals, 2021, 11(12): 3343[2022-04-02]. doi: 10.3390/ani11123343. [13] 何海燕, 王楠, 董路. 北京城市鸟类对食源植物利用规律[J]. 动物学杂志, 2021, 56(4): 491 − 499. HE Haiyan, WANG Nan, DONG Lu. A case study to investigate the foraging pattern of urban birds on edible plants in Beijing [J]. Chin J Zool, 2021, 56(4): 491 − 499. [14] LEI Boyu, CUI Jifa, NEWMAN C, et al. Seed dispersers shape the pulp nutrients of fleshy-fruited plants[J/OL]. Proc Royal Soc Biol Sci, 2021, 288(1953): 20210817[2022-04-02]. doi: 10.1098/rspb.2021.0817. [15] PIZO M A, MORALES J M, OVASKAINEN O, et al. Frugivory specialization in birds and fruit chemistry structure mutualistic networks across the neotropics [J]. Am Nat, 2021, 197(2): 236 − 249. [16] 中国教育在线. 3300种植物 400种入驻鸟类 浙农林大蝉联中国大学校园植物排行榜第一[EB/OL]. (2020-05-25)[2022-05-05]. https://www.eol.cn/zhejiang/zhejiang_news/202005/t20200525_1729048.shtml. China Education Online. 3300 species of plants, 400 species of birds settled in Zhejiang A&F University won the first place in the list of campus plants of Chinese universities [EB/OL]. (2020-05-25)[2022-05-05]. https://www.eol.cn/zhejiang/zhejiang_news/202005/t20200525_1729048.shtml. [17] 张宪政, 谭桂茹. 植物生理学实验技术[M]. 沈阳: 辽宁科学技术出版社, 1989. ZHANG Xianzheng, TAN Guiru. Experimental Techniques of Plant Physiology[M]. Shenyang: Liaoning Science and Technology Publishing House, 1989. [18] 肖望. 植物生理学实验指导[M]. 广州: 中山大学出版社, 2020. XIAO Wang. Experimental Guidance in Plant Physiology[M]. Guangzhou: Sun Yat-sen University Press, 2020. [19] PAKER Y, YOM-TOV Y, ALON-MOZES T, et al. The effect of plant richness and urban garden structure on bird species richness, diversity and community structure [J]. Landscape Urban Plann, 2014, 122: 186 − 195. [20] 罗祖奎, 李扬, 徐曦. 鸟类群落结构及分布研究——以贵阳市观山湖区为例[J]. 河南师范大学学报(自然科学版), 2021, 49(5): 104 − 111. LUO Zukui, LI Yang, XU Xi. Study on the bird community structure and distribution in Guanshanhu District of Guiyang City [J]. J Henan Norm Univ Nat Sci Ed, 2021, 49(5): 104 − 111. [21] 干靓, 吴志强, 郭光普. 高密度城区建成环境与城市生物多样性的关系研究——以上海浦东新区世纪大道地区为例[J]. 城市发展研究, 2018, 25(4): 97 − 106. GAN Jing, WU Zhiqiang, GUO Guangpu. The influence of built environment on urban biodiversity in high-density urban areas: case study in blocks along century avenue, Pudong New District, Shanghai [J]. Urban Dev Stud, 2018, 25(4): 97 − 106. [22] 史慧灵, 白皓天, 吴良早, 等. 昆明城市绿地结构对鸟类多样性的影响[J]. 四川动物, 2016, 35(5): 774 − 780. SHI Huiling, BAI Haotian, WU Liangzao, et al. The effect of urban space structure on bird diversity in Kunming [J]. Sichuan J Zool, 2016, 35(5): 774 − 780. [23] 隋金玲, 张志翔, 胡德夫, 等. 北京市区绿化带内鸟类食源树种研究[J]. 林业科学, 2006, 42(12): 83 − 89. SUI Jinling, ZHANG Zhixiang, HU Defu, et al. Studies on bird-feed trees at green belts of Beijing urban area [J]. Sci Silv Sin, 2006, 42(12): 83 − 89. [24] SINNOTT-ARMSTRONG M A, LEE C, CLEMENT W L, et al. Fruit syndromes in Viburnum: correlated evolution of color, nutritional content, and morphology in bird-dispersed fleshy fruits[J/OL]. BMC Evol Biol, 2020, 20(1): 7[2022-04-02]. doi: 10.1186/s12862-019-1546-5. [25] TEICHMANN M, THOROGOOD R, HAMALAINEN L. Seeing red? colour biases of foraging birds are context dependent [J]. Anim Cognition, 2020, 23(5): 1007 − 1018. [26] 宋小娟, 杨卫灵, 张清海, 等. 野生火棘果活性成分及抗氧化和抑菌性的研究[J]. 食品研究与开发, 2022, 43(7): 45 − 51, 133. SONG Xiaojuan, YANG Weiling, ZHANG Qinghai, et al. Study of active components in wild Pyracantha fortuneana fruit and their antioxidant and antibacterial properties [J]. Food Res Dev, 2022, 43(7): 45 − 51, 133. [27] ZENG Fagu, SU Qian, DI Yingtong, et al. Penylpropanoids with cytotoxic activity from the fruits of Melia azedarach [J]. Nat Prod Res Dev, 2016, 28(8): 1171 − 1175. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220359
 点击查看大图
点击查看大图

计量
- 文章访问数: 2390
- HTML全文浏览量: 474
- PDF下载量: 26
- 被引次数: 0
