




-
土壤动物及土壤微生物的活动能够改善土壤的通气状况、养分有效性, 在土壤形成、发育、演化及土壤肥力形成演变中发挥着重要的作用。土壤动物数量繁多, 通过参与一系列生命活动, 直接或间接改变土壤的性质[1], 对土壤形成、发育及生态系统物质循环都具有重要意义。稻草还田是一种有效的农田培肥措施[3], 在避免稻草焚烧造成环境污染的同时, 也为农业生产提供有机肥源。作为重要肥料来源和潜在的碳库能源[2], 还田的稻草在增加土壤养分[4], 培肥地力, 改善土壤理化性状[5], 优化农田生态环境, 提高作物产量与品质[6-9]等方面意义重大。本研究以未进行稻草还田的处理为对照, 设置不同质量稻草还田处理, 并调查不同处理下农田土壤动物群落特征, 分析不同稻草还田量对土壤动物群落结构的影响, 旨在为农田耕地环境的保护及可持续农业发展提供理论依据。
-
研究区四川省都江堰市浦阳镇金凤村(31°0′23″N, 103°37′18″E)属四川盆地亚热带湿润气候, 年平均气温为15.2 ℃, 年降水量为1 243.8 mm, 空间分布不均, 呈东南向西北减少, 无霜期269.0 d, 年蒸发量为930.1 mm。土壤类型为山地黄壤。样地为水稻Oryza sativa-油菜Brassica campestris轮作模式, 水稻品种为‘川优6203’‘Chuanyou 6203’, 油菜品种为‘绵油88’‘Mianyou 88’。
-
2016年4月, 在耕地内按照具有代表性的原则设置2个面积均为10 m × 30 m的样地; 各样地内设置面积为5 m × 5 m样方各4个, 保持各样方间间距大于2 m, 其中4个为重复样地。随机编号, 按照单位面积稻草的平均产生量, 选取最新收获的水稻稻草, 切成3~5段后按0(ck)、0.2(T5)、0.4(T10)、0.8 kg·m-2(T20)的稻草均匀铺撒在样方上进行还田。
分别在还田2个月和5个月时, 在各样方内按"品"字形布点, 随机设置3个大小为50 cm × 50 cm的小样方, 采集0~5、5~10、10~15 cm深度土层。手捡法分层收集土壤中的大型动物, 并用体积分数为75%的乙醇将动物杀死, 带回实验室。环刀法(r=5 cm, V=100 cm3)自上而下依次取土样, 每层各取2个, 用尼龙网包好贴上标签迅速放入黑布袋带回实验室; 用Tullgren[10]干漏斗和Baermann[11]湿漏斗分离土壤中的中小型土壤动物, 分离时间为48 h, 其中湿生土壤动物每4 h观察1次。干生分离出的土壤动物用盛有体积分数为75%乙醇的器皿收集, 湿生土壤动物用清水收集, 解剖镜下进行分类鉴定和数量统计。
所得土壤动物用双目解剖镜(Leica, EZ4HD)进行观察, 参照《中国土壤动物检索图鉴》[12]、《昆虫分类检索》[13]、《幼虫分类学》[14]进行分类鉴定, 一般鉴定到科, 同时统计个体数量。
-
土壤动物多样性分析:采用Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)[15]、Pielou均匀度指数(J)[16]、Simpson优势度指数(C)[17]。计算公式如下:Shannon-Wiener多样性指数${H^\prime } = - \sum\limits_{i = 1}^s {{P_i}} \ln {P_i}$。式中:Pi=ni/N, ni为第i个类群的个体数; N为所有类群的个体数。Margalef丰富度指数D=(S-1)/lnN。式中:S为类群数, N为所有类群的个体数。Pielou均匀度指数J=H′/lnS。式中:H′为Shannon-Wiener多样性指数, S为类群数。Simpson优势度指数$C = \sum\limits_{i = 1}^s {P_i^2} $。其中:ni为第i个类群的个体数; N为所有类群的个体数。
类群数量等级划分:个体数量大于捕获总量的10.0%者为优势类群(+++), 大于等于1.0%小于等于10.0%者为常见类群(++), 小于1.0%者为稀有类群(+)。
采用Excel 2010和SPSS 22.0完成数据处理和分析。用单因素方差分析(one-way ANOVA)对不同样方间土壤动物群落组成进行检验; 用LSD(方差齐性)法进行多重比较, 显著性水平设定为P=0.05[18]。
-
本研究共捕获土壤动物1 194只, 隶属于3门11纲19目44个类群, 类群数排序为T20>T10>T5>对照。优势类群为线虫纲Nematoda和懒甲螨科Nothridae, 分别占总捕获量的72.16%和11.09%;常见类群为蚁科Formicidae、线蚓科Enchytraeidae、绥螨科Sejidae、丽甲螨科Liacaridae、双翅目Diptera幼虫, 分别占总量的4.33%、4.00%、2.41%、1.95%和1.59%;棘科Onychiuridae、跳虫科Poduridae、露尾甲科Nitidulidae等构成稀有类群, 占总量的2.47%。
由表 1可见:还田2个月后各样地的优势类群都为线虫纲和懒甲螨科, 其中:T20样地2次捕获的土壤动物包含30个类群, 平均密度4.13×104只·m-2; T10样地2次捕获的土壤动物隶属于20科, 平均密度为4.83×104只·m-2; T5样地2次捕获的土壤动物隶属于21科, 平均密度4.73×104只·m-2; 对照样地2次捕获的土壤动物隶属于11科, 平均密度3.70×104只·m-2。还田5个月后T20、T10、T5样地的优势类群都为线虫纲, 对照样地优势类群为线虫纲和蚁科, 其中:T20样地2次捕获的土壤动物包含22科, 优势类群为线虫纲, 平均密度9.32×104只·m-2; T10样地2次捕获的土壤动物隶属于20科, 平均密度8.48×104只·m-2; T5样地2次捕获的土壤动物隶属于19科, 平均密度4.75×104只·m-2; 对照样地2次捕获的土壤动物隶属于11科, 平均密度4.60×104只·m-2。
表 1 不同样地土壤动物群落密度统计
Table 1. Compositions of soil fauna community in the different plots
类群 还田2个月/(只·m-2) 还田5个月/(只·m-2) 总计/(只·m-2) 多度 T20 T10 T5 ck T20 T10 T5 ck 线虫纲Nematoda 60.13 52.13 38.43 82.41 77.79 88.00 87.51 73.90 72.16 +++ 懒甲螺科Nothridae 26.03 27.45 49.36 5.40 0.09 0 0 0 11.09 +++ 蚁科Formicidae 0.84 0.01 6.35 0.46 9.03 0 0.35 15.58 4.33 ++ 线蚓科Enchytraeidae 0 0 0 1.80 6.08 8.45 6.31 2.90 4.00 ++ 绥螨科Sejidae 4.24 3.62 3.00 4.50 1.07 1.57 1.40 2.54 2.41 ++ 丽甲螨科Liacaridae 0.61 0 0 0 5.28 0.69 2.28 3.98 1.95 ++ 双翅目幼虫Diptera larvae 0.20 13.12 0 1.80 0 0 0 0 1.59 ++ 棘䖴科Onychiuridae 0.81 0.35 1.23 2.25 0.27 0.10 0.70 0.36 0.62 + 跳虫科Poduridae 1.21 0.17 0.53 0 0 0.10 0.35 0.72 0.32 + 露尾甲科Nitidulidae 1.21 0 0 0 0 0.79 0 0 0.26 + 白蚁科Termitidae 2.42 0 0 0 0 0 0 0 0.22 + 蝴蛛目Araneae 0.01 1.04 0.18 0 0 0.20 0 0 0.17 + 叶蝉科Cicadellidae 0 0.69 0.18 0.90 0 0 0 0 0.17 + 蚁甲亚科Pselaphidae 0 0 0.18 0.45 0 0 0.53 0 0.11 + 叶甲科Chrysomelidae 0.20 0.52 0.18 0 0 0 0 0 0.10 + 幺蚰科Scutigerellidae 0 0.17 0 0 0.27 0.10 0 0 0.09 + 长角长䖴科Orchesellidae 0.81 0 0 0 0 0 0 0 0.07 + 蜈蚣目Scolopendromorpha 0.21 0.17 0 0 0 0 0.18 0 0.06 + 圆䖴科Sminthuridae 0.20 0.35 0 0 0 0 0 0 0.06 + 蜚蠊科Blattidae 0.01 0 0 0 0 0 0.35 0 0.04 + 拟步甲科Tenebrionidae 0.41 0 0.01 0 0 0 0 0 0.04 + 派盾螨科Parholaspididae 0.20 0 0.18 0 0 0 0 0 0.04 + 木螱科Kalotermitidae 0.20 0.17 0 0 0 0 0 0 0.04 + 等节䖴科Isotomidae 0 0 0 0 0.09 0 0 0 0.02 + 蟋蟀科Gryllidae 0 0 0.18 0 0 0 0 0 0.02 + 小蚓类Microdrile oligochaetes 0.02 0.03 0.02 0.01 0 0 0 0 0.01 + 蠼螋科Labiduridae 0 0 0 0 0.01 0 0 0 0 + 鼠妇科Porcellionidae 0.01 0 0 0 0 0 0.01 0 0 + 蝼蛄科Gryllotalpidae 0 0 0 0 0 0 0.01 0 0 + 步甲科Carabidae 0 0 0 0 0 0 0.01 0 0 + 姬马陆科Julidae 0.01 0 0 0 0 0 0 0 0 + 金龟甲科幼虫Scarabaeidae larvae 0 0 0 0 0 0 0 0 0 + 石蜈蚣目Lithobiomorpha 0 0 0 0 0 0 0 0 0 + 猎蝽科Reduviidae 0 0 0 0 0 0 0 0 0 + 隐翅虫科Staphylinidae 0 0 0 0 0 0 0 0 0 + 蛭纲Hirudinea 0 0 0 0 0 0 0 0 0 + 叩甲科Elateridae 0 0 0 0 0 0 0 0 0 + 天牛科Cerambycidae 0 0 0 0 0 0 0 0 0 + 瓢甲科Coccinellidae 0 0 0 0 0 0 0 0 0 + 蝗科Acrididae 0 0 0 0 0 0 0 0 0 + 角板盲蛛科Ceratolasmatidae 0 0 0 0 0 0 0 0 0 + 缘蝽科Coreidae 0 0 0 0 0 0 0 0 0 + 夜蛾科Noctuidae 0 0 0 0 0 0 0 0 0 + 锹甲科Lucanidae 0 0 0 0 0 0 0 0 0 + 合计 41 302.01 48 277.34 47 269.99 37 009.33 93 197.35 84 846.68 47 524.66 46 007.99 445 435.35 总类群数 30 20 21 11 22 20 19 11 44 -
由图 1A可知:还田5个月后各样地的土壤动物密度均高于还田2个月后的土壤密度。还田2个月后, 各样地的平均密度排序为T10>T5>T20>对照, 各样地间差异不显著(F=0.105, P=0.956)。还田5个月后, 各样地的平均密度排序为T20>T10>T5>对照, 样地间差异极显著(F=6.702, P=0.003);其中, T20样地与对照、T5样地的平均密度均呈极显著差异(P<0.01), T10与T5样地的平均密度均呈极显著差异(P<0.01), 与对照样地呈显著差异(P<0.05)。
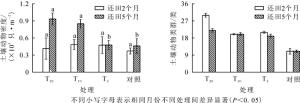
图 1 各生境土壤动物水平分布变化
Figure 1. Each horizontal distribution of soil fauna habitat change
T20土壤动物的类群数均高于T10、T5和对照样地的土壤动物。统计分析显示(图 1B):还田2个月后样地间土壤动物类群数差异性极显著(F=9.312, P=0.001), 其中, T20样地和T10、T5、对照样地分别呈极显著差异(P<0.01);还田5个月后T10和T20样地的土壤动物类群数呈显著差异(F=1.299, P=0.011)。
-
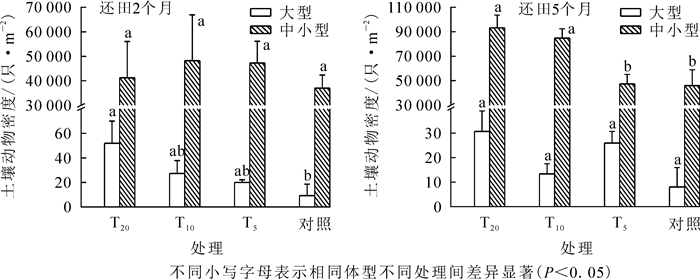
按照尹文英[12]对土壤动物体型的划分, 土壤动物可分为大型和中小型2类。由图 2可以看出:与对照相比, T20、T10、T5样地大型、中小型土壤动物的平均密度均较高。还田2个月后, 各处理大型土壤动物平均密度排序为T20>T10>T5>对照, 其中T20样地和对照差异显著(P<0.05), 其他样地间差异不显著; 中小型土壤动物以T10样地平均密度最高, 对照样地最低, 差异均不显著。还田5个月后, T20样地大型土壤动物密度最高, 占总密度的41.44%, 其余依次为T5(35.14%)、T10(18.02%)、对照(5.40%)。统计分析显示:还田5个月后, 各样地中小型土壤动物平均密度差异极显著(F=6.759, P=0.003), 对照分别与T10、T20呈显著差异(P<0.05), T5分别与T10、T20呈显著差异(P<0.05)。

图 2 各生境不同体型土壤动物分布变化
Figure 2. Each horizontal size distribution of soil fauna in the different levels of change
-
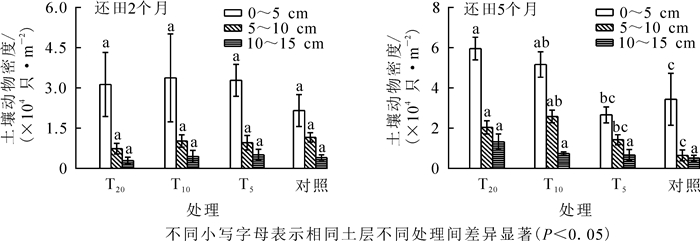
对不同稻草还田量下0~5、5~10和10~15 cm层的土壤动物密度进行比较, 结果显示: 0~5 cm层土壤动物平均密度最高, 随着土层加深土壤动物密度降低, 具有明显的表聚性(图 3A)。

图 3 各生境土壤动物垂直分布变化
Figure 3. Each vertical distribution of soil fauna habitat change
方差分析显示(图 3B):还田2个月后各样地所有土层土壤动物平均密度差异均不显著(P>0.05);还田5个月后各样地10~15 cm土层土壤动物平均密度差异不显著(F=1.556, P=0.237), 0~5 cm层(F=6.099, P=0.005)、5~10 cm层(F=5.942, P=0.006)差异显著。0~5 cm土层中, 对照与T20样地土壤动物密度差异显著(P<0.05), T5与T10、T20样地差异显著(P<0.05)。
-
稻草还田提高了土壤动物的多样性指数。还田2个月后各处理下土壤动物多样性指数(H′)、丰富度指数(D)均高于对照, 优势度指数(C)均低于对照, 均匀度指数T20和T5高于对照, 但T10低于对照。还田5个月后对照土壤动物多样性指数和均匀度指数最高, T10处理下土壤动物优势度指数最高, T20处理下土壤动物丰富度指数最高(表 2)。
表 2 稻草不同还田量下土壤动物群落的多样性特征
Table 2. Diversity characteristics of soil fauna in each habitat
处理 还田2个月 还田5个月 H' J C D H' J C D T20 1.15±0.12 a 0.47±0.05 a 0.44±0.12 a 1.06±0.10 a 0.68±0.15 a 0.29±0.05 ab 0.68±0.08 a 0.78±0.20 a T10 0.79±0.13 a 0.39±0.06 a 0.60±0.07 a 0.60±0.29 b 0.37±0.13 a 0.19±0.06 b 0.83±0.07 a 0.47±0.17 b T5 0.87±0.10 a 0.42±0.04 a 0.54±0.06 a 0.67±0.06 b 0.50±0.80 a 0.25±0.03 ab 0.78±0.04 a 0.66±0.10 ab ck 0.69±0.24 a 0.41±0.16 a 0.68±0.13 a 0.47±0.14 b 0.74±0.08 a 0.39±0.06 a 0.64±0.07 a 0.53±0.09 ab 说明:数据为平均值±标准误。同列不同小写字母表示处理间差异显著(P<0.05) 方差分析结果显示:还田2个月后, 各样地中土壤动物优势度指数(F=1.810, P=0.184)、多样性指数(F=2.049, P=0.145)、均匀性指数(F=0.270, P=0.846)的差异均不显著, 丰富度指数(F=9.764, P=0.001)T20样地与T10、T5、对照呈极显著差异(P<0.01)。还田5个月后, 各样地中土壤动物优势度指数(F=1.436, P=0.267)、多样性指数(F=1.795, P=0.186)差异均不显著, T20与T10样地的丰富度指数差异显著(P<0.05), 对照与T10样地的均匀度指数差异显著(P<0.05)。
-
目前, 稻草还田已经作为农业上培肥地力的一项技术而普遍利用。本研究发现:还田2个月和5个月后, 土壤动物类群数排序均为T20>T10>T5>对照, 土壤动物密度排序为T10>T5>T20>对照。整体来看, 土壤动物平均密度和类群数差异不显著, 原因可能有以下几点。首先, 实验在夏季进行, 土壤温度高、湿度大, 微生物活动旺盛, 分解快, 还田5个月后(9月)未发现明显稻草的分解物, 说明腐熟高峰已过, 土壤营养处于下降状态, 因此土壤动物个体数量和类群数降低。其次, 稻草覆盖的还田方式增加了农田表层腐殖质的土壤水分和有机质, 提高了土壤保水保墒能力, 喜湿土壤动物幼虫数量明显增加, 土壤动物类群数量也随之增加, 其中, 稻草还田处理的土壤线虫数量最高达到ck处理的2.5倍; 再次, 单位面积上稻草还田较多, 则分解后养分较多, 有利于创造适宜土壤动物生存繁衍的环境[19-21]。相比于稻草还田处理, 对照土壤贫瘠, 土壤肥力较差, 因此土壤动物类群数和土壤动物密度都最低[22]。这与在楠木人工林凋落物和草本层对土壤动物群落的影响中得到相似的结论[23]。
各样地土壤动物密度剖面及类群分布具明显的表聚性特征, 2次采样所采集到的大型、中小型土壤动物的数量有一定的差异, 随着季节的不同土壤动物的多样性也各不相同。还田2个月和还田5个月, 不同量稻草还田对土壤动物多样性的影响不同, 具体来说, 土壤动物数量与类群在不同还田量稻草处理下差异明显, 稻草还田量大的样地土壤动物数量与类群明显多于稻草还田量少的样地。稻草还田2个月后各样地上土壤动物多样性指数、均匀度指数和丰富度指数均高于还田5个月的样地, 以线虫纲密度增加最为明显, 可能是因为稻草还田后线虫等中小型土壤动物对土壤环境敏感性强、响应明显; 优势度指数则低于还田5个月后, 与此同时对照组不同月份各指数差异变化不大, 说明稻草还田后, 样地的土壤动物种群更加丰富, 土壤的生态环境相对稳定良好, 一段时间后优势种群开始突出。
综上所述, 与未进行稻草还田处理的普通样地相比, 稻草还田显著增加了土壤动物密度和类群数, 影响了区域内土壤动物群落结构; 稻草还田通过改变土壤理化性质, 促进了土壤动物的多样性和生态系统的稳定性, 提高了作物质量和产量[22, 24]。基于对生态系统稳定性及生产效率等因素的考虑, 建议农业生产上稻草切碎后覆盖还田选择0.8 kg·m-2还田量。本次研究历时较短, 仅研究了稻草覆盖还田对土壤动物群落的影响, 今后应继续对稻草还田腐烂程度等影响因素的土壤动物动态变化特征进行监测研究, 为农业生产提供更加科学的依据。
Effects of different rice straw returning quantities on soil fauna community structure
-
摘要:
目的 研究稻草不同还田量下土壤动物群落结构特征。 方法 在5 m×5 m的样方内,以未进行稻草还田的处理为对照(ck),采用手拣法和干、湿漏斗分离法,调查稻草还田后2个月和稻草还田后5个月,不同还田量[0.8(T20)、0.4(T10)、0.2(T5)和0 kg·m-2(ck)]处理下耕地土壤动物群落特征。 结果 试验共捕获土壤动物1 194只,隶属于3门11纲19目44科,平均密度4.45×105只·m-2;稻草还田处理后的样地土壤动物密度和类群数明显增加(P < 0.05),土壤动物类群数排序为T20(37)> T10(30)> T5(28)> ck(17)。土壤动物群落垂直分布明显,土壤动物密度随着土层加深而降低,具有明显的表聚性特征。稻草还田2个月后T20处理的土壤动物多样性指数、均匀度指数、丰富度指数均最高,稻草还田后5个月T10处理优势度指数最高。 结论 农业生产上稻草切碎后覆盖还田宜选择0.8 kg·m-2还田量。 Abstract:Objective The aim is to reveal the effects of different straw returning quantities on cropland soil fauna community. Method An experiment with four types of straw returning quantities[0.8(T20), 0.4(T10), 0.2(T5) kg·m-2 and no straw returning(ck)], were carried out in 5 m×5 m quadrats. Hand-picking method and dry and wet funnel separation method were used in the experiment. Result The total soil animals 1 194 were captured in this survey, belonging to 3 phyla, 11 classes, 19 orders and 44 families. The average density of soil animals was 4.45×105 ind·m-2. The density and variety of soil animals in the sample land increased significantly after straw returning(P < 0.05). The varieties were ranked as follows:T20(37) > T10(30) > T5(28) > ck(17). The vertical distribution of soil fauna community was obvious, and the density of soil fauna decreased with the deepening of soil layer, which had obvious surface aggregation. Two months after straw returning, T20 had the highest diversity index, evenness index and species richness. Five months after straw returning, T10 had the highest predominant index. Conclusion 0.8 kg·m-2 straw returning quantity is suggested. -
Key words:
- soil zoology /
- straw returning /
- soil fauna /
- community structure /
- diversity
-
图 2 各生境不同体型土壤动物分布变化
Figure 2 Each horizontal size distribution of soil fauna in the different levels of change
表 1 不同样地土壤动物群落密度统计
Table 1. Compositions of soil fauna community in the different plots
类群 还田2个月/(只·m-2) 还田5个月/(只·m-2) 总计/(只·m-2) 多度 T20 T10 T5 ck T20 T10 T5 ck 线虫纲Nematoda 60.13 52.13 38.43 82.41 77.79 88.00 87.51 73.90 72.16 +++ 懒甲螺科Nothridae 26.03 27.45 49.36 5.40 0.09 0 0 0 11.09 +++ 蚁科Formicidae 0.84 0.01 6.35 0.46 9.03 0 0.35 15.58 4.33 ++ 线蚓科Enchytraeidae 0 0 0 1.80 6.08 8.45 6.31 2.90 4.00 ++ 绥螨科Sejidae 4.24 3.62 3.00 4.50 1.07 1.57 1.40 2.54 2.41 ++ 丽甲螨科Liacaridae 0.61 0 0 0 5.28 0.69 2.28 3.98 1.95 ++ 双翅目幼虫Diptera larvae 0.20 13.12 0 1.80 0 0 0 0 1.59 ++ 棘䖴科Onychiuridae 0.81 0.35 1.23 2.25 0.27 0.10 0.70 0.36 0.62 + 跳虫科Poduridae 1.21 0.17 0.53 0 0 0.10 0.35 0.72 0.32 + 露尾甲科Nitidulidae 1.21 0 0 0 0 0.79 0 0 0.26 + 白蚁科Termitidae 2.42 0 0 0 0 0 0 0 0.22 + 蝴蛛目Araneae 0.01 1.04 0.18 0 0 0.20 0 0 0.17 + 叶蝉科Cicadellidae 0 0.69 0.18 0.90 0 0 0 0 0.17 + 蚁甲亚科Pselaphidae 0 0 0.18 0.45 0 0 0.53 0 0.11 + 叶甲科Chrysomelidae 0.20 0.52 0.18 0 0 0 0 0 0.10 + 幺蚰科Scutigerellidae 0 0.17 0 0 0.27 0.10 0 0 0.09 + 长角长䖴科Orchesellidae 0.81 0 0 0 0 0 0 0 0.07 + 蜈蚣目Scolopendromorpha 0.21 0.17 0 0 0 0 0.18 0 0.06 + 圆䖴科Sminthuridae 0.20 0.35 0 0 0 0 0 0 0.06 + 蜚蠊科Blattidae 0.01 0 0 0 0 0 0.35 0 0.04 + 拟步甲科Tenebrionidae 0.41 0 0.01 0 0 0 0 0 0.04 + 派盾螨科Parholaspididae 0.20 0 0.18 0 0 0 0 0 0.04 + 木螱科Kalotermitidae 0.20 0.17 0 0 0 0 0 0 0.04 + 等节䖴科Isotomidae 0 0 0 0 0.09 0 0 0 0.02 + 蟋蟀科Gryllidae 0 0 0.18 0 0 0 0 0 0.02 + 小蚓类Microdrile oligochaetes 0.02 0.03 0.02 0.01 0 0 0 0 0.01 + 蠼螋科Labiduridae 0 0 0 0 0.01 0 0 0 0 + 鼠妇科Porcellionidae 0.01 0 0 0 0 0 0.01 0 0 + 蝼蛄科Gryllotalpidae 0 0 0 0 0 0 0.01 0 0 + 步甲科Carabidae 0 0 0 0 0 0 0.01 0 0 + 姬马陆科Julidae 0.01 0 0 0 0 0 0 0 0 + 金龟甲科幼虫Scarabaeidae larvae 0 0 0 0 0 0 0 0 0 + 石蜈蚣目Lithobiomorpha 0 0 0 0 0 0 0 0 0 + 猎蝽科Reduviidae 0 0 0 0 0 0 0 0 0 + 隐翅虫科Staphylinidae 0 0 0 0 0 0 0 0 0 + 蛭纲Hirudinea 0 0 0 0 0 0 0 0 0 + 叩甲科Elateridae 0 0 0 0 0 0 0 0 0 + 天牛科Cerambycidae 0 0 0 0 0 0 0 0 0 + 瓢甲科Coccinellidae 0 0 0 0 0 0 0 0 0 + 蝗科Acrididae 0 0 0 0 0 0 0 0 0 + 角板盲蛛科Ceratolasmatidae 0 0 0 0 0 0 0 0 0 + 缘蝽科Coreidae 0 0 0 0 0 0 0 0 0 + 夜蛾科Noctuidae 0 0 0 0 0 0 0 0 0 + 锹甲科Lucanidae 0 0 0 0 0 0 0 0 0 + 合计 41 302.01 48 277.34 47 269.99 37 009.33 93 197.35 84 846.68 47 524.66 46 007.99 445 435.35 总类群数 30 20 21 11 22 20 19 11 44  下载: 导出CSV
下载: 导出CSV
表 2 稻草不同还田量下土壤动物群落的多样性特征
Table 2. Diversity characteristics of soil fauna in each habitat
处理 还田2个月 还田5个月 H' J C D H' J C D T20 1.15±0.12 a 0.47±0.05 a 0.44±0.12 a 1.06±0.10 a 0.68±0.15 a 0.29±0.05 ab 0.68±0.08 a 0.78±0.20 a T10 0.79±0.13 a 0.39±0.06 a 0.60±0.07 a 0.60±0.29 b 0.37±0.13 a 0.19±0.06 b 0.83±0.07 a 0.47±0.17 b T5 0.87±0.10 a 0.42±0.04 a 0.54±0.06 a 0.67±0.06 b 0.50±0.80 a 0.25±0.03 ab 0.78±0.04 a 0.66±0.10 ab ck 0.69±0.24 a 0.41±0.16 a 0.68±0.13 a 0.47±0.14 b 0.74±0.08 a 0.39±0.06 a 0.64±0.07 a 0.53±0.09 ab 说明:数据为平均值±标准误。同列不同小写字母表示处理间差异显著(P<0.05)
下载: 导出CSV
-
[1] 尹文英.中国土壤动物[M].北京:科学出版社, 2000:11-198. [2] 张桂玲.秸秆和生草覆盖对桃园土壤养分含量、微生物数量及土壤酶活性的影响[J].植物生态学报, 2011, 35(12):1236-1244. ZHANG Guiling. Effects of straw and living grass mulching on soil nutrients, soil microbial quantities and soil enzyme activities in a peach orchard[J]. Chin J Plant Ecol, 2011, 35(12):1236-1244. [3] 康轩, 黄景, 姜建初, 等.免耕稻草覆盖种植红薯对稻田土壤碳库及微生物数量的影响[J].广西农业科学, 2010, 41(3):236-239. KANG Xuan, HUANG Jing, JIANG Jianchu, et al. Effects of no-tillage and straw covering for sweet potato on paddy soil carbon pool and quantity of soil microorganism[J]. Guangxi Agric Sci, 2010, 41(3):236-239. [4] 吴建富, 曾研华, 潘晓华, 等.稻草还田方式对双季水稻产量和土壤碳库管理指数的影响[J].应用生态学报, 2013, 24(6):1572-1578. WU Jianfu, ZENG Yanhua, PAN Xiaohua, et al. Effects of rice straw returning mode on rice grain yield and soil carbon pool management index in double rice-cropping system[J]. Chin J Appl Ecol, 2013, 24(6):1572-1578. [5] 黄伟生, 黄道友, 汪立刚, 等.稻草覆盖对坡地红壤培肥及作物增产的效果[J].农业工程学报, 2006, 22(10):102-104. HUANG Weisheng, HUANG Daoyou, WANG Ligang, et al. Effects of straw mulching to slope red soil on fertility maintaining and crop yield[J]. Trans Chin Soc Agric Eng, 2006, 22(10):102-104. [6] 叶文培, 谢小立, 王凯荣, 等.不同时期秸秆还田对水稻生长发育及产量的影响[J].中国水稻科学, 2008, 22(1):65-70. YE Wenpei, XIE Xiaoli, WANG Kairong, et al. Effects of rice straw manuring in different periods on growth and yield of rice[J]. Chin J Rice Sci, 2008, 22(1):65-70. [7] 张水清, 钟旭华, 黄农荣, 等.稻草覆盖还田对水稻氮素吸收和氮肥利用率的影响[J].中国生态农业学报, 2010, 18(3):611-616. ZHANG Shuiqing, ZHONG Xuhua, HUANG Nongrong, et al. Effect of straw-mulch-incorporation on nitrogen uptake and N fertilizer use efficiency of rice (Oryza sativa L.)[J]. Chin J Eco-Agric, 2010, 18(3):611-616. [8] 张水清, 钟旭华, 黄农荣, 等.稻草覆盖还田对华南双季晚稻物质生产和产量的影响[J].中国水稻科学, 2011, 25(3):284-290. ZHANG Shuiqing, ZHONG Xuhua, HUANG Nongrong, et al. Effects of straw mulching on dry matter production and grain yield of double cropping late-season rice (Oryza sativa) in south China[J]. Chin J Rice Sci, 2011, 25(3):284-290. [9] 曾研华, 吴建富, 何虎, 等.机械化稻草全量还田下双季早稻生长发育、产量及品质的响应[J].江西农业大学学报, 2011, 33(5):840-844. ZENG Yanhua, WU Jianfu, HE Hu, et al. Effect of mechanized total returning of straw to field on growth, yield and quality of early rice[J]. Acta Agric Univ Jiangxi, 2011, 33(5):840-844. [10] 邢协加, 王振中, 张友梅, 等.杀虫双农药对土壤螨类和弹尾类影响的研究[J].湖南师范大学自然科学学报, 1997, 20(1):80-85. XING Xiejia, WANG Zhenzhong, ZHANG Youmei, et al. Studies on the effects of dimethypo pesticide on the soil nemaloda and collembola[J]. Acta Sci Nat Univ Norm Hunan, 1997, 20(1):80-85. [11] 王一华, 傅荣恕.辛硫磷农药对土壤螨类影响的研究[J].山东师范大学学报(自然科学版), 2003, 18(4):72-75. WANG Yihua, FU Rongshu. Simulating toxicity tests of phoxim pesticide to soil mites[J]. J Shandong Norm Univ Nat Sci, 2003, 18(4):72-75. [12] 尹文英.中国土壤动物检索图鉴[M].北京:科学出版社, 1998:236-239. [13] 李鸿兴, 隋敬之, 周世秀.昆虫分类检索[M].北京:农业出版社, 1987:1236-1244. [14] 钟觉民.幼虫分类学[M].北京:农业出版社, 1990:1527-1532. [15] 黄玉梅, 张健, 杨万勤.巨桉人工林中小型土壤动物类群分布规律[J].应用生态学报, 2006, 17(12):2327-2331. HUANG Yumei, ZHANG Jian, YANG Wanqin. Distribution pattern of meso-micro soil fauna in Eucalyptus grandis plantation[J]. Chin J Appl Ecol, 2006, 17(12):2327-2331. [16] 肖玖金, 黄晓丽, 朱万强, 等.猕猴桃园春季土壤动物群落结构特征[J].应用与环境生物学报, 2013, 19(3):454-458. XIAO Jiujin, HUANG Xiaoli, ZHU Wanqiang, et al. Community structure of soil fauna in kiwifruit orchards in spring[J]. Chin J Appl Environ Biol, 2013, 19(3):454-458. [17] 崔宁洁, 张丹桔, 刘洋, 等.马尾松人工林不同大小林窗植物多样性及其季节动态[J].植物生态学报, 2014, 38(5):477-490. CUI Ningjie, ZHANG Danju, LIU Yang, et al. Plant diversity and seasonal dynamics in forest gaps of varying sizes in Pinus massoniana plantations[J]. Chin J Plant Ecol, 2014, 38(5):477-490. [18] 申燕.茶园土壤动物群落结构特征及影响因素研究[D].成都: 四川农业大学, 2010. SHEN Yan. Study on Dynamic Change of the Characteristics of Soil Fauna Community Structure in Tea Plantation and Its Effect Factor[D]. Chengdu: Sichuan Agricultural University, 2010. [19] 董炜华, 殷秀琴, 顾卫, 等.农牧交错带不同土地类型土壤动物生态特征研究:以内蒙古卓资山为例[J].干旱区地理, 2008, 31(5):693-700. DONG Weihua, YIN Xiuqin, GU Wei, et al. Ecological characteristics of soil fauna in different soil types of agro-pasture ecotone:a case of Zhuozi Mountain, Inner Mongolia[J]. Arid Land Geogr, 2008, 31(5):693-700. [20] 刘迎新, 王凯荣, 谢小立, 等.稻草覆盖对亚热带红壤旱坡地玉米旱期生长的生理调节作用及其产量效应[J].生态与农村环境学报, 2007, 23(4):18-23, 56. LIU Yingxin, WANG Kairong, XIE Xiaoli, et al. Effect of straw mulching on physiological adjustment and output of maize growing on subtropical red soil slope-land in dry season[J]. J Ecol Rural Environ, 2007, 23(4):18-23, 56. [21] 赵睿宇, 李正才, 王斌, 等.毛竹林地表稻草覆盖后翻耕对土壤有机碳的影响[J].生态学杂志, 2017, 36(8):2118-2126. ZHAO Ruiyu, LI Zhengcai, WANG Bin, et al. Effects of straw mulching and scarification on soil labile organic carbon pool in a Phyllostachys edulis plantation[J]. Chin J Ecol, 2017, 36(8):2118-2126. [22] ZHU Xinyu, ZHU Bo. Diversity and abundance of soil fauna as influenced by long-term fertilization in cropland of purple soil, China[J]. Soil Tillage Res, 2015, 146:39-46. [23] 苟丽琼, 肖玖金, 黄进平, 等.土壤动物群落对楠木人工林凋落物和草本层去除的初期响应[J]. 2017, 34(5): 895-906. GOU Liqiong, XIAO Jiujin, HUANG Jinping, et al. Soil fauna community after removal of litter and herb layers in an artificial Phoebe zhennan plantaion[J]. 2017, 34(5): 895-906. [24] 连旭, 隋玉柱, 武海涛, 等.秸秆还田对黑土农田土壤甲螨群落结构的影响[J].农业环境科学学报, 2017, 36(1):134-142. LIAN Xu, SUI Yuzhu, WU Haitao, et al. Effect of on-site recycling of straw on community structure of soil Oribatida in black soil farmland[J]. J Agro-Environ Sci, 2017, 36(1):134-142. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.2020.01.011
 点击查看大图
点击查看大图

图(3) / 表(2)
计量
- 文章访问数: 2277
- HTML全文浏览量: 694
- PDF下载量: 39
- 被引次数: 0
