







-
液氨预处理法(liquid ammonia treatment,LAT)在氨纤维爆破预处理法(ammonia fiber explosion, AFEX)的基础上改进得来,通过提高反应温度、去除爆破工艺,将氨由快速释放转化为慢速释放,从而降低预处理工艺操作难度,具有非常广阔的发展前景[1]。赵相君等[2]研究了液氨预处理法和双氧水预处理法对稻草(水稻Oryza sativa)酶解转化率的影响,发现双氧水预处理后的葡聚糖和木聚糖的酶解转化率仅为61.6%和47.8%,显著低于液氨预处理法(88.6%和79.3%)。OLADI等[3]和ZHAO等[4]利用响应面法对液氨预处理工艺(温度、氨添加量、反应时间等)进行了优化,当反应温度为170 ℃、氨与原材料质量比为5∶1、载水与原材料质量比为0.8~1.6∶1.0、反应时间为10 min时,芦竹Arundo donax葡聚糖和木聚糖的酶解转化率最高,达到了94.2%和84.4%,总糖回收率高达530.9 g·kg−1。现有文献主要集中于研究LAT预处理工艺对单种生物质原料酶解率的影响[5-7],而对多种生物质经预处理后化学结构的变化规律(如微观形貌结构、表面化学官能团、纤维素结晶度等)的研究还鲜有报道。中国是农业生产大国,农业生物质资源丰富[8]。小麦Triticum aestivum、高粱Sorghum bicolor和苜蓿Lotus corniculatus作为传统且非常有潜力的农作物,具有栽植面积广,产量可观的优势[9-11]。本研究以麦秸秆、苜蓿草、高粱秸秆及其混合物(质量比为1∶1∶1)为原料,在不同温度条件下开展LAT预处理试验,利用热重分析仪(TGA)、傅里叶变化红外光谱仪(FTIR)、X-射线衍射仪(XRD)、扫描电镜(SEM)等对原料预处理前后的物理化学特性进行表征分析,并利用高效液相色谱(HPLC)对4类原料中葡聚糖/木聚糖的酶解转化率进行对比,探究最理想的LAT预处理温度及预处理前后材性的变化,为LAT预处理工艺设计提供理论依据,也为农作物转化为生物燃料建立基础。
-
麦秸秆取自浙江省杭州市临安区清凉峰镇,苜蓿草和高粱秸秆取材于甘肃省白银市会宁县,混合物由麦秸秆、苜蓿草、高粱秸秆按质量比1∶1∶1混合而成。先将原料烘干至含水率低于15%,再粉碎成粒径为50~100目的粉末。用水分分析仪(MA35,赛多利斯科学仪器有限公司)测试实际含水率,测试后用密封袋密封低温(−20 ℃)保存。纤维素酶(NS50013)和β-葡萄糖苷酶(NS50010)由诺维信公司(中国)提供。
-
称取各生物质原料20 g,加去离子水,使固液质量比达到70%,搅拌均匀并平衡10 min后转移到高压反应釜(GCF-1 L20/350,大连自控设备厂)中,并对反应釜抽真空。将液氨注入称量小钢瓶,使氨与原材料的质量比为1∶1;加热小钢瓶至50 ℃,用以提升氨的压力,再将氨注入反应釜。控制每组样品的反应釜温度在70、90、110、130、150和170 ℃下各反应5 min,反应结束后快速冷却反应釜并释放氨气。取出原料,40 ℃真空烘箱中干燥24 h后测量质量和含水率,计算预处理后原料得率,再将处理后的原料放入−20 ℃的冰箱冷藏备用。
-
采用元素分析仪(Vario EL Ⅲ,德国 Elementary 公司) 测定预处理前后原料的元素组成;根据国家可再生能源实验室(NREL)标准中的LAP002方法[12]测定碳水化合物相对含量,根据LAP003方法[12]测定酸不溶性木质素(Klason木质素)相对含量;采用高效液相色谱(HPLC-1200,美国安捷伦科技有限公司)测定各种单糖质量分数,并计算葡聚糖(纤维素)、木聚糖、阿拉伯聚糖等组分的相对含量;根据GB/T 2677.3−1993《造纸原料灰分的测定》标准测定原料灰分测定原料灰分。
采用热重分析仪(TG209F1,德国耐驰仪器制造有限公司)分析预处理前后的原料的热失重规律。称取10 mg原料放入三氧化二铝坩埚中,以20 ℃·min−1升温速率升至600 ℃;载气为高纯氮气(99.999%),载气流量为40 mL·min−1,保护气流量为20 mL·min−1。采用傅里叶红外光谱仪(Nicolet 6700,美国尼高力仪器公司)分析预处理前后原料的表面官能团。称取1 mg样品粉末与溴化钾(质量比1∶200)混合研磨均匀并压片,在4 000~500 cm−1波长范围内扫描。采用冷场发射扫描电镜(SEM,SU8010,日本日立公司)观察预处理前后样品微观结构。采用 X 射线衍射仪(XRD-6000,日本岛津公司)分析预处理前后原料的结晶度进行。称取200 mg粉末样品,压片处理后放入样品室,扫描范围为5°~40°,步长为0.05°,扫描速度为5°·min−1。根据公式计算纤维素结晶度ICr=(I002−Iam)/I002×100%。其中,I002为结晶峰的衍射强度,Iam为无定形区域衍射强度。
-
根据NREL标准中的LAP009[12]规程进行酶水解实验。称取150 mg葡聚糖当量的原料,用pH 4.8的蒸馏水-柠檬酸钠缓冲溶液溶解得到15 mL酶解液,加入抗生素(四环素和环己酰亚胺)防止菌落滋生。预混合1 h,添加纤维素酶和β-葡萄糖苷酶,放入摇床。酶解温度为50 ℃,摇床转速为150 r·min−1。
酶解24和72 h时取样,酶解液摇匀后用移液枪量取1 mL样品注入离心管,100 ℃下放置20 min,使酶蛋白变性完全失效,−20 ℃下冷却5 min,15 000 r·min−1转速下离心5 min,经0.22 μm聚醚砜膜过滤后注入HPLC分析瓶中,测定酶解液中的单糖相对含量。HPLC采用示差检测器,色谱柱(HPX-87H Ion Exclusion Column,Bio-Rad公司)条件:进样量为15 μL,每个样品运行20 min,柱温为50 ℃,流动相为0.005 mol·L−1稀硫酸,流速为0.5 mL·min−1。
用实际获得的单糖与原料中聚糖理论上可转换的同类单糖之比表示酶解率。计算其中葡聚糖和木聚糖的酶解率d=g×15/m×c×k×1 000×100%。其中:g为HPLC测得的酶解液中葡萄糖或木糖的质量浓度(g·L−1);m为加入原料的干基质量(g);c为原料成分分析(酸解法)葡聚糖或木聚糖相对含量(%);k为葡聚糖或木聚糖分别转化为葡萄糖或木糖的系数,取0.90和0.88。总糖回收量定义为1 000 g干基材料经LAT预处理72 h后酶解所释放的葡聚糖和木聚糖总质量之和。
-
由表1可知:预处理前高粱秸秆中葡聚糖、木聚糖以及阿拉伯聚糖的总相对含量(58.88%)高于麦秸秆和苜蓿草,而Klason木质素和灰分的总相对含量(17.20%)低于麦秸秆和苜蓿草,表明高粱秸秆中能有效转化为乙醇的多糖组分更多,其水解制备乙醇的潜力更大。经LAT预处理,3种原料的固体得率均出现少量下降;以高粱秸秆为例,葡聚糖、木聚糖以及阿拉伯聚糖相对含量分别从37.90%、17.63%和3.35%下降至33.72%、14.69%和3.28%,木质素从13.53%下降至10.53%;分析原因可能是液氨预处理过程中产生的诱导效应使木质素中的α-醚键发生断裂,引起木质素大分子结构部分断裂,木质素苯环上的甲基芳基醚键在氢氧根(OH−)作用下断裂,发生脱甲基反应,木质素的亲水性增强[13],使得部分小分子组分从生物质中脱落,溶解于预处理反应溶液中;与此同时,多糖组分的水解转化率得到提高[9]。如表2所示:预处理前,苜蓿草中碳和氢元素相对含量(43.79%和7.22%)及热值(22.24 MJ·kg−1)最高,麦秸秆中氧元素相对含量(53.16%)最高;经LAT预处理,3种原料中的碳元素相对含量都有所增加,而氢和氧相对含量下降,表明LAT预处理使得生物质原料中少量含氧和氢的官能团和联接键出现断裂和脱落,如羟基(—OH)、β—O—4芳基醚键,造成生物质结构疏松,有利于提高酶解率[14]。此外,预处理后原料中的氮元素相对含量增加,以苜蓿草为例,氮元素相对含量增加了1.2倍,表明在LAT预处理过程中,液氨中的氮元素与生物质中的官能团发生了化学反应,形成了少量的乙酰基[15]。
表 1 LAT预处理前后原料的化学组分分析
Table 1. Content of chemical components in biomass before and after LAT
样品 葡聚糖/% 木聚糖/% 阿拉伯聚糖/% Klason木质素/% 灰分/% 固体得率/% 未处理的麦秸秆 35.54 19.06 3.19 18.92 13.83 100 LAT预处理的麦秸秆 32.86 17.99 3.19 15.34 11.59 99.84 未处理的苜蓿草 27.38 12.88 3.02 21.38 5.90 100 LAT预处理的苜蓿草 25.81 11.53 3.01 20.09 5.04 99.77 未处理的高粱秸秆 37.90 17.63 3.35 13.53 3.67 100 LAT预处理的高粱秸秆 33.72 14.69 3.28 10.35 2.71 98.99 未处理的混合物 33.58 17.23 3.44 16.70 7.89 100 LAT预处理的混合物 30.32 16.81 3.43 13.61 7.22 99.46 表 2 LAT预处理前后原料的元素分析
Table 2. Ultimate analysis of biomass before and after LAT
样品 碳/% 氢/% 氧/% 氮/% 硫/% 高位热值/(MJ·kg−1) 未处理的麦秸秆 39.06 6.63 53.16 0.79 0.36 20.33 LAT预处理的麦秸秆 39.07 6.49 52.69 1.49 0.27 20.14 未处理的苜蓿草 43.79 7.22 47.47 1.36 0.16 22.24 LAT预处理的苜蓿草 43.87 6.90 46.83 3.10 0.29 21.29 未处理的高粱秸秆 43.53 7.09 48.21 1.01 0.17 22.29 LAT预处理的高粱秸秆 43.57 6.91 46.61 2.74 0.17 21.52 未处理的混合物 42.46 6.93 49.36 1.03 0.22 21.25 LAT预预处理的混合物 41.60 6.58 49.23 2.40 0.19 20.51 -
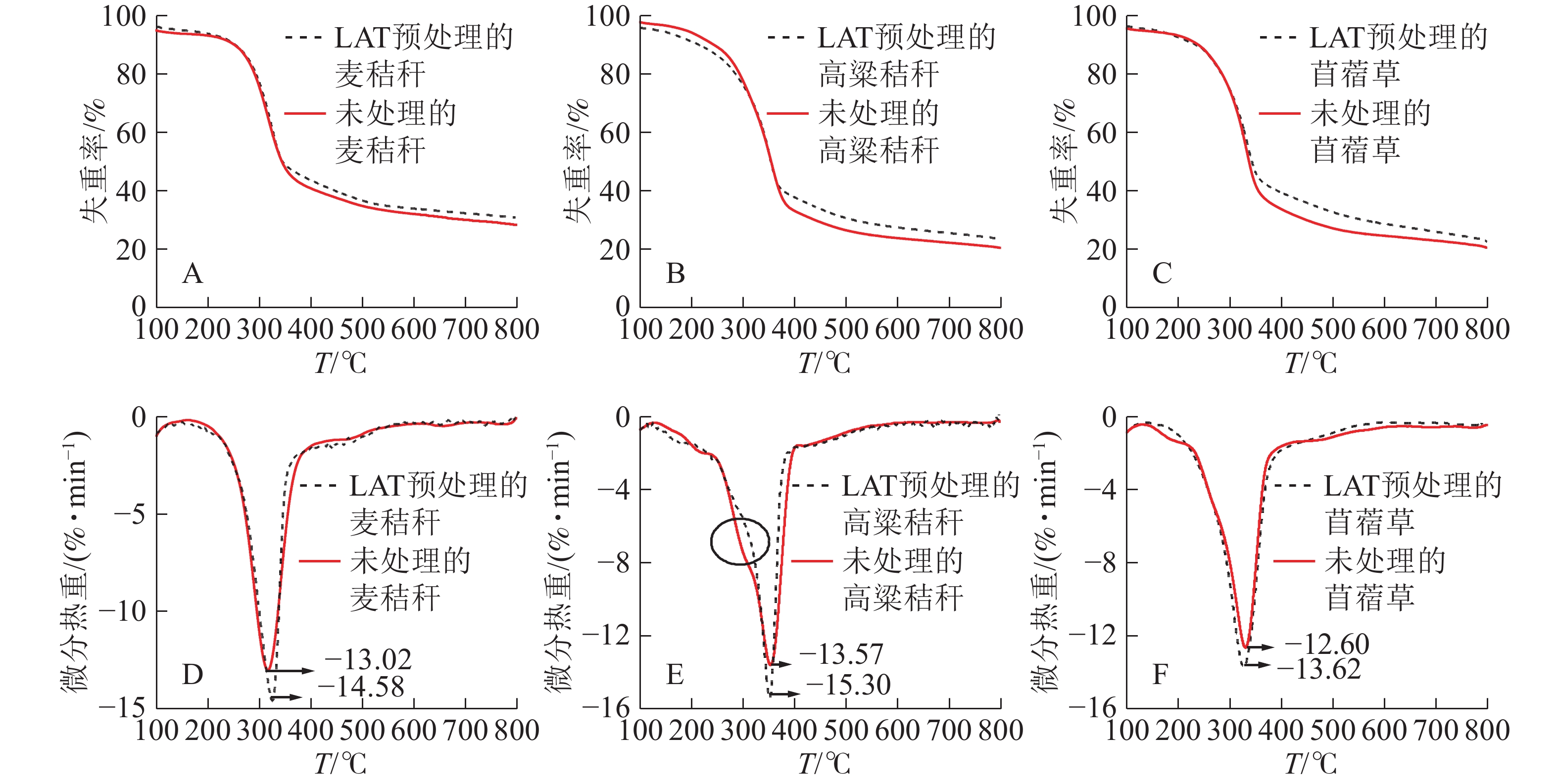
用20 ℃·min−1的升温速率对原料做热重分析,发现LAT预处理前后原料的热重(TG)曲线和微分热重(DTG)曲线不同。经LAT预处理的样品,热解后残余质量百分比增加,平均增幅为2%~3%(图1A、图1B和图1C)。由DTG曲线(图1D,图1E和图1F)可知:原料的热解过程可分为脱水、快速热解和炭化3个阶段[16-17]。30~200 ℃为脱水阶段,此时质量损失来源于游离水和结合水的脱除,由于样品已经经过烘干处理,因此原料失重率非常低[18-19]。当温度升至200~400 ℃时,生物质中的纤维素和半纤维素发生快速热解失重,预处理导致原料在此阶段的最大失重峰处的失重率下降;麦秸秆从13.02%·min−1下降至12.16%·min−1,高粱秸秆从13.57%·min−1下降至12.72%·min−1,苜蓿草从14.37%·min−1下降至13.62%·min−1。结合表1可知:LAT预处理造成生物质中的葡聚糖、木聚糖和阿拉伯糖相对含量下降,使得此阶段可降解物质相对含量下降,是本阶段失重率下降的原因[20]。此外,由图1E可知:高粱秸秆原料热解时存在1个热解失重肩峰,主要是由半纤维素热降解产生的;但LAT预处理后的高粱秸秆此肩峰消失,表明部分半纤维素(木聚糖和阿拉伯糖)已经在LAT预处理过程中移出,半纤维素的分解使得木质素-碳水化合物复合体(LCC)被破坏,提高了样品酶解可及度。400~800 ℃为炭化阶段,此阶段主要由木质素热降解造成,木质素经过慢速热解后逐步转化为生物质炭[21]。
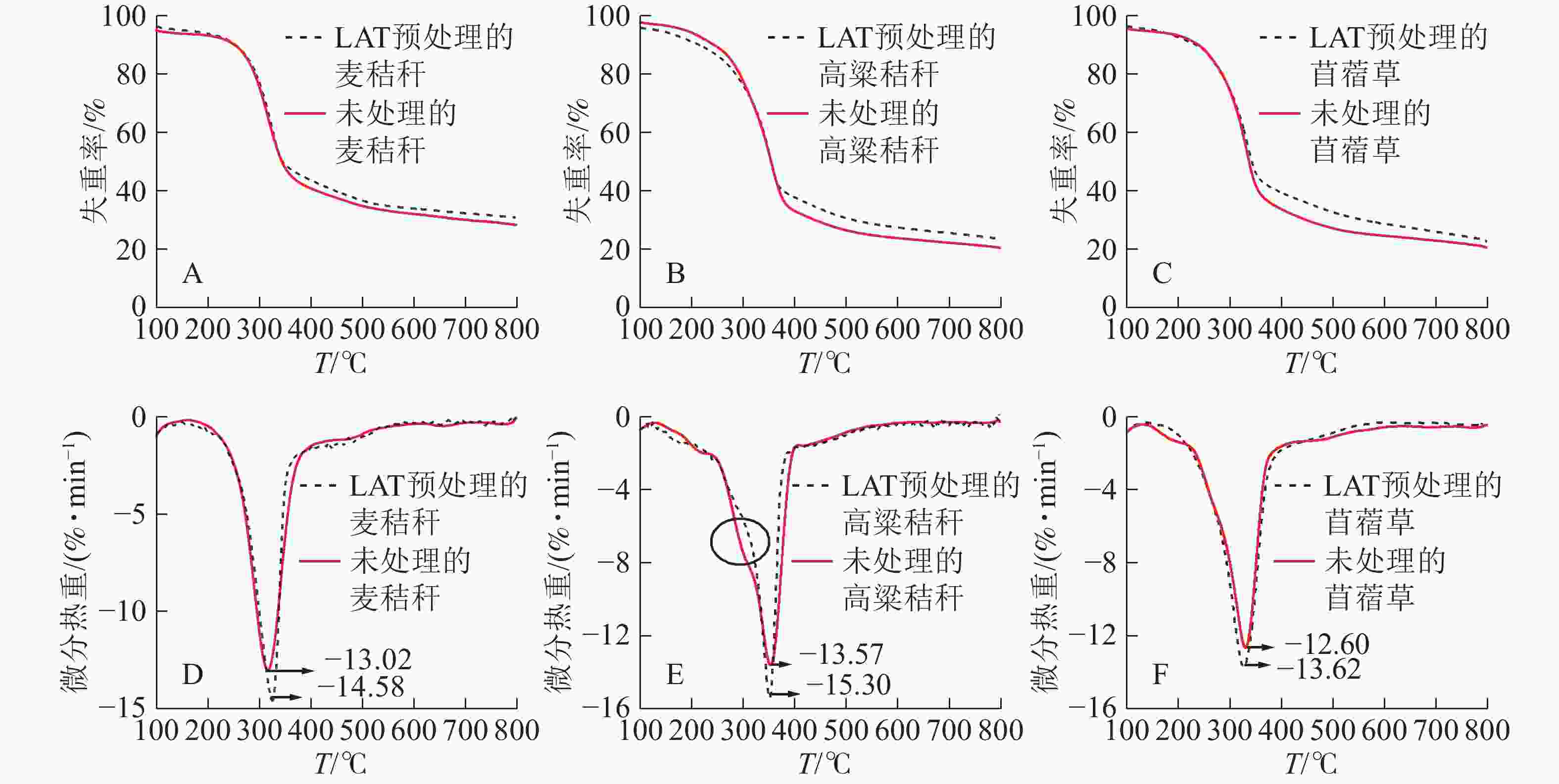
图 1 麦秸秆、高粱秸秆和苜蓿草在LAT预处理前后的热重和微分热重曲线
Figure 1. TG and DTG curves of wheat straw, sorghum straw and alfalfa before and after LAT
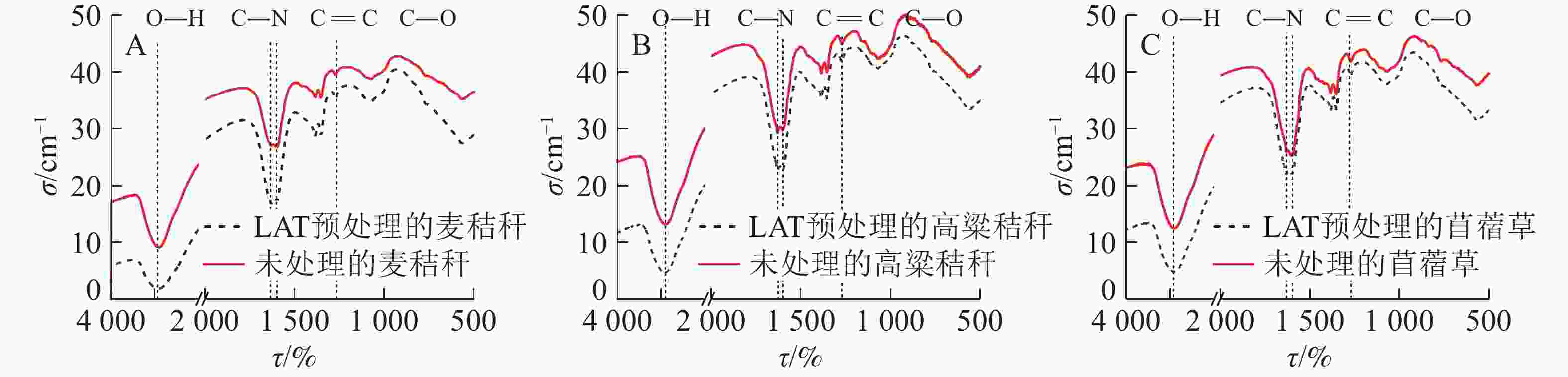
图 2 LAT预处理前后样品的红外光谱图
Figure 2. FTIR analysis of samples before and after LAT
-
由图2可知:LAT预处理前后样品的部分特征官能团的吸光度出现了轻微的变化。3 450 cm−1为羟基(—OH)的伸缩振动峰,主要来源于自由水以及纤维素、半纤维素和木质素化学结构上的羟基,经过LAT预处理后的样品在此处的吸光度强度出现一定程度下降,表明生物质中羟基含量下降,纤维素中的氢键被破坏,使得生物质结构更加疏松,与ZHAO等[8]的研究结论一致。1 631 cm−1处为碳—氮(C—N)伸缩振动吸收峰,经过LAT预处理后的样品在此处的吸光度强度出现一定程度增加,说明氨水与样品中的3大组分(纤维素、半纤维素和木质素)发生了化学反应,氮元素以酰胺的形式存在于样品中,与元素分析结果一致。1 595 cm−1处为苯环上碳=碳(C=C)键振动吸收峰,是木质素中苯丙烷基本结构单元的特征吸收峰,与原料谱图对比,LAT预处理后峰强度减弱,且有分峰出现,说明预处理使木质素结构发生改变,部分木质素溶解于液氨并脱除出生物质,使得生物质化学结构紧密程度降低,从而降低酶解顽抗性。3种原料吸收峰从大到小依次为苜蓿草、麦秸秆、高粱秸秆,侧面证明木质素相对含量大小依次为苜蓿草、麦秸秆、高粱秸秆。1 268 cm−1处吸收峰来自于纤维素和半纤维素的碳—氧(C—O )伸缩振动,对比发现,LAT预处理后该峰略微增强,归因于木质素和半纤维素联结生成LCC复合体,存在大量醚键,在碱性条件下,醚键断裂,LCC复合体破坏生成苯丙烷结构单元和多糖类物质[15,22-23]。
-
由图3可知:LAT预处理前后样品的微观形貌存在差异。未处理的麦秸秆纤维形状规则,排列整齐,纤维表面存在着规则排列的硅质细胞(图3A),预处理后纤维排列疏松膨胀,孔隙度增加(图3D),有利于进一步酶解反应。经过LAT预处理的高粱秸秆纤维表面暴露程度增加(图3B和图3E),苜蓿草表面结构发生扭曲和破坏,纤维暴露并出现裂纹和气孔,表面粗糙度变大,多孔性增加(图3C和图3F)。由此认为:LAT预处理能有效提高样品的比表面积,在压力作用下,氨水渗透进细胞腔和胞间层,引起纤维素的溶胀和结晶体的相变,LCC复合体被破坏,生成大量活性基团[24]。

图 3 LAT预处理前后3种原料的表观形貌
Figure 3. Micrograph of samples before and after LAT pretreatment
-
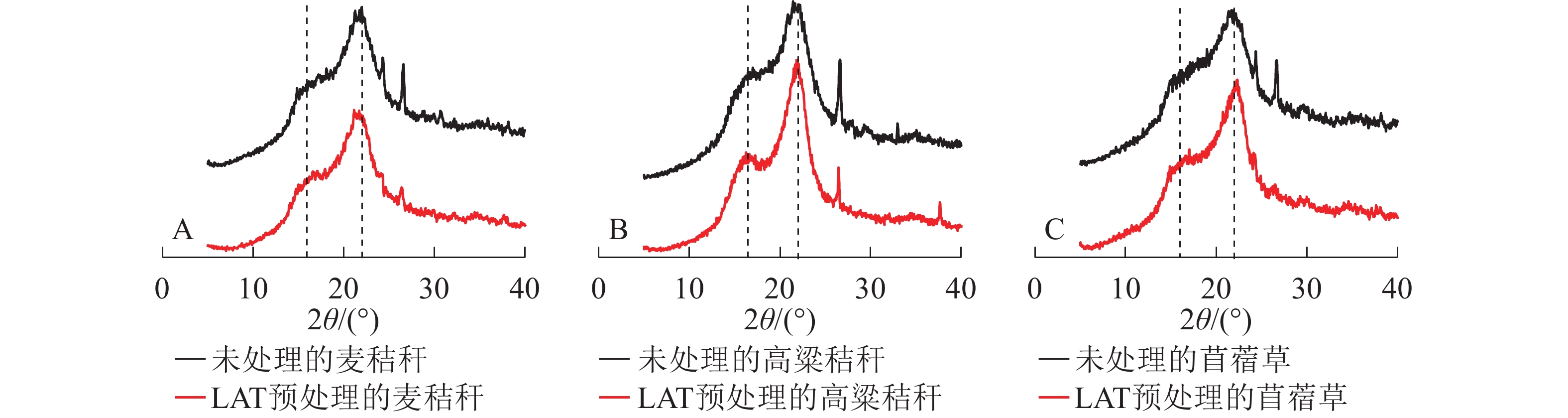
生物质的纤维素上存在结晶区和非结晶区,结晶度是用来表征纤维素结晶区含量的重要指标。结晶区域,纤维素链间以氢键紧密结合,纤维素大分子长链紧密排列,化学结构相对致密;非结晶区域,纤维素链随机排列、结构疏松,较有利于纤维素酶在其表面的键结。因此,纤维素的结晶度能够显著影响木质纤维素的酶解率。由图4可知:3种原料XRD谱图中主要存在2个峰,分别为16°和22°处纤维素的非结晶区和结晶区衍射峰;经过LAT预处理后纤维素结晶度下降,表明LAT预处理能破坏纤维素结晶区,氢键发生断裂,导致部分结晶区转化为无定形区,使得酶在纤维素化学结构上的可及度增加,从而提升酶解转化率[5]。
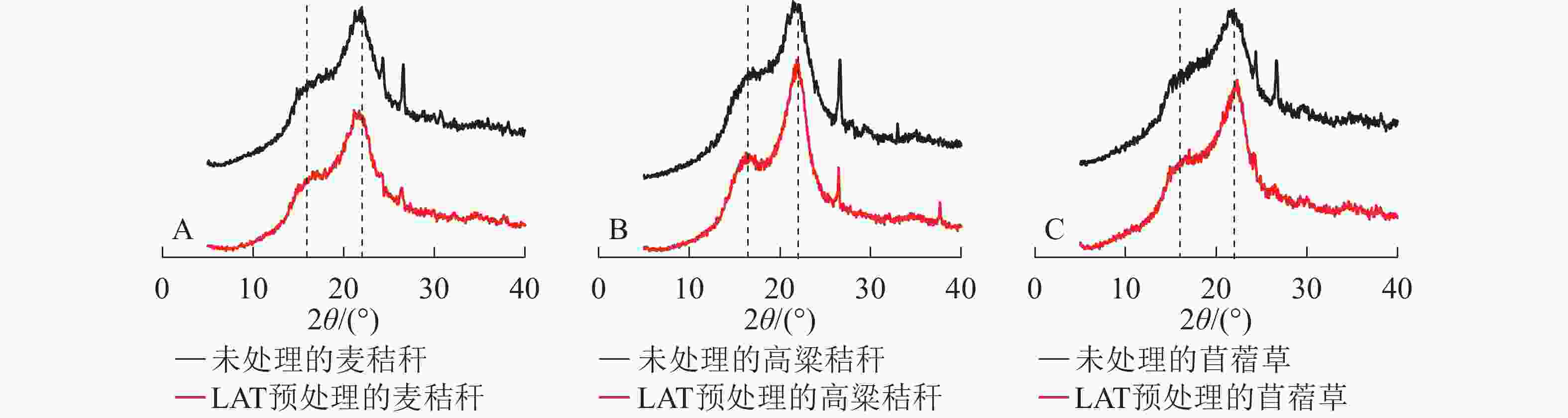
图 4 LAT预处理前后3种原料的XRD谱图
Figure 4. XRD analysis of samples before and after LAT
-
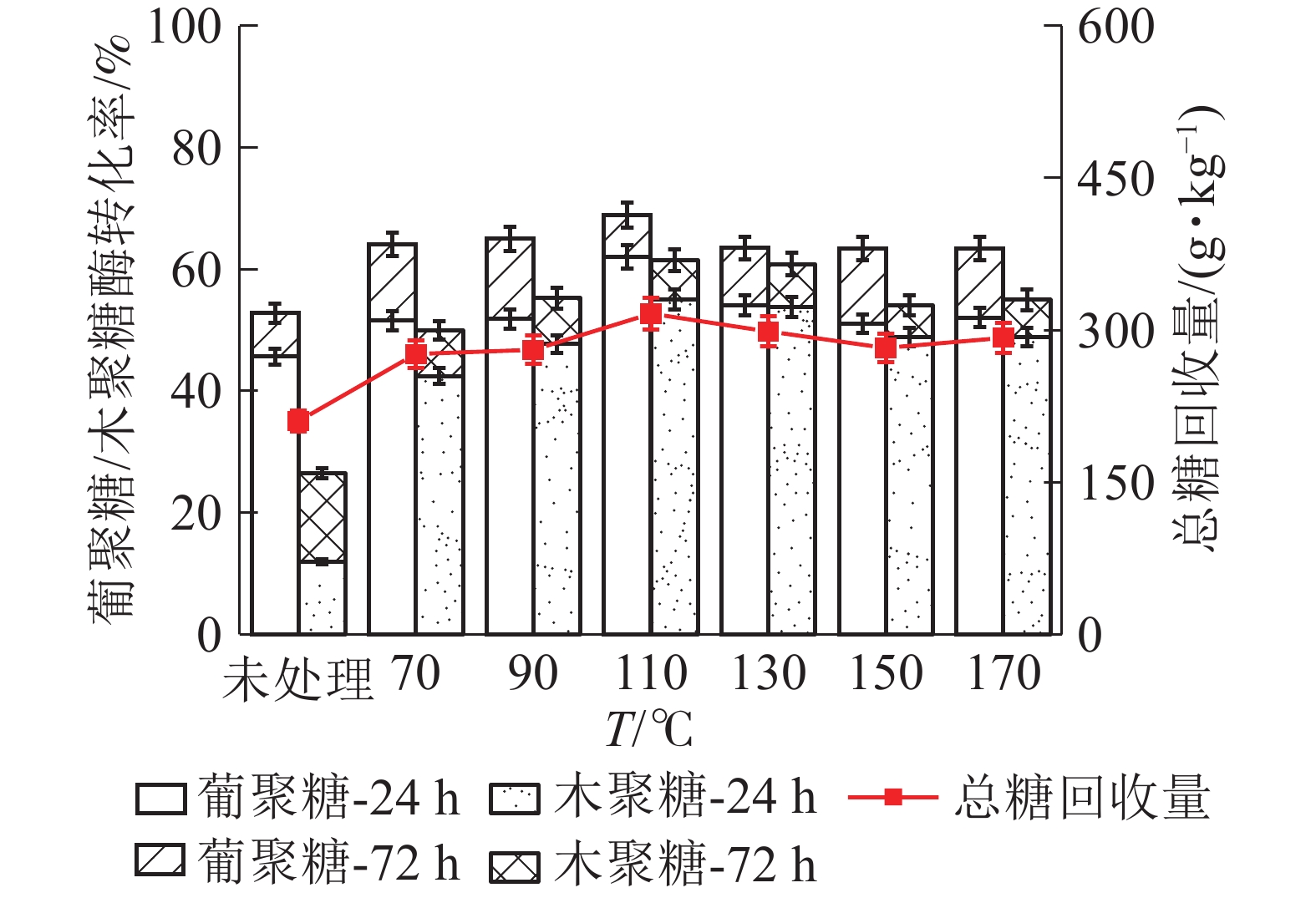
如图5所示:随着LAT预处理温度升高,葡聚糖的酶解转化率呈现先增加后减小的趋势,90 ℃时达到最大。在90 ℃下酶解24 h,葡聚糖酶解率为79.98%,是未经预处理原料的葡聚糖转化率(30.74%)的1.60倍;酶解至72 h时,进一步增至89.98%,达到最大值。木聚糖的酶解转化率也在90 ℃、72 h时达最高(79.03%),是未经处理原料的木聚糖转化率(14.87%)的4.34倍。认为90 ℃、酶解72 h为LAT预处理的最优条件。研究发现:HP预处理的最佳工艺条件下,麦秸秆总糖回收率为368.9 g;而利用优化条件下的LAT预处理,干基麦秸秆可转化单糖569.7 g·kg−1,是未经预处理原料产糖量(195.0 g)的2.92倍,是双氧水预处理的1.50倍,说明LAT预处理对麦秸秆酶解非常有效[11]。

图 5 LAT预处理温度对麦秸秆中葡聚糖/木聚糖酶解转化率的影响
Figure 5. Effect of temperatureof LAT on the glucan and xylan conversion of wheat straw
-
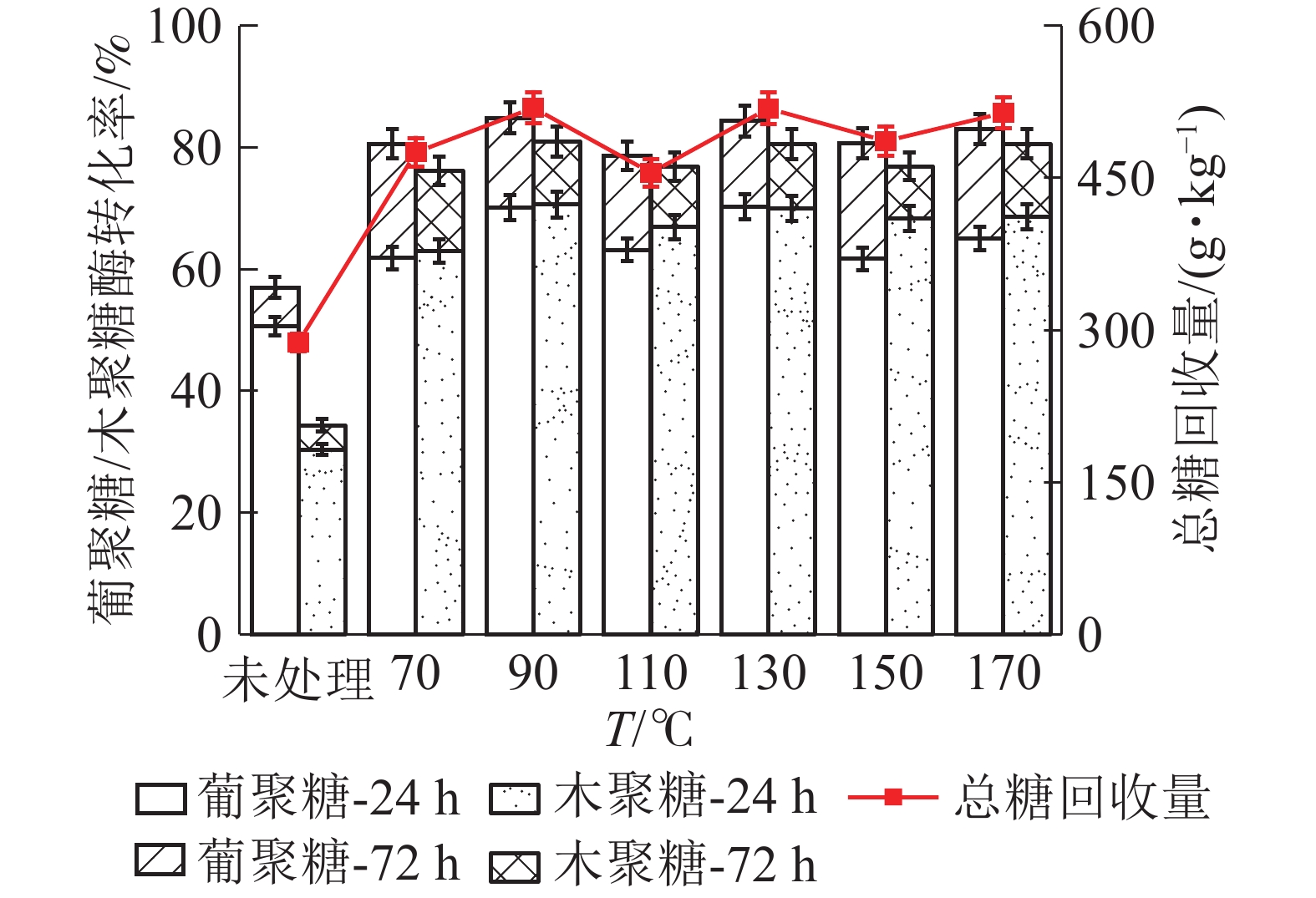
如图6所示:随着LAT预处理温度升高,苜蓿草葡聚糖的酶解转化率呈现先增加后减小趋势,110℃时达到最大值。在110 ℃下酶解24 h,葡聚糖酶解率为62.07%,比未经预处理原料的葡聚糖转化率(45.67%)提高了36.00%。继续酶解至72 h,葡聚糖酶解率最高可达到68.95%。木聚糖酶解转化率与葡聚糖大致相同,110 ℃下酶解72 h时,转化率达到最高值(68.22%),是未经处理原料的木聚糖转化率(11.96%)的4.70倍。110℃预处理后酶解72 h后,葡萄糖和木糖转化的单糖得率分别为244.0和102.6 g·kg−1,单糖总产量为316.3 g·kg−1。相比之下,低于麦秸秆的单糖产量(569.7 g·kg−1),推测原因是苜蓿草木质素相对含量较高,阻碍酶对纤维素和半纤维素的可及性,并与水解酶结合,导致酶水解速度降低,影响了酶解效果[8]。
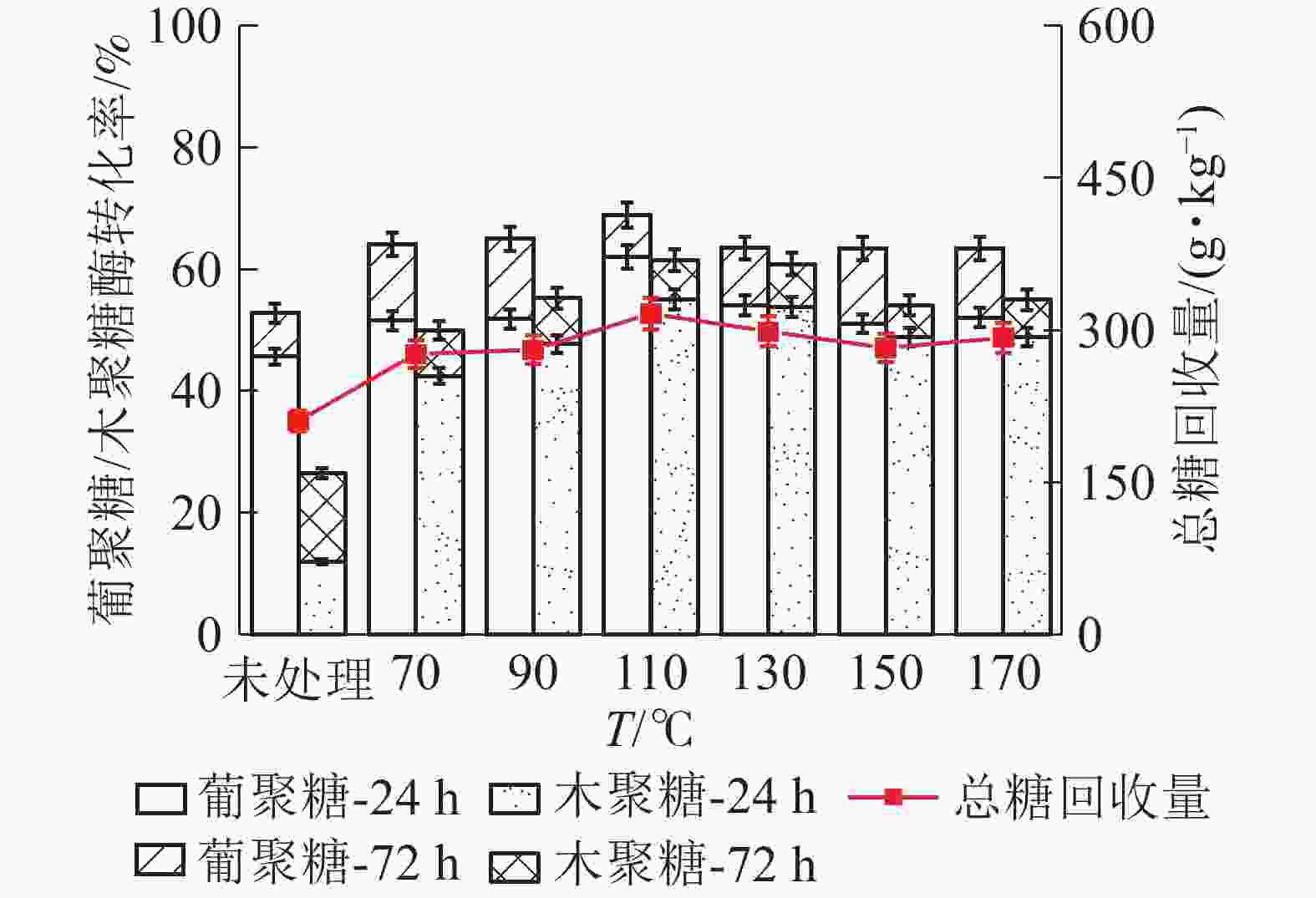
图 6 LAT预处理温度对苜蓿草中葡聚糖/木聚糖酶解转化率的影响
Figure 6. Effect of temperature of LAT on the glucan and xylan conversion of alfalfa
-
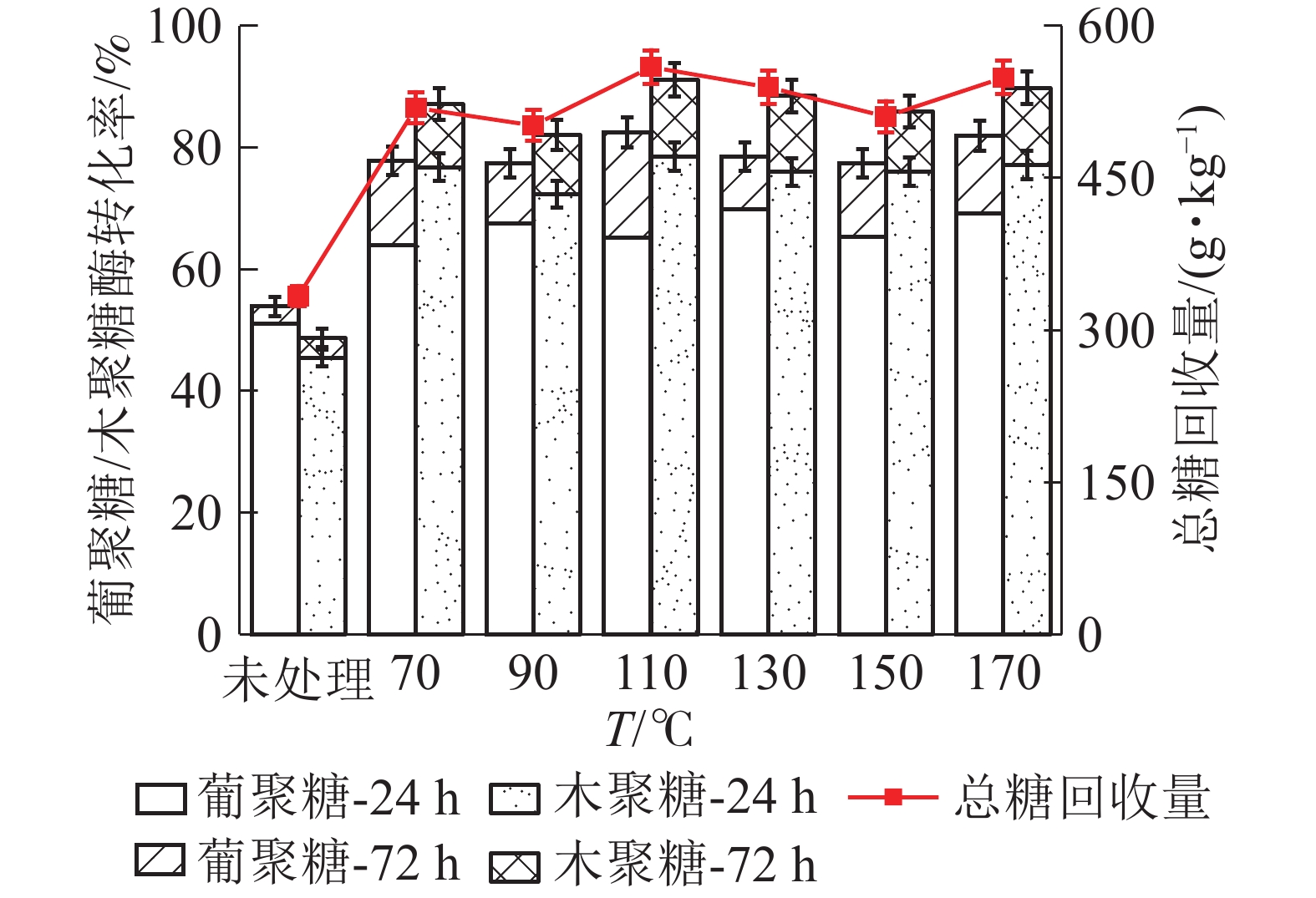
如图7所示:随着LAT预处理温度升高,高粱秸秆中葡聚糖酶解转化率呈现先增加后减小的趋势,110 ℃时转化率达到最大值。110 ℃下酶解24 h,葡聚糖酶解率为65.26%,比未经预处理原料(51.03%)增加了14.23%。继续酶解至72 h时,葡聚糖酶解率达到最大值(82.12%)。木聚糖酶解转化率与葡聚糖大致相同,但相同温度下大于葡聚糖。110 ℃酶解24 h,木聚糖转化为率为78.58%,比未处理样品(47.00%)提高了31.58%,酶解72 h时达到最大(91.21%)。预处理促进了原料中葡聚糖和木聚糖的酶解效果,高粱秸秆(干基)在110 ℃下酶解72 h,总糖回收量为559.5 g·kg−1,高于未处理原料(329.7 g·kg−1),说明液氨预处理提高了高粱秸秆单糖产量,LAT预处理有效。
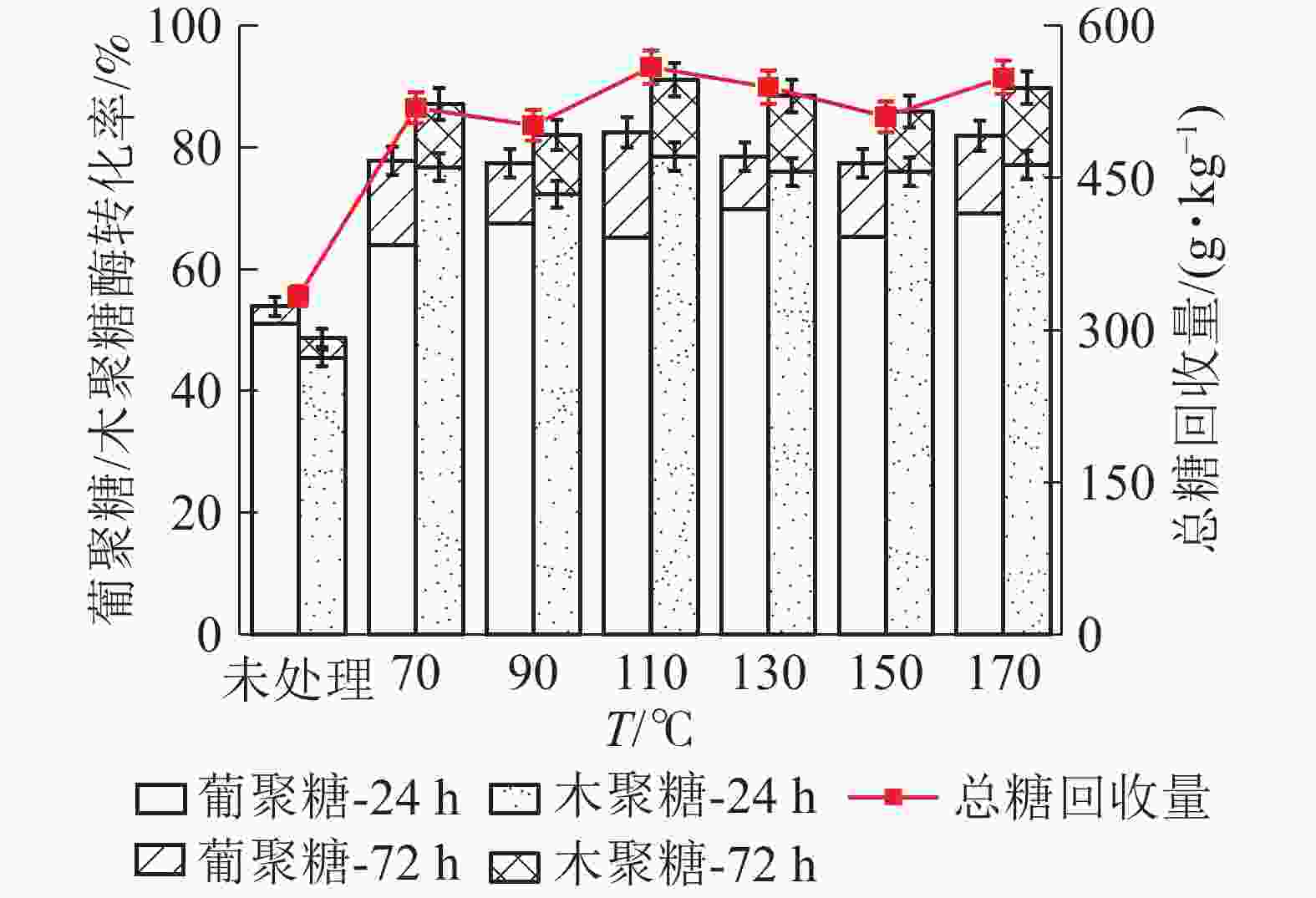
图 7 LAT预处理温度对高粱秸秆中葡聚糖/木聚糖酶解转化率的影响
Figure 7. Effect of temperature of LAT on the glucan and xylan conversion of sorghum straw
-
麦秸秆、苜蓿草、高粱秸秆按质量比1∶1∶1混合,混合物葡聚糖和木聚糖的酶解转化率随LAT预处理温度升高呈现先增加后减小的趋势(图8),在90 ℃时达到最大值;当温度由90℃升至170℃,混合物酶解转化率差异不明显。90℃下酶解24 h,混合物葡聚糖酶解率为70.19%,比未经预处理样品的葡聚糖酶解率(50.72%)提高了19.47%;继续酶解至72 h,葡聚糖酶解率为84.90%,比未处理样品提高了34.18%。木聚糖酶解转化率与葡聚糖大致相同,在90℃下酶解24 h,混合物木聚糖转化为率70.68%,比未经预处理样品的木聚糖转化率(30.39%)提高了40.29%,继续酶解至72 h,木聚糖转化率为81.02%,比未处理样品提高了50.63%。
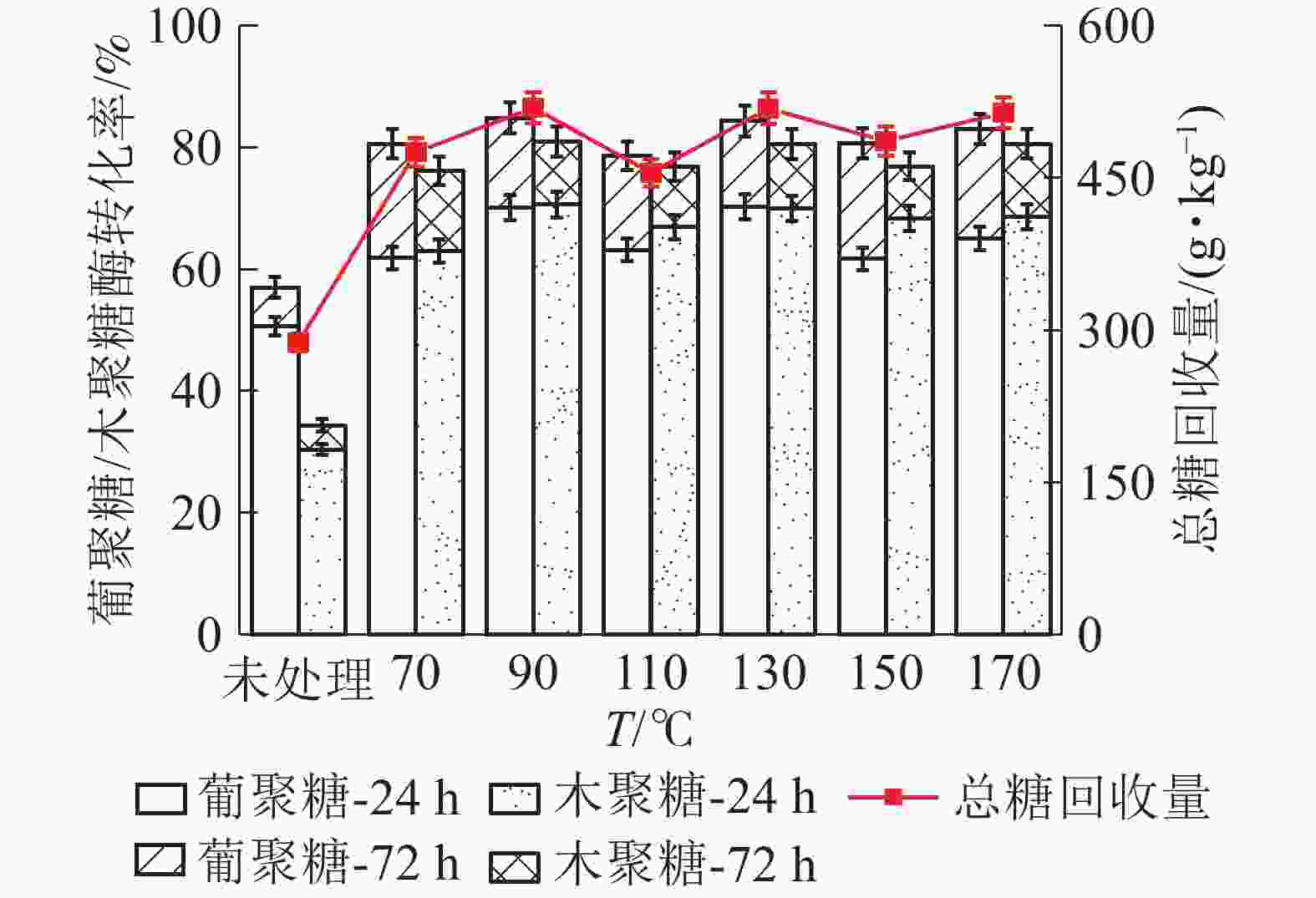
图 8 LAT预处理温度对混合物中葡聚糖/木聚糖酶解转化率的影响
Figure 8. Effect of temperature of LAT on the glucan and xylan conversion of mixture
-
采用LAT法对麦秸秆、苜蓿草、高粱秸秆以及三者混合物(质量比1∶1∶1)进行预处理,研究预处理温度与葡聚糖和木聚糖酶解转化率的关系,得到结论如下:①LAT预处理对生物质原料的化学结构影响显著。LAT预处理后,原料化学组分中葡聚糖、木聚糖和阿拉伯糖的相对含量下降;元素组分中的氧和氢相对含量下降,碳和氮相对含量上升;结晶度小幅下降,生物质表面孔隙结构增强,使得酶在生物质化学结构上的可及度增加。②麦秸秆和混合物中葡聚糖和木聚糖的酶解率在LAT预处理温度为90 ℃时达到最大值,苜蓿草和高粱秸秆在110 ℃时达到最大值。随着酶解时间延长,4种原料葡聚糖和木聚糖的酶解率均增加;葡聚糖的最大酶解率从大到小依次为麦秸秆、混合物、高粱秸秆、苜蓿草,木聚糖的最大酶解率从大到小依次为高粱秸秆、麦秸秆、混合物、苜蓿草。
Effects of LAT on chemical structure and enzymatic hydrolysis of crop straw
-
摘要:
目的 探索液氨预处理(liquid ammonia treatment,LAT)对生物质原料水解顽抗性和纤维素类生物质酶解效率的影响。 方法 采用LAT法对小麦Triticum aestivum秸秆(以下称麦秸秆)、高粱Sorghum bicolor秸秆、苜蓿Lotus corniculatus草及三者混合物(质量比为1∶1∶1)进行预处理,利用热重分析仪、傅里叶变化红外光谱仪、X-射线衍射仪和扫描电镜等对其预处理前后的化学结构变化进行表征,研究预处理温度和酶解时间对4种原料中葡聚糖和木聚糖的酶解转化率的影响。 结果 LAT预处理对生物质原料的化学结构影响显著。经LAT预处理后,葡聚糖、木聚糖和阿拉伯糖等化学组分的相对含量降低;氧(O)和氢(H)元素的相对含量降低,部分含氢(H)、氧(O)元素的官能团发生脱落;结晶度出现小幅下降,生物质表面孔隙结构增多,酶在生物质化学结构上的可及度增加。麦秸秆和混合物的最佳预处理温度为90 ℃,苜蓿草和高粱秸秆的最佳预处理温度为110 ℃;随酶解时间延长,4种原料中葡聚糖和木聚糖的酶解率都增加;葡聚糖的最大酶解率从大到小为麦秸秆、混合物、高粱秸秆、苜蓿草,木聚糖的最大酶解率从大到小依次为高粱秸秆、麦秸秆、混合物、苜蓿草。 结论 LAT预处理可以提高木质纤维素生物质尤其是麦秸秆和高粱秸秆的酶解效率。图8表2参24 Abstract:Objective This study aims to investigate the effects of liquid ammonia treatment (LAT) pretreatment on the hydrolysis resistance of biomass and the enzymatic hydrolysis rate of lignocellulosic biomass. Method Four different types of lignocelluloses biomass, namely wheat straw (Triticum aestivum), alfalfa (Lotus corniculatus), sorghum straw (Sorghum bicolor), and their mixture (mass ratio 1∶1∶1), were pretreated by LAT method, and the effect of LAT on their chemical structure changes was studied by using thermo gravimetric analysis (TGA), Fourier transform infrared spectrometer (FTIR), X-ray diffractometer (XRD), and scanning electron microscope (SEM). Then, the effect of pretreatment temperature and enzymatic hydrolysis time on the enzymatic hydrolysis conversion rates of glucan and xylan in the four raw materials was investigated. Result LAT had a significant effect on the chemical structure of biomass materials. After this pretreatment, the relative content of glucan, xylan, and arabinan in four types of lignocelluloses biomass slightly decreased. The relative content of O and H decreased because some functional groups containing O and H dropped off. The crystallinity decreased slightly, while the surface pore structure significantly increased, and the availability of enzymes in the chemical structure of biomass increased. The optimum pretreatment temperature of wheat straw and mixture was 90 ℃, while that of alfalfa and sorghum straw was 110 ℃. The enzymatic hydrolysis rates of glucan and xylan increased with the increase of enzymatic hydrolysis time. Among the four types of lignocelluloses biomass, the highest enzymatic hydrolysis rate of glucan obtained at the optimal enzymatic hydrolysis condition was wheat straw, followed by the mixture, sorghum straw, and alfalfa. The enzymatic hydrolysis rate of xylan ranging from large to small was sorghum straw, wheat straw, the mixture, and alfalfa. Conclusion LAT can improve the enzymatic hydrolysis efficiency of lignocellulosic biomass, especially that of wheat straw and sorghum straw. [Ch, 8 fig. 2 tab. 24 ref.] -
Key words:
- wheat straw /
- alfalfa /
- sorghum straw /
- liquid ammonia treatment /
- enzymatic hydrolysis /
- cellulose ethanol
-
图 1 麦秸秆、高粱秸秆和苜蓿草在LAT预处理前后的热重和微分热重曲线
Figure 1 TG and DTG curves of wheat straw, sorghum straw and alfalfa before and after LAT
图 3 LAT预处理前后3种原料的表观形貌
Figure 3 Micrograph of samples before and after LAT pretreatment
图 5 LAT预处理温度对麦秸秆中葡聚糖/木聚糖酶解转化率的影响
Figure 5 Effect of temperatureof LAT on the glucan and xylan conversion of wheat straw
图 6 LAT预处理温度对苜蓿草中葡聚糖/木聚糖酶解转化率的影响
Figure 6 Effect of temperature of LAT on the glucan and xylan conversion of alfalfa
图 7 LAT预处理温度对高粱秸秆中葡聚糖/木聚糖酶解转化率的影响
Figure 7 Effect of temperature of LAT on the glucan and xylan conversion of sorghum straw
图 8 LAT预处理温度对混合物中葡聚糖/木聚糖酶解转化率的影响
Figure 8 Effect of temperature of LAT on the glucan and xylan conversion of mixture
表 1 LAT预处理前后原料的化学组分分析
Table 1. Content of chemical components in biomass before and after LAT
样品 葡聚糖/% 木聚糖/% 阿拉伯聚糖/% Klason木质素/% 灰分/% 固体得率/% 未处理的麦秸秆 35.54 19.06 3.19 18.92 13.83 100 LAT预处理的麦秸秆 32.86 17.99 3.19 15.34 11.59 99.84 未处理的苜蓿草 27.38 12.88 3.02 21.38 5.90 100 LAT预处理的苜蓿草 25.81 11.53 3.01 20.09 5.04 99.77 未处理的高粱秸秆 37.90 17.63 3.35 13.53 3.67 100 LAT预处理的高粱秸秆 33.72 14.69 3.28 10.35 2.71 98.99 未处理的混合物 33.58 17.23 3.44 16.70 7.89 100 LAT预处理的混合物 30.32 16.81 3.43 13.61 7.22 99.46  下载: 导出CSV
下载: 导出CSV
表 2 LAT预处理前后原料的元素分析
Table 2. Ultimate analysis of biomass before and after LAT
样品 碳/% 氢/% 氧/% 氮/% 硫/% 高位热值/(MJ·kg−1) 未处理的麦秸秆 39.06 6.63 53.16 0.79 0.36 20.33 LAT预处理的麦秸秆 39.07 6.49 52.69 1.49 0.27 20.14 未处理的苜蓿草 43.79 7.22 47.47 1.36 0.16 22.24 LAT预处理的苜蓿草 43.87 6.90 46.83 3.10 0.29 21.29 未处理的高粱秸秆 43.53 7.09 48.21 1.01 0.17 22.29 LAT预处理的高粱秸秆 43.57 6.91 46.61 2.74 0.17 21.52 未处理的混合物 42.46 6.93 49.36 1.03 0.22 21.25 LAT预预处理的混合物 41.60 6.58 49.23 2.40 0.19 20.51
下载: 导出CSV
-
[1] 彭何欢, 邵千钧, CHANDIO F A, 等. 液氨预处理工艺对五节芒酶解糖化效果的影响[J]. 农业机械学报, 2016, 47(7): 196 − 202. PENG Hehuan, SHAO Qianjun, CHANDIO F A, et al. Effects of liquid ammonia treatment on enzymatic hydrolysis to fermentable sugars of miscanthus floridulus [J]. Trans Chin Soc Agric Mach, 2016, 47(7): 196 − 202. [2] 赵相君, 李琮, 彭何欢, 等. 液氨和过氧化氢预处理对稻草酶解效果的影响机制[J]. 农业工程学报, 2019, 35(10): 221 − 226. ZHAO Xiangjun, LI Cong, PENG Hehuan, et al. Mechanism on effect of liquid ammonia and hydrogen peroxide pretreatment on rice straw enzymatic hydrolysis [J]. Trans Chin Soc Agric Eng, 2019, 35(10): 221 − 226. [3] OLADI S, AITA G M. Optimization of liquid ammonia pretreatment variables for maximum enzymatic hydrolysis yield of energy cane bagasse [J]. Ind Crops Prod, 2017, 103: 122 − 132. [4] ZHAO Chao, CAO Yan, MA Zhongqing, et al. Optimization of liquid ammonia pretreatment conditions for maximizing sugar release from giant reed (Arundo donax L.) [J]. Biomass Bioenergy, 2017, 98: 61 − 69. [5] 王霞, 陈迪嘉, 叶广英, 等. 我国非粮作物燃料乙醇技术与产业发展现状[J]. 新能源进展, 2014, 2(2): 89 − 93. WANG Xia, CHEN Dijia, YE Guangying, et al. Development of technology and industrialization of non-grain crops fuel ethanol [J]. Adv New Renewable Energy, 2014, 2(2): 89 − 93. [6] 刘丁宁, 何春霞, 薛娇, 等. 理化预处理麦秸改善其聚丙烯复合材料抗霉菌腐蚀力学性能[J]. 农业工程学报, 2016, 32(17): 309 − 314. LIU Dingning, HE Chunxia, XUE Jiao, et al. Physicochemical pretreatments of wheat straw improving fungus corrossion resistance and mechanical properties of wheat straw/polypropylene composites [J]. Trans Chin Soc Agric Eng, 2016, 32(17): 309 − 314. [7] 陈朝儒, 王智, 马强, 等. 甜高粱茎汁及茎渣同步糖化发酵工艺优化[J]. 农业工程学报, 2016, 32(3): 253 − 258. CHEN Chaoru, WANG Zhi, MA Qiang, et al. Optimization of ethanol production from bagasse and juice of sweet sorghum stem by simultaneous saccharification and fermentation [J]. Trans Chin Soc Agric Eng, 2016, 32(3): 253 − 258. [8] 王文信, 蔡世攀, 王刚. 黄淮海地区农户苜蓿种植行为影响因素分析[J]. 农业工程学报, 2015, 31(增刊 1): 284 − 290. WANG Wenxin, CAI Shipan, WANG Gang. Empirical analysis of factors affecting alfalfa plantation behavior of farmers in Huang-huai-hai region [J]. Trans Chin Soc Agric Eng, 2015, 31(suppl 1): 284 − 290. [9] 刘建军, 彭何欢, 赵相君, 等. 斑茅酶解转化可发酵单糖的液氨预处理及参数优化[J]. 生物工程学报, 2013, 29(3): 333 − 341. LIU Jianjun, PENG Hehuan, ZHAO Xiangjun, et al. Optimization of liquid ammonia treatment (LAT) parameters for enzymatic hydrolysis of Saccharum arundinaceum to fermentable sugars [J]. Chin J Biotechnol, 2013, 29(3): 333 − 341. [10] 赵超, 邵千钧, 曹艳, 等. 液氨过氧化氢联合预处理对玉米芯酶解的影响[J]. 农业机械学报, 2015(6): 193 − 200. ZHAO Chao, SHAO Qianjun, CAO Yan, et al. Effects of combined hydrogen peroxide and liquid ammonia treatment on enzymatic hydrolysis of corn cob [J]. Trans Chin Soc Agric Mach, 2015(6): 193 − 200. [11] MOUTHIER T M B, de RINK B, van ERVEN G, et al. Low liquid ammonia treatment of wheat straw increased enzymatic cell wall polysaccharide degradability and decreased residual hydroxycinnamic acids [J]. Bioresour Technol, 2019, 272: 288 − 299. [12] National Renewable Energy Laboratory. Standard Biomass Analytical Procedures (LAPs) [R]. NREL(National Renewable Energy Laboratory), 2010. [13] 陈冰炜, 阚玉娜, 袁诚, 等. 乙醇预处理对芦竹细胞壁的影响及荧光可视化分析[J]. 林业工程学报, 2019, 4(4): 59 − 65. CHEN Bingwei, KAN Yuna, YUAN Chen, et al. Effects of ethanol pretreatment on the cell wall of Arundo donax and fluorescence visualization analysis [J]. J For Eng, 2019, 4(4): 59 − 65. [14] NGUYEN T A D, KIM K R, HAN S J, et al. Pretreatment of rice straw with ammonia and ionic liquid for lignocellulose conversion to fermentable sugars [J]. Bioresour Technol, 2010, 101(19): 7432 − 7438. [15] ZHAO Chao, MA Zongqing, SHAO Qianjun, et al. Enzymatic hydrolysis and physiochemical characterization of corn leaf after H-AFEX pretreatment [J]. Energy Fuels, 2016, 30(2): 1154 − 1161. [16] 王浚浩, 张雨, 杨优优, 等. 微藻种类对其热解质量损失规律和产物及动力学的影响[J]. 农业工程学报, 2018, 34(19): 239 − 247. WANG Junhao, ZHANG Yu, YANG Youyou, et al. Weight-loss characteristics, components of bio-oil and kinetics during pyrolysis from different types of microalgae [J]. Trans Chin Soc Agric Eng, 2018, 34(19): 239 − 247. [17] MA Zhongqing, YANG Youyou, WU Youlong, et al. In-depth comparison of the physicochemical characteristics of bio-char derived from biomass pseudo components: hemicellulose, cellulose, and lignin [J]. J Anal Appl Pyrolysis, 2019, 140: 195 − 204. [18] 张雨, 王浚浩, 马中青, 等. 温度对竹材烘焙过程中气固液三相产物组成及特性的影响[J]. 农业工程学报, 2018, 34(18): 242 − 251. ZHANG Yu, WANG Junhao, MA Zhongqing, et al. Effects of torrefaction temperature on composition and characteristics ofgas-solid-liquid three-phase products in bamboo torrefaction process [J]. Trans Chin Soc Agric Eng, 2018, 34(18): 242 − 251. [19] CHEN Dengyu, WANG Yun, LIU Yixuan, et al. Comparative study on the pyrolysis behaviors of rice straw under differentwashing pretreatments of water, acid solution, and aqueous phase bio-oil byusing TG-FTIR and Py-GC/MS [J]. Fuel, 2019, 252: 1 − 9. [20] MA Zhongqing, SUN Qingfeng, YE Jiewang, et al. Study on the thermal degradation behaviors and kinetics of alkali lignin for production of phenolic-rich bio-oil using TGA-FTIR and Py-GC/MS [J]. J Anal Appl Pyrolysis, 2016, 117: 116 − 124. [21] VO T K, LY H V, LEE O K, et al. Pyrolysis characteristics and kinetics of microalgal Aurantiochytrium sp. KRS101 [J]. Energy, 2017, 118: 369 − 376. [22] 向松明, 谢益民, 杨海涛, 等. 纤维素前驱物6-13C标记示踪研究纤维素与木质素连接方式[J]. 林产化学与工业, 2014, 34(1): 37 − 42. XIANG Songming, XIE Yimin, YANG Haitao, et al. Analysis of the association between cellulose and lignin by carbon 13 tracer method with cellulose precursor [J]. Chem Ind For Prod, 2014, 34(1): 37 − 42. [23] 曹轩, 陈晓娟, 邢沙沙, 等. 甲醚芳醛类化合物的脱甲基研究[J]. 当代化工研究, 2012(11): 31 − 33. CAO Xuan, CHEN Xiaojuan, XING Shasha, et al. Study on demethylation of methoxy-substituted aromatic aldehydes compounds [J]. Modern Chem Res, 2012(11): 31 − 33. [24] CHUNDAWAT S P S, BELLESIA G, UPPUGNDLA N, et al. Restructuring the crystalline cellulose hydrogen bond network enhances its depolymerization rate [J]. J Am Chem Soc, 2011, 133(29): 11163 − 11174. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20190575
 点击查看大图
点击查看大图
计量
- 文章访问数: 2001
- HTML全文浏览量: 733
- PDF下载量: 36
- 被引次数: 0
