






-
物种在时空中具有共存机制,种间势必发生相互吸引、相互排斥等关联[1−2],这种种间关联与群落环境异质性、资源供给及竞争强弱等有显著关系[3−4]。群落的种间关联在一定程度上反映了种群消长变化、群落动态演替进程以及群落稳定性[5−7]。群落稳定性是群落适应异质环境以维持自身稳定性等综合能力的集中体现,对群落结构功能和生态系统健康状况具有重要指示作用[8]。目前种间关联研究主要从乔灌草群落立地条件[9−10]、群落类型[3−4]、围封[11]或采伐[12]等不同干扰强度[13]展开,对不同营林措施产生的种间关联研究还未有报道。
截至2020年中国高速公路已有16.1 万km,位居世界第1位,各国高速公路路域森林绿化也从最初的单一绿化转变为多结构、多功能的复合体生态绿化[14−15],形成纵横交错、相互联系的路域森林网络,以改善路域森林脆弱性和敏感性,恢复植物多样性,提高其生态服务效益[15−16]。目前,路域森林主要对生态破坏和植被恢复[17]、营林措施[16]、公路绿化[14]等方面进行研究。
林下植物是路域森林系统的重要功能结构[18],通过调控土壤养分、能量流动等对立地质量、立地生产力以及林分环境产生重要影响,从而影响地上植物的生长、分布、数量和结构[18−19]。林下灌草层是对生境变化最敏感的群落,是衡量路域森林生态健康状况的重要指标[20],因此,研究路域森林林下植物种间关联具有重要意义。加杨Populus canadensis具有早期速生、耐贫瘠与适应力强等特点[21],现存在结构单一、营林粗放、群落稳定性差等问题[22]。鉴于此,本研究对不同营林措施下路域加杨人工林林下植物种间及群落产生的内在影响进行研究,为加杨人工林的植物恢复和城市路域森林多样性保育提高提供参考依据。
-
成都第一绕城高速G4202 (30°72′~30°78′N,103°93′~104°11′E)位于四川成都市,全长84.96 km,平均高程为500 m。该区属亚热带湿润季风气候,气候温和、四季分明、湿度大,年平均气温为16 ℃,年平均降水量为896.8 mm,土壤肥沃,主要为冲积土和稻土。林地类型多为人工栽植的乔木林且无樵采。
加杨因其早期速生性且适应能力强,现作为重要的人工用材林,栽植面积已位居中国首位[22]。本研究路域杨树人工林于2009年集中统一营造,初植密度适中且无倒木和枯立木,林分密度差异小。乔木层以加杨为主,间或润楠Machilus nanmu、八角枫Alangium chinense等;灌木层主要为天竺桂Cinnamomum japonicum、朴树Celtis sinensis、构树Broussonetia papyrifera等;草本层主要为葎草Humulus scandens、蜘蛛抱蛋Aspidistra elatior、求米草Oplismenus undulatifolius等。
-
于2020年7月中旬对成都G4202路域进行全面踏查,调查区域立地条件相近,地势平坦,没有明显坡度,坡度差异小于4°。参考方精云等[23]的样地选取方法,采用典型样地法,选取林相整齐、林层丰富、具有代表性的加杨人工纯林样地,5种营林措施分别为:种植(UP)、翻耕(PT)、割灌(SW)、弃耕(AC)、对照(CG,未发生任何经营活动的样地)。每个样地面积为20 m×30 m,共20个样地,总面积为12 000 m2,测其林分密度、郁闭度、平均树高以及平均胸径(DBH),各样地基本情况如表1。对胸径≥3 cm的乔木层所有个体记录种名、胸径、树高、冠幅等指标;在每个样地沿对角线设置5 m×5 m的6个灌木样方和1 m×1 m的12个草本样方进行调查,灌木层和更新幼苗幼树(胸径<3 cm)、草本层分别统计种名、高度、株数、冠幅(盖度)等指标。
表 1 样地基本情况
Table 1. Basic situation of the plot
样地
编号林分密度/
(株·hm−2)郁闭度 平均树高/m 平均胸径/cm 营林
措施备注 1~4 537.50 0.69 16.49±0.43 b 23.79±0.70 b 种植 种植样地:在林冠下充分开展农、林业的双向复合经营模式,是最典型的营林措施。本研究区于2018年3月集中种植蜘蛛抱蛋Aspidistra elatior,套种密度为30 株·m−2 5~8 537.50 0.80 16.84±0.39 b 28.11±0.92 a 翻耕 翻耕样地:用有壁犁等农具使土壤结构疏松的耕作方法,以改善土壤条件,在每年的春冬季去除地表残茬和杂草,留茬12~15 cm 9~12 516.67 0.70 12.97±0.43 c 19.42±1.08 c 割灌 割灌抚育样地:通常在冬季距离道路较近处进行割灌作业,多在当年初夏(集中在5—6月)进行,且割除的灌丛茬高不超过5 cm 13~16 550.00 0.78 18.09±0.28 a 25.41±0.69 b 弃耕 弃耕样地:原进行农业经营的土地不再进行农耕作业而撂荒的土地,该样地为林下种植后弃耕样地 17~20 562.50 0.71 18.27±0.14 a 24.80±0.50 b 对照 对照样地:造林开始后未发生任何经营活动的样地 说明:不同字母表示营林措施间数据差异显著(P<0.05)。 -
根据需求采取重要值(IV)[24]、方差比率(VR)[25]、种间联结系数(AC)[26]以及Spearman种间秩相关系数[7]计算不同营林措施下加杨人工林林下植物优势种的种间关联各测度。统计样方内种群存在与否的二元数据矩阵,运用方差比率法来检验种间总体关联性,用统计量(W)检验物种间关联强弱。由于本研究为非连续性取样,因此χ2值用Yates的连续性校正公式检验种对关联情况[6],用AC、Spearman秩相关系数补充计算种间关联强弱程度,AC为[−1, 1],AC越接近1表示物种间正联结越强,AC越接近−1,表示物种间负联结越强,AC为0表示物种间完全独立。Spearman为[−1, 1],正值表示为正相关,负值则为负相关,0表示物种间没有关联,且值越接近1表示正联结越强,越接近−1则表示负联结越强。采用郑元润[27]改进后的M. Godron贡献定律法对群落的稳定性进行分析。
-
采用Excel对重要值等数据进行整理和计算,用R 4.1.2软件对VR、χ2、AC、Spearman秩相关系数等指标与M. Gordon稳定性进行分析和绘图。
-
不同营林措施下加杨人工林林下共有乔、灌、草植物209种,隶属85科158属。林下灌木植物共30科36属47种,以樟科Lauraceae、大戟科Euphorbiaceae、蔷薇科Rosaceae等为主;林下草本植物156种,隶属于52科117属,以菊科Asteraceae、荨麻科Urticaceae、蓼科Polygonaceae等植物为主。不同营林措施下草本物种数从大到小依次为翻耕、种植、割灌、对照、弃耕,翻耕样地中物种数最多,弃耕和对照样地差异较小;灌木物种数在割灌营林措施下最多,种植、翻耕、弃耕及对照则无明显差异;草本层物种科属种数量明显多于灌木层,表明5种营林措施下草本群落占据了优势地位。方差分析表明:各样地灌草层出现的科属种数在不同营林措施下差异均不显著(P>0.05),但是草本层物种数量均值从大到小依次为翻耕、割灌、种植、对照、弃耕,而灌木层物种数量均值除割灌最多外,其他营林措施下则无明显差异(图1)。
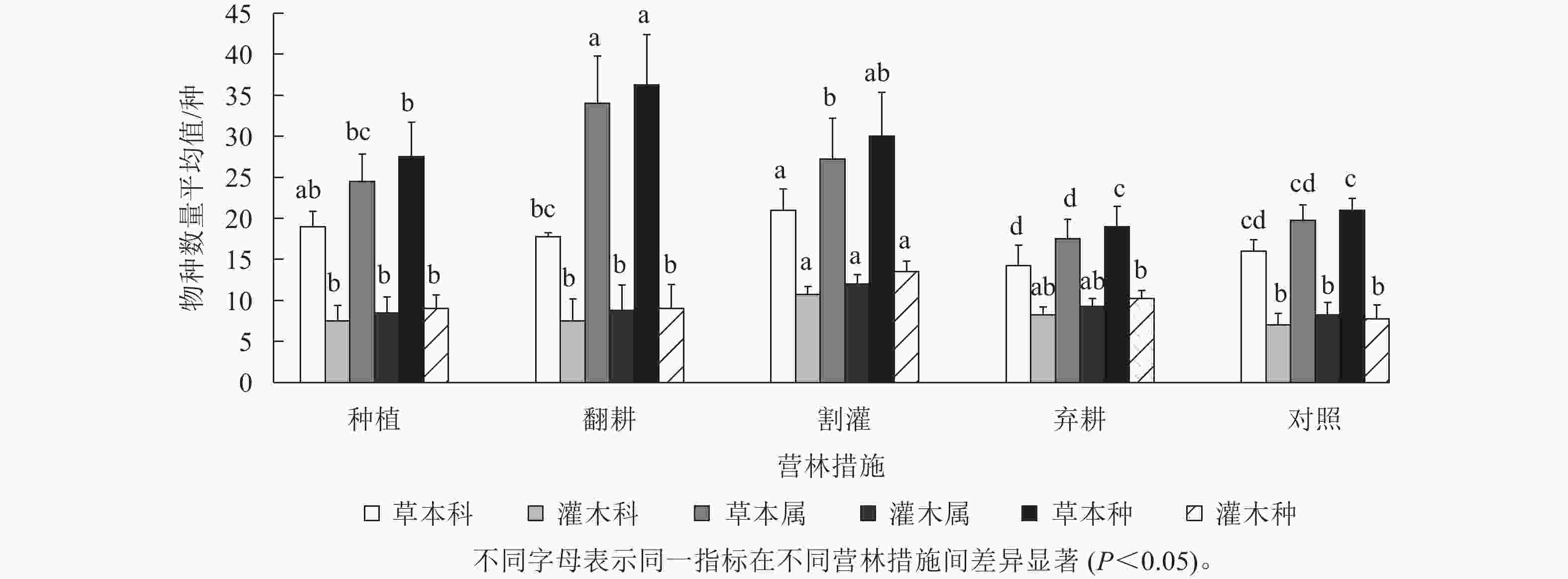
图 1 不同营林措施的杨树人工林林下植物物种组成
Figure 1. Species composition with the under canopy planting in P. canadensis plantation under different management measures
-
分别选取不同营林措施各样地中重要值之和大于0.6的前12位灌、草植物为优势种进行相关分析。研究表明:草本层优势种求米草、蜘蛛抱蛋、葎草、龙葵Solanum nigrum、乌蔹莓Cayratia japonica在5种营林措施中的重要值居于前列(表2),其中除种植样地之外,求米草与葎草在其他样地中的重要值均偏大,这可能由于葎草觅光能力强、生长周期短,使其能快速调整以适应光照环境变化的原因;杠板归Polygonum perfoliatum作为翻耕、割灌样地中1年生喜温喜光的优势种,在割灌样地中具有更强的适应性。
表 2 不同营林措施下加杨人工林草本层优势种的重要值和相对频度
Table 2. Importance value and relative frequency of dominant herbaceous species in P. canadensis plantation under different management measures
序号 物种 营林措施 种植 翻耕 割灌 弃耕 对照 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 H1 蜘蛛抱蛋 Aspidistra elatior 0.36 0.21 0.10 0.07 H2 求米草 Oplismenus undulatifolius 0.11 0.10 0.05 0.06 0.14 0.17 0.09 0.10 H3 空心莲子草 Alternanthera philoxeroides 0.06 0.08 0.04 0.04 H4 皱叶狗尾草 Setaria plicata 0.04 0.01 0.03 0.04 H5 狗尾草 Setaria viridis 0.03 0.01 0.03 0.03 0.03 0.03 H6 龙葵 Solanum nigrum 0.03 0.02 0.05 0.03 0.06 0.05 0.04 0.04 0.05 0.03 H7 乌蔹莓 Cayratia japonica 0.02 0.04 0.11 0.10 0.06 0.05 H8 吉祥草 Reineckia carnea 0.02 0.02 H9 冷水花 Pilea notata 0.02 0.03 0.02 0.03 0.04 0.02 H10 过路黄 Lysimachia christiniae 0.02 0.02 H11 蜈蚣草 Pteris vittata 0.02 0.01 0.02 0.04 H12 艾 Artemisia argyi 0.01 0.02 H13 葎草 Humulus scandens 0.15 0.08 0.06 0.08 0.18 0.21 0.27 0.32 H14 千金子 Leptochloa chinensis 0.05 0.03 H15 蛇莓 Duchesnea indica 0.05 0.05 H16 白车轴 Trifolium repens 0.04 0.03 H17 酢浆草 Oxalis corniculata 0.03 0.04 H18 野燕麦 Avena fatua 0.03 0.02 H19 杠板归 Polygonum perfoliatum 0.03 0.02 0.10 0.14 H20 狗娃花 Heteropappus hispidus 0.02 0.03 H21 牛膝 Achyranthes bidentata 0.03 0.06 H22 野茼蒿 Crassocephalum crepidioides 0.09 0.06 H23 土牛膝 Achyranthes aspera 0.04 0.04 H24 紫菀 Aster tataricus 0.07 0.04 H25 商陆 Phytolacca acinosa 0.03 0.04 H26 接骨草 Sambucus chinensis 0.03 0.03 0.07 0.07 0.04 0.05 H27 鸢尾 Iris tectorum 0.07 0.06 H28 序叶苎麻 Boehmeria clidemioides 0.04 0.04 H29 凹叶景天 Sedum emarginatum 0.01 0.03 H30 鬼针草 Bidens pilosa 0.03 0.03 0.02 0.03 H31 飞蓬 Erigeron acer 0.01 0.02 H32 辣椒 Capsicum annuum 0.03 0.05 H33 凤尾蕨 Pteris cretica 0.01 0.03 H34 雾水葛 Pouzolzia zeylanica 0.02 0.02 H35 铁苋菜 Acalypha australis 0.03 0.02 H36 牵牛花 Ipomoea nil 0.03 0.02 由表3可知:研究区内灌木层中桑科Moraceae与樟科植物优势明显,且灌木层优势种在不同营林措施样地中相似度较大,其中构树、天竺桂、八角枫、桑Morus alba在5种样地中都有分布,显示出了绝对的竞争优势,表明这4个物种在各样地中对资源占有具有优势力。其中构树重要值占比大,占据较宽生态位,其作为根系发达的落叶树种,具有速生、耐贫瘠和良好的滞尘效果,故推荐可在该路域林下大面积种植。对比灌草层优势种相对频度与重要值,并未发现其存在显著的线性关系,这与重要值还受到物种相对盖度和相对密度影响,并不单一受到相对频度影响的原因有关。
表 3 不同营林措施下加杨人工林灌木层优势种的重要值和相对频度
Table 3. Importance value and relative frequency of dominant shrub species in P. canadensis plantation under different management measures
序号 物种 营林措施 种植 翻耕 割灌 弃耕 对照 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 S1 构树Broussonetia papyrifera 0.32 0.31 0.23 0.20 0.34 0.25 0.10 0.12 0.25 0.30 S2 天竺桂Cinnamomum japonicum 0.19 0.25 0.01 0.01 0.20 0.21 0.23 0.17 0.09 0.11 S3 伞房决明Senna corymbosa 0.11 0.04 0.02 0.04 S4 八角枫Alangium chinense 0.09 0.07 0.02 0.01 0.01 0.02 0.27 0.17 0.04 0.06 S5 桑Morus alba 0.08 0.09 0.13 0.13 0.02 0.03 0.04 0.08 0.01 0.02 S6 加杨Populus canadensis 0.04 0.03 0.11 0.15 0.01 0.02 S7 朴树Celtis sinensis 0.03 0.05 0.05 0.06 0.09 0.16 0.04 0.05 S8 鸡屎藤Paederia scandens 0.02 0.03 0.04 0.05 0.02 0.02 0.02 0.03 S9 女贞Ligustrum lucidum 0.02 0.03 0.02 0.04 S10 银木Cinnamomum septentrionale 0.02 0.03 0.05 0.04 0.01 0.01 0.03 0.05 S11 蓖麻Ricinus communis 0.02 0.01 S12 枫杨Pterocarya stenoptera 0.02 0.01 0.12 0.09 S13 乌桕Sapium sebiferum 0.09 0.12 S14 细叶楠Phoebe hui 0.06 0.04 S15 海桐Pittosporum tobira 0.03 0.02 S16 二乔玉兰Yulania soulangeana 0.03 0.02 S17 栾树Koelreuteria paniculata 0.06 0.06 S18 野桐Mallotus japonicus 0.05 0.04 S19 茶树Camellia sinensis 0.04 0.06 S20 山麻杆Alchornea davidii 0.03 0.04 S21 覆盆子Rubus idaeus 0.03 0.03 S22 接骨木Sambucus williamsii 0.03 0.01 S23 四蕊朴Celtis tetrandra 0.03 0.04 S24 插田泡Rubus coreanus 0.03 0.04 S25 山柿Diospyros montana 0.02 0.02 S26 毛桐Mallotus japonicus 0.02 0.02 S27 臭牡丹Clerodendrum bungei 0.15 0.10 0.04 0.05 S28 小蜡Ligustrum sinense 0.04 0.06 0.01 0.02 S29 复羽叶栾树Koelreuteria bipinnata 0.01 0.02 0.18 0.14 S30 香椿Toona sinensis 0.01 0.01 0.02 0.03 S31 慈竹Neosinocalamus affinis 0.22 0.06 S32 大芽南蛇藤Celastrus gemmatus 0.01 0.01 0.02 0.03 S33 枇杷Eriobotrya japonica 0.01 0.01 0.01 0.02 -
从表4可见:不同营林措施下草本层优势种的方差比率(VR)从大到小依次为种植、割灌、翻耕、对照、弃耕。其中种植、割灌的VR>1,表明种植、割灌样地草本层总体关联性呈正关联,检验统计量(W)在χ20.95(47)与χ20.05(47)临界值内,总体关联性呈不显著正关联;弃耕、对照样地草本层VR<1,其中弃耕样地的W在χ20.95(47)临界值外,总体关联性呈显著负关联,对照样地的W在χ20.95(47)与χ20.05(47)临界值内,总体关联性呈不显著负关联,表明弃耕、对照样地草本群落总体联结性不强,演替更新缓慢。
表 4 不同营林措施下加杨人工林林下植物的总体关联性
Table 4. Overall correlation of the under canopy planting in P. canadensis plantation under different management measures
营林措施 草本层 灌木层 方差比率
(VR)检验统计量
(W)χ2临界值
[χ20.95(47), χ20.05(47)]检验结果 方差比率
(VR)检验统计量
(W)χ2临界值
[χ20.95(23), χ20.05(23)]检验结果 种植 1.26 60.45 32.27,64.00 不显著正关联 1.06 25.38 13.09,35.17 不显著正关联 翻耕 0.80 38.27 32.27,64.00 不显著负关联 3.02 72.52 13.09,35.17 显著正关联 割灌 1.08 52.74 32.27,64.00 不显著正关联 1.30 31.14 13.09,35.17 不显著正关联 弃耕 0.62 29.94 32.27,64.00 显著负关联 0.81 19.42 13.09,35.17 不显著负关联 对照 0.69 32.98 32.27,64.00 不显著负关联 1.17 28.01 13.09,35.17 不显著正关联 不同营林措施下灌木层优势种的VR从大到小依次为翻耕、割灌、对照、种植、弃耕。其中翻耕、割灌、对照、种植样地的VR>1,翻耕样地的W在χ20.95(23)临界值外,总体关联性呈显著正关联;割灌、对照、种植样地的W在χ20.95(23)与χ20.05(23)临界值内,总体关联性呈不显著正关联;弃耕样地灌木层优势种VR<1,W在χ20.95(23)与χ20.05(23)临界值内,总体关联性呈不显著负关联,因此表明除弃耕样地外,其他样地灌木层优势种种间联结强度高,群落均处于相对稳定状态。
-
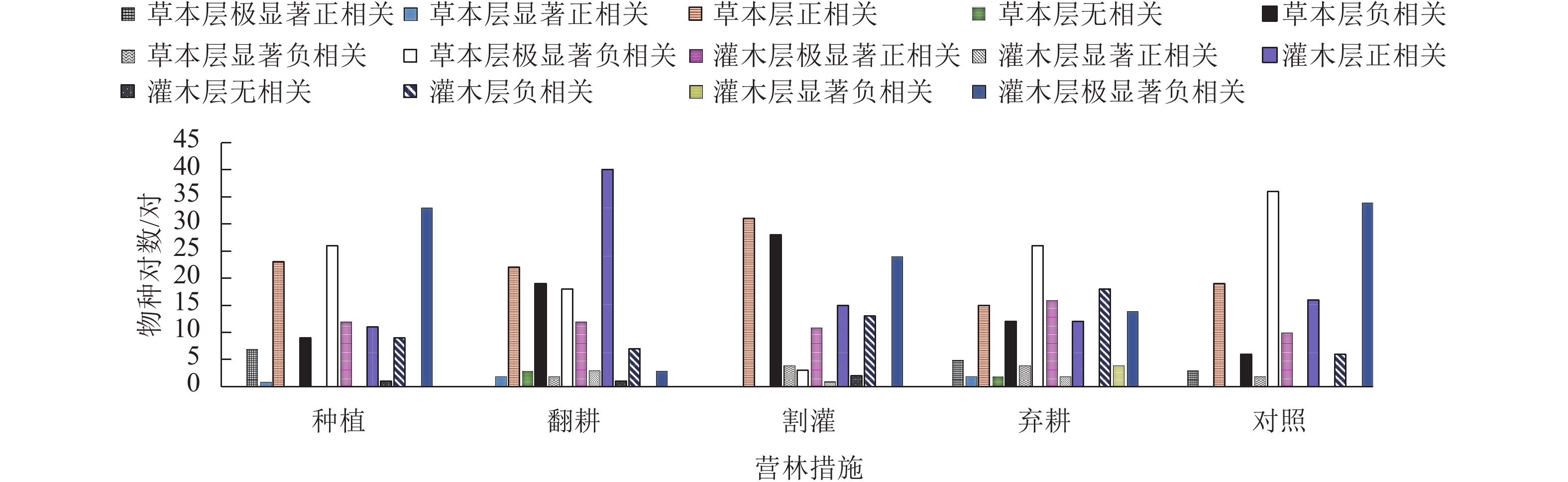
由图2可知:灌草层优势种对数均为78对,其种对间主要为正、负相关,且正相关种对数明显多于负相关。在不同营林措施下草本层优势种正相关种对数差异不大,但在翻耕样地中其负相关种对数明显减少为最小值,表明翻耕样地种对间竞争相对温和。灌木优势种在不同营林措施样地中正相关种对数差异不大,这与研究区草本层结论相似;不同营林措施下灌木优势种存在较大相似度,但种对间正负相关性却存在显著差别,例如种对构树和八角枫在种植样地中为负相关,在弃耕样地中为正相关。弃耕样地中灌草优势种正相关种对数均最少,负相关种对数均最多,表明其灌草层种间竞争强度均大。

图 2 加杨人工林林下植物优势种的χ2检验半矩阵图
Figure 2. χ2 test semi-matrix of dominant species in the under canopy planting of P. canadensis plantation
种间联结系数和Spearman秩相关系数因比χ2检验更灵敏,故可以作为χ2检验的有效补充[28]。由图3可知:不同营林措施样地中草本层优势种种对间主要为正负相关及极显著负相关,灌木层优势种种对间主要为正负相关及极显著正负相关,这较χ2检验结果更加敏感。比较不同营林措施样地中灌草层优势种种间联结系数可知:在弃耕样地中灌草层优势种极显著正相关及正相关种对数基本最少,极显著负相关及负相关种对数均最多,这与χ2检验结果一致;在翻耕与割灌样地中,灌草优势种种对关系出现显著差异,翻耕样地灌木优势种正相关种对数明显多于草本层,而割灌样地草本优势种正相关种对数显著高于灌木层,2种营林措施导致灌草层正负种对数呈相反变化趋势;在种植、对照样地中,灌草优势种极显著负相关种对数均较多,表明2种营林措施下,种对间均存在十分激烈的竞争现象。
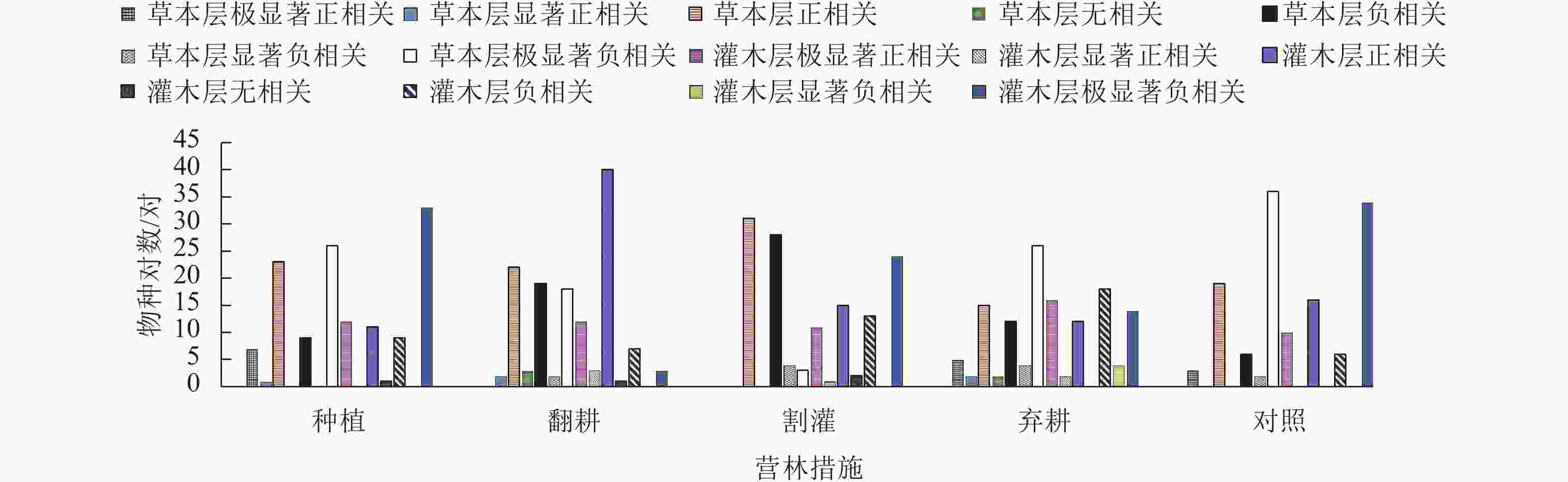
图 3 不同营林措施的加杨人工林林下植物种间联结系数(AC)结果统计
Figure 3. Statistics of connection coefficient (AC) of the under canopy planting in P. canadensis plantation under different management measures
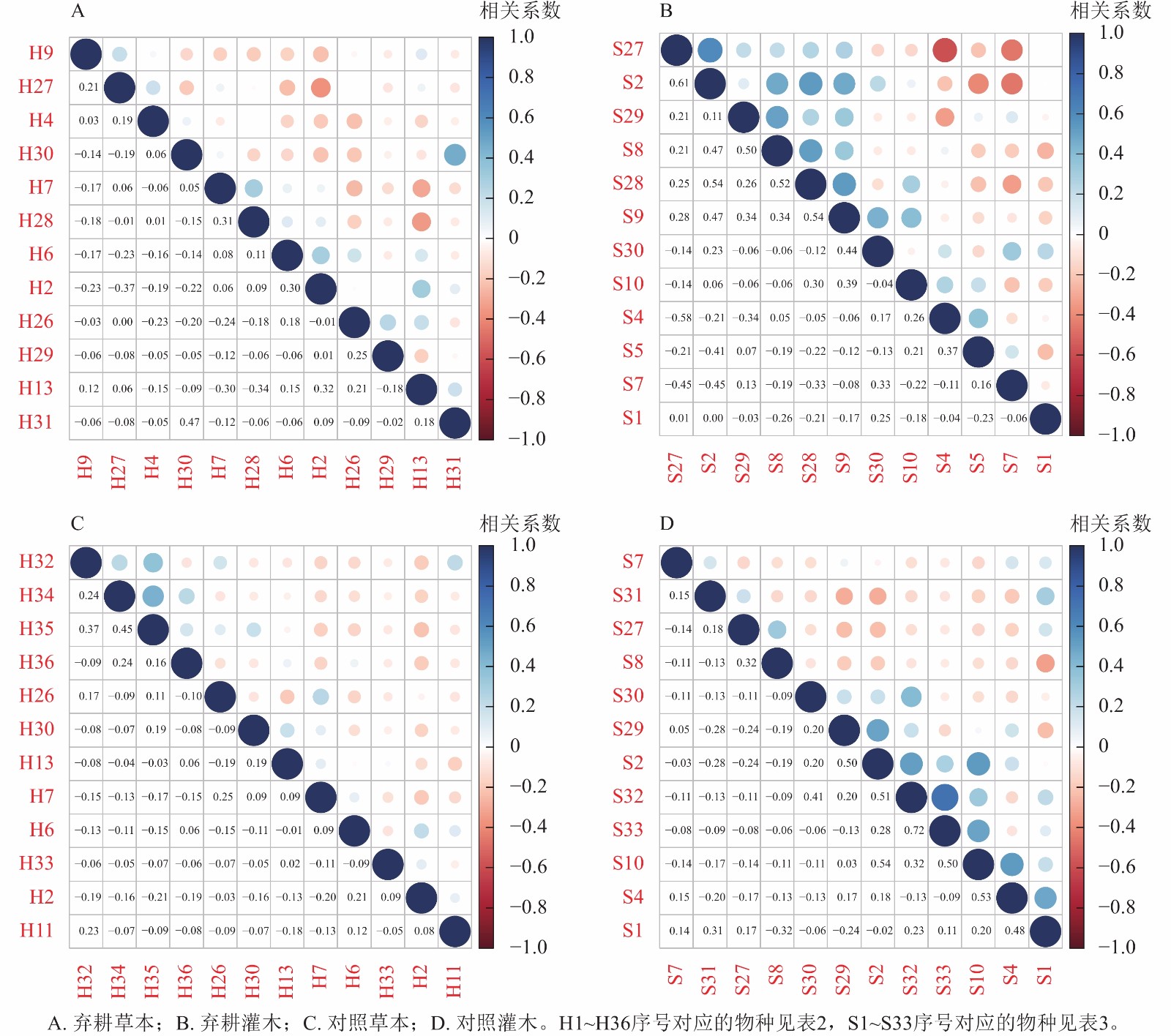
Spearman秩相关系数体现种间关联相对强弱。不同营林措施样地中灌木层优势种显著正相关种对数显著多于草本层。对比灌草层优势种Spearman秩相关系数可知:在种植样地中草本层优势种正相关种对数最多,负相关最少,灌木层优势种正相关种对数最少,负相关最多;在翻耕样地中,灌木层优势种正相关种对数最多,草本层则偏少;在割灌样地中草本层优势种正相关种对数偏多,灌木层则偏少,因此表明在翻耕、割灌样地中,灌草层优势种正负相关种对数变化趋势相反,这与种间联结系数的分析结果一致;在种植、对照样地中,灌木层正相关种对数差异不大,草本层正相关种对数出现明显区别,这可能与Spearman秩相关系数比种间联结系数反应更加敏捷有关[28]。在弃耕样地中草本层优势种求米草和乌蔹莓间存在正相关,而在对照样地中则表现为负相关(图4),表明同种种对间由于资源分配差异等因素,会产生不同的相关性甚至是相反关系,这与χ2检验结果相似。
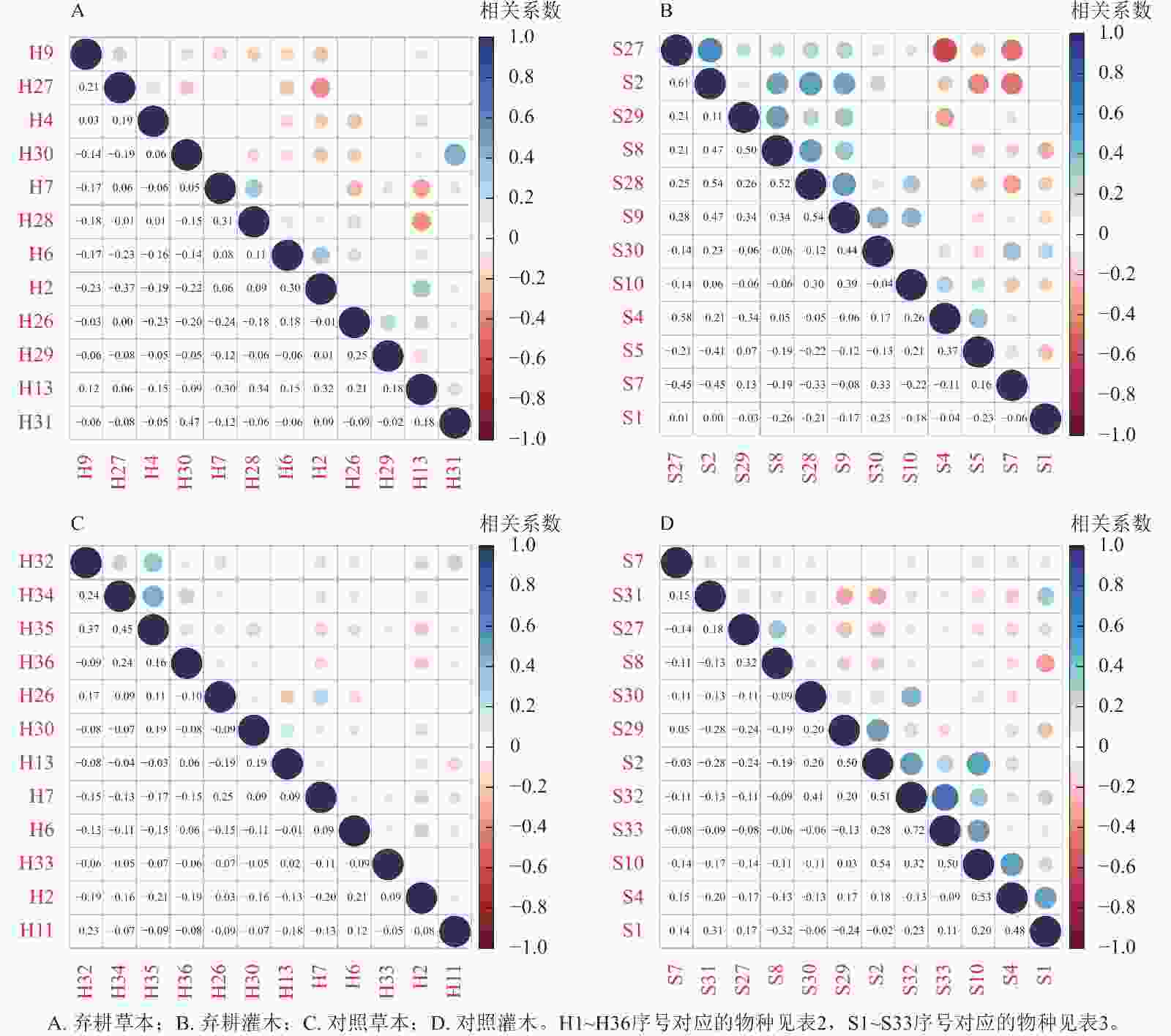
图 4 加杨人工林林下植物优势种的Spearman秩相关系数半矩阵图
Figure 4. Spearman rank correlation coefficient semi-matrix of dominant species to the under canopy planting in P. canadensis plantation
-
采用M. Gordon稳定性计算方法绘制不同营林措施下灌草层优势种稳定性图(图5)以及林下植物群落整体稳定性分析表(表5),其中欧氏距离可定量衡量群落的稳定性,距离越近,其稳定性越好。由此表明:在弃耕时,草本层群落稳定性最好,在种植时,灌木层群落稳定性最好,林下植物群落整体稳定性最好。
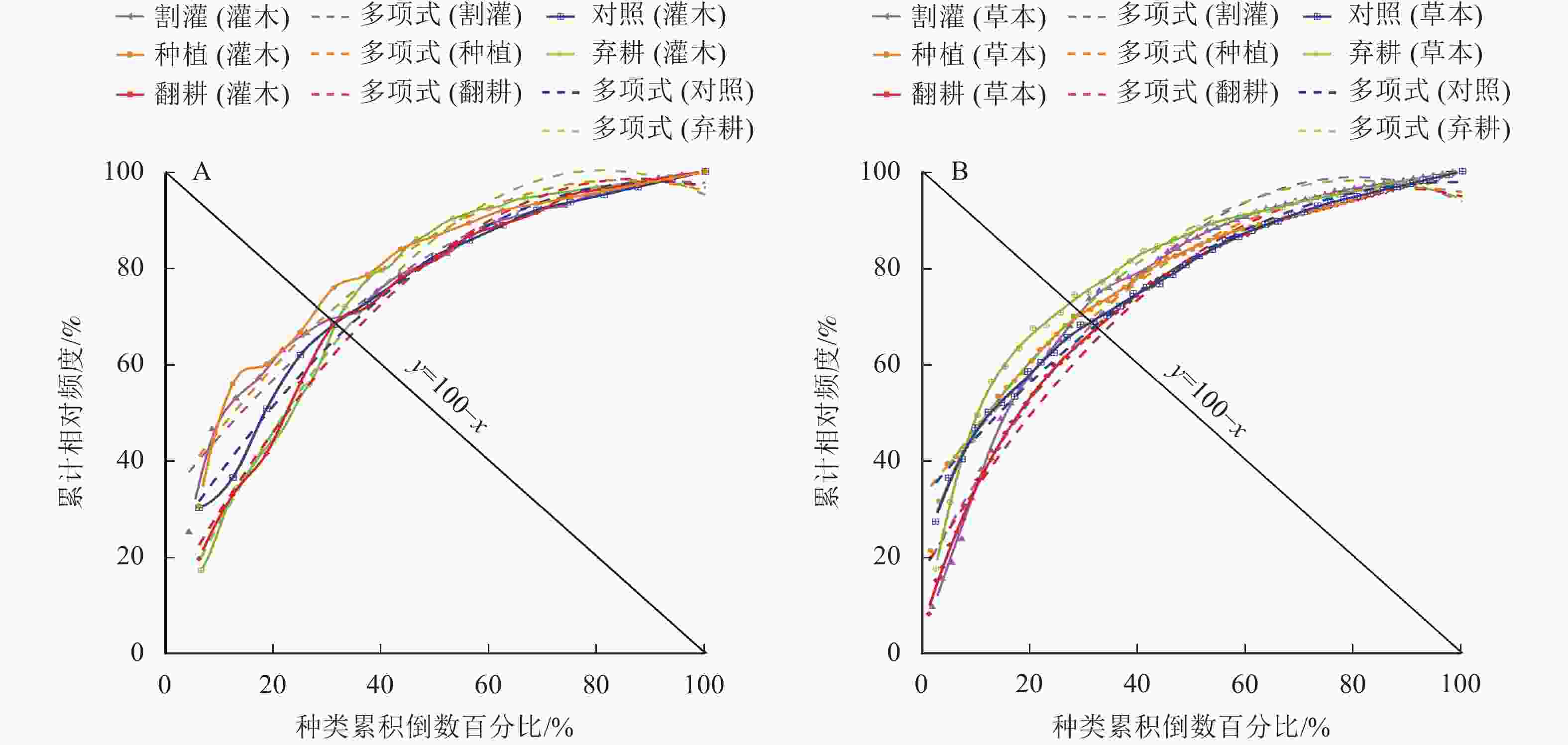
图 5 不同营林措施下加杨人工林灌木(A)和草本(B)的群落稳定性
Figure 5. Shrub (A) and herb (B) community stability of P. canadensis plantation under different management measures
表 5 不同营林措施下加杨人工林林下植物群落稳定性分析结果
Table 5. Results of community stability in the under canopy planting of P. canadensis plantation under different management measures
层级 营林措施 曲线类型 决定系数(R2) P 交点坐标 欧氏距离 结果 草本层 种植 y=−0.008 1x2+1.44x+32.3 0.98 <0.001 (30.90, 69.10) 15.42 不稳定 翻耕 y=−0.010 6x2+1.84x+16.8 0.99 <0.001 (33.48, 66.52) 19.06 不稳定 割灌 y=−0.012 9x2+2.06x+16.2 0.97 <0.001 (31.59, 68.41) 16.40 不稳定 弃耕 y=−0.010 3x2+1.66x+31.2 0.95 <0.001 (29.16, 70.84) 12.95 不稳定 对照 y=−0.006 9x2+1.34x+32.1 0.99 <0.001 (32.03, 67.97) 17.02 不稳定 灌木层 种植 y=−0.009 3x2+1.58x+31.4 0.96 <0.001 (29.80, 70.20) 13.86 不稳定 翻耕 y=−0.011 6x2+2.02x+10.2 0.99 <0.001 (34.24, 65.76) 20.14 不稳定 割灌 y=−0.007 5x2+1.41x+31.5 0.97 <0.001 (31.51, 68.49) 16.28 不稳定 弃耕 y=−0.014 5x2+2.35x+4.81 0.99 <0.001 (33.18, 66.82) 18.64 不稳定 对照 y=−0.009 3x2+1.69x+21.4 0.99 <0.001 (32.97, 67.03) 18.35 不稳定 林下植物 种植 y=−0.000 5x2+0.71x+31.1 0.93 <0.001 (40.82, 59.18) 29.44 不稳定 翻耕 y=−0.003 5x2+1.09x+19.7 0.94 <0.001 (41.31, 58.69) 30.14 不稳定 割灌 y=−0.002 3x2+1.01x+21.4 0.94 <0.001 (41.03, 58.97) 29.74 不稳定 弃耕 y=0.001 0x2+0.59x+31.1 0.91 <0.001 (42.18, 57.82) 31.37 不稳定 对照 y=−0.000 2x2+0.70x+29.4 0.97 <0.001 (41.40, 58.60) 30.27 不稳定 说明:x表示种类累积倒数百分比,y表示累计相对频度。 图5显示:不同营林措施下草本层稳定性从大到小依次为弃耕、种植、割灌、对照、翻耕,灌木层稳定性从大到小依次为种植、割灌、对照、弃耕、翻耕,林下植物整体稳定性从大到小依次为种植、割灌、翻耕、对照、弃耕。群落各层级均远离交点坐标(20, 80),表明群落均不稳定,演替处于初级阶段;其中种植、割灌下的灌草层、林下植物群落稳定性相对较高,表明林下种植与割灌措施均有利于群落演替更新,群落可更快演替为相对稳定状态;翻耕明显影响灌草层稳定性,演替更新缓慢;弃耕样地中,草本层稳定性最高,但灌木层、林下植物群落稳定性均最低。灌木层各营林措施稳定性差异大于草本层,表明灌木层对各营林措施的响应程度更强烈,草本层则相对更温和。
-
物种间并不是完全独立的,而是会产生正负等关联[29],这种关联强弱直接影响群落资源分配和利用,对群落可持续发展有重要影响[6, 11]。本研究对比不同营林措施下灌草层的总体关联性表明:翻耕对草本植物损伤更大,更有利于灌木层优势种对资源的占有和利用;种植作为一种典型的营林措施,能增强灌草层种群间总体关联性,促进灌草层向高层级演替更新,而割灌样地在进行割灌处理后,显著抑制了灌木层对资源的争夺能力,改善了草本层对水热资源的接受总量,平衡了灌草层种间竞争程度;对照样地灌草层整体关联强度偏低,说明适当的营林措施可促进加杨人工林林下植物的演替更新。本研究表明:营林措施翻耕、割灌可使灌草层资源分配更加合理,有利于种群的总体关联性。
-
物种间竞争、协同等相互关系是产生种间关联不同性质与关联程度的原因,主要受环境与自身生物学特性等影响,是群落稳定性的体现[3−4]。在对各样地进行χ2检验、种间联结系数和Spearman秩相关系数检验时出现差异可能与计算方法有关,因种间联结系数和Spearman秩相关系数相较于χ2检验更加灵敏,故本研究更多参考前两者检验结果[28]。种间关联研究表明:在翻耕、割灌样地中灌草层优势种正负相关种对数呈相反变化趋势,这可能与翻耕损伤了草本层根系,减弱草本层竞争优势,而割灌直接破坏了灌木生长过程,有利于草本层对资源的获取有关;在种植和对照样地中,Spearman秩相关系数检验结果显示:营林措施对灌木层影响小,主要是对草本层产生影响,这与GILLIAM[18]的研究结果相似,表明适当的营林措施可改变草本层优势种间正负关联性并提高其关联程度。因此,种间关联性研究结果显示:营林措施翻耕、割灌可平衡灌草层资源利用,有利于群落整体演替更新,提高群落整体的生态效益;营林措施种植可提高草本层关联强度,更有利于草本群落稳定性。
-
种间关联性与群落稳定性密切相关,可间接预测群落稳定性,群落稳定性是物种生态适应的综合表现[4],而M. Gordon稳定性计算方法可直接反映群落稳定性,但并不能具体反映种间关联性。物种从种间关联性到群落稳定性是一个漫长复杂的生态学过程,两者可对群落稳定性进行相互验证,具有一定的互补性和可靠性。M. Gordon稳定性计算方法表明:灌草各层级与林下植物整体群落均处于不稳定状态,群落处于演替的初级阶段。灌草层以及林下植物整体稳定性最好的分别为种植、弃耕、种植,其中营林措施翻耕明显影响灌草层群落稳定性,其稳定性均处于最低,这与总体关联性、种间关联性的分析结果不一致,特别是灌木层存在较大差异。这可能是由于总体关联性、种间关联性是通过种间关联强度间接预测群落稳定性,而M. Gordon稳定性计算方法是直接说明群落稳定性的指标,因此对于群落稳定性检验结果应更多参考后者。本研究表明:营林措施弃耕草本层稳定性最高,但灌木层、林下植物群落稳定性均最低,表明前期农耕等人为干扰活动会造成土壤养分流失,影响后期植物生长,并对群落整体稳定性有较大的负面影响,不利于群落的演替更新,这与苏天成等[16]的研究结果相似;在种植和割灌营林措施下,灌木、草本、林下植物群落稳定性均较高,表明林下种植与割灌抚育可促进群落演替更新并提高加杨人工林林下植物群落整体稳定性。
Effects of forest management of the Populus canadensis plantations on the interspecific association of the under canopy planting in expressway areas
-
摘要:
目的 路域森林已成为现代林业的重要组成部分,但仍存在脆弱性和敏感性等问题,研究不同营林措施对成都绕城高速路域加杨Populus canadensis人工林林下植物种间关联及群落稳定性的影响,可为高速路域人工林提出适当林分抚育措施,以提高其整体生态效益。 方法 以成都第一绕城高速路域加杨人工林为研究对象,采取典型抽样法设置20个样地,以造林开始后未发生任何经营活动的样地为对照,通过方差比率法、χ2检验、联结系数、Spearman秩相关系数、M. Godron稳定性等测度,比较不同营林措施(种植、翻耕、割灌、弃耕、对照)对高速路域加杨人工林林下植物种间关联和群落稳定性的影响。 结果 不同营林措施样地中草本层联结系数从大到小依次为种植、割灌、翻耕、对照、弃耕,其中种植、割灌样地的联结系数大于1,总体关联性为正关联;灌木层联结系数从大到小依次为种植、割灌、对照、种植、弃耕,仅弃耕样地的联结系数小于1,总体关联性为负关联。种间关联分析表明:种植、割灌的灌草层优势种正、负相关种对数呈相反变化趋势,种植、对照主要是对草本优势群落产生影响。M. Godron稳定性分析表明:5种营林措施样地群落整体均不稳定,为演替的初级阶段,其中种植、割灌群落整体稳定性相对较好。 结论 营林措施中,种植、割灌的灌草优势种种间关联强度更好,可促进群落演替更新及高速路域加杨人工林林下植物群落整体稳定性。图5表5参29 Abstract:Objective Expressway forest, as an important part of modern forestry, is undergoing common problems such as vulnerability and sensitivity. Therefore, this study, with an investigation into the effects of different forest management measures on the interspecific association of the under canopy planting and community stability, is aimed to put forward appropriate stand rearing measures for the Populus canadensis plantation in Chengdu ring expressway area so as to improve its overall ecological benefits. Method First, taking the P. canadensis plantation in the first expressway area of Chengdu as the research object, a total of 20 plots were set up by typical sampling method. Then, with the empployment of the variance ratio method (VR), χ2 test, connection coefficient (AC), Spearman rank correlation coefficient and M. Godron stability were measured before a comparative analysis was conducted of the effects of under canopy planting (UP), plow tillage (PT), swamping (SW), abandoned cultivation (AC) and control group (the sample land without any business activities after afforestation as a control group, CG) on interspecific association and community stability. Result The VR values of dominant herb species displayed a trend of UP>SW>PT>CG>AC under different management measures, and the VR (>1) values in UP and SW plots demonstrated positive correlation whereas the VR values of dominant shrub species showed a trend of PT>SW>CG>UP>AC and only the VR (<1) values in AC plots showed a negative correlation; Interspecific association analysis showed that the number of species of dominant species in shrub and herb layer with positive correlation displayed an opposite trend from that of those with negative correlation with managing methods PT and SW and the managing methods of UP and CG mainly affected dominant herb species; The stability analysis of M. Godron showed a relatively weak overall stability of the communities in the five forestry measures plots which were all in the primary stage of succession, with the UP and SW communities having the highest overall stability. Conclusion The interspecific association strength of the dominant shrub species is better in UP and SW which can help promote community succession and the overall stability of the under canopy planting community. [Ch, 5 fig. 5 tab. 29 ref.] -
图 1 不同营林措施的杨树人工林林下植物物种组成
Figure 1 Species composition with the under canopy planting in P. canadensis plantation under different management measures
图 2 加杨人工林林下植物优势种的χ2检验半矩阵图
Figure 2 χ2 test semi-matrix of dominant species in the under canopy planting of P. canadensis plantation
图 3 不同营林措施的加杨人工林林下植物种间联结系数(AC)结果统计
Figure 3 Statistics of connection coefficient (AC) of the under canopy planting in P. canadensis plantation under different management measures
图 4 加杨人工林林下植物优势种的Spearman秩相关系数半矩阵图
Figure 4 Spearman rank correlation coefficient semi-matrix of dominant species to the under canopy planting in P. canadensis plantation
图 5 不同营林措施下加杨人工林灌木(A)和草本(B)的群落稳定性
Figure 5 Shrub (A) and herb (B) community stability of P. canadensis plantation under different management measures
表 1 样地基本情况
Table 1. Basic situation of the plot
样地
编号林分密度/
(株·hm−2)郁闭度 平均树高/m 平均胸径/cm 营林
措施备注 1~4 537.50 0.69 16.49±0.43 b 23.79±0.70 b 种植 种植样地:在林冠下充分开展农、林业的双向复合经营模式,是最典型的营林措施。本研究区于2018年3月集中种植蜘蛛抱蛋Aspidistra elatior,套种密度为30 株·m−2 5~8 537.50 0.80 16.84±0.39 b 28.11±0.92 a 翻耕 翻耕样地:用有壁犁等农具使土壤结构疏松的耕作方法,以改善土壤条件,在每年的春冬季去除地表残茬和杂草,留茬12~15 cm 9~12 516.67 0.70 12.97±0.43 c 19.42±1.08 c 割灌 割灌抚育样地:通常在冬季距离道路较近处进行割灌作业,多在当年初夏(集中在5—6月)进行,且割除的灌丛茬高不超过5 cm 13~16 550.00 0.78 18.09±0.28 a 25.41±0.69 b 弃耕 弃耕样地:原进行农业经营的土地不再进行农耕作业而撂荒的土地,该样地为林下种植后弃耕样地 17~20 562.50 0.71 18.27±0.14 a 24.80±0.50 b 对照 对照样地:造林开始后未发生任何经营活动的样地 说明:不同字母表示营林措施间数据差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 不同营林措施下加杨人工林草本层优势种的重要值和相对频度
Table 2. Importance value and relative frequency of dominant herbaceous species in P. canadensis plantation under different management measures
序号 物种 营林措施 种植 翻耕 割灌 弃耕 对照 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 H1 蜘蛛抱蛋 Aspidistra elatior 0.36 0.21 0.10 0.07 H2 求米草 Oplismenus undulatifolius 0.11 0.10 0.05 0.06 0.14 0.17 0.09 0.10 H3 空心莲子草 Alternanthera philoxeroides 0.06 0.08 0.04 0.04 H4 皱叶狗尾草 Setaria plicata 0.04 0.01 0.03 0.04 H5 狗尾草 Setaria viridis 0.03 0.01 0.03 0.03 0.03 0.03 H6 龙葵 Solanum nigrum 0.03 0.02 0.05 0.03 0.06 0.05 0.04 0.04 0.05 0.03 H7 乌蔹莓 Cayratia japonica 0.02 0.04 0.11 0.10 0.06 0.05 H8 吉祥草 Reineckia carnea 0.02 0.02 H9 冷水花 Pilea notata 0.02 0.03 0.02 0.03 0.04 0.02 H10 过路黄 Lysimachia christiniae 0.02 0.02 H11 蜈蚣草 Pteris vittata 0.02 0.01 0.02 0.04 H12 艾 Artemisia argyi 0.01 0.02 H13 葎草 Humulus scandens 0.15 0.08 0.06 0.08 0.18 0.21 0.27 0.32 H14 千金子 Leptochloa chinensis 0.05 0.03 H15 蛇莓 Duchesnea indica 0.05 0.05 H16 白车轴 Trifolium repens 0.04 0.03 H17 酢浆草 Oxalis corniculata 0.03 0.04 H18 野燕麦 Avena fatua 0.03 0.02 H19 杠板归 Polygonum perfoliatum 0.03 0.02 0.10 0.14 H20 狗娃花 Heteropappus hispidus 0.02 0.03 H21 牛膝 Achyranthes bidentata 0.03 0.06 H22 野茼蒿 Crassocephalum crepidioides 0.09 0.06 H23 土牛膝 Achyranthes aspera 0.04 0.04 H24 紫菀 Aster tataricus 0.07 0.04 H25 商陆 Phytolacca acinosa 0.03 0.04 H26 接骨草 Sambucus chinensis 0.03 0.03 0.07 0.07 0.04 0.05 H27 鸢尾 Iris tectorum 0.07 0.06 H28 序叶苎麻 Boehmeria clidemioides 0.04 0.04 H29 凹叶景天 Sedum emarginatum 0.01 0.03 H30 鬼针草 Bidens pilosa 0.03 0.03 0.02 0.03 H31 飞蓬 Erigeron acer 0.01 0.02 H32 辣椒 Capsicum annuum 0.03 0.05 H33 凤尾蕨 Pteris cretica 0.01 0.03 H34 雾水葛 Pouzolzia zeylanica 0.02 0.02 H35 铁苋菜 Acalypha australis 0.03 0.02 H36 牵牛花 Ipomoea nil 0.03 0.02
下载: 导出CSV
表 3 不同营林措施下加杨人工林灌木层优势种的重要值和相对频度
Table 3. Importance value and relative frequency of dominant shrub species in P. canadensis plantation under different management measures
序号 物种 营林措施 种植 翻耕 割灌 弃耕 对照 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 重要值 相对频度 S1 构树Broussonetia papyrifera 0.32 0.31 0.23 0.20 0.34 0.25 0.10 0.12 0.25 0.30 S2 天竺桂Cinnamomum japonicum 0.19 0.25 0.01 0.01 0.20 0.21 0.23 0.17 0.09 0.11 S3 伞房决明Senna corymbosa 0.11 0.04 0.02 0.04 S4 八角枫Alangium chinense 0.09 0.07 0.02 0.01 0.01 0.02 0.27 0.17 0.04 0.06 S5 桑Morus alba 0.08 0.09 0.13 0.13 0.02 0.03 0.04 0.08 0.01 0.02 S6 加杨Populus canadensis 0.04 0.03 0.11 0.15 0.01 0.02 S7 朴树Celtis sinensis 0.03 0.05 0.05 0.06 0.09 0.16 0.04 0.05 S8 鸡屎藤Paederia scandens 0.02 0.03 0.04 0.05 0.02 0.02 0.02 0.03 S9 女贞Ligustrum lucidum 0.02 0.03 0.02 0.04 S10 银木Cinnamomum septentrionale 0.02 0.03 0.05 0.04 0.01 0.01 0.03 0.05 S11 蓖麻Ricinus communis 0.02 0.01 S12 枫杨Pterocarya stenoptera 0.02 0.01 0.12 0.09 S13 乌桕Sapium sebiferum 0.09 0.12 S14 细叶楠Phoebe hui 0.06 0.04 S15 海桐Pittosporum tobira 0.03 0.02 S16 二乔玉兰Yulania soulangeana 0.03 0.02 S17 栾树Koelreuteria paniculata 0.06 0.06 S18 野桐Mallotus japonicus 0.05 0.04 S19 茶树Camellia sinensis 0.04 0.06 S20 山麻杆Alchornea davidii 0.03 0.04 S21 覆盆子Rubus idaeus 0.03 0.03 S22 接骨木Sambucus williamsii 0.03 0.01 S23 四蕊朴Celtis tetrandra 0.03 0.04 S24 插田泡Rubus coreanus 0.03 0.04 S25 山柿Diospyros montana 0.02 0.02 S26 毛桐Mallotus japonicus 0.02 0.02 S27 臭牡丹Clerodendrum bungei 0.15 0.10 0.04 0.05 S28 小蜡Ligustrum sinense 0.04 0.06 0.01 0.02 S29 复羽叶栾树Koelreuteria bipinnata 0.01 0.02 0.18 0.14 S30 香椿Toona sinensis 0.01 0.01 0.02 0.03 S31 慈竹Neosinocalamus affinis 0.22 0.06 S32 大芽南蛇藤Celastrus gemmatus 0.01 0.01 0.02 0.03 S33 枇杷Eriobotrya japonica 0.01 0.01 0.01 0.02
下载: 导出CSV
表 4 不同营林措施下加杨人工林林下植物的总体关联性
Table 4. Overall correlation of the under canopy planting in P. canadensis plantation under different management measures
营林措施 草本层 灌木层 方差比率
(VR)检验统计量
(W)χ2临界值
[χ20.95(47), χ20.05(47)]检验结果 方差比率
(VR)检验统计量
(W)χ2临界值
[χ20.95(23), χ20.05(23)]检验结果 种植 1.26 60.45 32.27,64.00 不显著正关联 1.06 25.38 13.09,35.17 不显著正关联 翻耕 0.80 38.27 32.27,64.00 不显著负关联 3.02 72.52 13.09,35.17 显著正关联 割灌 1.08 52.74 32.27,64.00 不显著正关联 1.30 31.14 13.09,35.17 不显著正关联 弃耕 0.62 29.94 32.27,64.00 显著负关联 0.81 19.42 13.09,35.17 不显著负关联 对照 0.69 32.98 32.27,64.00 不显著负关联 1.17 28.01 13.09,35.17 不显著正关联
下载: 导出CSV
表 5 不同营林措施下加杨人工林林下植物群落稳定性分析结果
Table 5. Results of community stability in the under canopy planting of P. canadensis plantation under different management measures
层级 营林措施 曲线类型 决定系数(R2) P 交点坐标 欧氏距离 结果 草本层 种植 y=−0.008 1x2+1.44x+32.3 0.98 <0.001 (30.90, 69.10) 15.42 不稳定 翻耕 y=−0.010 6x2+1.84x+16.8 0.99 <0.001 (33.48, 66.52) 19.06 不稳定 割灌 y=−0.012 9x2+2.06x+16.2 0.97 <0.001 (31.59, 68.41) 16.40 不稳定 弃耕 y=−0.010 3x2+1.66x+31.2 0.95 <0.001 (29.16, 70.84) 12.95 不稳定 对照 y=−0.006 9x2+1.34x+32.1 0.99 <0.001 (32.03, 67.97) 17.02 不稳定 灌木层 种植 y=−0.009 3x2+1.58x+31.4 0.96 <0.001 (29.80, 70.20) 13.86 不稳定 翻耕 y=−0.011 6x2+2.02x+10.2 0.99 <0.001 (34.24, 65.76) 20.14 不稳定 割灌 y=−0.007 5x2+1.41x+31.5 0.97 <0.001 (31.51, 68.49) 16.28 不稳定 弃耕 y=−0.014 5x2+2.35x+4.81 0.99 <0.001 (33.18, 66.82) 18.64 不稳定 对照 y=−0.009 3x2+1.69x+21.4 0.99 <0.001 (32.97, 67.03) 18.35 不稳定 林下植物 种植 y=−0.000 5x2+0.71x+31.1 0.93 <0.001 (40.82, 59.18) 29.44 不稳定 翻耕 y=−0.003 5x2+1.09x+19.7 0.94 <0.001 (41.31, 58.69) 30.14 不稳定 割灌 y=−0.002 3x2+1.01x+21.4 0.94 <0.001 (41.03, 58.97) 29.74 不稳定 弃耕 y=0.001 0x2+0.59x+31.1 0.91 <0.001 (42.18, 57.82) 31.37 不稳定 对照 y=−0.000 2x2+0.70x+29.4 0.97 <0.001 (41.40, 58.60) 30.27 不稳定 说明:x表示种类累积倒数百分比,y表示累计相对频度。
下载: 导出CSV
-
[1] BROOKER R W, MAESTRE F T, CALLAWAY R M, et al. Facilitation in plant communities: the past, the present, and the future [J]. Journal of Ecology, 2008, 96(1): 18 − 34. [2] ARMAS C, PUGNAIRE F I. Plant interactions govern population dynamics in semiarid plant community [J]. Journal of Ecology, 2005, 93(5): 978 − 989. [3] 曹静, 白玉宏, 张钦弟, 等. 山西太岳山接骨木群落优势种种间关联性分析[J]. 生态学杂志, 2015, 34(5): 1313 − 1319. CAO Jing, BAI Yuhong, ZHANG Qindi, et al. Interspecific association analysis of dominant species in Sambucus williamsii community in Taiyue Mountain of Shanxi Province [J]. Chinese Journal of Ecology, 2015, 34(5): 1313 − 1319. [4] 涂洪润, 农娟丽, 朱军, 等. 桂林岩溶石山密花树群落主要物种的种间关联及群落稳定性[J]. 生态学报, 2022, 42(9): 3688 − 3705. TU Hongrun, NONG Juanli, ZHU Jun, et al. Interspecific associations of main species and community stability of Myrsine seguinii community in karst hills of Guilin, southwestern China [J]. Acta Ecologica Sinica, 2022, 42(9): 3688 − 3705. [5] GREIG-SMITH P. Quantitative Plant Ecology [M]. 3rd ed. Oxford: Blackwell Scientific Publications, 1983. [6] 张金屯. 数量生态学[M]. 3版. 北京: 科学出版社, 2018. ZHANG Jintun. Quantitative Ecology [M]. 3rd ed. Beijing: Science Press, 2018. [7] 龚直文, 亢新刚, 顾丽, 等. 长白山云冷杉针阔混交林两个演替阶段乔木的种间联结性[J]. 北京林业大学学报, 2011, 33(5): 28 − 33. GONG Zhiwen, KANG Xin’gang, GU Li, et al. Interspecific association among arbor species in two succession stages of spruce-fir conifer and broadleaved mixed forest in Changbai Mountains, northeastern China [J]. Journal of Beijing Forestry University, 2011, 33(5): 28 − 33. [8] MACARTHUR R. Fluctuations of animal populations and a measure of community stability [J]. Ecology, 1955, 36(3): 533 − 536. [9] 刘旻霞, 连依明, 李文. 微地形对优势种群点格局和关联性的影响[J]. 应用生态学报, 2018, 29(5): 1569 − 1575. LIU Minxia, LIAN Yiming, LI Wen. Effects of microtopography on the pattern and association of dominant population [J]. Chinese Journal of Applied Ecology, 2018, 29(5): 1569 − 1575. [10] 丁文慧, 李秀珍, 姜俊彦, 等. 崇明东滩南部河口盐沼植物群落种间关系的数量分析[J]. 应用生态学报, 2016, 27(5): 1417 − 1426. DING Wenhui, LI Xiuzhen, JIANG Junyan, et al. Numerical analysis of inter-specific relationships in the estuary salt marsh plant community of southern Chongming Dongtan, Shanghai [J]. Chinese Journal of Applied Ecology, 2016, 27(5): 1417 − 1426. [11] 董轲, 丁新峰, 郝广, 等. 围封年限对内蒙古灌丛化草原小叶锦鸡儿灌丛结构及群落种间关联的影响[J]. 生态学报, 2021, 41(14): 5775 − 5781. DONG Ke, DING Xinfeng, HAO Guang, et al. Effects of enclosure period on population structure of Caragana microphylla and interspecific associations in the Inner Mongolia shrub-encroached grasslands [J]. Acta Ecologica Sinica, 2021, 41(14): 5775 − 5781. [12] 齐麟, 赵福强. 不同采伐强度对阔叶红松林主要树种空间分布格局和物种空间关联性的影响[J]. 生态学报, 2015, 35(1): 46 − 55. QI Lin, ZHAO Fuqiang. Impact of logging intensity on the spatial distribution and association of dominant tree species in a broadleaved Korean pine mixed forest [J]. Acta Ecologica Sinica, 2015, 35(1): 46 − 55. [13] 崔玉华, 韩有志, 张梦弢, 等. 不同干扰强度下针阔混交林树种空间格局及种间关联性[J]. 应用生态学报, 2021, 32(6): 2053 − 2060. CUI Yuhua, HAN Youzhi, ZHANG Mengtao, et al. Spatial pattern and interspecific association of tree species in coniferous and deciduous broad-leaved mixed forest under different disturbance intensities [J]. Chinese Journal of Applied Ecology, 2021, 32(6): 2053 − 2060. [14] 张宝贵, 王耀斌, 刘光琇, 等. 甘肃省高速公路绿化策略研究[J]. 生态经济, 2014, 30(11): 170 − 176. ZHANG Baogui, WANG Yaobin, LIU Guangxiu, et al. Research on greening strategy of Gansu Province highways [J]. Ecological Economy, 2014, 30(11): 170 − 176. [15] CHRISTENSEN N L, BARTUSKA A M, BROWN J H, et al. The report of the ecological society of America committee on the scientific basis for ecosystem management [J]. Ecological Applications, 1996, 6(3): 665 − 691. [16] 苏天成, 王姚瑶, 向琳, 等. 营林措施对成都绕城高速路域杨树人工林群落结构和物种多样性的影响[J]. 应用与环境生物学报, 2022, 28(5): 1144 − 1150. SU Tiancheng, WANG Yaoyao, XIANG Lin, et al. Effects of forest management to the poplar plantations on the community structure and species diversity of the Chengdu ring expressway area [J]. Chinese Journal of Applied and Environmental Biology, 2022, 28(5): 1144 − 1150. [17] 胥晓刚, 杨冬生, 胡庭兴. 公路区域生态破坏及植被恢复技术应用与研究进展[J]. 中国园林, 2005(1): 54 − 57. XU Xiaogang, YANG Dongsheng, HU Tingxing. Discussion on the road environment destruction and vegetation restoration [J]. Chinese Landscape Architecture, 2005(1): 54 − 57. [18] GILLIAM F S. The ecological significance of the herbaceous layer in temperate forest ecosystems [J]. BioScience, 2007, 57(10): 845 − 858. [19] SU Xiangping, WANG Minhuang, HUANG Zhiqun, et al. Forest understorey vegetation: colonization and the availability and heterogeneity of resources [J/OL]. Forests, 2019, 10(11): 944[2022-09-10]. doi:10.3390/f10110944. [20] 李育中. 三种类型草地植物种间关联的测定与比较[J]. 生态学杂志, 1991, 10(6): 8 − 12. LI Yuzhong. Determination and comparison of plant species interconnections in three types of grassland [J]. Chinese Journal of Ecology, 1991, 10(6): 8 − 12. [21] KUKLOVA M, HNILIKOVA H, HNILIKA F, et al. Impact of expressway on physiology of plants and accumulation of risk elements in forest ecosystems [J]. Plant Soil &Environment, 2019, 65(1): 46 − 53. [22] 房莉. 杨树连栽对林木生长及其土壤肥力的影响[D]. 南京: 南京林业大学, 2005. FANG Li. Effects of Continuous Planting of Poplars on Forest Growth and Soil Fertility [D]. Nanjing: Nanjing Forestry University, 2005. [23] 方精云, 王襄平, 沈泽昊. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533 − 548. FANG Jingyun, WANG Xiangpin, SHEN Zehao. Main contents, methods and technical specifications of plant community inventory [J]. Biodiversity Science, 2009, 17(6): 533 − 548. [24] 王育松, 上官铁梁. 关于重要值计算方法的若干问题[J]. 山西大学学报(自然科学版), 2010, 33(2): 312 − 316. WANG Yusong, SHANGGUAN Tieliang. Some problems on calculation methods of important values [J]. Journal of Shanxi University (Natural Science Edition), 2010, 33(2): 312 − 316. [25] SCHULTER D. A variance test for detecting species association, with some example application [J]. Ecology, 1984, 65: 998 − 1005. [26] LUDWING J A, REYNOLDS J F. Statistical ecology: a primer on methods and computing [J]. Mathematical Biosciences, 1988, 95(2): 237 − 239. [27] 郑元润. 森林群落稳定性研究方法初探[J]. 林业科学, 2000, 36(5): 28 − 32. ZHEN Yuanrun. Comparison of methods for studying stability of forest community [J]. Scientia Silvae Sinicae, 2000, 36(5): 28 − 32. [28] 赵丽娅, 钟韩珊, 齐开, 等. 围封和放牧对科尔沁沙地植物群落种间关联的影响[J]. 生态学报, 2021, 41(9): 3724 − 3733. ZHAO Liya, ZHONG Hanshan, QI Kai, et al. Effects of enclosure and grazing on interspecific association of plant communities in Horqin sandy land, Inner Mongolia [J]. Acta Ecologica Sinica, 2021, 41(9): 3724 − 3733. [29] AGUIAR M R, SALA O E. Competition, facilitation, seed distribution and the origin of patches in a Patagonian steppe [J]. Oikos, 1994, 70(1): 26 − 34. -

-
链接本文:
https://zlxb.zafu.edu.cn/article/doi/10.11833/j.issn.2095-0756.20220604
 点击查看大图
点击查看大图

计量
- 文章访问数: 1106
- HTML全文浏览量: 211
- PDF下载量: 22
- 被引次数: 0
